

Genetic Diversity of Microcystin Producers (Cyanobacteria) and Microcystin Congeners in Aquatic Resources across Africa: A Review Paper

 , , ,
, , ,
Abstract
1. Introduction
2. Toxicosis of Microcystins in Africa
3. The Taxonomic Diversity of African Cyanobacteria Using Genetic Markers

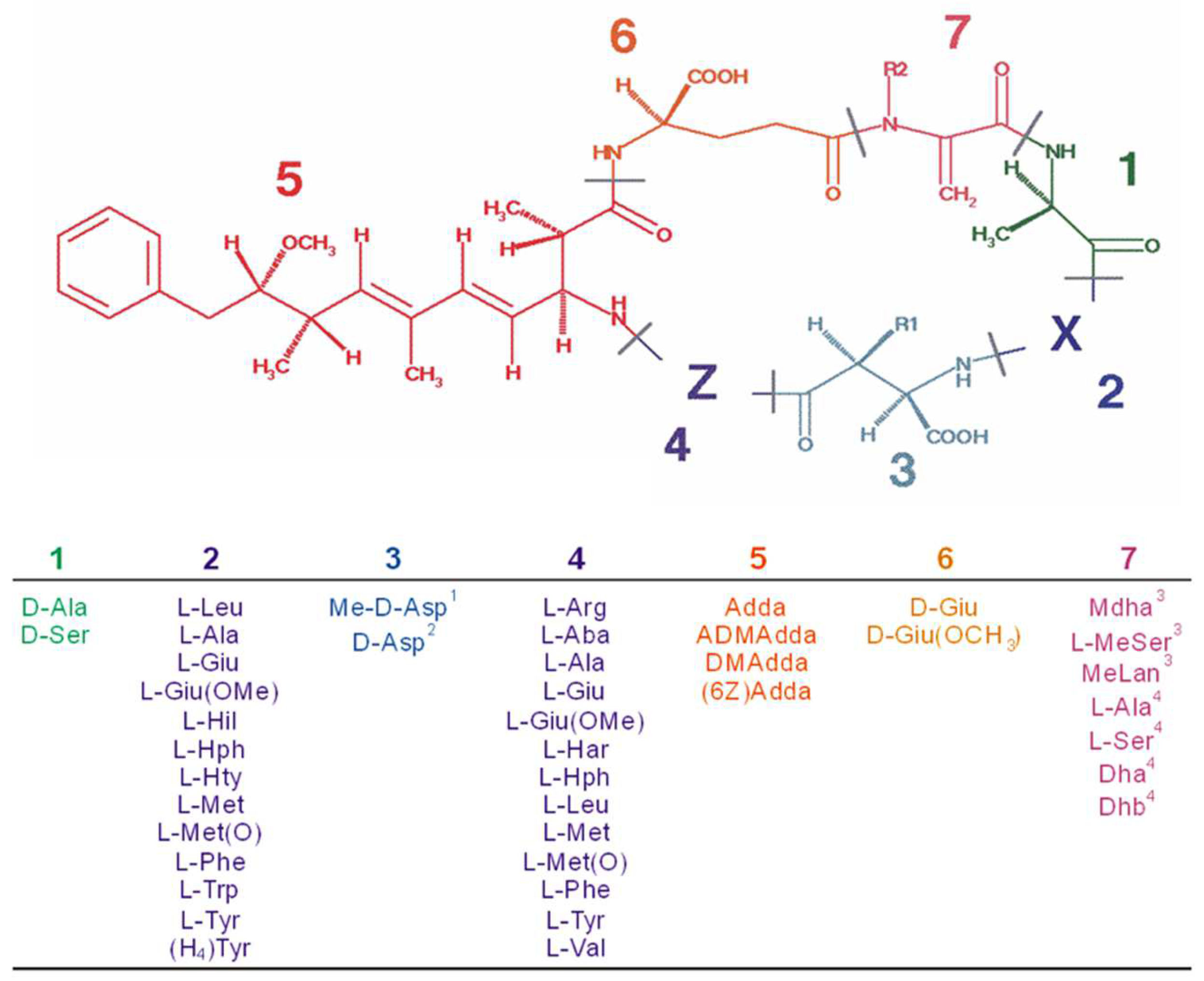{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sub-Region | Country/Sampling Sites | Identified/Isolated Species | Number of Strains/Sequences Analyzed | Method of Identification | Taxonomic Genetic Marker and/or Molecular Technique | Microcystin Detection—Molecular Markers | Microcystin Detection—Chemical Analysis | Location of Closer Related spp. /Genotype/Strain | Reference |
|---|---|---|---|---|---|---|---|---|---|
| North Africa | Wadi El-Natrun, Egypt | Spirulina, Oscillatoria, Microcystis | NA | SEM, Light microscopy | 16S rDNA, SDS-PAGE | 28 a | NA | NA | [28] |
| Tunisia | Microcystis aeruginosa, Raphidiopsis raciborskii, Planktothrix agardhii | 27 | NA | 16S–23S rRNA ITS, rpoC1 | NA | NA | South Africa, Sweden, and USA strains. Toxic Micro-HJ-02 strain clustered with a French strain | [31] | |
| BirM’cherga reservoir, Tunisia | Raphidiopsis raciborskii | 4 | NA | 16S rRNA, rpoC1 | mcyA, mcyE | HPLC and MALDI-TOF | Distinct from other African strains | [25] | |
| East Africa | Lake Bogori, Kenya | Arthrospira fusiformis, Anabaenopsis, Synechococcus, Arthrospira, Spirulina, Oscillatoria, Leptolyngbya | NA | Light microscopy | Multilocus 454- sequencing (16S rRNA and (cpcBA-IGS, Metagenomics | NA | NA | NA | [29,32] |
| Kenya, Tanzania, Uganda | Arthrospira | 18 | Width of trichome, types of helix, and apical shape of trichomes | 16S-23S ITS (31 *), cpcBA-IGS (23 *) | NA | NA | India, Asia | [33] | |
| Kenya, Uganda | Microcystis aeruginosa | 17 (8) | Light microscopy | PC-IGS, ITS1 rDNA | 4 (3 MC producing) | NA | Australia, Brazil | [34] | |
| Uganda | Raphidiopsis raciborskii | (44 **) | Morphological traits | ITS1, PC-IGS, nifH, rpoC1 | NA | NA | Germany | [35] | |
| Sediments and plankton of saline-alkaline and freshwater lakes of Kenya | Anabaenopsis, Umezakia, Arthrospira sp., Chroococcidiopsis sp. | Nucleotide sequences (8) from DGGE bands: Nostocales (5), Pleurocapsales (1), Oscillatoriales (2) | BLAST search analysis | PCR-based denaturing gradient gel electrophoresis (DGGE) analysis of 16S rRNA gene fragments | mcyE and ndaF | NA | Kenya | [33] | |
| Ugandan freshwaters | Anabaena, Aphanocapsa, Chroococcus, Merismopedia, Microcystis, Planktolyngbia, and Pseudanabaena | NA | Morphological keys | PC-IGS | mcyB and mcyE | HPLC-DAD, MALDI-TOF MS, Q-TOF | Uganda | [36] | |
| West Africa | Rivers, lagoons, and coastal waters, Nigeria | Microcystis aeruginosa, Planktothrix aghardii, Anabaena sp. | NA | Light microscopy | NA | mcyE | Abraxis microcystins /Nodularin Adda Kit (Abraxis Inc., Warminster, PA, USA) | Southern Germany, Korea, Japan | [12] |
| Freshwater lakes and reservoir, Senegal | Raphidiopsis raciborskii, Raphidiopsis africana Raphidiopsis sp. | 18 | Light microscopy, the ability to differentiate heterocysts when grown in a nitrogen-free medium | 16S rRNA, ITS1-S, ITS1-L, rpoCl, nifH [10 *] | NA | NA | America, Europe | [27] | |
| Southern Africa | Hartbeespoort Dam, South Africa | Sphaerospermopsis reniformis, Sphaerospermopsis aphanizomenoides, Raphidiopsis curvispora, Raphidiopsis curvata, Raphidiopsis mediterrranea, Microcystis aeruginosa | 27 (1 MC producing) | Light microscopy | NA | mcyE 41 b (35) MC (1 d; MC-AnaR) | NA | Tropical and subtropical regions of Africa | [26] |
| Crusts Karoo and Nama Karoo, South Africa | Consortia of cyanobacteria | 45 | Morphological features | 16S rRNA (140 *) | 103 a (11 d) | NA | NA | [37] | |
| South Africa | Microcystis aeruginosa | 23 | Light microscopy | AFLPs | NA | NA | Japan, Europe, North America | [38] | |
| Theewaterskloof Dam, South Africa | Anabaena ucrainica | NA | SEM | 16S rRNA | NA | NA | NA | [39] | |
| Phakalane ponds, Botswana | Microcystis novacekii | NA | Microscopic identification BLAST search analysis | 16S rRNA | mcyA, -B, -C, -D, -E and –G | LC-ESI-MS and solid-phase extraction (SPE) steps | Botswana | [40] |
3.1. 16S Ribosomal RNA and 16S–23S rDNA Internal Transcribed Spacer (ITS)
3.2. Amplified Fragment Length Polymorphism
3.3. Omics—Metagenomics for Characterization of Cyanobacteria in Africa
4. Detection of Potentially African Microcystin-Producing Cyanobacteria by Molecular Markers
5. Cyanobacterial Abundance and Microcystin Congeners Detected in African Waters
5.1. East Africa
5.2. North Africa
5.3. South Africa
5.4. West Africa
6. Relationship between Genetic Characterization and Geographical Origin within Toxic Cyanobacteria Strains across Africa
7. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Buratti, F.M.; Manganelli, M.; Vichi, S.; Stefanelli, M.; Scardala, S.; Testai, E.; Funari, E. Cyanotoxins: Producing Organisms, Occurrence, Toxicity, Mechanism of Action and Human Health Toxicological Risk Evaluation. Arch. Toxicol. 2017, 91, 1049–1130. [Google Scholar] [CrossRef] [PubMed]
- Paerl, H.W.; Huisman, J. Climate Change: A Catalyst for Global Expansion of Harmful Cyanobacterial Blooms. Environ. Microbiol. Rep. 2009, 1, 27–37. [Google Scholar] [CrossRef] [PubMed]
- Funari, E.; Testai, E. Human Health Risk Assessment Related to Cyanotoxins Exposure. Crit. Rev. Toxicol. 2008, 38, 97–125. [Google Scholar] [CrossRef] [PubMed]
- Puschner, B.; Hoff, B.; Tor, E.R. Diagnosis of Anatoxin-a Poisoning in Dogs from North America. J. Vet. Diagn. Investig. 2008, 20, 89–92. [Google Scholar] [CrossRef] [PubMed]
- Metcalf, J.S.; Codd, G.A. Cyanotoxins. In Ecology of Cyanobacteria II; Springer: Berlin/Heidelberg, Germany, 2012; pp. 651–675. [Google Scholar]
- Codd, G.A.; Azevedo, S.; Bagchi, M.D.; Burch, M.D.; Carmichael, W.W.; Harding, W.R.; Kaya, K.; Utkilen, H.C. CYANONET: A Global Network for Cyanobacterial Bloom and Toxin Risk Management: Initial Situation Assessment and Recommendations; UNESCO: Paris, France, 2005. [Google Scholar]
- Ndlela, L.L.; Oberholster, P.J.; Van Wyk, J.H.; Cheng, P.-H. An Overview of Cyanobacterial Bloom Occurrences and Research in Africa over the Last Decade. Harmful Algae 2016, 60, 11–26. [Google Scholar] [CrossRef]
- Moreira, C.; Vasconcelos, V.; Antunes, A. Phylogeny and Biogeography of Cyanobacteria and Their Produced Toxins. Mar. Drugs 2013, 11, 4350–4369. [Google Scholar] [CrossRef]
- Fewer, D.P.; Wahlsten, M.; Österholm, J.; Jokela, J.; Rouhiainen, L.; Kaasalainen, U.; Rikkinen, J.; Sivonen, K. The Genetic Basis for O-Acetylation of the Microcystin Toxin in Cyanobacteria. Chem. Biol. 2013, 20, 861–869. [Google Scholar] [CrossRef]
- Malik, J.K.; Bharti, V.K.; Rahal, A.; Kumar, D.; Gupta, R.C. Cyanobacterial (Blue-Green Algae) Toxins. In Handbook of Toxicology of Chemical Warfare Agents; Elsevier: Amsterdam, The Netherlands, 2020; pp. 467–478. [Google Scholar]
- Roy-Lachapelle, A.; Solliec, M.; Sauvé, S.; Gagnon, C. A Data-Independent Methodology for the Structural Characterization of Microcystins and Anabaenopeptins Leading to the Identification of Four New Congeners. Toxins 2019, 11, 619. [Google Scholar] [CrossRef]
- Kadiri, M.O.; Isagba, S.; Ogbebor, J.U.; Omoruyi, O.A.; Unusiotame-Owolagba, T.E.; Lorenzi, A.S.; Bittencourt-Oliveira, M.; Chia, M.A. The Presence of Microcystins in the Coastal Waters of Nigeria, from the Bights of Bonny and Benin, Gulf of Guinea. Environ. Sci. Pollut. Res. 2020, 27, 35284–35293. [Google Scholar] [CrossRef]
- Butler, N.; Carlisle, J.C.; Linville, R.; Washburn, B. Microcystins: A Brief Overview of Their Toxicity and Effects, with Special Reference to Fish, Wildlife, and Livestock; Ecotoxicology Program Integrated Risk Assessment Branch; Office of Environmental Health Hazard Assessment California Environmental Protection Agency: Sacramento, CA, USA, 2009; Volume 5. [Google Scholar]
- Hastie, C.J.; Borthwick, E.B.; Morrison, L.F.; Codd, G.A.; Cohen, P.T.W. Inhibition of Several Protein Phosphatases by a Non-Covalently Interacting Microcystin and a Novel Cyanobacterial Peptide, Nostocyclin. Biochim. Biophys. Acta (BBA)-Gen. Subj. 2005, 1726, 187–193. [Google Scholar] [CrossRef]
- Maynes, J.T.; Luu, H.A.; Cherney, M.M.; Andersen, R.J.; Williams, D.; Holmes, C.F.B.; James, M.N.G. Crystal Structures of Protein Phosphatase-1 Bound to Motuporin and Dihydromicrocystin-LA: Elucidation of the Mechanism of Enzyme Inhibition by Cyanobacterial Toxins. J. Mol. Biol. 2006, 356, 111–120. [Google Scholar] [CrossRef]
- Arman, T.; Clarke, J.D. Microcystin Toxicokinetics, Molecular Toxicology, and Pathophysiology in Preclinical Rodent Models and Humans. Toxins 2021, 13, 537. [Google Scholar] [CrossRef]
- Humbert, J.F. Toxins of Cyanobacteria. In Handbook of Toxicology of Chemical Warfare Agents; Academic Press: Cambridge, MA, USA, 2009. [Google Scholar] [CrossRef]
- Mrdjen, I.; Morse, M.A.; Ruch, R.J.; Knobloch, T.J.; Choudhary, S.; Weghorst, C.M.; Lee, J. Impact of Microcystin-LR on Liver Function Varies by Dose and Sex in Mice. Toxins 2018, 10, 435. [Google Scholar] [CrossRef]
- Chen, L.; Giesy, J.P.; Adamovsky, O.; Svirčev, Z.; Meriluoto, J.; Codd, G.A.; Mijovic, B.; Shi, T.; Tuo, X.; Li, S.-C. Challenges of Using Blooms of Microcystis Spp. in Animal Feeds: A Comprehensive Review of Nutritional, Toxicological and Microbial Health Evaluation. Sci. Total Environ. 2021, 764, 142319. [Google Scholar] [CrossRef]
- Van Halderen, A.; Harding, W.R.; Wessels, J.C.; Schneider, D.J.; Heine, E.W.P.; Van der Merwe, J.; Fourie, J.M. Cyanobacterial (Blue-Green Algae) Poisoning of Livestock in the Western Cape Province of South Africa. J. S. Afr. Vet. Assoc. 1995, 66, 260–264. [Google Scholar]
- Puschner, B.; Moore, C. Cyanobacteria. In Small Animal Toxicology, 3rd. ed; Elsevier Inc.: Amsterdam, The Netherlands, 2012; pp. 533–540. [Google Scholar]
- Krienitz, L.; Ballot, A.; Kotut, K.; Wiegand, C.; Pütz, S.; Metcalf, J.S.; Codd, G.A.; Stephan, P. Contribution of Hot Spring Cyanobacteria to the Mysterious Deaths of Lesser Flamingos at Lake Bogoria, Kenya. FEMS Microbiol. Ecol. 2003, 43, 141–148. [Google Scholar] [CrossRef]
- Miles, C.O.; Sandvik, M.; Nonga, H.E.; Rundberget, T.; Wilkins, A.L.; Rise, F.; Ballot, A. Identification of Microcystins in a Lake Victoria Cyanobacterial Bloom Using LC–MS with Thiol Derivatization. Toxicon 2013, 70, 21–31. [Google Scholar] [CrossRef]
- Masango, M.G.; Myburgh, J.G.; Labuschagne, L.; Govender, D.; Bengis, R.G.; Naicker, D. Assessment of Microcystis Bloom Toxicity Associated with Wildlife Mortality in the Kruger National Park, South Africa. J. Wildl. Dis. 2010, 46, 95–102. [Google Scholar] [CrossRef]
- Fathalli, A.; Jenhani, A.B.R.; Moreira, C.; Azevedo, J.; Welker, M.; Romdhane, M.; Antunes, A.; Vasconcelos, V. Genetic Variability of the Invasive Cyanobacteria Cylindrospermopsis Raciborskii from Bir M’cherga Reservoir (Tunisia). Arch. Microbiol. 2011, 193, 595–604. [Google Scholar] [CrossRef]
- Ballot, A.; Sandvik, M.; Rundberget, T.; Botha, C.J.; Miles, C.O. Diversity of Cyanobacteria and Cyanotoxins in Hartbeespoort Dam, South Africa. Mar. Freshw. Res. 2013, 65, 175–189. [Google Scholar] [CrossRef]
- Gugger, M.; Molica, R.; Le Berre, B.; Dufour, P.; Bernard, C.; Humbert, J.-F. Genetic Diversity of Cylindrospermopsis Strains (Cyanobacteria) Isolated from Four Continents. Appl. Environ. Microbiol. 2005, 71, 1097–1100. [Google Scholar] [CrossRef] [PubMed]
- Hoballah, E.M.; Attallah, A.G.; Abd-El-Aal, S.K. Genetic Diversity of Some New Local Strains of Cyanobacteria Isolated from Wadi El-Natrun, Egypt. Int. J. Acad. Res. 2012, 4, 314–326. [Google Scholar]
- Krienitz, L.; Dadheech, P.K.; Fastner, J.; Kotut, K. The Rise of Potentially Toxin Producing Cyanobacteria in Lake Naivasha, Great African Rift Valley, Kenya. Harmful Algae 2013, 27, 42–51. [Google Scholar] [CrossRef]
- Konur, O. The Pioneering Research on the Cyanobacterial Photosystems and Photosynthesis. In Handbook of Algal Science, Technology and Medicine; Elsevier: Amsterdam, The Netherlands, 2020; pp. 231–243. [Google Scholar]
- Moreira, C.; Fathalli, A.; Vasconcelos, V.; Antunes, A. Genetic Diversity and Structure of the Invasive Toxic Cyanobacterium Cylindrospermopsis Raciborskii. Curr. Microbiol. 2011, 62, 1590–1595. [Google Scholar] [CrossRef] [PubMed]
- Dadheech, P.K.; Glöckner, G.; Casper, P.; Kotut, K.; Mazzoni, C.J.; Mbedi, S.; Krienitz, L. Cyanobacterial Diversity in the Hot Spring, Pelagic and Benthic Habitats of a Tropical Soda Lake. FEMS Microbiol. Ecol. 2013, 85, 389–401. [Google Scholar] [CrossRef]
- Dadheech, P.K.; Ballot, A.; Casper, P.; Kotut, K.; Novelo, E.; Lemma, B.; Pröschold, T.; Krienitz, L. Phylogenetic Relationship and Divergence among Planktonic Strains of Arthrospira (Oscillatoriales, Cyanobacteria) of African, Asian and American Origin Deduced by 16S–23S ITS and Phycocyanin Operon Sequences. Phycologia 2010, 49, 361–372. [Google Scholar] [CrossRef]
- Haande, S.; Ballot, A.; Rohrlack, T.; Fastner, J.; Wiedner, C.; Edvardsen, B. Diversity of Microcystis Aeruginosa Isolates (Chroococcales, Cyanobacteria) from East-African Water Bodies. Arch. Microbiol. 2007, 188, 15–25. [Google Scholar] [CrossRef]
- Haande, S.; Rohrlack, T.; Ballot, A.; Røberg, K.; Skulberg, R.; Beck, M.; Wiedner, C. Genetic Characterisation of Cylindrospermopsis Raciborskii (Nostocales, Cyanobacteria) Isolates from Africa and Europe. Harmful Algae 2008, 7, 692–701. [Google Scholar] [CrossRef]
- Okello, W.; Kurmayer, R. Seasonal Development of Cyanobacteria and Microcystin Production in Ugandan Freshwater Lakes. Lakes Reserv. Res. Manag. 2011, 16, 123–135. [Google Scholar] [CrossRef]
- Dojani, S.; Kauff, F.; Weber, B.; Büdel, B. Genotypic and Phenotypic Diversity of Cyanobacteria in Biological Soil Crusts of the Succulent Karoo and Nama Karoo of Southern Africa. Microb. Ecol. 2014, 67, 286–301. [Google Scholar] [CrossRef]
- Oberholster, P.J.; Botha, A.-M.; Cloete, T.E. An Overview of Toxic Freshwater Cyanobacteria in South Africa with Special Reference to Risk, Impact and Detection by Molecular Marker Tools. Biokemistri 2005, 17, 57–71. [Google Scholar]
- Oberholster, P.J.; Jappie, S.; Cheng, P.H.; Botha, A.M.; Matthews, M.W. First Report of an Anabaena Bory Strain Containing Microcystin-LR in a Freshwater Body in Africa First Report of an Anabaena Bory Strain Containing Microcystin-LR in a Freshwater Body in Africa. Afr. J. Aquat. Sci. 2015, 40, 37–41. [Google Scholar] [CrossRef]
- Mbukwa, E.; Msagati, T.A.; Mamba, B.B.; Boussiba, S.; Wepener, V.; Leu, S.; Kaye, Y. Toxic Microcystis Novacekii T20-3 from Phakalane Ponds, Botswana: PCR Amplifications of Microcystin Synthetase (Mcy) Genes, Extraction and LC-ESI-MS Identification of Microcystins. J. Environ. Anal. Toxicol. 2015, 525–2161. [Google Scholar]
- Woese, C.R.; Fox, G.E. Phylogenetic Structure of the Prokaryotic Domain: The Primary Kingdoms. Proc. Natl. Acad. Sci. USA 1977, 74, 5088–5090. [Google Scholar] [CrossRef]
- Wuyts, J.; Van de Peer, Y.; De Wachter, R. Distribution of Substitution Rates and Location of Insertion Sites in the Tertiary Structure of Ribosomal RNA. Nucleic Acids Res. 2001, 29, 5017–5028. [Google Scholar] [CrossRef]
- Wilson, K.M.; Schembri, M.A.; Baker, P.D.; Saint, C.P. Molecular Characterization of the Toxic Cyanobacterium Cylindrospermopsis Raciborskii and Design of a Species-Specific PCR. Appl. Environ. Microbiol. 2000, 66, 332–338. [Google Scholar] [CrossRef]
- Padisak, J. Cylindrospermopsis Raciborskii (Woloszynska) Seenayya et Subba Raju, an Expanding, Highly Adaptive Cyanobacterium: Worldwide Distribution and Review of Its Ecology. Arch. Für Hydrobiol. Suppl. Monogr. Beitrage 1997, 107, 563–593. [Google Scholar]
- Neilan, B.A.; Saker, M.L.; Fastner, J.; Törökné, A.; Burns, B.P. Phylogeography of the Invasive Cyanobacterium Cylindrospermopsis Raciborskii. Mol. Ecol. 2003, 12, 133–140. [Google Scholar] [CrossRef]
- van Gremberghe, I.; Van der Gucht, K.; Vanormelingen, P.; Asmelash, T.; Dejenie, T.; D’hondt, S.; Declerck, S.; De Meester, L.; Vyverman, W. Genetic Diversity of Microcystis Blooms (Cyanobacteria) in Recently Constructed Reservoirs in Tigray (Northern Ethiopia) Assessed by RDNA ITS. Aquat. Ecol. 2011, 45, 289–306. [Google Scholar] [CrossRef]
- Moreira, C.; Spillane, C.; Fathalli, A.; Vasconcelos, V.; Antunes, A. African Origin and Europe-Mediated Global Dispersal of the Cyanobacterium Microcystis Aeruginosa. Curr. Microbiol. 2014, 69, 628–633. [Google Scholar] [CrossRef] [PubMed]
- Palinska, K.A.; Liesack, W.; Rhiel, E.; Krumbein, W.E. Phenotype Variability of Identical Genotypes: The Need for a Combined Approach in Cyanobacterial Taxonomy Demonstrated on Merismopedia-like Isolates. Arch. Microbiol. 1996, 166, 224–233. [Google Scholar] [CrossRef] [PubMed]
- Tillett, D.; Dittmann, E.; Erhard, M.; Von Döhren, H.; Börner, T.; Neilan, B.A. Structural Organization of Microcystin Biosynthesis in Microcystis Aeruginosa PCC7806: An Integrated Peptide-Polyketide Synthetase System. Chem. Biol. 2000, 7, 753–764. [Google Scholar] [CrossRef] [PubMed]
- Rouhiainen, L.; Vakkilainen, T.; Siemer, B.L.; Buikema, W.; Haselkorn, R.; Sivonen, K. Genes Coding for Hepatotoxic Heptapeptides (Microcystins) in the Cyanobacterium Anabaena Strain 90. Appl. Environ. Microbiol. 2004, 70, 686–692. [Google Scholar] [CrossRef] [PubMed]
- Christiansen, G.; Kurmayer, R.; Liu, Q.; Börner, T. Transposons Inactivate Biosynthesis of the Nonribosomal Peptide Microcystin in Naturally Occurring Planktothrix spp. Appl. Environ. Microbiol. 2006, 72, 117–123. [Google Scholar] [CrossRef] [PubMed]
- Heck, K.; Alvarenga, D.O.; Shishido, T.K.; Varani, A.M.; Dörr, F.A.; Pinto, E.; Rouhiainen, L.; Jokela, J.; Sivonen, K.; Fiore, M.F. Biosynthesis of Microcystin Hepatotoxins in the Cyanobacterial Genus Fischerella. Toxicon 2018, 141, 43–50. [Google Scholar] [CrossRef]
- Mbukwa, E.A.; Msagati, T.A.M.; Mamba, B.B. Quantitative Variations of Intracellular Microcystin-LR,-RR and-YR in Samples Collected from Four Locations in Hartbeespoort Dam in North West Province (South Africa) during the 2010/2011 Summer Season. Int. J. Environ. Res. Public Health 2012, 9, 3484–3505. [Google Scholar] [CrossRef]
- Kurmayer, R.; Blom, J.F.; Deng, L.; Pernthaler, J. Integrating Phylogeny, Geographic Niche Partitioning and Secondary Metabolite Synthesis in Bloom-Forming Planktothrix. ISME J. 2015, 9, 909–921. [Google Scholar] [CrossRef]
- Douma, M.; Ouahid, Y.; Loudiki, M.; Del Campo, F.F.; Oudra, B. The First Detection of Potentially Toxic Microcystis Strains in Two Middle Atlas Mountains Natural Lakes (Morocco). Environ. Monit. Assess. 2017, 189, 1–9. [Google Scholar] [CrossRef]
- Fathalli, A.; Jenhani, A.B.R.; Moreira, C.; Welker, M.; Romdhane, M.; Antunes, A.; Vasconcelos, V. Molecular and Phylogenetic Characterization of Potentially Toxic Cyanobacteria in Tunisian Freshwaters. Syst. Appl. Microbiol. 2011, 34, 303–310. [Google Scholar] [CrossRef]
- Oberholster, P.J.; Botha, A.-M.; Ashton, P.J. The Influence of a Toxic Cyanobacterial Bloom and Water Hydrology on Algal Populations and Macroinvertebrate Abundance in the Upper Littoral Zone of Lake Krugersdrift, South Africa. Ecotoxicology 2009, 18, 34–46. [Google Scholar] [CrossRef]
- Bouhaddada, R.; Nélieu, S.; Nasri, H.; Delarue, G.; Bouaïcha, N. High Diversity of Microcystins in a Microcystis Bloom from an Algerian Lake. Environ. Pollut. 2016, 216, 836–844. [Google Scholar] [CrossRef]
- Nasri, H.; El Herry, S.; Bouaïcha, N. First Reported Case of Turtle Deaths during a Toxic Microcystis Spp. Bloom in Lake Oubeira, Algeria. Ecotoxicol. Environ. Saf. 2008, 71, 535–544. [Google Scholar] [CrossRef]
- Nasri, A.-B.; Bouaïcha, N.; Fastner, J. First Report of a Microcystin-Containing Bloom of the Cyanobacteria Microcystis Spp. in Lake Oubeira, Eastern Algeria. Arch. Environ. Contam. Toxicol. 2004, 46, 197–202. [Google Scholar] [CrossRef]
- Nasri, H.; Bouaïcha, N.; Harche, M.K. A New Morphospecies of Microcystis Sp. Forming Bloom in the Cheffia Dam (Algeria): Seasonal Variation of Microcystin Concentrations in Raw Water and Their Removal in a Full-scale Treatment Plant. Environ. Toxicol. Int. J. 2007, 22, 347–356. [Google Scholar] [CrossRef]
- Brittain, S.; Mohamed, Z.A.; Wang, J.; Lehmann, V.K.B.; Carmichael, W.W.; Rinehart, K.L. Isolation and Characterization of Microcystins from a River Nile Strain of Oscillatoria Tenuis Agardh Ex Gomont. Toxicon 2000, 38, 1759–1771. [Google Scholar] [CrossRef]
- Mohamed, Z.A.; Carmichael, W.W. Seasonal Variation in Microcystin Levels of River Nile Water at Sohag City, Egypt. Int. J. Limnol. 2000, 36, 227–234. [Google Scholar] [CrossRef]
- Mohamed, Z.A. Accumulation of Cyanobacterial Hepatotoxins by Daphnia in Some Egyptian Irrigation Canals. Ecotoxicol. Environ. Saf. 2001, 50, 4–8. [Google Scholar] [CrossRef]
- Mohamed, Z.A.; Deyab, M.A.; Abou-Dobara, M.I.; El-Sayed, A.K.; El-Raghi, W.M. Occurrence of Cyanobacteria and Microcystin Toxins in Raw and Treated Waters of the Nile River, Egypt: Implication for Water Treatment and Human Health. Environ. Sci. Pollut. Res. 2015, 22, 11716–11727. [Google Scholar] [CrossRef]
- Mohamed, Z.A. First Report of Toxic Cylindrospermopsis Raciborskii and Raphidiopsis Mediterranea (Cyanoprokaryota) in Egyptian Fresh Waters. FEMS Microbiol. Ecol. 2007, 59, 749–761. [Google Scholar] [CrossRef]
- Amer, R.; Díez, B.; El-Shehawy, R. Diversity of Hepatotoxic Cyanobacteria in the Nile Delta, Egypt. J. Environ. Monit. 2009, 11, 126–133. [Google Scholar] [CrossRef]
- Oudra, B.; El Andaloussi, M.; Franca, S.; Barros, P.; Martins, R.; Oufdou, K.; Sbiyyaa, B.; Loudiki, M.; Mezrioui, N.; Vasconcelos, V. Harmful Cyanobacterial Toxic Blooms in Waste Stabilisation Ponds. Water Sci. Technol. 2000, 42, 179–186. [Google Scholar] [CrossRef]
- Oudra, B.; Loudiki, M.; Sbiyyaa, B.; Martins, R.; Vasconcelos, V.; Namikoshi, N. Isolation, Characterization and Quantification of Microcystins (Heptapeptides Hepatotoxins) in Microcystis Aeruginosa Dominated Bloom of Lalla Takerkoust Lake–Reservoir (Morocco). Toxicon 2001, 39, 1375–1381. [Google Scholar] [CrossRef] [PubMed]
- Douma, M.; Manaut, N.; Oudra, B.; Loudiki, M. First Report of Cyanobacterial Diversity and Microcystins in a Microcystis Strain from Sidi Boughaba, a Moroccan Coastal Lagoon. Afr. J. Aquat. Sci. 2016, 41, 445–452. [Google Scholar] [CrossRef]
- Hammou, H.A.; Latour, D.; Samoudi, S.; Mouhri, K.; Douma, M.; Robin, J.; Loudiki, M. Occurrence of the First Toxic Microcystis Bloom in a Recent Moroccan Reservoir. Water Resour. 2018, 45, 409–417. [Google Scholar] [CrossRef]
- El Herry-Allani, S.; Bouaïcha, N. Cyanobacterial Blooms in Dams: Environmental Factors, Toxins, Public Health, and Remedial Measures. In Dams: Structure, Performance and Safety Management; Nova Science Publishers, Inc.: Hauppauge, NY, USA, 2013; pp. 227–264. [Google Scholar]
- Willén, E.; Ahlgren, G.; Tilahun, G.; Spoof, L.; Neffling, M.-R.; Meriluoto, J. Cyanotoxin Production in Seven Ethiopian Rift Valley Lakes. Inl. Waters 2011, 1, 81–91. [Google Scholar] [CrossRef]
- Major, Y.; Kifle, D.; Spoof, L.; Meriluoto, J. Cyanobacteria and Microcystins in Koka Reservoir (Ethiopia). Environ. Sci. Pollut. Res. 2018, 25, 26861–26873. [Google Scholar] [CrossRef]
- Admas, A.; Agida, A.; Melese, S.; Genetu, A. Isolation and Characterization of Toxic Cyanobacterial Communities Distribution in Lake Tana, Amhara Regional State, Ethiopia. bioRxiv 2019. preprint. [Google Scholar] [CrossRef]
- Tilahun, S.; Kifle, D.; Zewde, T.W.; Johansen, J.A.; Demissie, T.B.; Hansen, J.H. Temporal Dynamics of Intra-and Extra-Cellular Microcystins Concentrations in Koka Reservoir (Ethiopia): Implications for Public Health Risk. Toxicon 2019, 168, 83–92. [Google Scholar] [CrossRef]
- Krienitz, L.; Ballot, A.; Wiegand, C.; Kotut, K.; Codd, G.A.; Pflugmacher, S. Cyanotoxin-Producing Bloom of Anabaena Flos-Aquae, Anabaena Discoidea and Microcystis aeruginosa (Cyanobacteria) in Nyanza Gulf of Lake Victoria, Kenya. Angew. Bot. 2002. [Google Scholar]
- Krienitz, L.; Ballot, A.; Casper, P.; Kotut, K.; Wiegand, C.; Pflugmacher, S. Cyanobacteria in Hot Springs of East Africa and Their Potential Toxicity. Algol. Stud. Hydrobiol. Suppl. Vol. 2005, 297–306. [Google Scholar] [CrossRef]
- Ballot, A.; Krienitz, L.; Kotut, K.; Wiegand, C.; Pflugmacher, S. Cyanobacteria and Cyanobacterial Toxins in the Alkaline Crater Lakes Sonachi and Simbi, Kenya. Harmful Algae 2005, 4, 139–150. [Google Scholar] [CrossRef]
- Kotut, K.; Ballot, A.; Wiegand, C.; Krienitz, L. Toxic Cyanobacteria at Nakuru Sewage Oxidation Ponds–A Potential Threat to Wildlife. Limnologica 2010, 40, 47–53. [Google Scholar] [CrossRef]
- Mwaura, F.; Koyo, A.O.; Zech, B. Cyanobacterial Blooms and the Presence of Cyanotoxins in Small High Altitude Tropical Headwater Reservoirs in Kenya. J. Water Health 2004, 2, 49–57. [Google Scholar] [CrossRef]
- Kaggwa, M.N.; Straubinger-Gansberger, N.; Schagerl, M. Cyanotoxins in Small Artificial Dams in Kenya Utilised for Cage Fish Farming–a Threat to Local People? Afr. J. Aquat. Sci. 2018, 43, 123–129. [Google Scholar] [CrossRef]
- Sekadende, B.C.; Lyimo, T.J.; Kurmayer, R. Microcystin Production by Cyanobacteria in the Mwanza Gulf (Lake Victoria, Tanzania). Hydrobiologia 2005, 543, 299–304. [Google Scholar] [CrossRef]
- Kimambo, O.N.; Gumbo, J.R.; Chikoore, H. The Occurrence of Cyanobacteria Blooms in Freshwater Ecosystems and Their Link with Hydro-Meteorological and Environmental Variations in Tanzania. Heliyon 2019, 5, e01312. [Google Scholar] [CrossRef]
- Green, J. Geographical Variation in Rotifers Associated with Microcystis Blooms. Hydrobiologia 2011, 662, 197–204. [Google Scholar] [CrossRef]
- Okello, W.; Ostermaier, V.; Portmann, C.; Gademann, K.; Kurmayer, R. Spatial Isolation Favours the Divergence in Microcystin Net Production by Microcystis in Ugandan Freshwater Lakes. Water Res. 2010, 44, 2803–2814. [Google Scholar] [CrossRef]
- Okello, W.; Portmann, C.; Erhard, M.; Gademann, K.; Kurmayer, R. Occurrence of Microcystin-producing Cyanobacteria in Ugandan Freshwater Habitats. Environ. Toxicol. 2010, 25, 367–380. [Google Scholar] [CrossRef]
- Poste, A.E.; Hecky, R.E.; Guildford, S.J. Phosphorus Enrichment and Carbon Depletion Contribute to High Microcystis Biomass and Microcystin Concentrations in Ugandan Lakes. Limnol. Oceanogr. 2013, 58, 1075–1088. [Google Scholar] [CrossRef]
- Addico, G.; Hardege, J.; Komarek, J.; Babica, P.; de Graft-Johnson, K.A.A. Cyanobacteria Species Identified in the Weija and Kpong Reservoirs, Ghana, and Their Implications for Drinking Water Quality with Respect to Microcystin. Afr. J. Mar. Sci. 2006, 28, 451–456. [Google Scholar] [CrossRef]
- Addico, G.N.D.; Lawton, L.; Edwards, C. Hepatotoxic-Microcystins in Two Drinking Water Reservoirs in the Central Region of Ghana. Hepatotoxic-Microcystins in Two Drinking. Toxicol. Forensic Med. Open J. 2017, 2, 1–11. [Google Scholar] [CrossRef]
- Akin-Oriola, G.A.; Anetekhai, M.A.; Oriola, A. Algal Blooms in Nigerian Waters: An Overview. Afr. J. Mar. Sci. 2006, 28, 219–224. [Google Scholar] [CrossRef]
- Odokuma, L.O.; Isirima, J.C. Distribution of Cyanotoxins in Aquatic Environments in the Niger Delta. Afr. J. Biotechnol. 2007, 6. [Google Scholar] [CrossRef]
- Chia, A.M.; Oniye, S.J.; Ladan, Z.; Lado, Z.; Pila, A.E.; Inekwe, V.U.; Mmerole, J.U. A Survey for the Presence of Microcystins in Aquaculture Ponds in Zaria, Northern-Nigeria: Possible Public Health Implication. Afr. J. Biotechnol. 2009, 8, 6282–6289. [Google Scholar] [CrossRef]
- Chia, M.A.; Kwaghe, M.J. Microcystins Contamination of Surface Water Supply Sources in Zaria-Nigeria. Environ. Monit. Assess. 2015, 187, 606. [Google Scholar] [CrossRef]
- Indabawa, I. Detection of Variants of Microcystin Produced by Microcystis Aeruginosa in Some Burrow Pits of Kano, Nigeria. Bayero J. Pure Appl. Sci. 2010, 2, 189–197. [Google Scholar] [CrossRef]
- Okechukwu I, O. Cyanobacteria Abundance and Its Relationship to Water Quality in the Mid-Cross River Floodplain, Nigeria. Rev. Biol. Trop. 2009, 57, 33–43. [Google Scholar] [CrossRef]
- Onyema, I.C. Phytoplankton Diversity and Succession in the Iyagbe Lagoon, Lagos. Eur. J. Sci. Res. 2010, 43, 61–74. [Google Scholar]
- Mussagy, A.; Annadotter, H.; Cronberg, G. An Experimental Study of Toxin Production in Arthrospira Fusiformis (Cyanophyceae) Isolated from African Waters. Toxicon 2006, 48, 1027–1034. [Google Scholar] [CrossRef]
- Pedro, O.; Correia, D.; Lie, E.; Skåre, J.U.; Leão, J.; Sandvik, M.; Berdal, K.G. Polymerase Chain Reaction (PCR) Detection of the Predominant Microcystin-Producing Genotype of Cyanobacteria in Mozambican Lakes. Afr. J. Biotechnol. 2011, 10, 19299–19308. [Google Scholar] [CrossRef]
- Pedro, O.; Rundberget, T.; Lie, E.; Correia, D.; Skaare, J.U.; Berdal, K.G.; Neves, L.; Sandvik, M. Occurrence of Microcystins in Freshwater Bodies in Southern Mozambique. J. Res. Environ. Sci. Toxicol. 2012, 1, 58–65. [Google Scholar]
- Pedro, O.; Lie, E.; Correia, D.; Neves, L.; Skaare, J.U.; Sandvik, M.; Berdal, K.G. Quantification of Microcystin-Producing Microcystis in Freshwater Bodies in the Southern Mozambique Using Quantitative Real Time Polymerase Chain Reaction. Afr. J. Biotechnol. 2013, 12, 4850–4857. [Google Scholar] [CrossRef]
- Magonono, M.; Oberholster, P.J.; Shonhai, A.; Makumire, S.; Gumbo, J.R. The Presence of Toxic and Non-Toxic Cyanobacteria in the Sediments of the Limpopo River Basin: Implications for Human Health. Toxins 2018, 10, 269. [Google Scholar] [CrossRef]
- Eguzozie, K.U.; Mavumengwana, V.; Nkosi, D.; Kayitesi, E.; Nnabuo-Eguzozie, E.C. Screening of Cyanobacterial Peptide Toxin, Microcystins in Hyperscum Water Samples from an Inland Sub Saharan Drinking Freshwater Reservoir. Bull. Environ. Contam. Toxicol. 2016, 97, 728–736. [Google Scholar] [CrossRef]
- Conradie, K.R.; Barnard, S. The Dynamics of Toxic Microcystis Strains and Microcystin Production in Two Hypertrofic South African Reservoirs. Harmful Algae 2012, 20, 1–10. [Google Scholar] [CrossRef]
- Van Ginkel, C.E.; Silberbauer, M.J.; Du Plessis, S.; Carelsen, C.I.C. Monitoring Microcystin Toxin and Chlorophyll in Five South African Impoundments. Int. Ver. Theor. Angew. Limnol. Verh. 2006, 29, 1611–1616. [Google Scholar] [CrossRef]
- Nchabeleng, T.; Cheng, P.; Oberholster, P.J.; Botha, A.M.; Smit, W.J.; Luus-Powell, W.J. Microcystin-LR Equivalent Concentrations in Fish Tissue during a Postbloom Microcystis Exposure in Loskop Dam, South Africa. Afr. J. Aquat. Sci. 2014, 39, 459–466. [Google Scholar] [CrossRef]
- Wicks, R.J.; Thiel, P.G. Environmental Factors Affecting the Production of Peptide Toxins in Floating Scums of the Cyanobacterium Microcystis Aeruginosa in a Hypertrophic African Reservoir. Environ. Sci. Technol. 1990, 24, 1413–1418. [Google Scholar] [CrossRef]
- Oberholster, P.J.; Botha, A.-M. Use of PCR Based Technologies for Risk Assessment of a Winter Cyanobacterial Bloom in Lake Midmar, South Africa. Afr. J. Biotechnol. 2007, 6. [Google Scholar] [CrossRef]
- Mokoena, M.M.; Mukhola, M.S. Current Effects of Cyanobacteria Toxin in Water Sources and Containers in the Hartbeespoort Dam Area, South Africa. Int. J. Environ. Res. Public Health 2019, 16, 4468. [Google Scholar] [CrossRef] [PubMed]
- Van Ginkel, C.E.; Conradie, B. Potentially Toxic Algal Incident in the Orange River, Northern Cape, 2000; Internal Report No. N/D801/12/DEQ/0800; Institute for Water Quality Studies, Department of Water Affairs and Forestry: Pretoria, South Africa, 2001. [Google Scholar]
- Oberholster, P.J.; Myburgh, J.G.; Govender, D.; Bengis, R.; Botha, A.-M. Identification of Toxigenic Microcystis Strains after Incidents of Wild Animal Mortalities in the Kruger National Park, South Africa. Ecotoxicol. Environ. Saf. 2009, 72, 1177–1182. [Google Scholar] [CrossRef] [PubMed]
- Mhlanga, L.; Day, J.; Cronberg, G.; Chimbari, M.; Siziba, N.; Annadotter, H. Cyanobacteria and Cyanotoxins in the Source Water from Lake Chivero, Harare, Zimbabwe, and the Presence of Cyanotoxins in Drinking Water. Afr. J. Aquat. Sci. 2006, 31, 165–173. [Google Scholar] [CrossRef]
- Magadza, C.H.D. 2009. Re-Eutrophication and Pathenogenic Contamination of Lake Chivero. Integrated Lake Basin Management; International Lake Environment Committee: Shiga, Japan, 2008. [Google Scholar]
- Kunz, M.J. Effect of Large Dams in the Zambezi River Basin: Changes in Sediment, Carbon and Nutrient Fluxes. Ph.D. Thesis, Swiss Federal Institute of Technology, Zürich, Switzerland, 2011. [Google Scholar]
- Tendaupenyu, P. Nutrient Limitation of Phytoplankton in Five Impoundments on the Manyame River, Zimbabwe. Water SA 2012, 38, 97–104. [Google Scholar] [CrossRef]
- Mankiewicz-Boczek, J.; Gągała, I.; Jurczak, T.; Urbaniak, M.; Negussie, Y.Z.; Zalewski, M. Incidence of Microcystin-producing Cyanobacteria in Lake Tana, the Largest Waterbody in Ethiopia. Afr. J. Ecol. 2015, 53, 54–63. [Google Scholar] [CrossRef]
- Oudra, B.; Dadi-El Andaloussi, M.; Vasconcelos, V.M. Identification and Quantification of Microcystins from a Nostoc Muscorum Bloom Occurring in Oukaïmeden River (High-Atlas Mountains of Marrakech, Morocco). Environ. Monit. Assess. 2009, 149, 437–444. [Google Scholar] [CrossRef]
- Mohamed, Z.A. Harmful Cyanobacteria and Their Cyanotoxins in Egyptian Fresh Waters–State of Knowledge and Research Needs. Afr. J. Aquat. Sci. 2016, 41, 361–368. [Google Scholar] [CrossRef]
- Mohamed, Z.A. Breakthrough of Oscillatoria Limnetica and Microcystin Toxins into Drinking Water Treatment Plants-Examples from the Nile River, Egypt. Water SA 2016, 42, 161–165. [Google Scholar] [CrossRef]
- Mokoena, M.M.; Mukhola, M.S.; Okonkwo, O.J. Hazard Assessment of Microcystins from the Household’s Drinking Water. Appl. Ecol. Environ. Res. 2016, 14, 695–710. [Google Scholar] [CrossRef]
- Chia, A.M.; Abolude, D.S.; Ladan, Z.; Akanbi, O.; Kalaboms, A. The Presence of Microcystins in Aquatic Ecosystems in Northern Nigeria: Zaria as a Case Study. Res. J. Environ. Toxicol. 2009, 3, 170–178. [Google Scholar] [CrossRef]
- Addico, G.N.D.; Hardege, J.D.; Babica, P.; Kohoutek, J.; Johnson, K.A.A.D.G. Microcystin-RR Like Toxin Identified in the Cyano- Bacterium Anabaena Flos-Aquae Strain CCAP 1403/13B Culture. West Afr. J. Appl. Ecol. 2011, 19. [Google Scholar]
- Kouassi, B.A.T.; Adon, M.P.; Komoé, K.; Ouattara, A. Cyanobacteria from a Shallow Reservoir in Côte d’Ivoire UFR Biological Sciences, Peleforo Gon Coulibaly University, Korhogo, Côte d’Ivoire. J. Biodivers. Environ. Sci. 2015, 7, 136–149. [Google Scholar]
- Kaebernick, M.; Dittmann, E.; Börner, T.; Neilan, B.A. Multiple Alternate Transcripts Direct the Biosynthesis of Microcystin, a Cyanobacterial. Appl. Environ. Microbiol. 2002, 68, 449–455. [Google Scholar] [CrossRef]
- Christiansen, G.; Molitor, C.; Philmus, B.; Kurmayer, R. Nontoxic Strains of Cyanobacteria Are the Result of Major Gene Deletion Events Induced by a Transposable Element. Mol. Biol. Evol. 2008, 25, 1695–1704. [Google Scholar] [CrossRef]





| Gene | Protein | Functions |
|---|---|---|
| mcyH | McyH | ABC transporter |
| mcyI | McyI | Putative dehydrogenase |
| mcyF | McyF | Amino acid racemase |
| mcyE | McyE | NRPS-PKS (KS-AT-ACP-AMT-C-A-PCP-C) |
| mcyD | McyD | PKS (KS-DH-CM-KR-ACP-KS-AT-DH-KR-ACP) |
| mcyG | McyG | NRPS-PKS (A-PCP-KS-AT-CM-KR-ACP) |
| mcyA | McyA | NRPS (A-NMT-PCP-C-A-PCP-E) |
| mcyB | McyB | NRPS (C-A-PCP-C-A-PCP) |
| mcyC | McyC | NRPS (C-A-PCP-TE) |
| mcyJ | McyJ | O-acetyltransferase |
| Sub-region | Country | Type of Water Body | Major Toxin-Producing Species | Detected Cyanotoxins | Concentration of Detected Cyanotoxins | Detection Method | References |
|---|---|---|---|---|---|---|---|
| North Africa | Algeria | Freshwater lakes | Microcystis spp. | MC-RR, MC-LR, MC-FR, MC-WR, MC-YR, MC-LA, MC-(H4)YR, MC-HilR, [Asp3]MCRAba, [Glu(OCH3)6]MC-LR, and [Glu(OCH3)6]MC-FR | 4590 μg g−1 DW total microcystins | LCMS/MS, HPLC | [58,59,60,61] |
| Egypt | Nile river; irrigation canals; upper Egypt fishponds | Microcystis aeruginosa, Oscillatoria tenuis, Anabaena subcylindrica, Anabaena variables, Nostoc spongiaeforme, Oscillatoria limnetica, Phormidium corium, Planktothrix agardhii, Anabaena variables, Plectonema boryanum, Raphidiopsis raciborskii | Total MCs, MC-LR, MC-RR, MC-YR | 300 to 877 µg L−1 total microcystins | HPLC, NMR, ESIMS, ESIMS-CID-MS, ELISA | [62,63,64,65,66,67] | |
| Morocco | Reservoirs, ponds, waste stabilization ponds, and rivers | Nostoc muscorum | Total MCs, MC-LR, MC-RR, and MC-WR | 229.4 μg/g−1 DW | ELISA, HPLC, FAB-MS | [68,69,70,71] | |
| Tunisia | Dams, reservoirs | Microcystis spp., Oscillatoria tenuis, M. aeruginosa, R. raciborskii, Planktothrix agardhii | MC-LR equivalent MC-LR (MC-FR, MC-RR, MC-YR, and MCWR) | 0.008–5.57 μg L−1 | GC/MS, PP2 inhibition assay | [31,59,72] | |
| East Africa | Ethiopia | Reservoirs and lakes | Microcystis spp., Microcystis aeruginosa, Microcystis botrys, Microcystis flos-aquae Microcystis novacekii, Microcystis panniformis, Anabaena spp., Nostoc spp., Raphidiopsis spp., Raphidiopsis africana, Raphidiopsis. Raciborskii | MC-LR, MC-YR, MC-RR, MC-dmLR, and MC-LA | 0.58- 1547.28 μg L−1 total microcystins | ELISA, LC-ESI-MS-MRM, HPLC-DAD | [46,73,74,75,76] |
| Kenya | Freshwater lakes and saline lakes | Microcystis aeruginosa, Arthrospira fusiformis, Microcystis flos-aquae, Phormidium terebriformis, Oscillatoria willei, Spirulina subsalsa | MC-LR;MC-RR; MC-LF; MC-YR | 1.6–19,800 μg g−1 DW | HPLC-PDA, MALDI-TOF, ELISA | [29,34,77,78,79,80,81,82] | |
| Tanzania | Freshwater lakes | Microcystis sp. | MC-RR | 0–1.0 μg L−1 | HPLC-DAD | [83,84] | |
| Uganda | Freshwater lakes | Microcystis aeruginosa, M. flos-aquae, Anabaenopsis spp., Aphanizomenon sp., Anabaena sp., Raphidiopsis raciborskii | MC-RR, (Asp3) MC-RR, MC-YR, (Asp3) MC-YR, MC-LR, MC-RY, (Asp3) MC-RY | 0.02–10.00 μg MC-LR eq./L 0.03 to 144 fg cell−1 total microcystins | HPLC-DAD, LC-MS/MS, MALDI-TOF MS | [34,35,36,85,86,87,88] | |
| West Africa | Ghana | Freshwater reservoirs | Microcystis aeruginosa | MC-RR | 0.03–3.21 μg L−1 total microcystins, 0.1–0.79 μg L−1, 10.60 μg g−1 DW MC-RR | HPLC-UV | [89,90] |
| Nigeria | Lakes, reservoirs, lagoons, rivers, ponds, floodplain | Raphidiopsis sp., Microcystis sp., Microcystis aeruginosa, Microcystis flos-aquae, Microcystis wesenbergii, Lyngbya sp. | MCs, MC-LR, MC-RR | 0.19–7.75, 3.8, 1.68–3.94, 0.6–5.89 μg L−1 total microcystins | ELISA, HPLC-DAD | [12,91,92,93,94,95,96,97] | |
| South Africa | Mozambique | Wastewater treatment ponds, dams, lakes, irrigation channels | Arthrospira fusiformis, Microcystis spp. | MC-LR, MC-YR, MC-RR | 0.10–- 7.89 μg L−1 2.1–159.4 ng g−1 DW | LCMS | [98,99,100,101] |
| Botswana | Ponds, rivers, and lakes | Microcystis novacekii, Raphidiopsis raciborskii, Phormidium spp., Planktothrix spp. | MC-RR, MC-YR, MC-LR, and MC-WR | 53.62 μg g−1 DW MC-RR; 12.114 μg g−1 DW MC-LR | LC-MS/MS | [102,103] | |
| South Africa | Reservoirs, lakes, and rivers | Microcystis spp., Anabaena spp., Microcystis aeruginosa, Microcystis flos-aquae, Raphidiopsis raciborskii, Phormidium spp., Planktothrix spp. | MC-LR, MC-YR, MC-RR, MC-(H4)YR and (D-Asp3, Dha7) MC-RR; MC-LA, MC-LF | 14.10 to 270.7 μg g−1 MC-LR; 0.15 to 72.28 μg g−1 MC-YR | HPLC-DAD, HPLC-UV, ELISA, LC-MS | [20,24,26,39,53,57,102,103,104,105,106,107,108,109,110,111] | |
| Zimbabwe | Rivers and lakes | Microcystis spp., Raphidiopsis raciborskii, Phormidium spp., Planktothrix spp. | MC-LR | 0.2–22.48 μg L−1 | ELISA, HPLC | [102,112,113,114,115] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chia, M.A.; Ameh, I.; George, K.C.; Balogun, E.O.; Akinyemi, S.A.; Lorenzi, A.S. Genetic Diversity of Microcystin Producers (Cyanobacteria) and Microcystin Congeners in Aquatic Resources across Africa: A Review Paper. Toxics 2022, 10, 772. https://doi.org/10.3390/toxics10120772
Chia MA, Ameh I, George KC, Balogun EO, Akinyemi SA, Lorenzi AS. Genetic Diversity of Microcystin Producers (Cyanobacteria) and Microcystin Congeners in Aquatic Resources across Africa: A Review Paper. Toxics. 2022; 10(12):772. https://doi.org/10.3390/toxics10120772
Chicago/Turabian StyleChia, Mathias Ahii, Ilu Ameh, Korie Chibuike George, Emmanuel Oluwadare Balogun, Suwebat Ayanronke Akinyemi, and Adriana Sturion Lorenzi. 2022. "Genetic Diversity of Microcystin Producers (Cyanobacteria) and Microcystin Congeners in Aquatic Resources across Africa: A Review Paper" Toxics 10, no. 12: 772. https://doi.org/10.3390/toxics10120772
APA StyleChia, M. A., Ameh, I., George, K. C., Balogun, E. O., Akinyemi, S. A., & Lorenzi, A. S. (2022). Genetic Diversity of Microcystin Producers (Cyanobacteria) and Microcystin Congeners in Aquatic Resources across Africa: A Review Paper. Toxics, 10(12), 772. https://doi.org/10.3390/toxics10120772