Effect of Rearing System on the Straight and Branched Fatty Acids of Goat Milk and Meat of Suckling Kids

 ,
,  , and
, and 
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Milk Sampling
2.3. Carcass Sampling
2.4. Fatty Acid Analyses
2.5. Statistical Analysis
3. Results
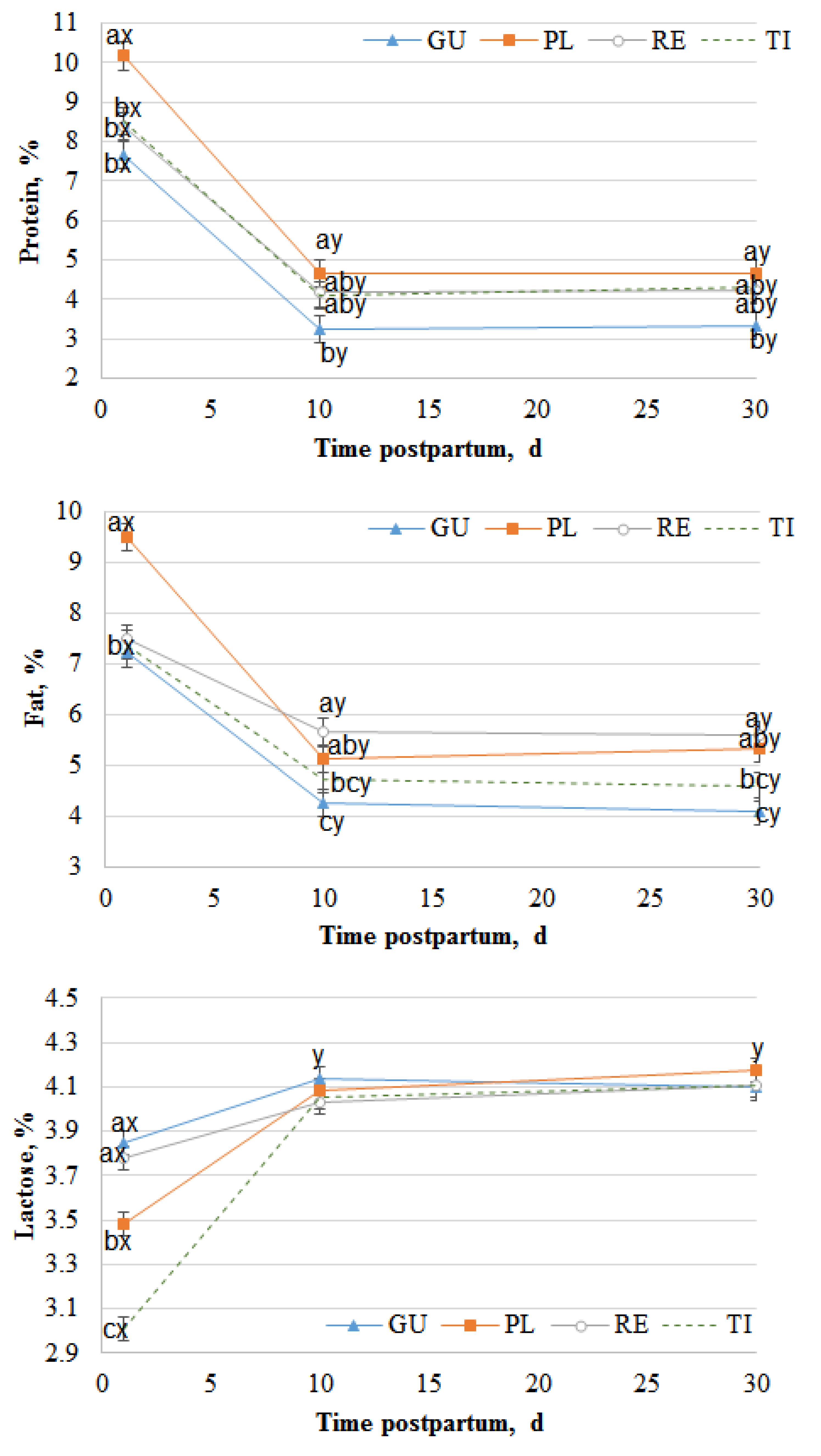
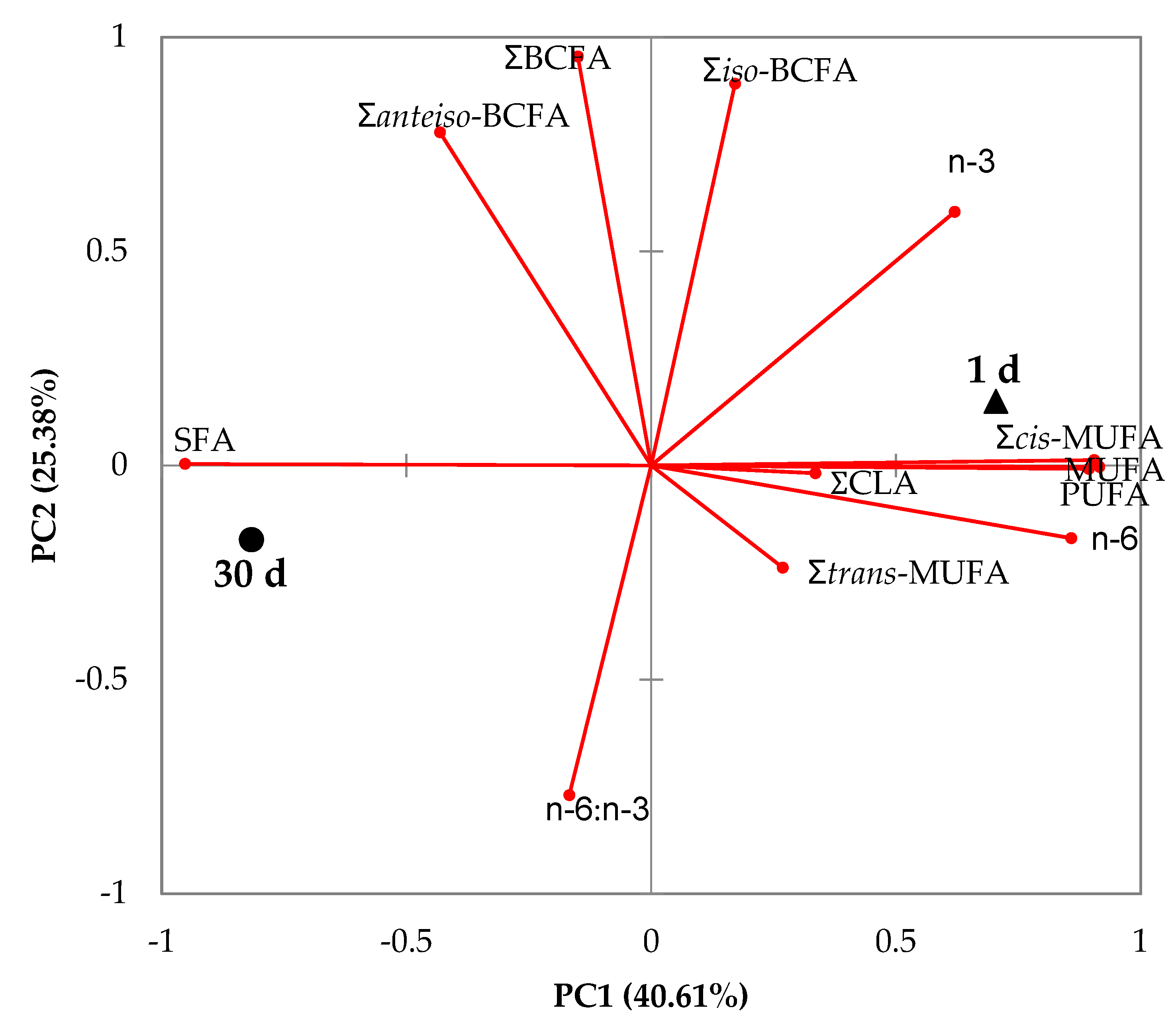
3.1. Milk
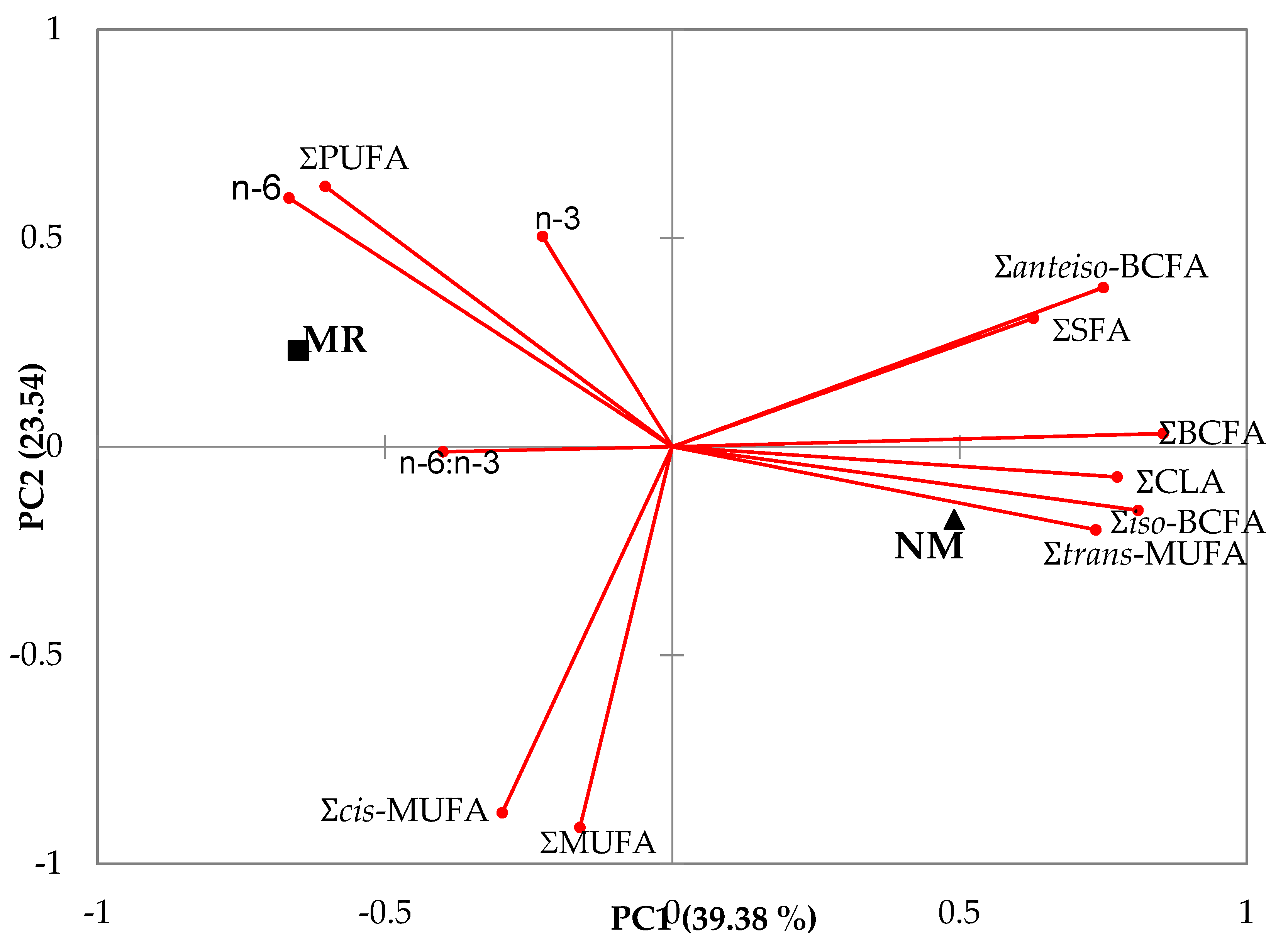
3.2. Meat
4. Discussion
4.1. Milk
4.2. Meat
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- FAO. FAOSTAT Livestock Primary; Food and Agriculture Organization: Rome, Italy, 2017. [Google Scholar]
- Yalcintan, H.; Akin, P.D.; Ozturk, N.; Ekiz, B.; Kocak, O.; Yilmaz, A. Carcass and meat quality traits of Saanen goat kids reared under natural and artificial systems and slaughtered at different ages. Acta Vet. Brno 2018, 87, 293–300. [Google Scholar] [CrossRef]
- Aparnathi, K.; Mehta, B.; Velpula, S. Goat Milk in Human Nutrition and Health—A Review. Int. J. Curr. Microbiol. Appl. Sci. 2017, 6, 1781–1792. [Google Scholar] [CrossRef]
- Castel, J.M.; Mena, Y.; Ruiz, F.A.; Gutiérrez, R. Situación y evolución de los sistemas de producción caprina en España. Tierras Caprino 2012, 1, 24–37. [Google Scholar]
- Marichal, A.; Castro, N.; Capote, J.; Zamorano, N.; Arguello, A. Effects of live weight at slaughter (6, 10 and 25 kg) on kid carcass and meat quality. Livest. Prod. Sci. 2003, 83, 247–256. [Google Scholar] [CrossRef]
- Argüello, A.; Castro, N.; Capote, J.; Solomon, M. Effects of diet and live weight at slaughter on kid meat quality. Meat Sci. 2005, 70, 173–179. [Google Scholar] [CrossRef]
- Ripoll, G.; Alcalde, M.J.; Argüello, A.; Córdoba, M.G.; Panea, B. Effect of the rearing system on the color of four muscles of suckling kids. Food Sci. Nutr. 2019, 7, 1502–1511. [Google Scholar] [CrossRef]
- Ripoll, G.; Alcalde, M.J.; Argüello, A.; Córdoba, M.G.; Panea, B. Consumer visual appraisal and shelf life of leg chops from suckling kids raised with natural milk or milk replacer. J. Sci. Food Agric. 2018, 98, 2651–2657. [Google Scholar] [CrossRef]
- Ripoll, G.; Alcalde, M.J.; Argüello, A.; Panea, B. Web-based survey of consumer preferences for the visual appearance of meat from suckling kids. Ital. J. Anim. Sci. 2019, 18, 284–1293. [Google Scholar] [CrossRef]
- Ripoll, G.; Córdoba, M.G.; Alcalde, M.J.; Martín, A.; Argüello, A.; Casquete, R.; Panea, B. Volatile organic compounds and consumer preference for meat from suckling goat kids raised with natural or replacers milk. Ital. J. Anim. Sci. 2019, 18, 1259–1270. [Google Scholar] [CrossRef]
- Ribeiro, R.D.X.; Medeiros, A.N.; Oliveira, R.L.; de Araújo, G.G.L.; Queiroga, R.d.C.d.E.; Ribeiro, M.D.; Silva, T.M.; Bezerra, L.R.; Oliveira, R.L. Palm kernel cake from the biodiesel industry in goat kid diets. Part 2: Physicochemical composition, fatty acid profile and sensory attributes of meat. Small Rumin. Res. 2018, 165, 1–7. [Google Scholar] [CrossRef]
- PAHO/WHO. Trans. fats free Americas. In Conclusions and Recommendations; PAHO/WHO: Washington, DC, USA, 2007; p. 5. [Google Scholar]
- Alonso, L.; Fontecha, F.J.; Fraga, M.J.; Juárez, M.; Lozada, L. Fatty acid composition of caprine milk: Major, branched-chain, and trans fatty acids. J. Dairy Sci. 1999, 82, 878–884. [Google Scholar] [CrossRef]
- Woo, A.; Lindsay, R. Concentrations of major free fatty acids and flavor development in Italian cheese varieties. J. Dairy Sci. 1984, 67, 960–968. [Google Scholar] [CrossRef]
- Watkins, P.J.; Kearney, G.; Rose, G.; Allen, D.; Ball, A.J.; Pethick, D.W.; Warner, R.D. Effect of branched-chain fatty acids, 3-methylindole and 4-methylphenol on consumer sensory scores of grilled lamb meat. Meat Sci. 2014, 96, 1088–1094. [Google Scholar] [CrossRef] [PubMed]
- Brennand, C.P.; Ha, J.K.; Lindsay, R.C. Aroma properties and thresholds of some branched-chaiin and other minor volatile fatty acids occurring in milk fat and meat lipids. J. Sens. Stud. 1989, 4, 105–120. [Google Scholar] [CrossRef]
- Ran-Ressler, R.R.; Devapatla, S.; Lawrence, P.; Brenna, J.T. Branched chain fatty acids are constituents of the normal healthy newborn gastrointestinal tract. Pediatric Res. 2008, 64, 605. [Google Scholar] [CrossRef] [PubMed]
- Yang, Z.; Liu, S.; Chen, X.; Chen, H.; Huang, M.; Zheng, J. Induction of apoptotic cell death and in vivo growth inhibition of human cancer cells by a saturated branched-chain fatty acid, 13-methyltetradecanoic acid. Cancer Res. 2000, 60, 505–509. [Google Scholar] [PubMed]
- Duncan, W.R.H.; Garton, G.A. Differences in the proportions of branched-chain fatty acids in subcutaneous triacylglycerols of barley-fed ruminants. Br. J. Nutr. 2007, 40, 29–33. [Google Scholar] [CrossRef]
- Serra, A.; Mele, M.; La Comba, F.; Conte, G.; Buccioni, A.; Secchiari, P. Conjugated Linoleic Acid (CLA) content of meat from three muscles of Massese suckling lambs slaughtered at different weights. Meat Sci. 2009, 81, 396–404. [Google Scholar] [CrossRef][Green Version]
- E.U. Directive 2010/63/EU of the European Parliament and of the Council of 22 September 2010 on the protection of animals used for scientific purposes. Off. J. Eur. Union L 2010, 276, 33–79. [Google Scholar]
- E.U. Council Regulation (EC) No 1099/2009 of 24 September 2009 on the protection of animals at the time of killing. Off. J. Eur. Union 2009, 303, 1–30. [Google Scholar]
- Molkentin, J.; Precht, D. Validation of a gas-chromatographic method for the determination of milk fat contents in mixed fats by butyric acid analysis. Eur. J. Lipid Sci. Technol. 2000, 102, 194–201. [Google Scholar] [CrossRef]
- Folch, J.; Lees, M.; Stanley, G. A simple method for the isolation and purification of lipids from animal tissues. J. Biol. Chem. 1957, 226, 4977509. [Google Scholar]
- Huerta-Leidenz, N.; Cross, H.; Lunt, D.; Pelton, L.; Savell, J.; Smith, S. Growth, carcass traits, and fatty acid profiles of adipose tissues from steers fed whole cottonseed. J. Anim. Sci. 1991, 69, 3665–3672. [Google Scholar] [CrossRef] [PubMed]
- Sanchez-Macias, D.; Moreno-Indias, I.; Castro, N.; Morales-Delanuez, A.; Arguello, A. From goat colostrum to milk: Physical, chemical, and immune evolution from partum to 90 days postpartum. J. Dairy Sci. 2014, 97, 10–16. [Google Scholar] [CrossRef]
- Sanchez-Macias, D.; Fresno, M.; Moreno-Indias, I.; Castro, N.; Morales-delaNuez, A.; Alvarez, S.; Arguello, A. Physicochemical analysis of full-fat, reduced-fat, and low-fat artisan-style goat cheese. J. Dairy Sci. 2010, 93, 3950–3956. [Google Scholar] [CrossRef]
- Linzell, J.; Peaker, M. Changes in colostrum composition and in the permeability of the mammary epithelium at about the time of parturition in the goat. J. Physiol. 1974, 243, 129–151. [Google Scholar] [CrossRef]
- Pecka-Kiełb, E.; Zachwieja, A.; Wojtas, E.; Zawadzki, W. Influence of nutrition on the quality of colostrum and milk of ruminants. Mljekarstvo Časopis za Unaprjeđenje Proizvodnje i Prerade Mlijeka 2018, 68, 169–181. [Google Scholar] [CrossRef]
- Almeida, O.C.; Ferraz, M.V.C.; Susin, I.; Gentil, R.S.; Polizel, D.M.; Ferreira, E.M.; Barroso, J.P.R.; Pires, A.V. Plasma and milk fatty acid profiles in goats fed diets supplemented with oils from soybean, linseed or fish. Small Rumin. Res. 2019, 170, 125–130. [Google Scholar] [CrossRef]
- Bobe, G.; Zimmerman, S.; Hammond, E.G.; Freeman, A.E.; Porter, P.A.; Luhman, C.M.; Beitz, D.C. Butter Composition and Texture from Cows with Different Milk Fatty Acid Compositions Fed Fish Oil or Roasted Soybeans. J. Dairy Sci. 2007, 90, 2596–2603. [Google Scholar] [CrossRef]
- Hernandez-Castellano, L.E.; Arguello, A.; Almeida, A.M.; Castro, N.; Bendixen, E. Colostrum protein uptake in neonatal lambs examined by descriptive and quantitative liquid chromatography-tandem mass spectrometry. J. Dairy Sci. 2015, 98, 135–147. [Google Scholar] [CrossRef]
- Palma, M.; Alves, S.P.; Hernandez-Castellano, L.E.; Capote, J.; Castro, N.; Arguello, A.; Matzapetakis, M.; Bessa, R.J.B.; de Almeida, A.M. Mammary gland and milk fatty acid composition of two dairy goat breeds under feed-restriction. J. Dairy Res. 2017, 84, 264–271. [Google Scholar] [CrossRef] [PubMed]
- Moreno-Indias, I.; Sánchez-Macías, D.; Castro, N.; Morales-delaNuez, A.; Hernández-Castellano, L.E.; Capote, J.; Argüello, A. Chemical composition and immune status of dairy goat colostrum fractions during the first 10h after partum. Small Rumin. Res. 2012, 103, 220–224. [Google Scholar] [CrossRef]
- Argüello, A.; Castro, N.; Álvarez, S.; Capote, J. Effects of the number of lactations and litter size on chemical composition and physical characteristics of goat colostrum. Small Rumin. Res. 2006, 64, 53–59. [Google Scholar] [CrossRef]
- Romero, T.; Beltrán, M.C.; Rodríguez, M.; De Olives, A.M.; Molina, M.P. Short communication: Goat colostrum quality: Litter size and lactation number effects. J. Dairy Res. 2013, 96, 7526–7531. [Google Scholar] [CrossRef] [PubMed]
- Capote, J.; Castro, N.; Caja, G.; Fernández, G.; Briggs, H.; Argüello, A. Effects of the frequency of milking and lactation stage on milk fractions and milk composition in Tinerfeña dairy goats. Small Rumin. Res. 2008, 75, 252–255. [Google Scholar] [CrossRef]
- Delgado-Pertíñez, M.; Guzmán-Guerrero, J.L.; Caravaca, F.P.; Castel, J.M.; Ruiz, F.A.; González-Redondo, P.; Alcalde, M.J. Effect of artificial vs. natural rearing on milk yield, kid growth and cost in Payoya autochthonous dairy goats. Small Rumin. Res. 2009, 84, 108–115. [Google Scholar] [CrossRef]
- Nudda, A.; Battacone, G.; Bee, G.; Boe, R.; Castanares, N.; Lovicu, M.; Pulina, G. Effect of linseed supplementation of the gestation and lactation diets of dairy ewes on the growth performance and the intramuscular fatty acid composition of their lambs. Animal 2015, 9, 800–809. [Google Scholar] [CrossRef]
- Lou, X.; Li, J.; Zhang, X.; Wang, J.; Wang, C. Variations in fatty acid composition of Laoshan goat milk from partum to 135 days postpartum. Anim. Sci. J. 2018, 89, 1628–1638. [Google Scholar] [CrossRef]
- Marziali, S.; Guerra, E.; Cerdán-Garcia, C.; Segura-Carretero, A.; Caboni, M.F.; Verardo, V. Effect of early lactation stage on goat colostrum: Assessment of lipid and oligosaccharide compounds. Int. Dairy J. 2018, 77, 65–72. [Google Scholar] [CrossRef]
- LeDoux, M.; Rouzeau, A.; Bas, P.; Sauvant, D. Occurrence of trans-C18: 1 fatty acid isomers in goat milk: Effect of two dietary regimens. J. Dairy Sci. 2002, 85, 190–197. [Google Scholar] [CrossRef]
- Andueza, D.; Rouel, J.; Chilliard, Y.; Leroux, C.; Ferlay, A. Prediction of the goat milk fatty acids by near infrared reflectance spectroscopy. Eur. J. Lipid Sci. Technol. 2013, 115, 612–620. [Google Scholar] [CrossRef]
- Ha, J.K.; Lindsay, R. Release of volatile branched-chain and other fatty acids from ruminant milk fats by various lipases. J. Dairy Sci. 1993, 76, 677–690. [Google Scholar] [PubMed]
- Haenlein, G. Goat milk in human nutrition. Small Rumin. Res. 2004, 51, 155–163. [Google Scholar] [CrossRef]
- Massart-Leën, A.M.; De Pooter, H.; Decloedt, M.; Schamp, N. Composition and variability of the branched-chain fatty acid fraction in the milk of goats and cows. Lipids 1981, 16, 286–292. [Google Scholar] [CrossRef]
- Vlaeminck, B.; Fievez, V.; Cabrita, A.R.J.; Fonseca, A.J.M.; Dewhurst, R.J. Factors affecting odd- and branched-chain fatty acids in milk: A review. Anim. Feed Sci. Technol. 2006, 131, 389–417. [Google Scholar] [CrossRef]
- Albenzio, M.; Santillo, A.; Avondo, M.; Nudda, A.; Chessa, S.; Pirisi, A.; Banni, S. Nutritional properties of small ruminant food products and their role on human health. Small Rumin. Res. 2016, 135, 3–12. [Google Scholar] [CrossRef]
- Calder, P.C. Fatty acids and inflammation: The cutting edge between food and pharma. Eur. J. Pharmacol. 2011, 668, S50–S58. [Google Scholar] [CrossRef]
- Lordan, R.; Tsoupras, A.; Mitra, B.; Zabetakis, I. Dairy fats and cardiovascular disease: Do we really need to be concerned? Foods 2018, 7, 29. [Google Scholar] [CrossRef]
- Van Soest, P. Function of the ruminant forestomach. In Nutritional Ecology of the Ruminant, 2nd ed.; Cornell University Press: Ithaca, NY, USA, 1994; pp. 230–252. [Google Scholar]
- Valvo, M.A.; Lanza, M.; Bella, M.; Fasone, V.; Scerra, M.; Biondi, L.; Priolo, A. Effect of ewe feeding system (grass v. concentrate) on intramuscular fatty acids of lambs raised exclusively on maternal milk. Anim. Sci. 2005, 81, 431–436. [Google Scholar] [CrossRef]
- Osorio, M.T.; Zumalacarregui, J.M.; Figueira, A.; Mateo, J. Fatty acid composition in subcutaneous, intermuscular and intramuscular fat deposits of suckling lamb meat: Effect of milk source. Small Rumin. Res. 2007, 73, 127–134. [Google Scholar] [CrossRef]
- Wood, J.D.; Enser, M.; Fisher, A.V.; Nute, G.R.; Sheard, P.R.; Richardson, R.I.; Hughes, S.I.; Whittington, F.M. Fat deposition, fatty acid composition and meat quality: A review. Meat Sci. 2008, 78, 343–358. [Google Scholar] [CrossRef]
- Horcada, A.; Ripoll, G.; Alcalde, M.J.; Sanudo, C.; Teixeira, A.; Panea, B. Fatty acid profile of three adipose depots in seven Spanish breeds of suckling kids. Meat Sci. 2012, 92, 89–96. [Google Scholar] [CrossRef] [PubMed]
- Chilliard, Y.; Rouel, J.; Leroux, C. Goat’s alpha-s1 casein genotype influences its milk fatty acid composition and delta-9 desaturation ratios. Anim. Feed Sci. Technol. 2006, 131, 474–487. [Google Scholar] [CrossRef]
- Horcada, A.; Campo, M.D.M.; Polvillo, O.; Alcalde, M.J.; Cilla, I.; Sañudo, C. A comparative study of fatty acid profiles of fat in commercial Spanish suckling kids and lambs. Span. J. Agric. Res. 2014, 12, 427. [Google Scholar] [CrossRef]
- Adeyemi, K.D.; Shittu, R.M.; Sabow, A.B.; Karim, R.; Sazili, A.Q. Myofibrillar protein, lipid and myoglobin oxidation, antioxidant profile, physicochemical and sensory properties of caprine longissimus thoracis during postmortem conditioning. J. Food Process. Preserv. 2017, 41, e13076. [Google Scholar] [CrossRef]
- Binkoski, A.E.; Kris-Etherton, P.M.; Wilson, T.A.; Mountain, M.L.; Nicolosi, R.J. Balance of unsaturated fatty acids is important to a cholesterol-lowering diet: Comparison of mid-oleic sunflower oil and olive oil on cardiovascular disease risk factors. J. Am. Diet. Assoc. 2005, 105, 1080–1086. [Google Scholar] [CrossRef] [PubMed]
- Pariza, M.W.; Park, Y.; Cook, M.E. The biologically active isomers of conjugated linoleic acid. Prog. Lipid Res. 2001, 40, 283–298. [Google Scholar] [CrossRef]
- Marventano, S.; Kolacz, P.; Castellano, S.; Galvano, F.; Buscemi, S.; Mistretta, A.; Grosso, G. A review of recent evidence in human studies of n–3 and n–6 PUFA intake on cardiovascular disease, cancer, and depressive disorders: Does the ratio really matter? Int. J. Food Sci. Nutr. 2015, 66, 611–622. [Google Scholar] [CrossRef] [PubMed]
- Scollan, N.; Hocquette, J.F.; Nuernberg, K.; Dannenberger, D.; Richardson, I.; Moloney, A. Innovations in beef production systems that enhance the nutritional and health value of beef lipids and their relationship with meat quality. Meat Sci. 2006, 74, 17–33. [Google Scholar] [CrossRef]
- Manso, T.; Bodas, R.; Vieira, C.; Mantecon, A.R.; Castro, T. Feeding vegetable oils to lactating ewes modifies the fatty acid profile of suckling lambs. Animal 2011, 5, 1659–1667. [Google Scholar] [CrossRef]
- Bas, P.; Morand-Fehr, P. Effect of nutritional factors on fatty acid composition of lamb fat deposits. Livest. Prod. Sci. 2000, 64, 61–79. [Google Scholar] [CrossRef]
- Bañón, S.; Vila, R.; Price, A.; Ferrandini, E.; Garrido, M.D. Effects of goat milk or milk replacer diet on meat quality and fat composition of suckling goat kids. Meat Sci. 2006, 72, 216–221. [Google Scholar] [CrossRef] [PubMed]
- Tsiplakou, E.; Papadomichelakis, G.; Sparaggis, D.; Sotirakoglou, K.; Georgiadou, M.; Zervas, G. The effect of maternal or artificial milk, age and sex on three muscles fatty acid profile of Damascus breed goat kids. Livest. Sci. 2016, 188, 142–152. [Google Scholar] [CrossRef]
- De Palo, P.; Maggiolino, A.; Centoducati, N.; Tateo, A. Effects of different milk replacers on carcass traits, meat quality, meat color and fatty acids profile of dairy goat kids. Small Rumin. Res. 2015, 131, 6–11. [Google Scholar] [CrossRef]
- Joy, M.; Ripoll, G.; Molino, F.; Dervishi, E.; Alvarez-Rodriguez, J. Influence of the type of forage supplied to ewes in pre- and post-partum periods on the meat fatty acids of suckling lambs. Meat Sci. 2012, 90, 775–782. [Google Scholar] [CrossRef] [PubMed]
- Nudda, A.; Palmquist, D.L.; Battacone, G.; Fancellu, S.; Rassu, S.P.G.; Pulina, G. Relationships between the contents of vaccenic acid, CLA and n–3 fatty acids of goat milk and the muscle of their suckling kids. Livest. Sci. 2008, 118, 195–203. [Google Scholar] [CrossRef]
- Sanz-Sampelayo, M.; Fernández, J.; Ramos, E.; Hermoso, R.; Extremera, F.G.; Boza, J. Effect of providing a polyunsaturated fatty acid-rich protected fat to lactating goats on growth and body composition of suckling goat kids. Anim. Sci. 2006, 82, 337–344. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Time (T) | 1 d of Lactation | 30 d of Lactation | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Breed (B) | GU † | PL | RE | TI | GU | PL | RE | TI | s.e. | B | T | B × T |
| C4:0 | 1.48c | 1.06d | 1.72b | 1.27cd | 2.03a | 1.62bc | 1.77ab | 2.02a | 0.103 | 0.005 | 0.0001 | 0.002 |
| C6:0 | 1.44d | 1.01e | 1.79c | 1.21de | 2.37ab | 2.22ab | 2.20b | 2.54a | 0.110 | 0.042 | 0.0001 | 0.0001 |
| C7:0 | 0.012bc | 0.010bc | 0.012bc | 0.005c | 0.051a | 0.048a | 0.023b | 0.062a | 0.006 | 0.018 | 0.0001 | 0.001 |
| C8:0 | 1.57c | 1.08d | 2.07b | 1.18d | 2.85a | 2.94a | 2.78a | 2.86a | 0.135 | 0.005 | 0.0001 | 0.0001 |
| C9:0 | 0.035cd | 0.031cd | 0.040cd | 0.025d | 0.112b | 0.130ab | 0.068c | 0.146a | 0.012 | 0.031 | 0.0001 | 0.0001 |
| C10:0 | 4.76d | 3.42d | 6.35c | 3.51d | 8.75b | 11.18a | 9.53ab | 10.13a | 0.483 | 0.020 | 0.0001 | 0.0001 |
| C11:0 | 0.125e | 0.091e | 0.176d | 0.080e | 0.273bc | 0.376a | 0.252c | 0.314ab | 0.020 | 0.40 | 0.0001 | 0.0001 |
| C12:0 | 2.48de | 1.85ef | 2.92d | 1.73f | 3.52c | 4.98a | 4.18ab | 4.00bc | 0.241 | 0.007 | 0.0001 | 0.0001 |
| C13:0 | 0.087c | 0.059cd | 0.077c | 0.041d | 0.153ab | 0.193a | 0.131b | 0.157ab | 0.013 | 0.12 | 0.0001 | 0.007 |
| C14:0 | 11.29ab | 9.90ab | 11.84a | 9.50b | 10.06ab | 12.52a | 11.76a | 12.07a | 0.694 | 0.23 | 0.059 | 0.007 |
| C15:0 | 0.593b | 0.530b | 0.618b | 0.521b | 0.635b | 0.945a | 0.818a | 0.954a | 0.049 | 0.012 | 0.0001 | 0.0001 |
| C16:0 | 24.37b | 27.88a | 24.47b | 26.86a | 19.62c | 27.83a | 23.85b | 28.24a | 0.718 | 0.0001 | 0.057 | 0.0001 |
| C17:0 | 0.804bcd | 0.955ab | 0.879abc | 0.965a | 0.540e | 0.746cd | 0.708d | 0.698d | 0.047 | 0.0001 | 0.0001 | 0.60 |
| C18:0 | 13.30b | 12.31bcd | 10.28de | 12.91bc | 17.64a | 9.25e | 12.29bcd | 10.87cde | 0.663 | 0.0001 | 0.52 | 0.0001 |
| C20:0 | 0.212b | 0.190bcd | 0.171cd | 0.221b | 0.338a | 0.150cd | 0.191bc | 0.140d | 0.015 | 0.0001 | 0.56 | 0.0001 |
| C22:0 | 0.050bc | 0.047bcd | 0.048bcd | 0.057bc | 0.074a | 0.037cd | 0.060b | 0.034d | 0.005 | 0.001 | 0.83 | 0.0001 |
| C23:0 | 0.006c | 0.008c | 0.015b | 0.017b | 0.006c | 0.008c | 0.022a | 0.014b | 0.002 | 0.0001 | 0.47 | 0.002 |
| Time (T) | 1 d of Lactation | 30 d of Lactation | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Breed (B) | GU † | PL | RE | TI | GU | PL | RE | TI | s.e. | B | T | B × T |
| C12:1 | 0.050 | 0.032 | 0.034 | 0.023 | 0.037 | 0.050 | 0.047 | 0.035 | 0.015 | 0.47 | 0.31 | 0.38 |
| C14:1c9 | 0.078bc | 0.066bcd | 0.103a | 0.062cd | 0.055d | 0.084abc | 0.084ab | 0.063cd | 0.007 | 0.0001 | 0.242 | 0.025 |
| C15:1 | 0.011 | 0.004 | 0.005 | 0.005 | 0.012 | 0.005 | 0.005 | 0.005a | 0.005 | 0.33 | 0.942 | 0.99 |
| C16:1t9 | 0.361cd | 0.445ab | 0.487a | 0.444ab | 0.272e | 0.377cd | 0.404bc | 0.336d | 0.049 | 0.0001 | 0.0001 | 0.79 |
| C16:1c7 | 0.338abc | 0.358ab | 0.344abc | 0.385a | 0.293bcd | 0.264d | 0.274cd | 0.237d | 0.022 | 0.99 | 0.0001 | 0.061 |
| C16:1c9 | 1.185b | 1.487a | 1.495a | 1.588a | 0.727d | 1.070bc | 1.056bc | 0.965c | 0.063 | 0.0001 | 0.0001 | 0.323 |
| C17:1c9 | 0.337b | 0.353b | 0.537a | 0.512a | 0.142d | 0.205cd | 0.290bc | 0.252bcd | 0.037 | 0.0001 | 0.0001 | 0.50 |
| C18:1t11 | 0.839c | 1.257a | 0.683c | 0.717c | 1.157ab | 0.674c | 0.817c | 0.901bc | 0.104 | 0.024 | 0.86 | 0.004 |
| C18:1c9 | 27.217b | 29.100ab | 27.559ab | 30.274a | 22.953c | 17.745de | 21.396cd | 17.713e | 1.186 | 0.58 | 0.0001 | 0.0001 |
| C18:1t15 | 0.125bc | 0.120bc | 0.128bc | 0.121bc | 0.156ab | 0.089c | 0.171a | 0.116c | 0.011 | 0.000 | 0.22 | 0.007 |
| C18:1c11 | 0.173b | 0.123bc | 0.117c | 0.114c | 0.239a | 0.142bc | 0.163b | 0.136bc | 0.015 | 0.0001 | 0.001 | 0.32 |
| C18:1c12 | 0.139ab | 0.139ab | 0.074c | 0.082c | 0.159a | 0.105bc | 0.121b | 0.079c | 0.010 | 0.0001 | 0.33 | 0.005 |
| C18:1c13 | 0.047 | 0.055 | 0.037 | 0.041 | 0.047 | 0.046 | 0.057 | 0.039 | 0.006 | 0.41 | 0.631 | 0.11 |
| C18:1t16 | 0.146ab | 0.139ab | 0.134b | 0.130b | 0.153ab | 0.116bc | 0.168a | 0.100c | 0.011 | 0.001 | 0.69 | 0.010 |
| C18:1c15 | 0.066ab | 0.058ab | 0.060ab | 0.059ab | 0.059ab | 0.048b | 0.070a | 0.050b | 0.005 | 0.038 | 0.23 | 0.12 |
| C20:1n–9 | 0.007 | 0.002 | 0.002 | 0.001 | 0.001 | 0.001 | 0.002 | 0.001a | 0.003 | 0.49 | 0.30 | 0.47 |
| C22:1 | 0.012d | 0.015cd | 0.022ab | 0.026a | 0.003e | 0.007e | 0.019bc | 0.015cd | 0.002 | 0.0001 | 0.0001 | 0.065 |
| Time (T) | 1 d of Lactation | 30 d of Lactation | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Breed (B) | GU † | PL | RE | TI | GU | PL | RE | TI | s.e. | B | T | B × T |
| C18:2 n–6 t9,12 | 0.122abc | 0.111bcd | 0.107bcd | 0.097cd | 0.130ab | 0.087d | 0.149a | 0.083d | 0.009 | 0.0001 | 0.67 | 0.002 |
| C18:2 n–6 | 3.37a | 3.31a | 2.13c | 2.77b | 2.62b | 1.88cd | 1.78d | 2.00cd | 0.120 | 0.0001 | 0.0001 | 0.003 |
| C18:2 c9, t11 | 0.221cd | 0.464a | 0.307b | 0.254bc | 0.176d | 0.248bcd | 0.293bc | 0.236bcd | 0.027 | 0.0001 | 0.0001 | 0.015 |
| C18:2 t9, c11 | 0.042b | 0.052ab | 0.051ab | 0.047b | 0.041b | 0.048b | 0.059a | 0.045b | 0.003 | 0.0001 | 0.94 | 0.20 |
| C18:2 t10, c12 | 0.019ab | 0.022ab | 0.019ab | 0.019ab | 0.017ab | 0.013b | 0.023a | 0.017ab | 0.002 | 0.35 | 0.17 | 0.065 |
| C18:3 n–6 | 0.031a | 0.020ab | 0.012b | 0.021ab | 0.009b | 0.012ab | 0.005b | 0.012b | 0.005 | 0.11 | 0.005 | 0.30 |
| C18:3 n–3 | 0.418a | 0.245bcd | 0.350ab | 0.265bc | 0.383a | 0.223cd | 0.420a | 0.171d | 0.031 | 0.0001 | 0.365 | 0.030 |
| C20:2 n–6 | 0.027b | 0.041a | 0.018c | 0.029b | 0.010d | 0.013cd | 0.012d | 0.011d | 0.002 | 0.0001 | 0.0001 | 0.0001 |
| C20:3 n–9 | 0.032b | 0.036ab | 0.026cd | 0.042a | 0.014f | 0.018ef | 0.021de | 0.028bc | 0.002 | 0.0001 | 0.0001 | 0.016 |
| C20:3 n–6 | 0.001c | 0.002c | 0.006b | 0.009a | 0.001c | 0.003c | 0.008ab | 0.008ab | 0.001 | 0.0001 | 0.63 | 0.43 |
| C20:4 n–6 | 0.521a | 0.483a | 0.356b | 0.526a | 0.164c | 0.190c | 0.170c | 0.215c | 0.029 | 0.001 | 0.0001 | 0.008 |
| C20:5 n–3 | 0.096a | 0.071b | 0.075b | 0.078b | 0.048c | 0.041c | 0.066b | 0.036c | 0.006 | 0.006 | 0.0001 | 0.001 |
| C22:3 n–3 | 0.013b | 0.018ab | 0.015b | 0.020a | 0.004d | 0.012bc | 0.022a | 0.006cd | 0.002 | 0.0001 | 0.0001 | 0.0001 |
| C22:4 n–6 | 0.084a | 0.109a | 0.045b | 0.089a | 0.028bc | 0.026bc | 0.018c | 0.025c | 0.008 | 0.0001 | 0.0001 | 0.007 |
| C22:5 n–3 | 0.314a | 0.158c | 0.201bc | 0.227b | 0.074d | 0.051d | 0.088d | 0.053d | 0.017 | 0.0001 | 0.0001 | 0.0001 |
| C22:6 n–3 | 0.054a | 0.016c | 0.033b | 0.015c | 0.014c | 0.005c | 0.018c | 0.006c | 0.004 | 0.0001 | 0.0001 | 0.0001 |
| Time (T) | 1 d of Lactation | 30 d of Lactation | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Breed (B) | GU † | PL | RE | TI | GU | PL | RE | TI | s.e. | B | T | B × T |
| SFA | 63.48c | 61.03c | 64.37c | 60.85c | 69.79b | 76.07a | 71.64b | 75.96a | 1.384 | 0.40 | 0.0001 | 0.0001 |
| ƩBCFA | 0.678b | 0.475c | 0.730ab | 0.564c | 0.569c | 0.696ab | 0.814a | 0.575c | 0.035 | 0.0001 | 0.046 | 0.0001 |
| Ʃiso-BCFA | 0.404a | 0.238c | 0.423a | 0.320b | 0.311b | 0.305bc | 0.407a | 0.279bc | 0.018 | 0.0001 | 0.120 | 0.002 |
| Ʃanteiso-BCFA | 0.274b | 0.237b | 0.307b | 0.244b | 0.258b | 0.391a | 0.407a | 0.296b | 0.021 | 0.0001 | 0.0001 | 0.001 |
| MUFA | 31.13b | 33.75ab | 31.82ab | 34.58a | 26.47c | 21.03d | 25.14cd | 21.04d | 1.292 | 0.72 | 0.0001 | 0.0001 |
| Ʃcis-MUFA | 29.66b | 31.79ab | 30.39ab | 33.17a | 24.73c | 19.77de | 23.58cd | 19.59e | 1.269 | 0.74 | 0.0001 | 0.001 |
| Ʃtrans-MUFA | 1.47bc | 1.96a | 1.43bc | 1.41bc | 1.74ab | 1.26c | 1.56abc | 1.45bc | 0.122 | 0.34 | 0.45 | 0.009 |
| ƩCLA | 0.283de | 0.538a | 0.377b | 0.320bcd | 0.235e | 0.309bcde | 0.376bc | 0.298cde | 0.030 | 0.0001 | 0.001 | 0.013 |
| PUFA | 5.36a | 5.16a | 3.75c | 4.51b | 3.73c | 2.86d | 3.15d | 2.95d | 0.175 | 0.0001 | 0.0001 | 0.000 |
| n–6 | 4.15a | 4.080a | 2.68cd | 3.54b | 2.96c | 2.21de | 2.14e | 2.35de | 0.139 | 0.0001 | 0.0001 | 0.001 |
| n–3 | 0.895a | 0.508cd | 0.674b | 0.605bc | 0.523c | 0.331de | 0.613bc | 0.273e | 0.044 | 0.0001 | 0.0001 | 0.0001 |
| n–6:n–3 | 4.78c | 8.14a | 4.08cd | 5.95b | 5.80b | 6.69b | 3.80d | 8.95a | 0.360 | 0.0001 | 0.031 | 0.0001 |
| RS † | Milk Replacer | Natural Milk | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Breed (B) | GU | PL | RE | TI | GU | PL | RE | TI | s.e. | B | RS | B × RS |
| C6:0 | 0.002a | 0.005a | 0.002a | 0.002a | 0.002a | 0.003a | 0.004a | 0.003a | 0.001 | 0.49 | 0.72 | 0.17 |
| C8:0 | 0.004b | 0.011a | 0.005ab | 0.007ab | 0.006ab | 0.007ab | 0.006ab | 0.009ab | 0.002 | 0.401 | 0.83 | 0.30 |
| C9:0 | 0.013b | 0.077a | 0.009b | 0.001b | 0.030ab | 0.048a | 0.025ab | 0.049a | 0.012 | 0.008 | 0.16 | 0.008 |
| C10:0 | 0.081d | 0.154cd | 0.142cd | 0.196cd | 0.239c | 0.511a | 0.274bc | 0.407ab | 0.048 | 0.034 | 0.0001 | 0.10 |
| C11:0 | 0.018c | 0.066ab | 0.012c | 0.011c | 0.045abc | 0.073ab | 0.036bc | 0.080a | 0.012 | 0.07 | 0.001 | 0.036 |
| C12:0 | 3.14a | 4.25a | 0.36c | 3.75a | 0.79bc | 1.12bc | 0.90bc | 1.56b | 0.360 | 0.0001 | 0.0001 | 0.0001 |
| C13:0 | 0.046de | 0.091abc | 0.021e | 0.065cd | 0.067bcd | 0.093ab | 0.054cd | 0.107a | 0.010 | 0.0001 | 0.003 | 0.16 |
| C14:0 | 12.18a | 10.78abc | 7.66d | 11.65ab | 8.98cd | 9.26cd | 9.79bc | 8.61cd | 0.641 | 0.013 | 0.006 | 0.0001 |
| C15:0 | 0.246c | 0.403bc | 0.199c | 0.452b | 0.425b | 0.634a | 0.455b | 0.715a | 0.054 | 0.0001 | 0.0001 | 0.78 |
| C16:0 | 22.60cd | 25.12b | 21.12d | 24.73b | 21.21d | 27.49a | 23.99bc | 27.64a | 0.697 | 0.0001 | 0.003 | 0.001 |
| C17:0 | 0.448d | 0.562bcd | 0.530cd | 0.691bc | 0.693bc | 1.021a | 0.794b | 0.977a | 0.062 | 0.002 | 0.0001 | 0.30 |
| C18:0 | 10.36c | 9.48c | 13.15ab | 9.79c | 14.07a | 13.57a | 13.09ab | 11.875b | 0.640 | 0.017 | 0.0001 | 0.003 |
| C20:0 | 0.062ab | 0.050b | 0.066ab | 0.061ab | 0.081a | 0.063ab | 0.071ab | 0.067ab | 0.006 | 0.34 | 0.028 | 0.54 |
| C22:0 | 0.056a | 0.018b | 0.020b | 0.011b | 0.010b | 0.018b | 0.023b | 0.014b | 0.007 | 0.035 | 0.07 | 0.000 |
| C23:0 | 0.205a | 0.072bc | 0.057c | 0.043c | 0.173ab | 0.049c | 0.098bc | 0.047c | 0.023 | 0.0001 | 0.90 | 0.31 |
| RS † | Milk Replacer | Natural Milk | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Breed (B) | GU | PL | RE | TI | GU | PL | RE | TI | s.e. | B | RS | B × RS |
| C14:1c9 | 0.229bc | 0.266ab | 0.063g | 0.276a | 0.107f | 0.175de | 0.143e | 0.200cd | 0.015 | 0.0001 | 0.0001 | 0.0001 |
| C15:1 | 0.002b | 0.013a | 0.001b | 0.003b | 0.003b | 0.003b | 0.004b | 0.004b | 0.002 | 0.14 | 0.36 | 0.009 |
| C16:1c7 | 1.92a | 0.28b | 0.85b | 0.34b | 1.40ab | 0.31b | 0.82b | 0.31b | 0.317 | 0.001 | 0.59 | 0.71 |
| C16:1c9 | 2.86a | 2.37ab | 2.55a | 2.64a | 2.10b | 2.42ab | 2.58a | 2.65a | 0.132 | 0.18 | 0.1 | 0.001 |
| C17:1c9 | 0.147e | 0.324cde | 0.238de | 0.425bc | 0.182e | 0.537ab | 0.379bcd | 0.626a | 0.054 | 0.0001 | 0.001 | 0.22 |
| C18:1c11 | 0.079d | 0.125bcd | 0.088cd | 0.127bc | 0.191ab | 0.192a | 0.150ab | 0.135b | 0.014 | 0.12 | 0.0001 | 0.0001 |
| C18:1c12 | 0.042d | 0.097cd | 0.069d | 0.162b | 0.145bc | 0.228a | 0.154bc | 0.164b | 0.018 | 0.005 | 0.0001 | 0.0001 |
| C18:1c13 | 0.010c | 0.035b | 0.021bc | 0.040b | 0.026bc | 0.065a | 0.036b | 0.035b | 0.006 | 0.001 | 0.004 | 0.017 |
| C18:1c15 | 0.026c | 0.069ab | 0.016c | 0.075ab | 0.032c | 0.080a | 0.052b | 0.053b | 0.007 | 0.0001 | 0.86 | 0.0001 |
| C18:1t11 | 0.349c | 0.680abc | 0.303c | 0.929ab | 0.651abc | 1.207a | 0.592bc | 1.049ab | 0.169 | 0.031 | 0.022 | 0.63 |
| C18:1t15 | 0.131a | 0.137a | 0.134a | 0.141a | 0.173a | 0.159a | 0.139a | 0.141a | 0.014 | 0.55 | 0.12 | 0.26 |
| C18:1t16 | 0.018c | 0.079bc | 0.035c | 0.120b | 0.134b | 0.216a | 0.125b | 0.136b | 0.021 | 0.054 | 0.0001 | 0.006 |
| C18:1c9 | 30.51c | 33.78b | 41.53a | 35.65b | 36.30b | 33.61b | 36.477b | 35.661b | 1.088 | 0.0001 | 0.86 | 0.0001 |
| C20:1n–9 | 0.002a | 0.003a | 0.002a | 0.002a | 0.004a | 0.001a | 0.003a | 0.004a | 0.001 | 0.81 | 0.71 | 0.32 |
| C22:1 | 0.010 a | 0.003 b | 0.010 ab | 0.006 b | 0.010 ab | 0.004 b | 0.007 ab | 0.004 b | 0.002 | 0.010 | 0.44 | 0.71 |
| RS † | Milk Replacer | Natural Milk | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Breed (B) | GU | PL | RE | TI | GU | PL | RE | TI | s.e. | B | RS | B × RS |
| C18:2c9,t11 | 0.174cd | 0.264bcd | 0.097d | 0.343b | 0.159d | 0.500a | 0.270bc | 0.353b | 0.046 | 0.002 | 0.006 | 0.007 |
| C18:2 n–6 | 7.275a | 6.048ab | 7.428a | 4.465b | 6.339a | 2.765c | 4.283bc | 3.188c | 0.468 | 0.0001 | 0.0001 | 0.007 |
| C18:2n–6t9,12 | 0.024c | 0.088b | 0.023c | 0.118b | 0.110b | 0.173a | 0.108b | 0.115b | 0.016 | 0.010 | 0.0001 | 0.001 |
| C18:2t10,c12 | 0.027a | 0.026ab | 0.017b | 0.025ab | 0.016b | 0.021ab | 0.021ab | 0.027a | 0.003 | 0.177 | 0.218 | 0.005 |
| C18:2t9,c11 | 0.042a | 0.014b | 0.023b | 0.014b | 0.026b | 0.011b | 0.016b | 0.014b | 0.004 | 0.0001 | 0.077 | 0.152 |
| C18:3 n–3 | 0.372c | 0.230d | 0.764a | 0.227d | 0.435c | 0.191d | 0.529b | 0.193d | 0.033 | 0.0001 | 0.019 | 0.0001 |
| C18:3 n–6 | 0.035a | 0.023bc | 0.018bc | 0.025b | 0.024bc | 0.020bc | 0.014c | 0.023bc | 0.004 | 0.000 | 0.087 | 0.494 |
| C20:2 n–6 | 0.125b | 0.062c | 0.182a | 0.020d | 0.073c | 0.022d | 0.059c | 0.021d | 0.009 | 0.0001 | 0.0001 | 0.0001 |
| C20:3 n–6 | 0.003abc | 0.002bcd | 0.005a | 0.001d | 0.004ab | 0.001d | 0.003abcd | 0.002cd | 0.001 | 0.033 | 0.447 | 0.303 |
| C20:3 n–9 | 0.252a | 0.163b | 0.093bcd | 0.074d | 0.151bc | 0.078cd | 0.083bcd | 0.073d | 0.022 | 0.0001 | 0.005 | 0.013 |
| C20:4 n–6 | 3.23a | 1.95bc | 0.93c | 0.87c | 2.38b | 1.15c | 1.25c | 0.94c | 0.321 | 0.0001 | 0.215 | 0.097 |
| C20:5 n–3 | 0.234a | 0.140ab | 0.090b | 0.078b | 0.169ab | 0.107b | 0.179ab | 0.091b | 0.029 | 0.001 | 0.954 | 0.020 |
| C22:3 n–3 | 0.161a | 0.0001b | 0.036b | 0.0001b | 0.075b | 0.0001b | 0.044b | 0.001b | 0.025 | 0.0001 | 0.386 | 0.071 |
| C22:4 n–6 | 0.463a | 0.264b | 0.149b | 0.167b | 0.227b | 0.178b | 0.135b | 0.171b | 0.042 | 0.0001 | 0.013 | 0.003 |
| C22:5 n–3 | 0.598a | 0.336bc | 0.235c | 0.197c | 0.527ab | 0.293c | 0.410bc | 0.252c | 0.061 | 0.0001 | 0.543 | 0.117 |
| C22:6 n–3 | 0.093a | 0.045bc | 0.044c | 0.029c | 0.109a | 0.040c | 0.091ab | 0.037c | 0.011 | 0.0001 | 0.072 | 0.127 |
| RS † | Milk Replacer | Natural Milk | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Breed (B) | GU | PL | RE | TI | GU | PL | RE | TI | s.e. | B | RS | B × RS |
| ƩSFA | 49.87bc | 51.62ab | 43.68d | 51.96ab | 47.34c | 54.58a | 50.21b | 52.69ab | 1.216 | 0.0001 | 0.047 | 0.001 |
| ƩBCFA | 0.41c | 0.46bc | 0.31d | 0.49abc | 0.48abc | 0.60a | 0.58ab | 0.51ab | 0.035 | 0.30 | 0.0001 | 0.001 |
| Ʃiso-BCFA | 0.23cd | 0.27bc | 0.21d | 0.31b | 0.31b | 0.39a | 0.39a | 0.33ab | 0.024 | 0.22 | 0.0001 | 0.004 |
| Ʃanteiso-BCFA | 0.18a | 0.19a | 0.11b | 0.18a | 0.17a | 0.21a | 0.19a | 0.18a | 0.015 | 0.073 | 0.06 | 0.001 |
| ƩMUFA | 36.48c | 38.49bc | 46.03a | 41.20b | 41.66b | 39.65b | 42.01b | 41.57b | 1.126 | 0.0001 | 0.45 | 0.0001 |
| Ʃcis-MUFA | 35.84c | 37.36bc | 45.43a | 39.74b | 40.50b | 37.63bc | 40.80b | 39.85b | 1.131 | 0.0001 | 0.90 | 0.000 |
| Ʃtrans-MUFA | 0.65c | 1.13bc | 0.59c | 1.47b | 1.16bc | 2.02a | 1.21bc | 1.72ab | 0.212 | 0.017 | 0.001 | 0.42 |
| ƩCLA | 0.24cd | 0.30bcd | 0.14d | 0.38b | 0.20cd | 0.53a | 0.31bc | 0.39b | 0.047 | 0.005 | 0.014 | 0.005 |
| ƩPUFA | 13.11a | 9.64bc | 10.13b | 6.66d | 10.83b | 5.55d | 7.94cd | 5.50d | 0.844 | 0.0001 | 0.0001 | 0.31 |
| n–6 | 11.16a | 8.43bc | 8.73b | 5.67d | 9.16b | 4.31d | 5.85cd | 4.46d | 0.744 | 0.0001 | 0.0001 | 0.18 |
| n–3 | 1.46a | 0.74b | 1.17a | 0.53b | 1.32a | 0.63b | 1.25a | 0.57b | 0.117 | 0.0001 | 0.73 | 0.62 |
| n–6:n–3 | 7.88b | 13.00a | 7.46b | 12.08a | 6.88b | 7.82b | 4.78c | 8.88b | 0.708 | 0.0001 | 0.0001 | 0.02 |
| Desirable FA | 59.97cd | 58.51cd | 69.24a | 58.20d | 65.62b | 59.28cd | 61.80c | 59.28cd | 0.970 | 0.0001 | 0.98 | 0.0001 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ripoll, G.; Alcalde, M.J.; Argüello, A.; Córdoba, M.d.G.; Panea, B. Effect of Rearing System on the Straight and Branched Fatty Acids of Goat Milk and Meat of Suckling Kids. Foods 2020, 9, 471. https://doi.org/10.3390/foods9040471
Ripoll G, Alcalde MJ, Argüello A, Córdoba MdG, Panea B. Effect of Rearing System on the Straight and Branched Fatty Acids of Goat Milk and Meat of Suckling Kids. Foods. 2020; 9(4):471. https://doi.org/10.3390/foods9040471
Chicago/Turabian StyleRipoll, Guillermo, María Jesús Alcalde, Anastasio Argüello, María de Guía Córdoba, and Begoña Panea. 2020. "Effect of Rearing System on the Straight and Branched Fatty Acids of Goat Milk and Meat of Suckling Kids" Foods 9, no. 4: 471. https://doi.org/10.3390/foods9040471
APA StyleRipoll, G., Alcalde, M. J., Argüello, A., Córdoba, M. d. G., & Panea, B. (2020). Effect of Rearing System on the Straight and Branched Fatty Acids of Goat Milk and Meat of Suckling Kids. Foods, 9(4), 471. https://doi.org/10.3390/foods9040471