
Neurotoxicity Assessment of Four Different Pesticides Using In Vitro Enzymatic Inhibition Assays

 , , , and
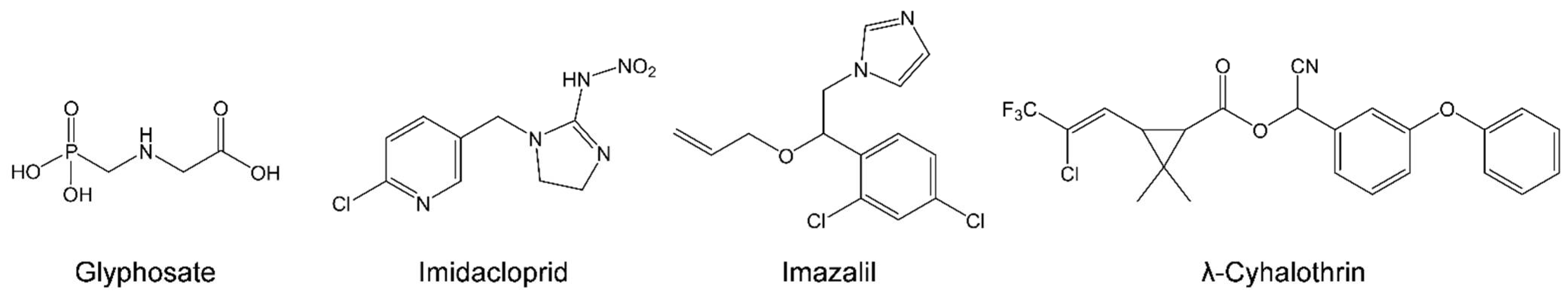
, , , and 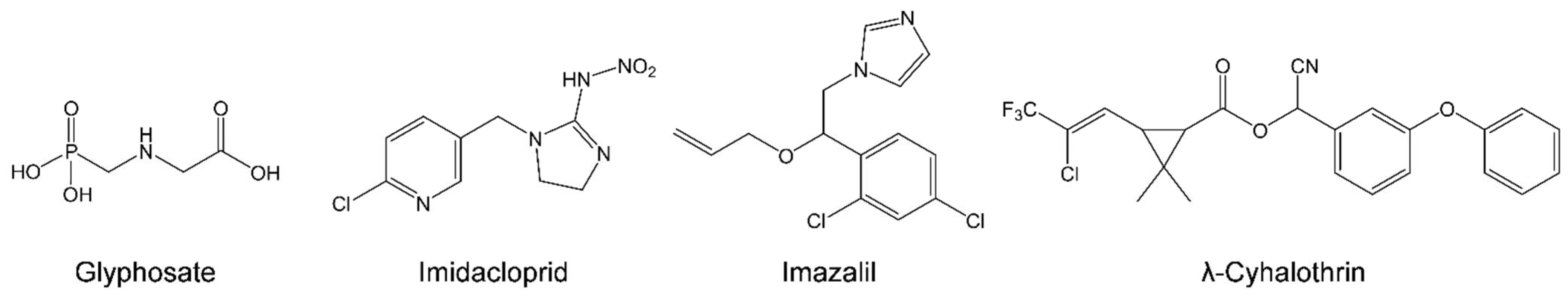{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials and Reagents
2.2. Enzyme Inhibition Assays
2.2.1. Cholinesterase Inhibition Assay
2.2.2. Tyrosinase Inhibition Assay
2.3. Data and Statistical Analysis
3. Results and Discussion
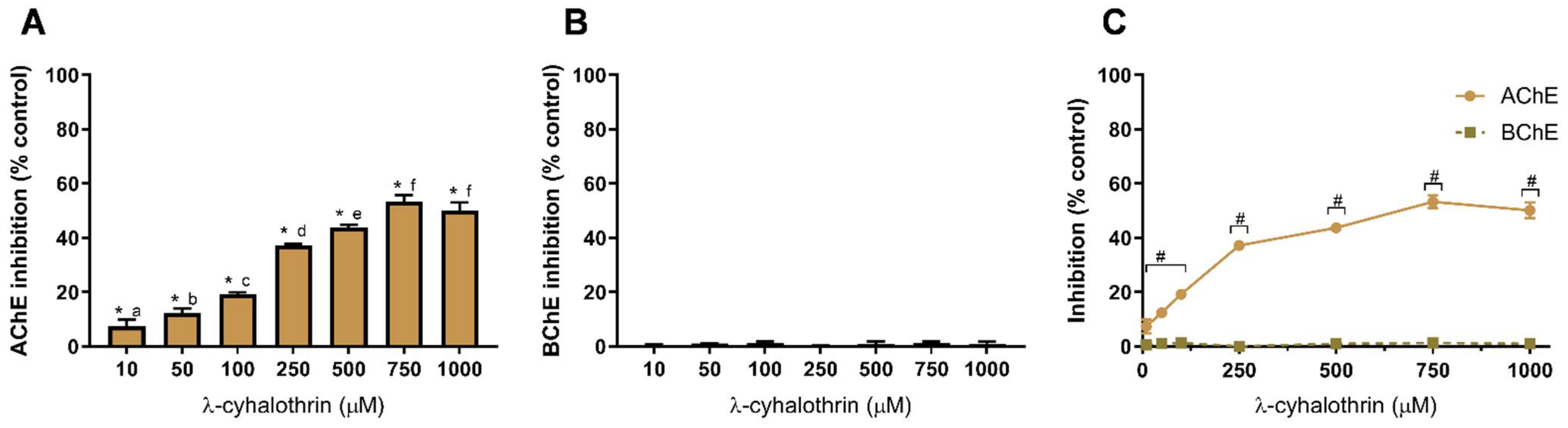
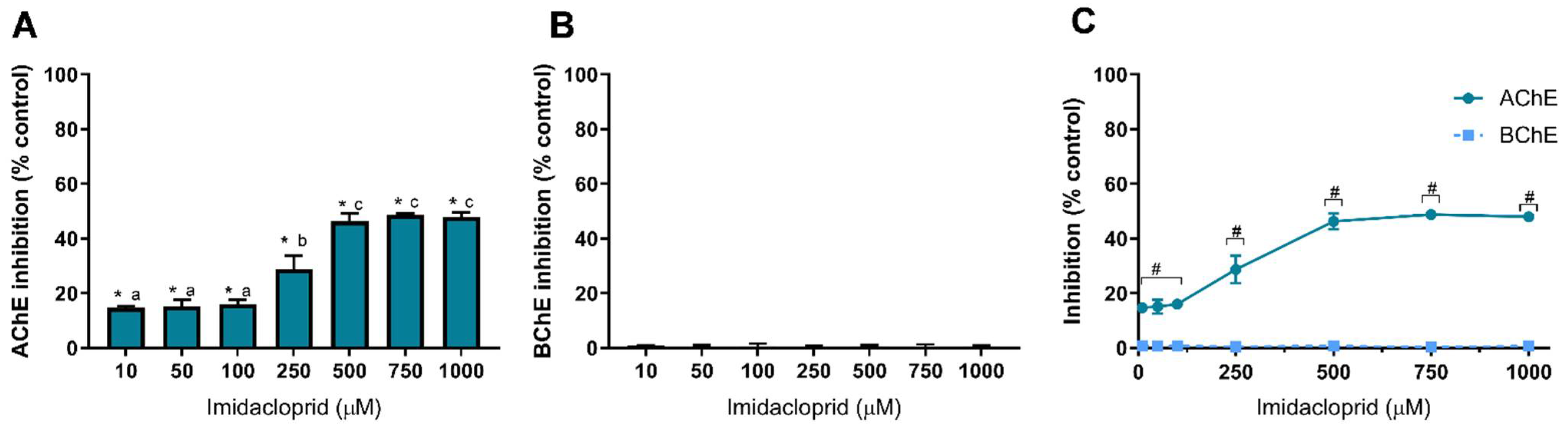
3.1. Pesticide-Induced Cholinesterase Inhibition
3.2. Pesticide-Induced Tyrosinase Inhibition
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Caldas, E.D. Toxicological Aspects of Pesticides. In Sustainable Agrochemistry: A Compendium of Technologies; Sílvio, V., Jr., Ed.; Springer International Publishing: Cham, Switzerland, 2019; pp. 275–305. [Google Scholar]
- Aktar, W.; Sengupta, D.; Chowdhury, A. Impact of pesticides use in agriculture: Their benefits and hazards. Interdiscip. Toxicol. 2009, 2, 1–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martins-Gomes, C.; Silva, T.L.; Andreani, T.; Silva, A.M. Glyphosate vs. Glyphosate-Based Herbicides Exposure: A Review on Their Toxicity. J. Xenobiotics 2022, 12, 21–40. [Google Scholar] [CrossRef] [PubMed]
- Damalas, C.A.; Ilias, G.E. Pesticide exposure, safety issues, and risk assessment indicators. Int. J. Environ. Res. Public Health 2011, 8, 402–419. [Google Scholar] [CrossRef] [PubMed]
- Freire, C.; Koifman, S. Pesticide exposure and Parkinson’s disease: Epidemiological evidence of association. NeuroToxicology 2012, 33, 947–971. [Google Scholar] [CrossRef]
- Tang, B.L. Neuropathological Mechanisms Associated with Pesticides in Alzheimer’s Disease. Toxics 2020, 8, 21. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yan, D.; Zhang, Y.; Liu, L.; Yan, H. Pesticide exposure and risk of Alzheimer’s disease: A systematic review and meta-analysis. Sci. Rep. 2016, 6, 32222. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martínez, M.-A.; Ares, I.; Rodríguez, J.-L.; Martínez, M.; Martínez-Larrañaga, M.-R.; Anadón, A. Neurotransmitter changes in rat brain regions following glyphosate exposure. Environ. Res. 2018, 161, 212–219. [Google Scholar] [CrossRef] [PubMed]
- Limberger, C.; Ferreira, P.C.; Fontella, F.U.; Oliveira, A.C.L.J.; Salles, G.B.; Souza, D.O.; Zimmer, E.R.; De Souza, D.G. Glyphosate-based herbicide alters brain amino acid metabolism without affecting blood-brain barrier integrity. Alzheimer’s Dement. 2020, 16, e043847. [Google Scholar] [CrossRef]
- Kafula, Y.A.; Philippe, C.; Pinceel, T.; Munishi, L.K.; Moyo, F.; Vanschoenwinkel, B.; Brendonck, L.; Thoré, E.S. Pesticide sensitivity of Nothobranchius neumanni, a temporary pond predator with a non-generic life-history. Chemosphere 2021, 291, 132823. [Google Scholar] [CrossRef]
- Martínez, M.A.; Rodríguez, J.L.; Lopez-Torres, B.; Martínez, M.; Martínez-Larrañaga, M.R.; Maximiliano, J.E.; Anadón, A.; Ares, I. Use of human neuroblastoma SH-SY5Y cells to evaluate glyphosate-induced effects on oxidative stress, neuronal development and cell death signaling pathways. Environ. Int. 2020, 135, 105414. [Google Scholar] [CrossRef]
- Jin, Y.; Zhu, Z.; Wang, Y.; Yang, E.; Feng, X.; Fu, Z. The fungicide imazalil induces developmental abnormalities and alters locomotor activity during early developmental stages in zebrafish. Chemosphere 2016, 153, 455–461. [Google Scholar] [CrossRef] [PubMed]
- Heusinkveld, H.J.; Westerink, R.H. Comparison of different in vitro cell models for the assessment of pesticide-induced dopaminergic neurotoxicity. Toxicol. Vitr. 2017, 45, 81–88. [Google Scholar] [CrossRef]
- Tomizawa, M.; Casida, J.E. Molecular Recognition of Neonicotinoid Insecticides: The Determinants of Life or Death. Accounts Chem. Res. 2008, 42, 260–269. [Google Scholar] [CrossRef] [PubMed]
- Abd-Elhakim, Y.M.; Mohammed, H.H.; Mohamed, W.A.M. Imidacloprid Impacts on Neurobehavioral Performance, Oxidative Stress, and Apoptotic Events in the Brain of Adolescent and Adult Rats. J. Agric. Food Chem. 2018, 66, 13513–13524. [Google Scholar] [CrossRef] [PubMed]
- Vohra, P.; Khera, K.S.; Sangha, G.K. Physiological, biochemical and histological alterations induced by administration of imidacloprid in female albino rats. Pestic. Biochem. Physiol. 2014, 110, 50–56. [Google Scholar] [CrossRef]
- Sheets, L.P. Chapter 95—Imidacloprid: A Neonicotinoid Insecticide. In Hayes’ Handbook of Pesticide Toxicology, 3rd ed.; Krieger, R., Ed.; Academic Press: New York, NY, USA, 2010. [Google Scholar]
- Lonare, M.; Kumar, M.; Raut, S.; Badgujar, P.; Doltade, S.; Telang, A. Evaluation of imidacloprid-induced neurotoxicity in male rats: A protective effect of curcumin. Neurochem. Int. 2014, 78, 122–129. [Google Scholar] [CrossRef] [PubMed]
- Tian, X.; Liu, J.; Guo, Z.; Hu, B.; Kibe, M.D.; Wang, S.; Wei, Q.; Su, J. The characteristics of voltage-gated sodium channel and the association with lambda cyhalothrin resistance in Spodoptera exigua. J. Asia-Pacific Èntomol. 2018, 21, 1020–1027. [Google Scholar] [CrossRef]
- Ali, Z.Y. Neurotoxic effect of lambda-cyhalothrin, a synthetic pyrethroid pesticide: Involvement of oxidative stress and protective role of antioxidant mixture. NY Sci. J. 2012, 9, 93–103. [Google Scholar]
- Ansari, R.W.; Shukla, R.K.; Yadav, R.S.; Seth, K.; Pant, A.B.; Singh, D.; Agrawal, A.K.; Islam, F.; Khanna, V.K. Cholinergic Dysfunctions and Enhanced Oxidative Stress in the Neurobehavioral Toxicity of Lambda-Cyhalothrin in Developing Rats. Neurotox. Res. 2012, 22, 292–309. [Google Scholar] [CrossRef]
- Kumar, A.; Rai, D.K.; Sharma, B.; Pandey, R.S. λ-cyhalothrin and cypermethrin induced in vivo alterations in the activity of acetylcholinesterase in a freshwater fish, Channa punctatus (Bloch). Pestic. Biochem. Physiol. 2009, 93, 96–99. [Google Scholar] [CrossRef]
- Pohanka, M. Acetylcholinesterase inhibitors: A patent review (2008–present). Expert Opin. Ther. Patents 2012, 22, 871–886. [Google Scholar] [CrossRef] [PubMed]
- Winek, K.; Soreq, H.; Meisel, A. Regulators of cholinergic signaling in disorders of the central nervous system. J. Neurochem. 2021, 158, 1425–1438. [Google Scholar] [CrossRef] [PubMed]
- Saldanha, C. Human Erythrocyte Acetylcholinesterase in Health and Disease. Molecules 2017, 22, 1499. [Google Scholar] [CrossRef] [Green Version]
- Umar, A.M.; Aisami, A. Acetylcholinesterase Enzyme (AChE) as a Biosensor and Biomarker for Pesticides: A Mini Review. Bull. Environ. Sci. Sustain. Manag. 2020, 4, 7–12. [Google Scholar] [CrossRef]
- Lionetto, M.G.; Caricato, R.; Calisi, A.; Giordano, M.E.; Schettino, T. Acetylcholinesterase as a Biomarker in Environmental and Occupational Medicine: New Insights and Future Perspectives. BioMed Res. Int. 2013, 2013, 321213. [Google Scholar] [CrossRef] [PubMed]
- Lotti, M.; Johnson, M.K. Neurotoxicity of organophosphorus pesticides: Predictions can be based on in vitro studies with hen and human enzymes. Arch. Toxicol. 1978, 41, 215–221. [Google Scholar] [CrossRef] [PubMed]
- Pundir, C.S.; Chauhan, N. Acetylcholinesterase inhibition-based biosensors for pesticide determination: A review. Anal. Biochem. 2012, 429, 19–31. [Google Scholar] [CrossRef]
- Carballo-Carbajal, I.; Laguna, A.; Romero-Giménez, J.; Cuadros, T.; Bové, J.; Martinez-Vicente, M.; Parent, A.; Gonzalez-Sepúlveda, M.; Peñuelas, N.; Torra, A.; et al. Brain tyrosinase overexpression implicates age-dependent neuromelanin production in Parkinson’s disease pathogenesis. Nat. Commun. 2019, 10, 973. [Google Scholar] [CrossRef] [Green Version]
- Wilson, B.W. Chapter 68—Cholinesterases. In Hayes’ Handbook of Pesticide Toxicology, 3rd ed.; Krieger, R., Ed.; Academic Press: New York, NY, USA, 2010. [Google Scholar]
- Mushtaq, G.; Greig, N.H.; Khan, J.; Kamal, M.A. Status of Acetylcholinesterase and Butyrylcholinesterase in Alzheimer’s Disease and Type 2 Diabetes Mellitus. CNS Neurol. Disord.-Drug Targets 2014, 13, 1432–1439. [Google Scholar] [CrossRef]
- Colovic, M.B.; Krstic, D.Z.; Lazarevic-Pasti, T.D.; Bondzic, A.M.; Vasic, V.M. Acetylcholinesterase Inhibitors: Pharmacology and Toxicology. Curr. Neuropharmacol. 2013, 11, 315–335. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Darvesh, S.; Hopkins, D.A.; Geula, C. Neurobiology of butyrylcholinesterase. Nat. Rev. Neurosci. 2003, 4, 131–138. [Google Scholar] [CrossRef]
- Nordberg, A.; Ballard, C.; Bullock, R.; Darreh-Shori, T.; Somogyi, M. A Review of Butyrylcholinesterase as a Therapeutic Target in the Treatment of Alzheimer’s Disease. Prim. Care Companion CNS Disord. 2013, 15, PCC.12r01412. [Google Scholar] [CrossRef]
- Dvir, H.; Silman, I.; Harel, M.; Rosenberry, T.L.; Sussman, J.L. Acetylcholinesterase: From 3D structure to function. Chem. Interact. 2010, 187, 10–22. [Google Scholar] [CrossRef] [Green Version]
- Soares, S.F.D.C.X.; Vieira, A.A.; Delfino, R.T.; Figueroa-Villar, J.D. NMR determination of Electrophorus electricus acetylcholinesterase inhibition and reactivation by neutral oximes. Bioorganic Med. Chem. 2013, 21, 5923–5930. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Boer, D.; Nguyen, N.; Mao, J.; Moore, J.; Sorin, E. A Comprehensive Review of Cholinesterase Modeling and Simulation. Biomolecules 2021, 11, 580. [Google Scholar] [CrossRef] [PubMed]
- Wiesner, J.; Kříž, Z.; Kuča, K.; Jun, D.; Koča, J. Acetylcholinesterases—The structural similarities and differences. J. Enzym. Inhib. Med. Chem. 2007, 22, 417–424. [Google Scholar] [CrossRef] [PubMed]
- Moorad, D.; Chunyuan, L.; Ashima, S.; Bhupendra, D.; Gregory, G.; Moorad, C.L.D.R. Purification and determination of the amino acid sequence of equine serum butyrylcholinesterase. Toxicol. Methods 1999, 9, 219–227. [Google Scholar] [CrossRef]
- Greggio, E.; Bergantino, E.; Carter, D.; Ahmad, R.; Costin, G.-E.; Hearing, V.J.; Clarimón, J.; Singleton, A.; Eerola, J.; Hellstrom, O.; et al. Tyrosinase exacerbates dopamine toxicity but is not genetically associated with Parkinson’s disease. J. Neurochem. 2005, 93, 246–256. [Google Scholar] [CrossRef]
- Ellman, G.L.; Courtney, K.D.; Andres, V., Jr.; Featherstone, R.M. A new and rapid colorimetric determination of acetylcholinesterase activity. Biochem. Pharmacol. 1961, 7, 88–95. [Google Scholar] [CrossRef]
- Taghouti, M.; Martins-Gomes, C.; Schäfer, J.; Félix, L.M.; Santos, J.A.; Bunzel, M.; Nunes, F.M.; Silva, A.M. Thymus pulegioides L. as a rich source of antioxidant, anti-proliferative and neuroprotective phenolic compounds. Food Funct. 2018, 9, 3617–3629. [Google Scholar] [CrossRef]
- Agarwal, P.; Singh, M.; Singh, J.; Singh, R.P. Chapter 1—Microbial Tyrosinases: A Novel Enzyme, Structural Features, and Applications. In Applied Microbiology and Bioengineering; Pratyoosh, S., Ed.; Academic Press: New York, NY, USA, 2019. [Google Scholar]
- Sam, C.; Bordoni, B. Physiology, Acetylcholine. In StatPearls; StatPearls Publishing: Treasure Island, FL, USA, 2022. [Google Scholar]
- Yoshioka, N.; Asano, M.; Kuse, A.; Mitsuhashi, T.; Nagasaki, Y.; Ueno, Y. Rapid determination of glyphosate, glufosinate, bialaphos, and their major metabolites in serum by liquid chromatography-tandem mass spectrometry using hydrophilic interaction chromatography. J. Chromatogr. A 2011, 1218, 3675–3680. [Google Scholar] [CrossRef] [PubMed]
- Louie, F.; Jacobs, N.F.; Yang, L.G.; Park, C.; Monnot, A.D.; Bandara, S.B. A comparative evaluation of dietary exposure to glyphosate resulting from recommended U.S. diets. Food Chem. Toxicol. 2021, 158, 112670. [Google Scholar] [CrossRef] [PubMed]
- Gotti, R.; Fiori, J.; Bosi, S.; Dinelli, G. Field-amplified sample injection and sweeping micellar electrokinetic chromatography in analysis of glyphosate and aminomethylphosphonic acid in wheat. J. Chromatogr. A 2019, 1601, 357–364. [Google Scholar] [CrossRef]
- Vass, A.; Korpics, E.; Dernovics, M. Follow-up of the fate of imazalil from post-harvest lemon surface treatment to a baking experiment. Food Addit. Contam. Part A 2015, 32, 1875–1884. [Google Scholar] [CrossRef] [PubMed]
- Kapoor, U.; Srivastava, M.; Srivastava, A.K.; Patel, D.; Garg, V.; Srivastava, L. Analysis of imidacloprid residues in fruits, vegetables, cereals, fruit juices, and baby foods, and daily intake estimation in and around Lucknow, India. Environ. Toxicol. Chem. 2012, 32, 723–727. [Google Scholar] [CrossRef] [PubMed]
- Silva, A.M.; Martins-Gomes, C.; Silva, T.L.; Coutinho, T.E.; Souto, E.B.; Andreani, T. In Vitro Assessment of Pesticides Toxicity and Data Correlation with Pesticides Physicochemical Properties for Prediction of Toxicity in Gastrointestinal and Skin Contact Exposure. Toxics 2022, 10, 378. [Google Scholar] [CrossRef] [PubMed]
- Silva, A.M.; Martins-Gomes, C.; Ferreira, S.S.; Souto, E.B.; Andreani, T. Molecular Physicochemical Properties of Selected Pesticides as Predictive Factors for Oxidative Stress and Apoptosis-Dependent Cell Death in Caco-2 and HepG2 Cells. Int. J. Mol. Sci. 2022, 23, 8107. [Google Scholar] [CrossRef] [PubMed]
- Samanta, P.; Pal, S.; Mukherjee, A.K.; Ghosh, A.R. Biochemical effects of glyphosate based herbicide, Excel Mera 71 on enzyme activities of acetylcholinesterase (AChE), lipid peroxidation (LPO), catalase (CAT), glutathione-S-transferase (GST) and protein content on teleostean fishes. Ecotoxicol. Environ. Saf. 2014, 107, 120–125. [Google Scholar] [CrossRef] [PubMed]
- Braz-Mota, S.; Sadauskas-Henrique, H.; Duarte, R.M.; Val, A.L.; Almeida-Val, V.M. Roundup® exposure promotes gills and liver impairments, DNA damage and inhibition of brain cholinergic activity in the Amazon teleost fish Colossoma macropomum. Chemosphere 2015, 135, 53–60. [Google Scholar] [CrossRef] [PubMed]
- Glusczak, L.; dos Santos Miron, D.; Crestani, M.; da Fonseca, M.B.; de Araújo Pedron, F.; Duarte, M.F.; Vieira, V.L.P. Effect of glyphosate herbicide on acetylcholinesterase activity and metabolic and hematological parameters in piava (Leporinus obtusidens). Ecotoxicol. Environ. Saf. 2006, 65, 237–241. [Google Scholar] [CrossRef] [PubMed]
- Gholami-Seyedkolaei, S.J.; Mirvaghefi, A.; Farahmand, H.; Kosari, A.A. Effect of a glyphosate-based herbicide in Cyprinus carpio: Assessment of acetylcholinesterase activity, hematological responses and serum biochemical parameters. Ecotoxicol. Environ. Saf. 2013, 98, 135–141. [Google Scholar] [CrossRef] [PubMed]
- Kwiatkowska, M.; Nowacka-Krukowska, H.; Bukowska, B. The effect of glyphosate, its metabolites and impurities on erythrocyte acetylcholinesterase activity. Environ. Toxicol. Pharmacol. 2014, 37, 1101–1108. [Google Scholar] [CrossRef] [PubMed]
- El-Demerdash, F.M.; Yousef, M.I.; Elagamy, E.I. Influence of paraquat, glyphosate, and cadmium on the activity of some serum enzymes and protein electrophoretic behavior (in vitro). J. Environ. Sci. Health Part B 2001, 36, 29–42. [Google Scholar] [CrossRef] [PubMed]
- Takeuchi, I.; Yanagawa, Y.; Nagasawa, H.; Jitsuiki, K.; Madokoro, S.; Takahashi, N.; Ohsaka, H.; Ishikawa, K.; Omori, K. Decrease in Butyrylcholinesterase Accompanied by Intermediate-like Syndrome after Massive Ingestion of a Glyphosate-surfactant. Intern. Med. 2019, 58, 3057–3059. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Geetha, N. Mitigatory role of butyrylcholinesterase in freshwater fish Labeo rohita exposed to glyphosate based herbicide Roundup®. Mater. Today Proc. 2021, 47, 2030–2035. [Google Scholar] [CrossRef]
- Lajmanovich, R.C.; Attademo, A.M.; Peltzer, P.M.; Junges, C.M.; Cabagna, M.C. Toxicity of Four Herbicide Formulations with Glyphosate on Rhinella arenarum (Anura: Bufonidae) Tadpoles: B-esterases and Glutathione S-transferase Inhibitors. Arch. Environ. Contam. Toxicol. 2010, 60, 681–689. [Google Scholar] [CrossRef]
- Saxena, A.; Redman, A.M.; Jiang, X.; Lockridge, O.; Doctor, B.P. Differences in active-site gorge dimensions of cholinesterases revealed by binding of inhibitors to human butyrylcholinesterase. Chem. Interact. 1999, 119–120, 61–69. [Google Scholar] [CrossRef]
- Katić, A.; Kašuba, V.; Kopjar, N.; Lovaković, B.T.; Čermak, A.M.M.; Mendaš, G.; Micek, V.; Milić, M.; Pavičić, I.; Pizent, A.; et al. Effects of low-level imidacloprid oral exposure on cholinesterase activity, oxidative stress responses, and primary DNA damage in the blood and brain of male Wistar rats. Chem.-Biol. Interact. 2021, 338, 109287. [Google Scholar] [CrossRef] [PubMed]
- Topal, A.; Alak, G.; Ozkaraca, M.; Yeltekin, A.C.; Comaklı, S.; Acıl, G.; Kokturk, M.; Atamanalp, M. Neurotoxic responses in brain tissues of rainbow trout exposed to imidacloprid pesticide: Assessment of 8-hydroxy-2-deoxyguanosine activity, oxidative stress and acetylcholinesterase activity. Chemosphere 2017, 175, 186–191. [Google Scholar] [CrossRef] [PubMed]
- Butcherine, P.; Kelaher, B.P.; Benkendorff, K. Assessment of acetylcholinesterase, catalase, and glutathione S-transferase as biomarkers for imidacloprid exposure in penaeid shrimp. Aquat. Toxicol. 2021, 242, 106050. [Google Scholar] [CrossRef]
- Shadnia, S.; Moghaddam, H.H. Fatal intoxication with imidacloprid insecticide. Am. J. Emerg. Med. 2008, 26, 634.e1–634.e4. [Google Scholar] [CrossRef]
- Proença, P.; Teixeira, H.; Castanheira, F.; Pinheiro, J.; Monsanto, P.V.; Marques, E.P.; Vieira, D.N. Two fatal intoxication cases with imidacloprid: LC/MS analysis. Forensic Sci. Int. 2005, 153, 75–80. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, A.; Mahai, G.; Wan, Y.; Yang, Z.; He, Z.; Xu, S.; Xia, W. Assessment of imidacloprid related exposure using imidacloprid-olefin and desnitro-imidacloprid: Neonicotinoid insecticides in human urine in Wuhan, China. Environ. Int. 2020, 141, 105785. [Google Scholar] [CrossRef]
- Dankyi, E.; Gordon, C.; Carboo, D.; Fomsgaard, I.S. Quantification of neonicotinoid insecticide residues in soils from cocoa plantations using a QuEChERS extraction procedure and LC-MS/MS. Sci. Total Environ. 2014, 499, 276–283. [Google Scholar] [CrossRef] [PubMed]
- Kittelmann, A.; Müller, C.; Rohn, S.; Michalski, B. Transfer of Pesticide Residues from Grapes (Vitis vinifera) into Wine—Correlation with Selected Physicochemical Properties of the Active Substances. Toxics 2022, 10, 248. [Google Scholar] [CrossRef] [PubMed]
- Khan, M.Z.; Zaheer, M.; Fatima, F. Effect of lambda cyhalothrin (pyrethroid) and monocrotophos (organophosphate) on cholinesterase activity in liver, kidney and brain of Rana cyanophlyctis. Korean J. Biol. Sci. 2003, 7, 165–168. [Google Scholar] [CrossRef]
- Eisenhofer, G.; Tian, H.; Holmes, C.; Matsunaga, J.; Roffler-Tarlov, S.; Hearing, V.J. Tyrosinase: A developmentally specific major determinant of peripheral dopamine. FASEB J. 2003, 17, 1248–1255. [Google Scholar] [CrossRef]
- Smith, D.F.Q.; Camacho, E.; Thakur, R.; Barron, A.J.; Dong, Y.; Dimopoulos, G.; Broderick, N.A.; Casadevall, A. Glyphosate inhibits melanization and increases susceptibility to infection in insects. PLoS Biol. 2021, 19, e3001182. [Google Scholar] [CrossRef] [PubMed]
- Ford, K.A.; Gulevich, A.G.; Swenson, T.L.; Casida, J.E. Neonicotinoid Insecticides: Oxidative Stress in Planta and Metallo-oxidase Inhibition. J. Agric. Food Chem. 2011, 59, 4860–4867. [Google Scholar] [CrossRef]
- Şişman, T.; Türkez, H. Toxicologic evaluation of imazalil with particular reference to genotoxic and teratogenic potentials. Toxicol. Ind. Health 2010, 26, 641–648. [Google Scholar] [CrossRef] [PubMed]
- Tang, F.; Shen, X.; Gao, X.-W. In Vitro Inhibition of the Diphenolase Activity of Tyrosinase by Insecticides and Allelochemicals in Micromelalopha troglodyta (Lepidoptera: Notodontidae). J. Èntomol. Sci. 2009, 44, 111–119. [Google Scholar] [CrossRef]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Martins-Gomes, C.; Coutinho, T.E.; Silva, T.L.; Andreani, T.; Silva, A.M. Neurotoxicity Assessment of Four Different Pesticides Using In Vitro Enzymatic Inhibition Assays. Toxics 2022, 10, 448. https://doi.org/10.3390/toxics10080448
Martins-Gomes C, Coutinho TE, Silva TL, Andreani T, Silva AM. Neurotoxicity Assessment of Four Different Pesticides Using In Vitro Enzymatic Inhibition Assays. Toxics. 2022; 10(8):448. https://doi.org/10.3390/toxics10080448
Chicago/Turabian StyleMartins-Gomes, Carlos, Tiago E. Coutinho, Tânia L. Silva, Tatiana Andreani, and Amélia M. Silva. 2022. "Neurotoxicity Assessment of Four Different Pesticides Using In Vitro Enzymatic Inhibition Assays" Toxics 10, no. 8: 448. https://doi.org/10.3390/toxics10080448
APA StyleMartins-Gomes, C., Coutinho, T. E., Silva, T. L., Andreani, T., & Silva, A. M. (2022). Neurotoxicity Assessment of Four Different Pesticides Using In Vitro Enzymatic Inhibition Assays. Toxics, 10(8), 448. https://doi.org/10.3390/toxics10080448