Effects of Farm Management Practices and Transport Time on Post-Mortem Changes of Longissimus lumborum Muscle Proteins in Suckling Goat Kids

 , ,
, ,
Abstract
:
1. Introduction
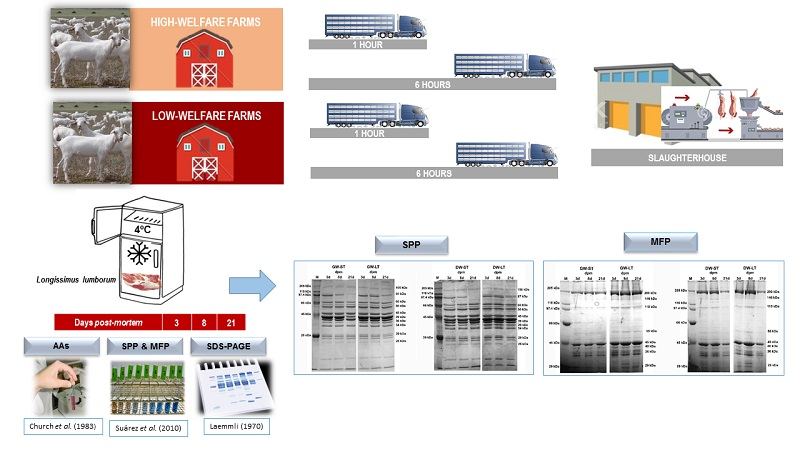
2. Materials and Methods
2.1. Animal Handling and Slaughtering
2.2. Experimental Design
2.3. Extraction of Sarcoplasmic (SPP) and Myofibrillar (MFP) Fractions
2.4. Determination of Soluble Protein
2.5. Total Amino Acid Determination
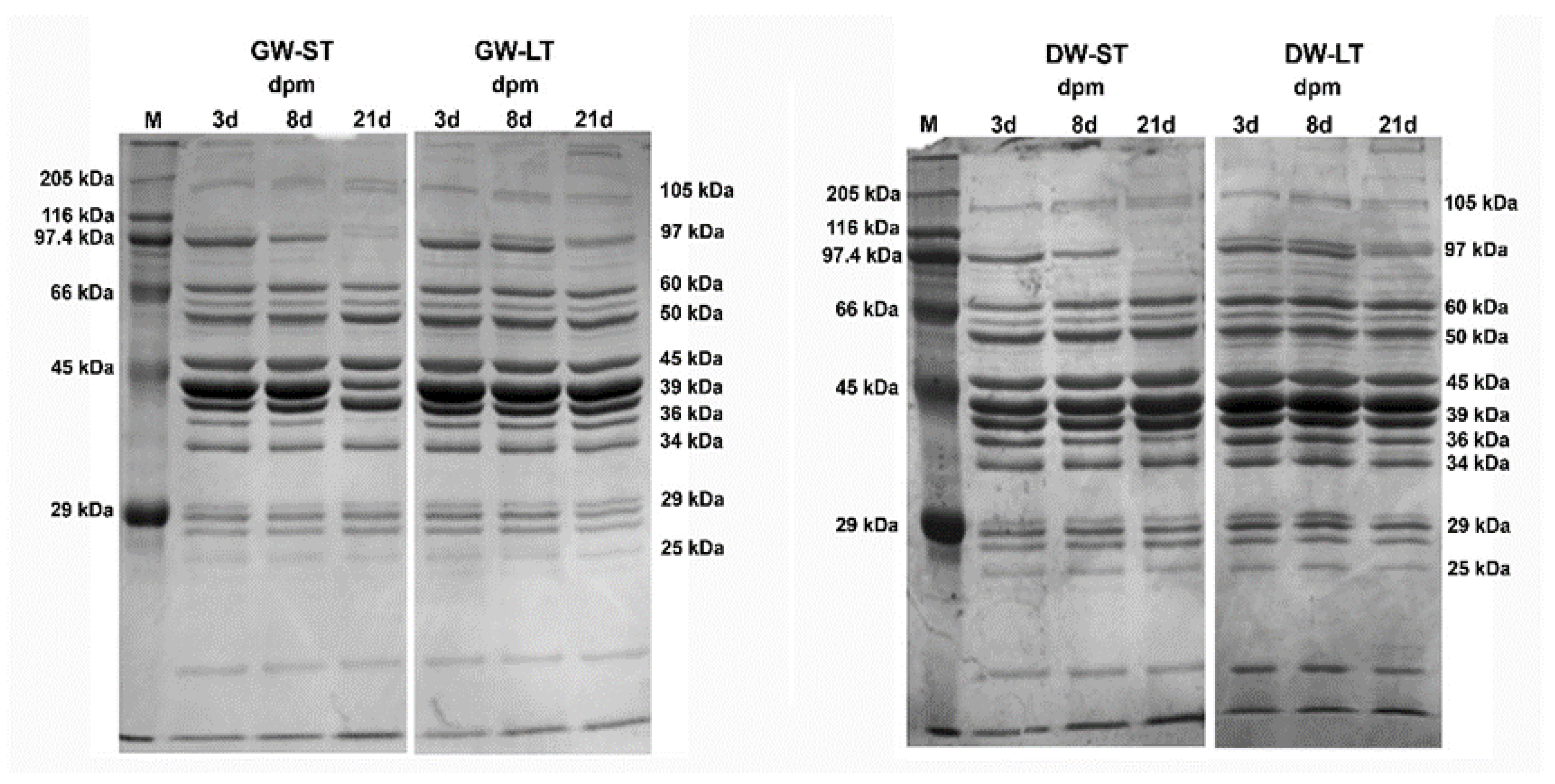
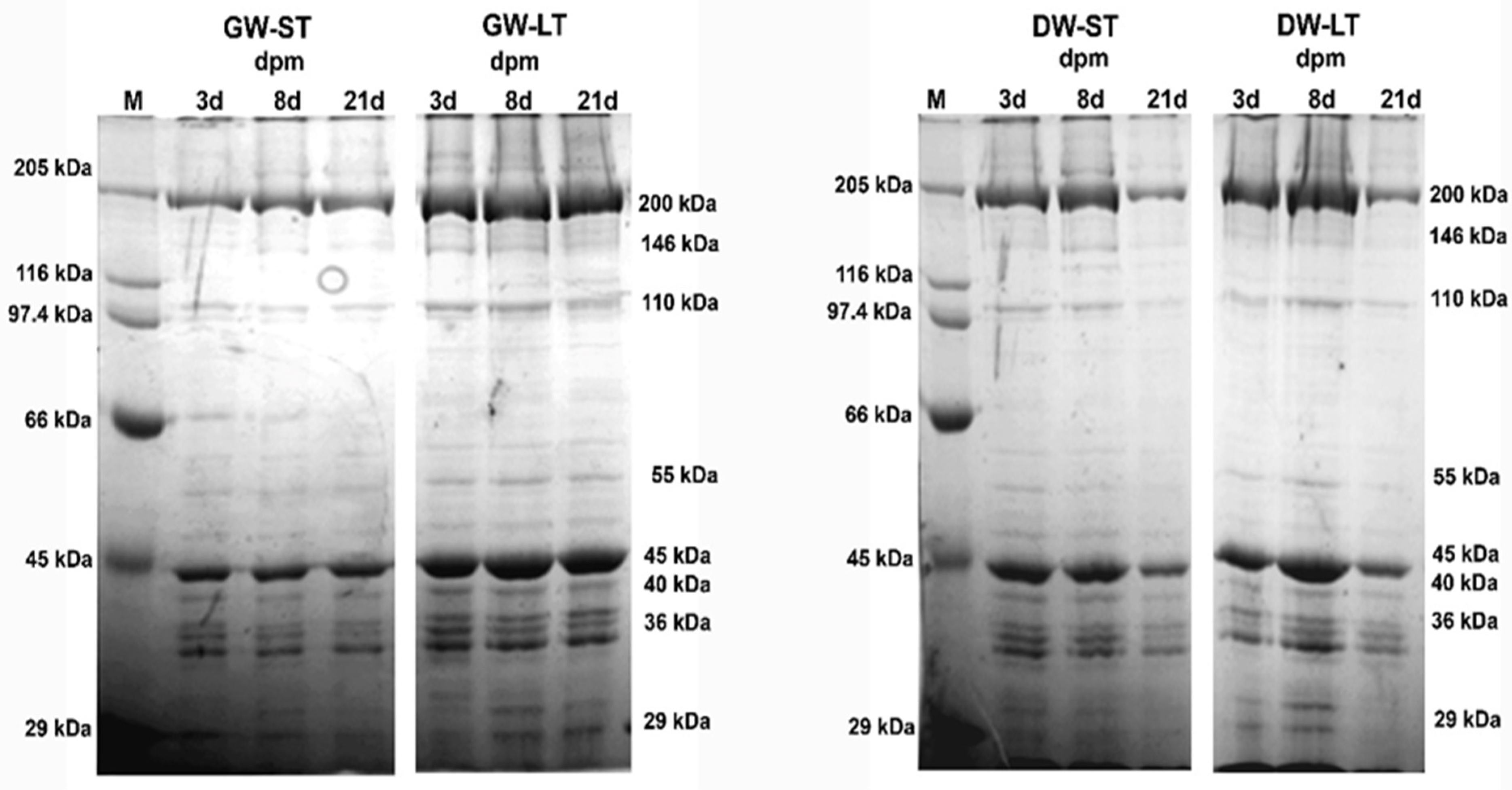
2.6. Electrophoresis of Muscle Proteins
2.7. Densitometric Analysis
2.8. Statistical Analysis
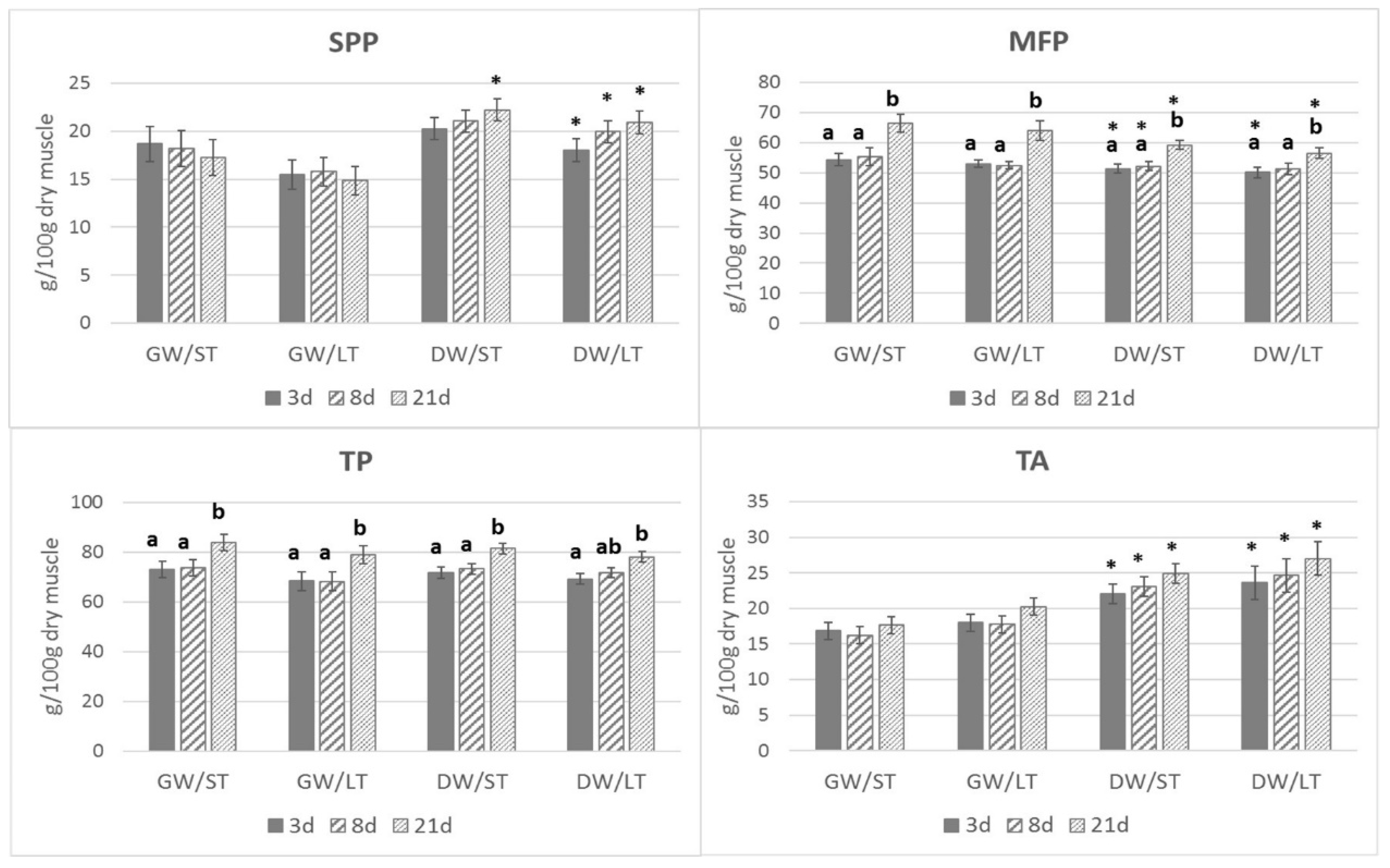
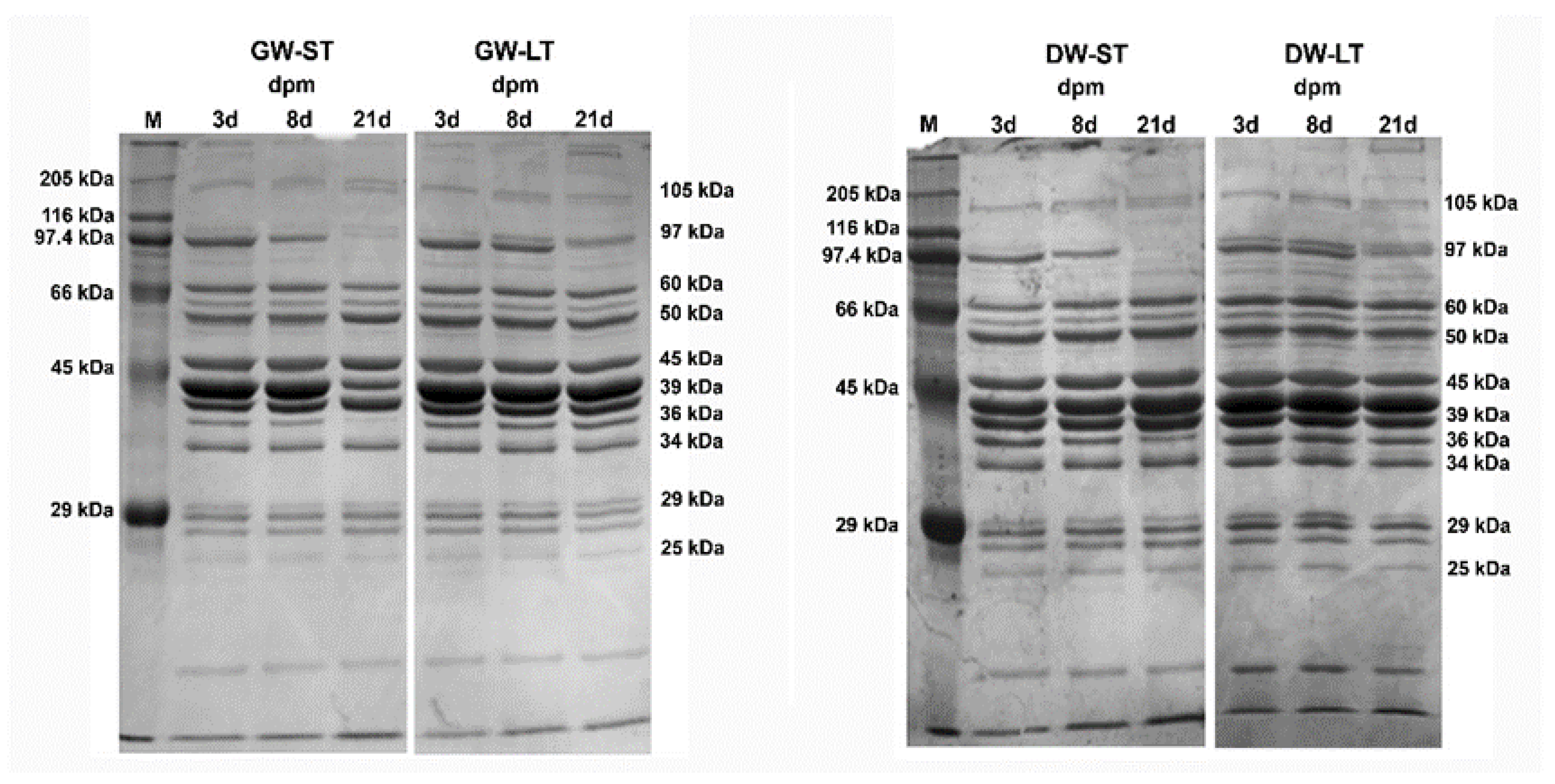
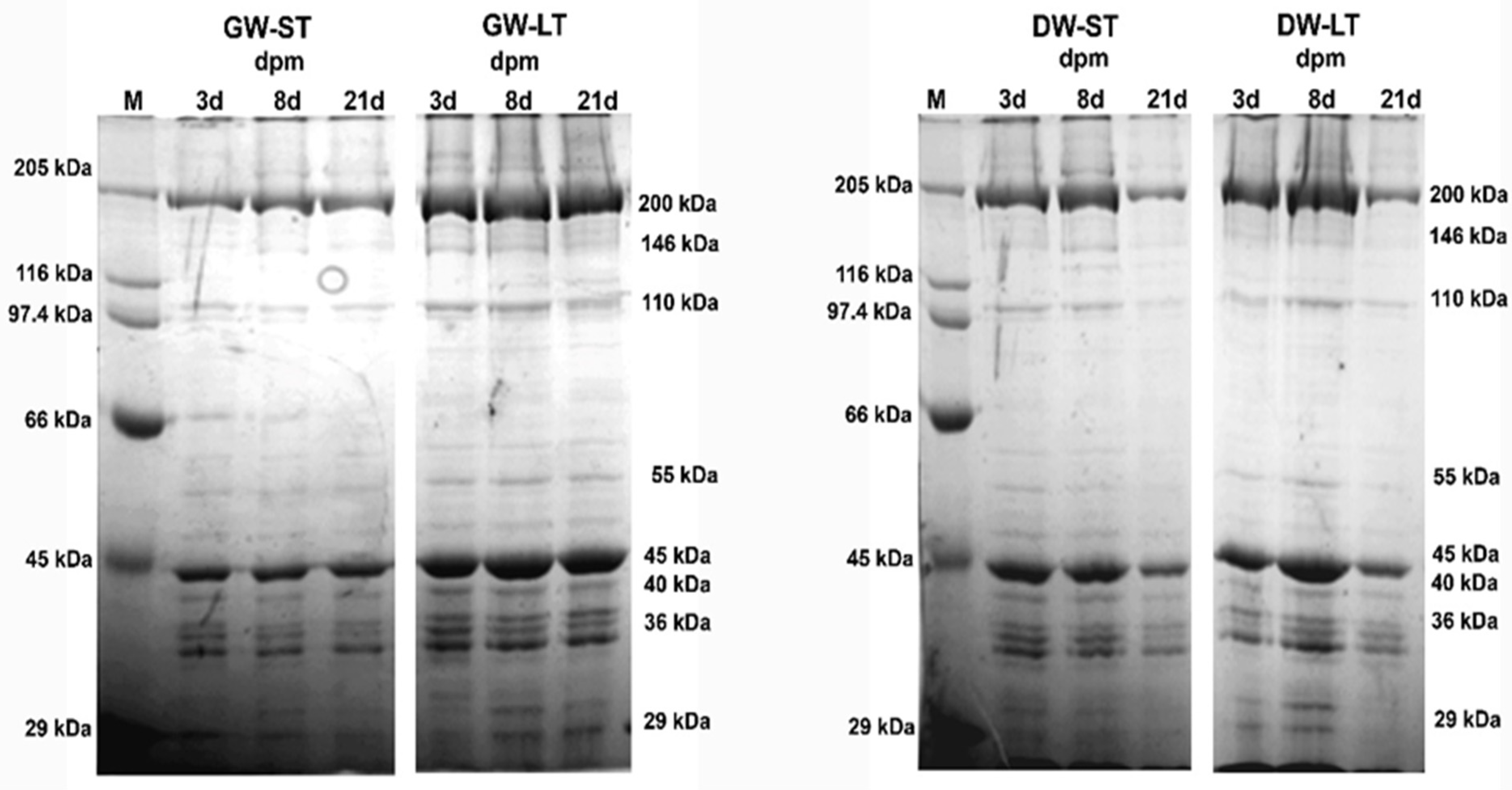
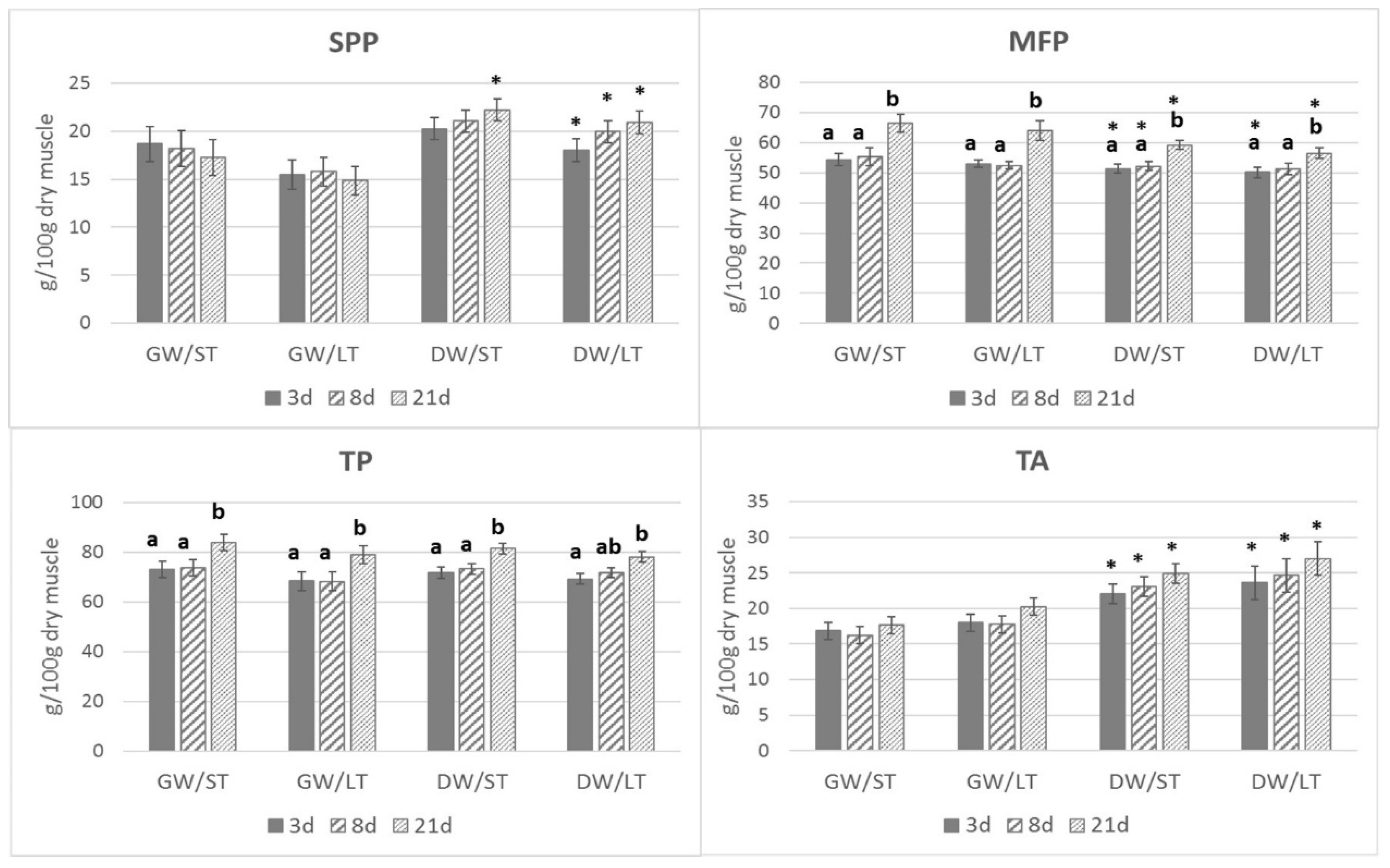
3. Results
4. Discussion
4.1. Proteins and Aminoacids Content
4.2. Electrophoresis of Sarcoplasmic Proteins
4.3. Electrophoresis of Myofibrillar Proteins
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Etim, N.N.; Williams, M.E.; Evans, E.I.; Offiong, E.E.A. Physiological and behavioural responses of farm animals to stress, implications to animal productivity. Am. J. Adv. Agric. Res. 2013, 1, 53–61. [Google Scholar]
- De la Fuente, J.; González de Chávarri, E.; Sánchez, M.; Vieira, C.; Lauzurica, S.; Díaz, M.T.; Pérez, C. The effects of journey duration and space allowance on the behavioural and biochemical measurements of stress responses in suckling lambs during transport to an abattoir. Appl. Anim. Behav. Sci. 2012, 142, 30–41. [Google Scholar] [CrossRef]
- Zając, M.; Palka, K.; Mikołajczak, B.; Pospiech, E. The effect of muscle type and time of storage on myofibrillar protein proportion in beef. Ann. Anim. Sci. 2016, 16, 585–600. [Google Scholar] [CrossRef] [Green Version]
- Santos, C.C.; Delgado, E.F.; Menten, J.F.M.; Pedreira, A.C.M.; Castilho, C.J.C.; Mourão, G.B.; Brossi, C.; Silva, I.J.O. Sarcoplasmatic and myofibrillar protein changes caused by acute heat stress in broiler chicken. Sci. Agric. 2008, 65, 453–458. [Google Scholar] [CrossRef] [Green Version]
- Lametsch, R.; Roepstorff, P.; Bendixen, E. Identification of protein degradation during post-mortem storage of pig meat. J. Agric. Food Chem. 2002, 50, 5508–5512. [Google Scholar] [CrossRef]
- Morzel, M.; Chambon, C.; Hamelin, M.; SantéLhoutellier, V.; Sayd, T.; Monin, G. Proteome changes during pork meat ageing following use of two different pre-slaughter handling procedures. Meat Sci. 2004, 67, 689–696. [Google Scholar] [CrossRef]
- Laville, E.; Sayd, T.; Terlouw, C.; Chambon, C.; Damon, M.; Larzul, C.; Leroy, P.; Glénisson, J.; Chérel, P. Comparison of sarcoplasmic proteomes between two groups of pig muscles selected for shear force of cooked meat. J. Agric. Food Chem. 2007, 55, 5834–5841. [Google Scholar] [CrossRef]
- Jia, X.; Veiseth-Kent, E.; Grove, H.; Kuziora, P.; Aass, L.; Hildrum, K.I.; Hollung, K. Peroxiredoxin-6- A potential protein marker for meat tenderness in bovine longissimus thoracis muscle. J. Anim. Sci. 2009, 87, 2391–2399. [Google Scholar] [CrossRef] [Green Version]
- Sierra, V.; Fernández-Suárez, V.; Castro, P.; Osoro, K.; Vega-Naredo, I.; García-Macia, M. Identification of biomarkers of meat tenderization and its use for early classification of Asturian beef into fast and late tenderizing meat. J. Sci. Food Agric. 2012, 92, 2727–2740. [Google Scholar] [CrossRef]
- Lametsch, R.; Karlsson, A.; Rosenvold, K.; Andersen, H.J.; Roepstorff, P.; Bendixen, E. Postmortem proteome changes of porcine muscle related to tenderness. J. Agric. Food Chem. 2003, 51, 6992–6997. [Google Scholar] [CrossRef]
- Picard, B.; Gagaoua, M. Proteomic investigations of beef tenderness. In Proteomics in Food Science: From Farm to Fork; Colgrave, M.L., Ed.; Academic Press: London, UK, 2017; pp. 177–197. [Google Scholar]
- Gagaoua, M.; Monteils, V.; Picard, B. Data from the farmgate-to-meat continuum including omics-based biomarkers to better understand the variability of beef tenderness, an integromics approach. J. Agric. Food Chem. 2018, 66, 13552–13563. [Google Scholar] [CrossRef] [PubMed]
- Xing, T.; Gao, F.; Tume, R.K.; Zhou, G.; Xu, X. Stress effects on meat quality, a mechanistic perspective. Comp. Rev. Food Sci. Food Saf. 2019, 18, 380–401. [Google Scholar] [CrossRef] [Green Version]
- Franco, D.; Mato, A.; Salgado, F.J.; López-Pedrouso, M.; Carrera, M.; Bravo, S.; Zapata, C. Tackling proteome changes in the longissimus thoracis bovine muscle in response to pre-slaughter stress. J. Proteom. 2015, 122, 73–85. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oliván, M.; Fernández-Suárez, V.; Díaz-Martínez, F.; Sierra, V.; Coto-Montes, A.; de Luxán-Delgado, B.; Peña, R.; Bassols, A.; Fàbrega, E.; Dalmau, A.; et al. Identification of biomarkers of stress in meat of pigs managed under different mixing treatments. Br. Biotech. J. 2016, 11, 1–13. [Google Scholar] [CrossRef]
- Guerrero, A.; Campo, M.M.; Olleta, J.L.; Sañudo, C. Carcass and Meat Quality in Goat, Goat Science, Sándor Kukovics; IntechOpen: London, UK, 2017; Available online: https://www.intechopen.com/books/goat-science/carcass-and-meat-quality-in-goat (accessed on 28 April 2020). [CrossRef] [Green Version]
- Casey, N.H.; Webb, E.C. Managing goat production for meat quality. Small Rum. Res. 2010, 89, 218–224. [Google Scholar] [CrossRef] [Green Version]
- Kannan, G.; Kouakou, B.; Terrill, T.H.; Gelaye, S. Endocrine, blood metabolite, and meat quality changes in goats as influenced by short-term, pre-slaughter stress. J. Anim. Sci. 2003, 81, 1499–1507. [Google Scholar] [CrossRef]
- Greenwood, P.L.; Finn, J.A.; May, T.J.; Nicholls, P.J. Pre-slaughter management practices influence carcass characteristics of young goats. Aust. J. Exp. Agric. 2008, 48, 910–915. [Google Scholar] [CrossRef]
- Kadim, I.T.; Mahgoub, O.; Al-Kindi, A.; Al-Marzooqi, W.; Al-Saqri, N.M. Effects of transportation at high ambient temperatures on physiological responses, carcass and meat quality characteristics of three breeds of Omani goats. Meat Sci. 2006, 73, 626–634. [Google Scholar] [CrossRef]
- Kadim, I.T.; Mahgoub, O.; Al-Marzoogi, W.; Khalaf, S.; Al-Sinawi, S.H.; Al-Amri, I. Effects of transportation during the hot season breed and electrical stimulation on histochemical and meat quality characteristics of goat longissimus muscle. Anim. Sci. J. 2010, 81, 352–361. [Google Scholar] [CrossRef]
- De la Fuente, J.; Sánchez, M.; Pérez, C.; Lauzurica, S.; Vieira, C.; González De Chávarri, E.; Díaz, M.T. Physiological response and carcass and meat quality of suckling lambs in relation to transport time and stocking density during transport by road. Animal 2010, 4, 250–258. [Google Scholar] [CrossRef] [Green Version]
- Alcalde, M.J.; Suárez, M.D.; Rodero, E.; Álvarez, R.; Sáez, M.I.; Martínez, T.F. Effects of farm management practices and transport duration on stress response and meat quality traits of suckling goat kids. Animal 2017, 11, 1626–1635. [Google Scholar] [CrossRef] [PubMed]
- Welfare Quality. Welfare Quality Assessment Protocol for Cattle; Welfare Quality® Consortium: Lelystad, The Netherlands, 2009. [Google Scholar]
- Battini, M.; Vieira, A.; Barbieri, S.; Ajuda, I.; Stilwell, G.; Mattiello, S. Animal-based indicators for on-farm welfare assessment for dairy goats, a review. J. Dairy Sci. 2014, 97, 6625–6648. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Gornall, A.G.; Bardawill, C.J. Determination of serum proteins by means of the biuret reaction. J. Biol. Chem. 1949, 177, 751–766. [Google Scholar] [PubMed]
- Church, F.C.; Swaisgood, H.E.; Porter, D.H.; Catignan, G.L. Spectrophotometric assay using o-phthaldialdehyde for determination of proteolysis in milk and isolated milk proteins. J. Dairy Sci. 1983, 66, 1219–1227. [Google Scholar] [CrossRef]
- Laemmli, U.K. Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature 1970, 227, 680–685. [Google Scholar] [CrossRef]
- Huff–Lonergan, E.; Lonergan, S.M. Mechanism of water–holding capacity of meat, The role of postmortem biochemical and structural changes. Meat Sci. 2005, 71, 194–204. [Google Scholar] [CrossRef]
- Picariello, G.; De Martino, A.; Mamone, G.; Ferranti, P.; Addeo, F.; Faccia, M.; SpagnaMusso, S.; Di Luccia, A. Proteomic study of muscle sarcoplasmic proteins using AUT-PAGE/SDS-PAGE as two-dimensional gel electrophoresis. J. Chrom. B Anal. Technol. Biom. Life Sci. 2006, 833, 101–108. [Google Scholar] [CrossRef]
- Ryu, Y.C.; Kim, B.C. The relationship between muscle fiber characteristics, postmortem metabolic rate, and meat quality of pig longissimus lumborum muscle. Meat Sci. 2005, 71, 351–357. [Google Scholar] [CrossRef]
- Gagaoua, M.; Terlouw, E.M.; Micol, D.; Boudjellal, A.; Hocquette, J.F.; Picard, B. Understanding early post-mortem biochemical processes underlying meat color and pH decline in the Longissimus thoracis muscle of young Blond d’Aquitaine bulls using protein biomarkers. J. Agric. Food Chem. 2015, 63, 6799–6809. [Google Scholar] [CrossRef]
- Nair, M.N.; Li, S.; Beach, C.M.; Rentfrow, G.; Suman, S.P. Changes in the sarcoplasmic proteome of beef muscles with differential color stability during postmortem aging. Meat Muscle Biol. 2018, 2, 1–17. [Google Scholar] [CrossRef] [Green Version]
- Oliván, M.; Gonzalez, J.; Bassols, A.; Diaz, F.; Carreras, R.; Mainau, E.; Arroyo, L.; Pena, R.; Potes, Y.; Coto-Montes, A.; et al. Effect of sex and ryr1 gene mutation on the muscle proteomic profile and main physiological biomarkers in pigs at slaughter. Meat Sci. 2018, 141, 81–90. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sabow, A.B.; Zulkifli, I.; Goh, Y.M.; Ab Kadir, M.Z.A.; Kaka, U.; Imlan, J.C.; Abubakar, A.A.; Adeyemi, K.D.; Sazili, A.Q. Bleeding efficiency, microbiological quality and oxidative stability of meat from goats subjected to slaughter without stunning in comparison with different methods of pre-slaughter electrical stunning. PLoS ONE 2016, 12, e0178890. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Adeyemi, K.D.; Shittu, R.M.; Sabow, A.B.; Ebrahimi, M.; Sazili, A.Q. Influence of diet and postmortem ageing on oxidative stability of lipids, myoglobin and myofibrillar proteins and quality attributes of gluteus medius muscle in goats. PLoS ONE 2016, 115, 1–15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gadiyaram, K.M.; Kannan, G.; Pringle, T.D.; Kouakou, B.; McMillin, K.V.; Park, Y.V. Effects of postmortem carcass electrical stimulation on goat meat quality characteristics. Small Rum. Res. 2008, 78, 106–114. [Google Scholar] [CrossRef]
- Schulte, M.D.; Johnson, L.G.; Zuber, E.A.; Patterson, B.M.; Outhouse, A.C.; Fedler, C.A.; Steadham, E.M.; King, D.A.; Prusa, K.J.; Huff-Lonergan, E.; et al. Influence of postmortem aging and post-aging freezing on pork loin quality attributes. Meat Muscle Biol. 2019, 3, 313–323. [Google Scholar] [CrossRef]
- Li, P.; Wang, T.; Mao, Y.; Zhang, Y.; Niu, L.; Liang, R.; Zhu, L.; Luo, X. Effect of ultimate pH on postmortem myofibrillar protein degradation and meat quality characteristics of chinese yellow crossbreed cattle. Sci. World J. 2014, 55, 5834–5841. [Google Scholar] [CrossRef] [Green Version]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| ST | LT | |||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Farm Origin | Post-Mortem Time (days) | Post-Mortem Time (days) | FO | TD | AT | FO*TD | FO*AT | TD*AT | FO*TD*AT | |||||
| 3 | 8 | 21 | 3 | 8 | 21 | |||||||||
| B-97 kDa | GW | 5.07 b ± 0.57 | 4.59 b ± 0.51 | 2.66 a ± 0.33 | 7.84 c# ± 0.45 | 5.35 b ± 0.46 | 1.89 a ± 0.51 | * | ns | *** | ns | * | ns | * |
| DW | 6.05 b ± 0.57 | 4.13 ab ± 0.51 | 3.03 a ± 0.33 | 5.21 b* ± 0.45 | 3.72 ab* ± 0.46 | 3.16 a ± 0.51 | ||||||||
| B-65 kDa | GW | 5.77 ± 0.53 | 5.52 ± 0.58 | 5.99 ± 0.47 | 7.02 ± 0.40 | 7.07 ± 0.38 | 6.38 ± 0.40 | ns | * | ns | ns | ns | ns | ns |
| DW | 5.44 ± 0.53 | 6.13 ± 0.58 | 7.14 ± 0.47 | 6.59 ± 0.40 | 6.36 ± 0.38 | 6.77 ± 0.40 | ||||||||
| B-58 kDa | GW | 8.34 ± 0.17 | 7.45 ± 0.68 | 8.48 ± 0.19 | 7.66 a ± 0.69 | 9.24 b ± 0.58 | 9.88 b# ± 0.48 | ns | ns | ns | * | ns | ns | ns |
| DW | 8.24 ± 0.19 | 9.04 ± 0.68 | 8.44 ± 0.22 | 8.47 ± 0.69 | 7.81 ± 0.58 | 8.51 ± 0.48 | ||||||||
| B-46 kDa | GW | 10.25 ± 0.49 | 9.07 ± 0.50 | 9.71 ± 0.46 | 9.75 ± 0.49 | 10.09 ± 0.73 | 11.85 # ± 0.62 | ** | ns | * | * | ns | ns | ns |
| DW | 11.00 ± 0.49 | 11.12 * ± 0.50 | 12.49 * ± 0.46 | 10.17 ± 0.49 | 10.54 ± 0.73 | 11.40 ± 0.62 | ||||||||
| B-43 kDa | GW | 23.45 ± 0.81 | 21.43 ± 1.08 | 20.80 ± 0.32 | 21.37 ± 1.54 | 22.17 ± 1.48 | 19.06 ± 0.58 | ns | ns | ns | ns | ns | ns | ns |
| DW | 25.84 ± 0.93 | 22.95 ± 1.08 | 21.59 ± 0.28 | 21.91 ± 1.54 | 21.09 ± 1.48 | 22.56 #* ± 0.58 | ||||||||
| B-40 kDa | GW | 11.88 ± 1.13 | 11.61 ± 0.91 | 13.93 ± 1.64 | 12.14 a ± 1.02 | 11.60 a ± 0.85 | 15.74 b ± 0.65 | ns | ns | ns | ns | ns | ns | ns |
| DW | 12.07 ± 1.13 | 10.81 ± 0.91 | 10.74 ± 1.64 | 11.60 ± 1.02 | 11.66 ± 0.85 | 12.17 * ± 0.65 | ||||||||
| B-38 kDa | GW | 6.59 c ± 0.66 | 4.47 b ± 0.27 | 1.74 a ± 0.24 | 6.51 b ± 0.46 | 5.45 b ± 0.54 | 1.52 a ± 0.40 | ** | ns | *** | ns | ** | ns | ns |
| DW | 7.37 c ± 0.66 | 5.88 b* ± 0.24 | 3.38 a* ± 0.24 | 6.14 c ± 0.40 | 5.20 ab ± 0.54 | 3.98 a* ± 0.40 | ||||||||
| B-36 kDa | GW | 7.47 ± 0.59 | 7.50 ± 0.70 | 7.48 ± 0.52 | 5.73 a ± 0.47 | 6.48 a ± 0.49 | 8.54 b ± 0.50 | * | ns | ns | ns | ns | ns | ns |
| DW | 6.64 ± 0.59 | 6.84 ± 0.70 | 6.48 ± 0.52 | 6.13 ± 0.40 | 6.16 ± 0.49 | 6.78 ± 0.50 | ||||||||
| B-30 kDa | GW | 1.96 ± 0.67 | 2.17 ± 0.37 | 2.41 ± 0.26 | 2.33 ± 0.17 | 2.75 ± 0.38 | 2.16 ± 0.38 | ns | ns | ns | ns | ns | ns | ns |
| DW | 2.10 ± 0.67 | 2.05 ± 0.37 | 2.24 ± 0.26 | 2.39 ± 0.15 | 2.94 ± 0.38 | 2.33 ± 0.38 | ||||||||
| B-28 kDa | GW | 4.33 ± 0.38 | 5.41 ± 0.25 | 4.45 ± 0.18 | 4.14 ± 0.26 | 4.60 ± 0.32 | 5.02 ± 0.50 | ns | ns | ns | ns | ns | ns | ns |
| DW | 4.75 ± 0.38 | 4.70 ± 0.21 | 4.75 ± 0.15 | 5.01 ± 0.26 | 4.91 ± 0.28 | 5.36 ± 0.50 | ||||||||
| B-26 kDa | GW | 2.25 a ± 0.29 | 4.33 ab ± 0.30 | 4.56 b ± 0.46 | 2.58 a ± 0.31 | 3.39 b ± 0.39 | 4.80 b ± 0.47 | ** | ns | *** | ns | ns | ns | ns |
| DW | 2.05 a ± 0.29 | 2.71 a* ± 0.26 | 3.98 b ± 0.40 | 2.23 a ± 0.27 | 3.58 b ± 0.34 | 3.74 b ± 0.47 | ||||||||
| B-16 kDa | GW | 2.67 ± 0.48 | 3.04 ± 0.24 | 2.10 ± 0.44 | 3.17 ± 0.32 | 3.54 ± 0.41 | 2.54 ± 0.39 | ns | ns | ns | ns | ns | ns | ns |
| DW | 2.75 ± 0.48 | 2.30 ± 0.21 | 2.83 ± 0.44 | 3.11 ± 0.32 | 2.56 ± 0.41 | 2.47 ± 0.39 | ||||||||
| ST | LT | |||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Farm Origin | Post-Mortem Time (days) | Post-Mortem Time (days) | FO | TD | AT | FO*TD | FO*AT | TD*AT | FO*TD*AT | |||||
| 3 | 8 | 21 | 3 | 8 | 21 | |||||||||
| MHC † | GW | 31.05 ± 0.65 | 30.48 ± 0.53 | 31.42 ± 0.91 | 31.76 ± 0.23 | 31.25 ± 0.26 | 30.40 ± 0.56 | ns | ns | ns | ns | ns | ns | ns |
| DW | 30.83 ± 0.65 | 29.87 ± 0.62 | 29.74 ± 0.91 | 31.05 b ± 0.26 | 29.99 a ± 0.22 | 29.82 a ± 0.56 | ||||||||
| Protein C ‡ | GW | 1.42 ± 0.32 | 1.11 ± 0.26 | 1.11 ± 0.23 | 1.91 ± 0.20 | 2.05 ± 0.41 | 1.93 ± 0.12 | ns | ** | ns | ns | ns | ns | ns |
| DW | 1.66 ± 0.32 | 0.85 ± 0.30 | 0.97 ± 0.23 | 1.64 * ± 0.20 | 1.12 * ± 0.41 | 1.19 * ± 0.12 | ||||||||
| α-actinin § | GW | 2.89 ± 0.27 | 2.55 ± 0.35 | 2.38 ± 0.21 | 2.65 ± 0.20 | 2.32 ± 0.16 | 2.41 ± 0.18 | ns | ns | ns | ns | ns | ns | ns |
| DW | 1.88 ± 0.27 | 2.04 ± 0.35 | 2.67 ± 0.21 | 2.51 ± 0.20 | 2.74 ± 0.29 | 2.07 ± 0.18 | ||||||||
| Desmin | GW | 2.75 ± 0.17 | 1.98 ± 0.13 | 2.21 ± 0.28 | 2.39 ± 0.13 | 1.53 ± 0.32 | 2.07 ± 0.24 | * | ns | ns | ns | ns | ns | ns |
| DW | 1.99 * ± 0.17 | 1.63 * ± 0.11 | 1.45 * ± 0.28 | 1.71 * ± 0.23 | 1.58 ± 0.32 | 1.55 ± 0.24 | ||||||||
| Actin ϒ | GW | 31.03 ± 0.23 | 31.08 ± 0.60 | 30.28 ± 0.32 | 31.77 ± 0.34 | 30.97 ± 0.53 | 31.16 ± 0.30 | ns | ns | ns | ns | ns | ns | ns |
| DW | 30.59 ± 0.27 | 30.50 ± 0.60 | 29.40 ± 0.32 | 30.91 ± 0.34 | 29.88 ± 0.53 | 29.07 ± 0.26 | ||||||||
| TnT Ϯ | GW | 8.47 b ± 0.69 | 5.94 a ± 0.69 | 6.04 a ± 0.69 | 7.94 b ± 0.36 | 6.68 a ± 0.36 | 6.10 a ± 0.36 | *** | ns | *** | ns | ns | ns | ns |
| DW | 5.85 b* ± 0.45 | 3.74 a* ± 0.45 | 4.37 ab* ± 0.45 | 5.32 * ± 0.65 | 4.34 * ± 0.65 | 4.57 * ± 0.65 | ||||||||
| 35–37 kDa group ψ | GW | 9.96 a ± 0.57 | 12.88 b ± 0.57 | 13.40 b ± 0.57 | 9.85 a ± 0.89 | 12.31 b ± 0.89 | 10.84 b ± 0.89 | *** | * | ** | ns | ns | ns | ns |
| DW | 14.76 * ± 0.46 | 16.30 * ± 0.46 | 15.58 * ± 0.46 | 12.68 a#* ± 0.83 | 14.86 b* ± 0.83 | 15.10 b* ± 0.83 | ||||||||
| 26–30 kDa Group ϐ | GW | 11.91 a ± 0.48 | 13.20 ab ± 0.51 | 14.21 b ± 0.42 | 11.8 a ± 0.52 | 13.9 b ± 0.58 | 14.2 b ± 0.46 | *** | ns | *** | ns | ns | ns | ns |
| DW | 13.10 a ± 0.48 | 16.12 b* ± 0.51 | 15.90 b* ± 0.42 | 12.3 a ± 0.52 | 15.7 b* ± 0.58 | 16.6 b* ± 0.46 | ||||||||
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Martínez, T.F.; Alcalde, M.J.; Sáez, M.I.; Suárez, M.D. Effects of Farm Management Practices and Transport Time on Post-Mortem Changes of Longissimus lumborum Muscle Proteins in Suckling Goat Kids. Foods 2020, 9, 934. https://doi.org/10.3390/foods9070934
Martínez TF, Alcalde MJ, Sáez MI, Suárez MD. Effects of Farm Management Practices and Transport Time on Post-Mortem Changes of Longissimus lumborum Muscle Proteins in Suckling Goat Kids. Foods. 2020; 9(7):934. https://doi.org/10.3390/foods9070934
Chicago/Turabian StyleMartínez, Tomás Francisco, María Jesús Alcalde, María Isabel Sáez, and María Dolores Suárez. 2020. "Effects of Farm Management Practices and Transport Time on Post-Mortem Changes of Longissimus lumborum Muscle Proteins in Suckling Goat Kids" Foods 9, no. 7: 934. https://doi.org/10.3390/foods9070934
APA StyleMartínez, T. F., Alcalde, M. J., Sáez, M. I., & Suárez, M. D. (2020). Effects of Farm Management Practices and Transport Time on Post-Mortem Changes of Longissimus lumborum Muscle Proteins in Suckling Goat Kids. Foods, 9(7), 934. https://doi.org/10.3390/foods9070934