In Vitro Modulation of Gut Microbiota and Metabolism by Cooked Cowpea and Black Bean

 ,
,  and
and
Abstract

1. Introduction
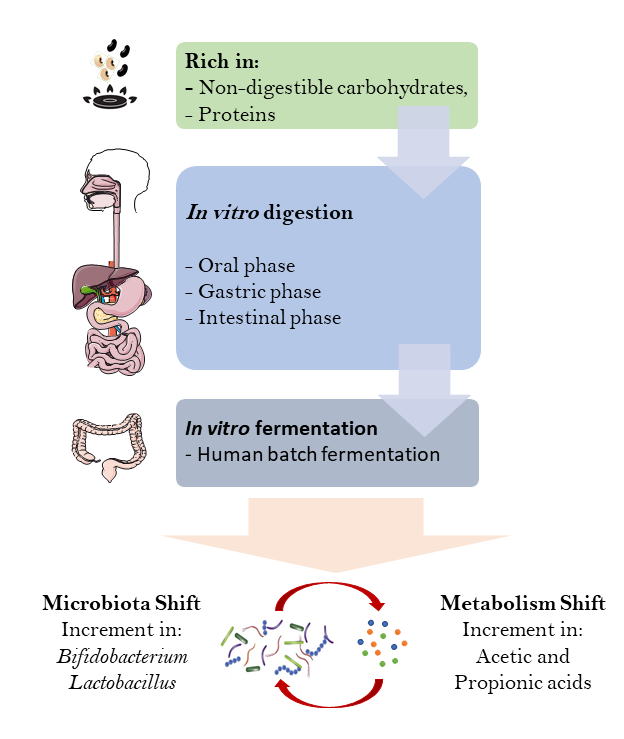
2. Materials and Methods
2.1. Sample Preparation
2.2. Static In Vitro Simulation of Gastrointestinal Food Digestion
2.3. Fecal Sample Preparation
2.4. DNA Extraction and Microbiota Analysis from Fermented Samples
2.5. SCFA Quantification by Gas Chromatography
2.6. pH and Gas Production Assessment
2.7. Statistical Analysis
3. Results
3.1. Changes in Bacteria Population in Fecal Batch Culture
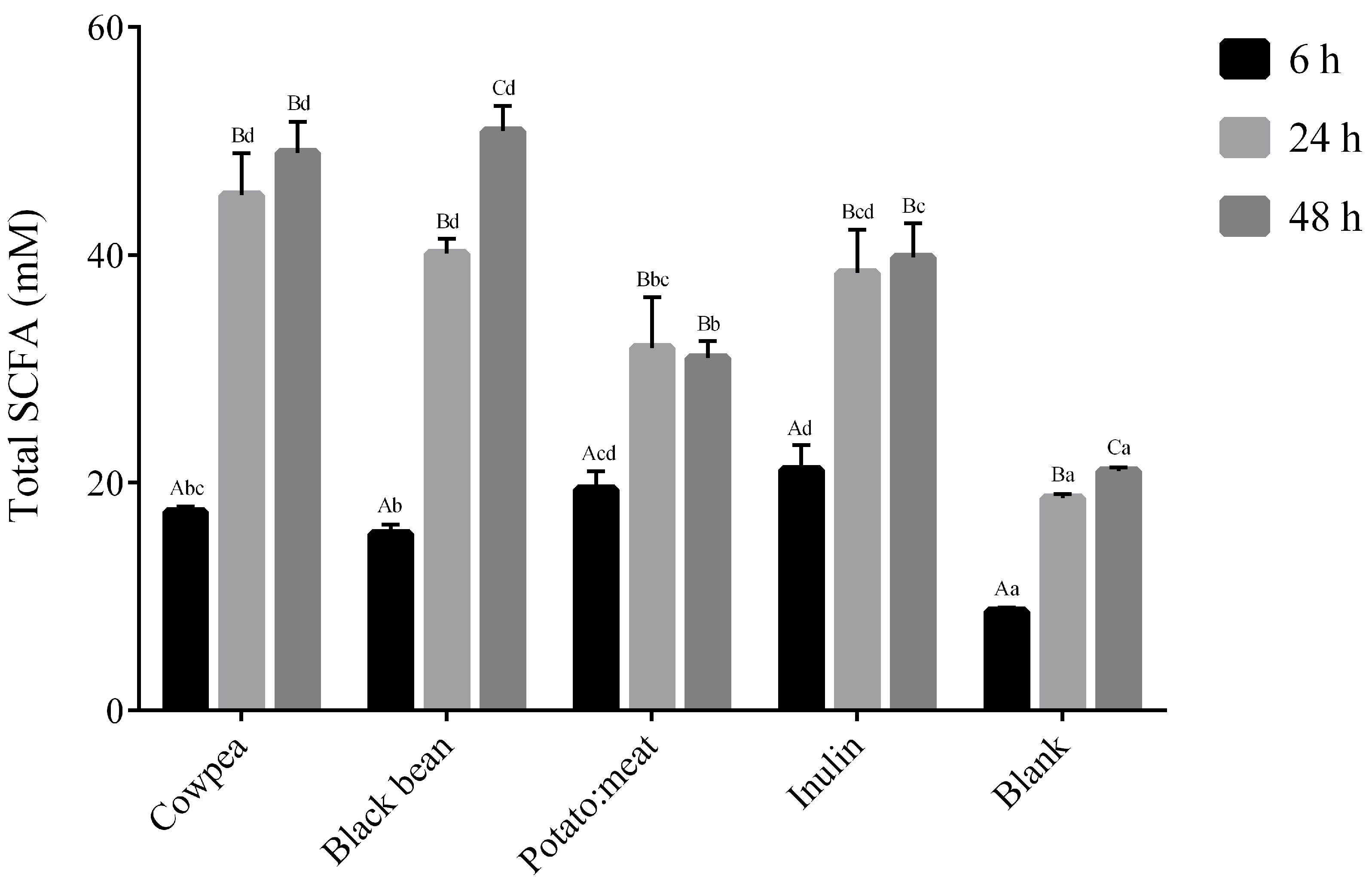
3.2. Production of SCFA, Gas and pH Shift
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Ley, R.E.; Peterson, D.A.; Gordon, J.I. Ecological and evolutionary forces shaping microbial diversity in the human intestine. Cell 2006, 124, 837–848. [Google Scholar] [CrossRef]
- Wang, B.; Yao, M.; Lv, L.; Ling, Z.; Li, L. The Human Microbiota in Health and Disease. Engineering 2017, 3, 71–82. [Google Scholar] [CrossRef]
- Turnbaugh, P.J.; Ley, R.E.; Mahowald, M.A.; Magrini, V.; Mardis, E.R.; Gordon, J.I. An obesity-associated gut microbiome with increased capacity for energy harvest. Nature 2006, 444, 1027–1031. [Google Scholar] [CrossRef] [PubMed]
- Cani, P.D.; Dewever, C.; Delzenne, N.M. Inulin-type fructans modulate gastrointestinal peptides involved in appetite regulation (glucagon-like peptide-1 and ghrelin) in rats. Br. J. Nutr. 2004, 92, 521. [Google Scholar] [CrossRef]
- Bäckhed, F.; Fraser, C.M.; Ringel, Y.; Sanders, M.E.; Sartor, R.B.; Sherman, P.M.; Versalovic, J.; Young, V.; Finlay, B.B. Defining a healthy human gut microbiome: Current concepts, future directions, and clinical applications. Cell Host Microbe 2012, 12, 611–622. [Google Scholar] [CrossRef] [PubMed]
- Bouskra, D.; Brézillon, C.; Bérard, M.; Werts, C.; Varona, R.; Boneca, I.G.; Eberl, G. Lymphoid tissue genesis induced by commensals through NOD1 regulates intestinal homeostasis. Nature 2008, 456, 507–510. [Google Scholar] [CrossRef]
- Hooper, L.V.; Stappenbeck, T.S.; Hong, C.V.; Gordon, J.I. Angiogenins: A new class of microbicidal proteins involved in innate immunity. Nat. Immunol. 2003, 4, 269–273. [Google Scholar] [CrossRef]
- Boon, N.; Top, E.M.; Verstraete, W.; Siciliano, S.D. Bioaugmentation as a tool to protect the structure and function of an activated-sludge microbial community against a 3-chloroaniline shock load. Appl. Environ. Microbiol. 2003, 69, 1511–1520. [Google Scholar] [CrossRef]
- Raskov, H.; Burcharth, J.; Pommergaard, H.C. Linking gut microbiota to colorectal cancer. J. Cancer 2017, 8, 3378–3395. [Google Scholar] [CrossRef]
- Kho, Z.Y.; Lal, S.K. The human gut microbiome—A potential controller of wellness and disease. Front. Microbiol. 2018, 9, 1–23. [Google Scholar] [CrossRef]
- Maccaferri, S.; Klinder, A.; Cacciatore, S.; Chitarrari, R.; Honda, H.; Luchinat, C.; Bertini, I.; Carnevali, P.; Gibson, G.R.; Brigidi, P.; et al. In vitro fermentation of potential prebiotic flours from natural sources: Impact on the human colonic microbiota and metabolome. Mol. Nutr. Food Res. 2012, 56, 1342–1352. [Google Scholar] [CrossRef]
- Roberfroid, M.R.; Gibson, G.; Hoyles, L.; McCartney, A.L.; Rastall, R.; Rowland, I.; Wolvers, D.; Watzl, B.; Szajewska, H.; Stahl, B.; et al. Prebiotic effects metabolic and health benefits. Br. J. Nutr. 2010, 104, S1–S63. [Google Scholar] [CrossRef] [PubMed]
- Gibson, G.R.; Hutkins, R.; Sanders, M.E.; Prescott, S.L.; Reimer, R.A.; Salminen, S.J.; Scott, K.; Stanton, C.; Swanson, K.S.; Cani, P.D.; et al. Expert consensus document: The International Scientific Association for Probiotics and Prebiotics (ISAPP) consensus statement on the definition and scope of prebiotics. Nat. Rev. Gastroenterol. Hepatol. 2017, 14, 491–502. [Google Scholar] [CrossRef] [PubMed]
- Gullón, P.; Gullón, B.; Tavaria, F.; Vasconcelos, M.; Gomes, A.M. In vitro fermentation of lupin seeds (Lupinus albus) and broad beans (Vicia faba): Dynamic modulation of the intestinal microbiota and metabolomic output. Food Funct. 2015, 6, 3316–3322. [Google Scholar] [CrossRef] [PubMed]
- Beards, E.; Tuohy, K.; Gibson, G. Bacterial, SCFA and gas profiles of a range of food ingredients following in vitro fermentation by human colonic microbiota. Anaerobe 2010, 16, 420–425. [Google Scholar] [CrossRef] [PubMed]
- Scott, K.P.; Duncan, S.H.; Flint, H.J. Dietary fibre and the gut microbiota. Br. Nutr. Fund. 2008, 33, 201–211. [Google Scholar] [CrossRef]
- Johnson, C.R.; Thavarajah, D.; Combs, G.F.; Thavarajah, P. Lentil (Lens culinaris L.): A prebiotic-rich whole food legume. Food Res. Int. 2013, 51, 107–113. [Google Scholar] [CrossRef]
- Siah, S.D.; Konczak, I.; Agboola, S.; Wood, J.A.; Blanchard, C.L. In vitro investigations of the potential health benefits of Australian-grown faba beans (Vicia faba L.): Chemopreventative capacity and inhibitory effects on the angiotensin-converting enzyme, α-glucosidase and lipase. Br. J. Nutr. 2012, 108, S123–S134. [Google Scholar] [CrossRef]
- Kalogeropoulos, N.; Chiou, A.; Ioannou, M.; Karathanos, V.T.; Hassapidou, M.; Andrikopoulos, N.K. Nutritional evaluation and bioactive microconstituents (phytosterols, tocopherols, polyphenols, triterpenic acids) in cooked dry legumes usually consumed in the Mediterranean countries. Food Chem. 2010, 121, 682–690. [Google Scholar] [CrossRef]
- Luna-Vital, D.A.; Liang, K.; González de Mejía, E.; Loarca-Piña, G. Dietary peptides from the non-digestible fraction of: Phaseolus vulgaris L. decrease angiotensin II-dependent proliferation in HCT116 human colorectal cancer cells through the blockade of the renin-angiotensin system. Food Funct. 2016, 7, 1–11. [Google Scholar] [CrossRef]
- Chávez-Mendoza, C.; Sánchez, E. Bioactive compounds from mexican varieties of the common bean (Phaseolus vulgaris): Implications for health. Molecules 2017, 22, 1–32. [Google Scholar] [CrossRef] [PubMed]
- Ganesan, K.; Xu, B. Polyphenol-rich dry common beans (Phaseolus vulgaris L.) and their health benefits. Int. J. Mol. Sci. 2017, 18, 2331. [Google Scholar] [CrossRef] [PubMed]
- Tiwari, B.; Gowen, A.; McKnna, B. Pulse Foods Processing, Quality and Nutraceutical Applications, 1st ed.; Tiwari, B., Gowen, A., McKenna, B., Eds.; Academic Press: Cambridge, MA, USA, 2011; ISBN 9780123724861. [Google Scholar]
- Singh, J.; Basu, P.S. Non-Nutritive Bioactive Compounds in Pulses and Their Impact on Human Health: An Overview. Food Nutr. Sci. 2012, 03, 1664–1672. [Google Scholar] [CrossRef]
- Singh, R.K.; Chang, H.W.; Yan, D.; Lee, K.M.; Ucmak, D.; Wong, K.; Abrouk, M.; Farahnik, B.; Nakamura, M.; Zhu, T.H.; et al. Influence of diet on the gut microbiome and implications for human health. J. Transl. Med. 2017, 15, 1–17. [Google Scholar] [CrossRef]
- Borresen, E.C.; Zhang, L.; Trehan, I.; Nealon, N.J.; Maleta, K.M.; Manary, M.J.; Ryan, E.P. The Nutrient and Metabolite Profile of 3 Complementary Legume Foods with Potential to Improve Gut Health in Rural Malawian Children. Curr. Dev. Nutr. 2017, 1, e001610. [Google Scholar] [CrossRef]
- Minekus, M.; Alminger, M.; Alvito, P.; Ballance, S.; Bohn, T.; Bourlieu, C.; Carrière, F.; Boutrou, R.; Corredig, M.; Dupont, D.; et al. A standardised static in vitro digestion method suitable for food—An international consensus. Food Funct. 2014, 5, 1113–1124. [Google Scholar] [CrossRef]
- Sánchez-Moya, T.; López-Nicolás, R.; Planes, D.; González-Bermúdez, C.A.; Ros-Berruezo, G.; Frontela-Saseta, C. In vitro modulation of gut microbiota by whey protein to preserve intestinal health. Food Funct. 2017, 8, 3053–3063. [Google Scholar] [CrossRef] [PubMed]
- González-Bermúdez, C.A.; López-Nicolás, R.; Peso-Echarri, P.; Frontela-Saseta, C.; Martínez-Graciá, C. Effects of different thickening agents on infant gut microbiota. Food Funct. 2018, 9, 1768–1778. [Google Scholar] [CrossRef] [PubMed]
- Lauber, C.; Zhou, N.; Gordon, J.; Knight, R.; Fierer, N. Effect of storage conditions on the assessment of bacterial community structure in soil and human-associated samples. FEMS Microbiol Lett. 2010, 307, 80–86. [Google Scholar] [CrossRef]
- Arboleya, S.; Salazar, N.; Solís, G.; Fernández, N.; Gueimonde, M.; de los Reyes-Gavilán, C.G. In vitro evaluation of the impact of human background microbiota on the response to Bifidobacterium strains and fructo-oligosaccharides. Br. J. Nutr. 2013, 110, 2030–2036. [Google Scholar] [CrossRef]
- Abdul Rahim, M.B.H.; Chilloux, J.; Martinez-Gili, L.; Neves, A.L.; Myridakis, A.; Gooderham, N.; Dumas, M.E. Diet-induced metabolic changes of the human gut microbiome: Importance of short-chain fatty acids, methylamines and indoles. Acta Diabetol. 2019, 56, 493–500. [Google Scholar] [CrossRef] [PubMed]
- Rinninella, E.; Raoul, P.; Cintoni, M.; Franceschi, F.; Miggiano, G.A.D.; Gasbarrini, A.; Mele, M.C. What is the healthy gut microbiota composition? A changing ecosystem across age, environment, diet, and diseases. Microorganisms 2019, 7, 14. [Google Scholar] [CrossRef] [PubMed]
- Siva, N.; Thavarajah, P.; Kumar, S.; Thavarajah, D. Variability in prebiotic carbohydrates in different market classes of chickpea, common bean, and lentil collected from the american local market. Front. Nutr. 2019, 6, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Shen, Q.; Chen, Y.A.; Tuohy, K.M. A comparative in vitro investigation into the effects of cooked meats on the human faecal microbiota. Anaerobe 2010, 16, 572–577. [Google Scholar] [CrossRef]
- Wang, M.; Wichienchot, S.; He, X.; Fu, X.; Huang, Q.; Zhang, B. In vitro colonic fermentation of dietary fibers: Fermentation rate, short-chain fatty acid production and changes in microbiota. Trends Food Sci. Technol. 2019, 88, 1–9. [Google Scholar] [CrossRef]
- Tsitko, I.; Wiik-Miettinen, F.; Mattila, O.; Rosa-Sibakov, N.; Seppänen-Laakso, T.; Maukonen, J.; Nordlund, E.; Saarela, M. A small in vitro fermentation model for screening the gut microbiota effects of different fiber preparations. Int. J. Mol. Sci. 2019, 20, 1–16. [Google Scholar] [CrossRef]
- Jian, C.; Luukkonen, P.; Yki-Järvinen, H.; Salonen, A.; Korpela, K. Quantitative PCR provides a simple and accessible method for quantitative microbiota profiling. PLoS ONE 2020, 15, 1–10. [Google Scholar] [CrossRef]
- Rodriguez-Colinas, B.; Kolida, S.; Baran, M.; Ballesteros, A.O.; Rastall, R.A.; Plou, F.J. Analysis of fermentation selectivity of purified galacto-oligosaccharides by in vitro human faecal fermentation. Appl. Microbiol. Biotechnol. 2013, 97, 5743–5752. [Google Scholar] [CrossRef]
- Hernandez-Hernandez, O.; Côté, G.L.; Kolida, S.; Rastall, R.A.; Sanz, M.L. In vitro fermentation of alternansucrase raffinose-derived oligosaccharides by human gut bacteria. J. Agric. Food Chem. 2011, 59, 10901–10906. [Google Scholar] [CrossRef]
- Wallace, R.J.; McKain, N. Peptidase activity of human colonic bacteria. Anaerobe 1997, 3, 251–257. [Google Scholar] [CrossRef]
- Walker, A.W.; Duncan, S.H.; Carol McWilliam Leitch, E.; Child, M.W.; Flint, H.J. pH and peptide supply can radically alter bacterial populations and short-chain fatty acid ratios within microbial communities from the human colon. Appl. Environ. Microbiol. 2005, 71, 3692–3700. [Google Scholar] [CrossRef] [PubMed]
- Rowland, I.; Gibson, G.; Heinken, A.; Scott, K.; Swann, J.; Thiele, I.; Tuohy, K. Gut microbiota functions: Metabolism of nutrients and other food components. Eur. J. Nutr. 2018, 57, 1–24. [Google Scholar] [CrossRef] [PubMed]
- Cruz-Bravo, R.K.; Guevara-González, R.G.; Ramos-Gómez, M.; Oomah, B.D.; Wiersma, P.; Campos-Vega, R.; Loarca-Piña, G. The fermented non-digestible fraction of common bean (Phaseolus vulgaris L.) triggers cell cycle arrest and apoptosis in human colon adenocarcinoma cells. Genes Nutr. 2014, 9, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Puertollano, E.; Kolida, S.; Yaqoob, P. Biological significance of short-chain fatty acid metabolism by the intestinal microbiome. Curr. Opin. Clin. Nutr. Metab. Care 2014, 17, 139–144. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Bacteria | Primers |
|---|---|
| Bifidobacterium | Fwd: GATTCTGGCTCAGGATGAACGC |
| Rv: CTGATAGGACGCGACCCCAT | |
| Lactobacillus | Fwd: AGCAGTAGGGAATCTTCCA |
| Rv: CATGGAGTTCCACTGTCCTC | |
| Enterobacteriaceae | Fwd: TGCCGTAACTTCGGGAGAAGGCA |
| Rv: TCAAGGACCAGTGTTCAGTGTC | |
| Firmicutes | Fwd: GGAGYATGTGGTTTAATTCGAAGCA |
| Rv: AGCTGACGACAACCATGCAC | |
| Bacteroides–Prevotella | Fwd: GAGAGGAAGGTCCCCCAC |
| Rv: CGKACTTGGCTGGTTCAG |
| Time (h) | Bifidobacterium | Lactobacillus | Enterobacteriaceae | Firmicutes | Bacteroides | |
|---|---|---|---|---|---|---|
| Cowpea | 0 | 5.34 ± 0.05Aa | 1.48 ± 0.13Aa | 4.19 ± 0.03Aa | 5.38 ± 0.09Aa | 5.81 ± 0.10Ba |
| 6 | 6.73 ± 0.11Bbc | 3.74 ± 0.14Cb | 5.04 ± 0.04Cb | 5.90 ± 0.10BCab | 6.04 ± 0.09Ba | |
| 24 | 7.55 ± 0.24Dd | 3.77 ± 0.17Cb | 4.95 ± 0.07Cc | 6.02 ± 0.13Cc | 4.87 ± 0.20Ab | |
| 48 | 7.20 ± 0.16Cd | 3.40 ± 0.12Bb | 4.67 ± 0.07Bd | 5.70 ± 0.11Bbc | 4.61 ± 0.21Ab | |
| Black bean | 0 | 5.34 ± 0.05Aa | 1.48 ± 0.13Aa | 4.19 ± 0.03Aa | 5.38 ± 0.09Aa | 5.81 ± 0.10Ba |
| 6 | 6.67 ± 0.21Bbc | 3.53 ± 0.29Bb | 4.99 ± 0.13Cb | 5.78 ± 0.29Cab | 6.06 ± 0.31Ba | |
| 24 | 7.31 ± 0.04Ccd | 3.56 ± 0.02Bb | 4.69 ± 0.08Bb | 5.75 ± 0.03BCbc | 4.71 ± 0.02Ab | |
| 48 | 6.89 ± 0.13Bc | 3.43 ± 0.09Bb | 4.35 ± 0.17Ac | 5.42 ± 0.11ABab | 4.66 ± 0.17Ab | |
| Potato:beef | 0 | 5.34 ± 0.05Aa | 1.48 ± 0.13Aa | 4.19 ± 0.03Aa | 5.38 ± 0.09Aa | 5.81 ± 0.10Ba |
| 6 | 7.10 ± 0.18Cc | 4.20 ± 0.27Cb | 5.36 ± 0.17Bc | 6.28 ± 0.20Bb | 6.23 ± 0.20Ba | |
| 24 | 6.63 ± 0.12Bb | 3.43 ± 0.25Bb | 5.20 ± 0.06Bc | 5.55 ± 0.25Ab | 3.84 ± 0.14Aa | |
| 48 | 6.58 ± 0.08Bb | 3.84 ± 0.06BCc | 5.11 ± 0.21Be | 5.37 ± 0.16Aa | 3.53 ± 0.49Aa | |
| Inulin | 0 | 5.34 ± 0.05Aa | 1.48 ± 0.13Aa | 4.19 ± 0.03Aa | 5.38 ± 0.09Aa | 5.81 ± 0.10ABa |
| 6 | 6.76 ± 0.52BCbc | 2.45 ± 0.61Ba | 4.89 ± 0.22Bb | 6.06 ± 0.44BCab | 6.18 ± 0.53Ba | |
| 24 | 7.05 ± 0.11Cc | 4.19 ± 0.11Cc | 4.72 ± 0.15Bb | 6.50 ± 0.15Cd | 5.97 ± 0.15ABd | |
| 48 | 6.46 ± 0.11Bb | 3.83 ± 0.22Cc | 3.96 ± 0.17Aab | 5.92 ± 0.13Bc | 5.39 ± 0.12Ac | |
| Blank | 0 | 5.34 ± 0.05Aa | 1.48 ± 0.13Aa | 4.19 ± 0.03Ba | 5.38 ± 0.09Aa | 5.81 ± 0.10Aa |
| 6 | 5.54 ± 0.26Aa | 1.76 ± 0.22Aa | 4.84 ± 0.19Cb | 5.65 ± 0.20Ba | 6.23 ± 0.25Ba | |
| 24 | 5.26 ± 0.12Aa | 1.50 ± 0.20Aa | 4.27 ± 0.04Ba | 5.18 ± 0.02Aa | 5.64 ± 0.05Ac | |
| 48 | 4.93 ± 0.07Aa | 1.63 ± 0.09Aa | 3.72 ± 0.06Aa | 5.19 ± 0.05Aa | 5.52 ± 0.10Ac |
| Time (h) | Acetate (mM) | Propionate (mM) | Butyrate (mM) | |
|---|---|---|---|---|
| 6 | 11.79 ± 0.44Abc | 4.48 ± 0.09Ac | 1.17 ± 0.04Aab | |
| Cowpea | 24 | 38.09 ± 3.64Bd | 5.67 ± 0.10Bb | 1.45 ± 0.10Aab |
| 48 | 41.45 ± 2.92Bd | 5.97 ± 0.46Bb | 1.55 ± 0.31Aa | |
| 6 | 10.28 ± 0.60Ab | 4.18 ± 0.22Ac | 1.04 ± 0.01Aa | |
| Black bean | 24 | 34.09 ± 3.64Bcd | 5.67 ± 0.10Bb | 1.35 ± 0.09Ba |
| 48 | 43.45 ± 2.20Cd | 5.80 ± 0.03Cb | 1.61 ± 0.02Ba | |
| 6 | 13.13 ± 1.54Acd | 4.85 ± 0.42Ac | 1.45 ± 0.14Abc | |
| Potato:meat | 24 | 24.07 ± 3.22Bb | 6.00 ± 0.89Ab | 1.77 ± 0.34Abc |
| 48 | 23.14 ± 1.19Ba | 5.89 ± 0.64Ab | 1.91 ± 0.32Aa | |
| 6 | 14.46 ± 1.59Ad | 4.24 ± 0.40Ac | 2.43 ± 0.18Ad | |
| Inulin | 24 | 29.19 ± 2.5Bbc | 5.36 ± 0.37Ba | 3.59 ± 0.20Bd |
| 48 | 30.83 ± 2.89Bc | 5.24 ± 0.15Bb | 3.69 ± 0.05Bc | |
| 6 | 5.11 ± 0.03Aa | 2.20 ± 0.27Aa | 1.05 ± 0.10Aa | |
| Blank | 24 | 11.36 ± 0.55Ba | 3.04 ± 0.02Ba | 2.01 ± 0.04Bc |
| 48 | 12.57 ± 0.01Ca | 3.66 ± 0.26Ca | 2.49 ± 0.03Cb |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Teixeira-Guedes, C.; Sánchez-Moya, T.; Pereira-Wilson, C.; Ros-Berruezo, G.; López-Nicolás, R. In Vitro Modulation of Gut Microbiota and Metabolism by Cooked Cowpea and Black Bean. Foods 2020, 9, 861. https://doi.org/10.3390/foods9070861
Teixeira-Guedes C, Sánchez-Moya T, Pereira-Wilson C, Ros-Berruezo G, López-Nicolás R. In Vitro Modulation of Gut Microbiota and Metabolism by Cooked Cowpea and Black Bean. Foods. 2020; 9(7):861. https://doi.org/10.3390/foods9070861
Chicago/Turabian StyleTeixeira-Guedes, Catarina, Tereza Sánchez-Moya, Cristina Pereira-Wilson, Gaspar Ros-Berruezo, and Rubén López-Nicolás. 2020. "In Vitro Modulation of Gut Microbiota and Metabolism by Cooked Cowpea and Black Bean" Foods 9, no. 7: 861. https://doi.org/10.3390/foods9070861
APA StyleTeixeira-Guedes, C., Sánchez-Moya, T., Pereira-Wilson, C., Ros-Berruezo, G., & López-Nicolás, R. (2020). In Vitro Modulation of Gut Microbiota and Metabolism by Cooked Cowpea and Black Bean. Foods, 9(7), 861. https://doi.org/10.3390/foods9070861