Bifidobacterium longum subsp. infantis ATCC 15697 and Goat Milk Oligosaccharides Show Synergism In Vitro as Anti-Infectives against Campylobacter jejuni



Abstract
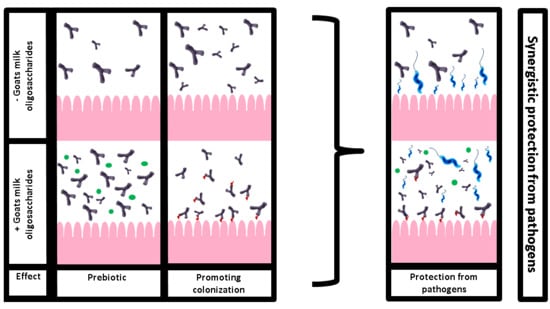
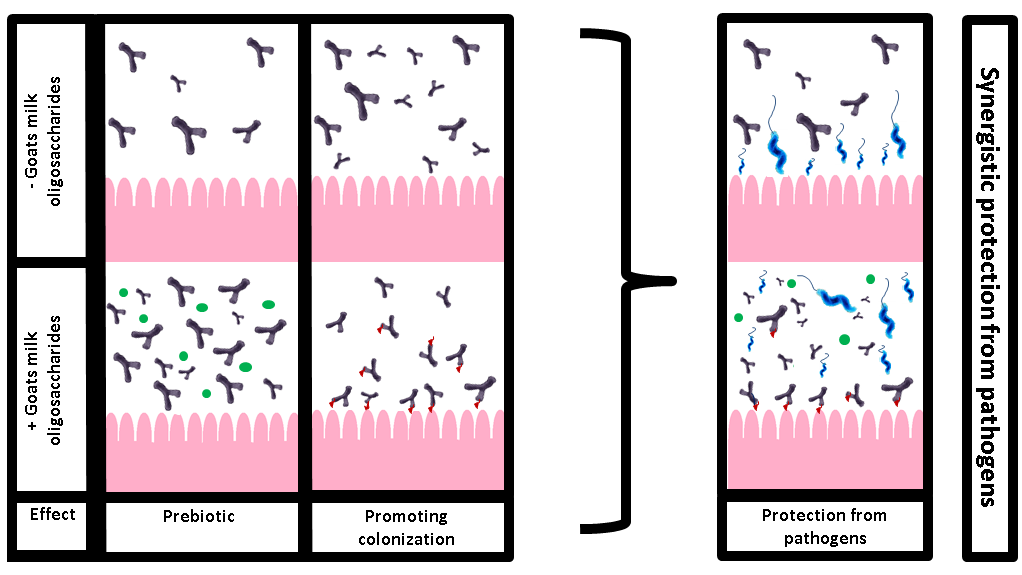
1. Introduction
2. Materials and Methods
2.1. Oligosaccharides Standards
2.2. Isolation of Goat Milk Oligosaccharides
2.3. Milk Oligosaccharide Analysis
2.4. Bifidobacterium longum subsp. infantis Culture Conditions
2.5. Campylobacter jejuni Culture Conditions
2.6. Exposure of B. infantis to Goat Milk Oligosaccharides
2.7. Mammalian Cell Culture Conditions
2.8. Adhesion Assays with B. infantis
2.9. Anti-Infective Assays and Exclusion Assay
2.10. Effect of GMO on the Growth of B. infantis
2.11. GMO Consumption by B. infantis
2.12. B. infantis Metabolite Analysis
2.13. Statistical Analysis
3. Results and Discussion
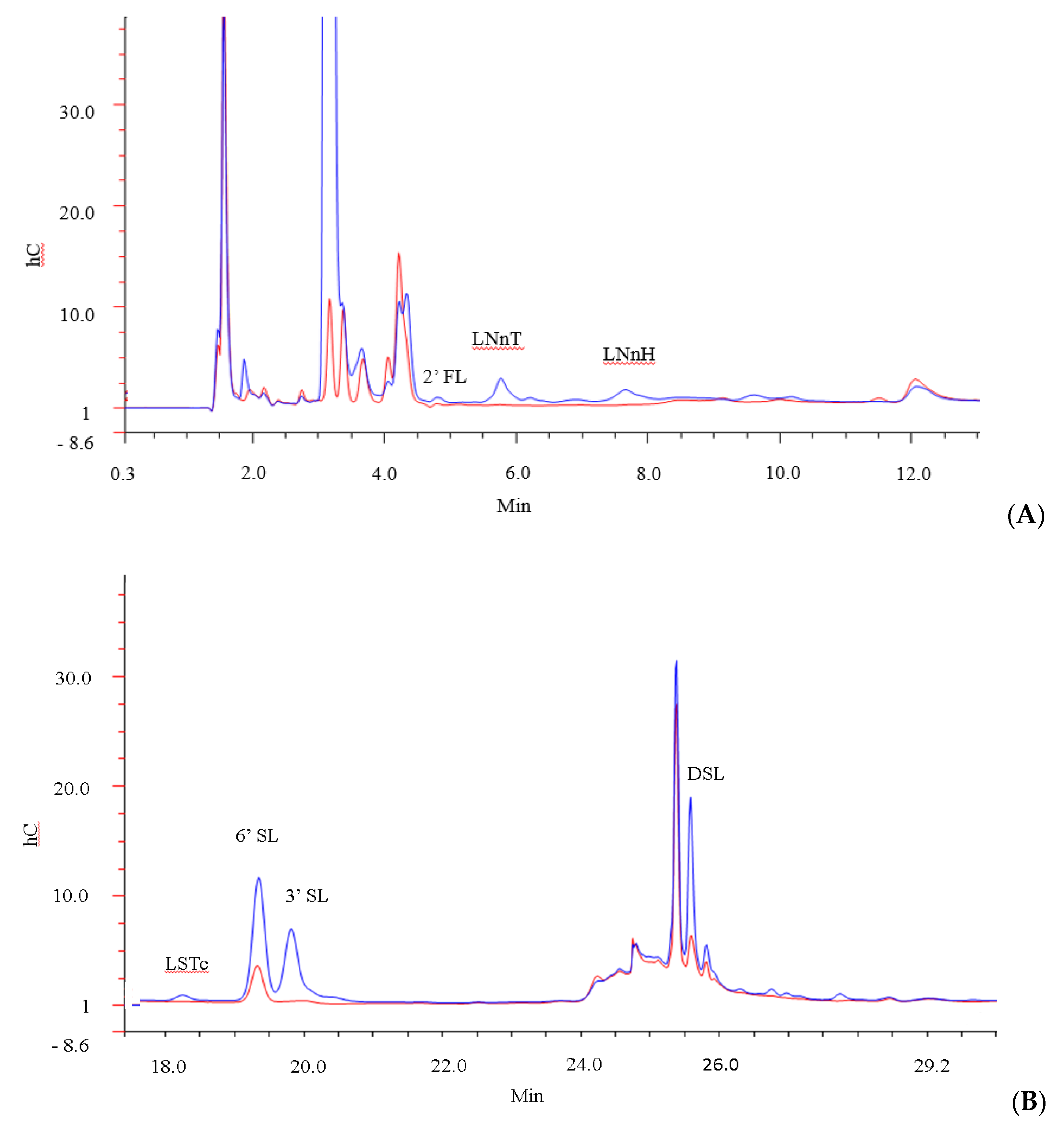
3.1. Characterisation of the Goat Milk Oligosaccharides
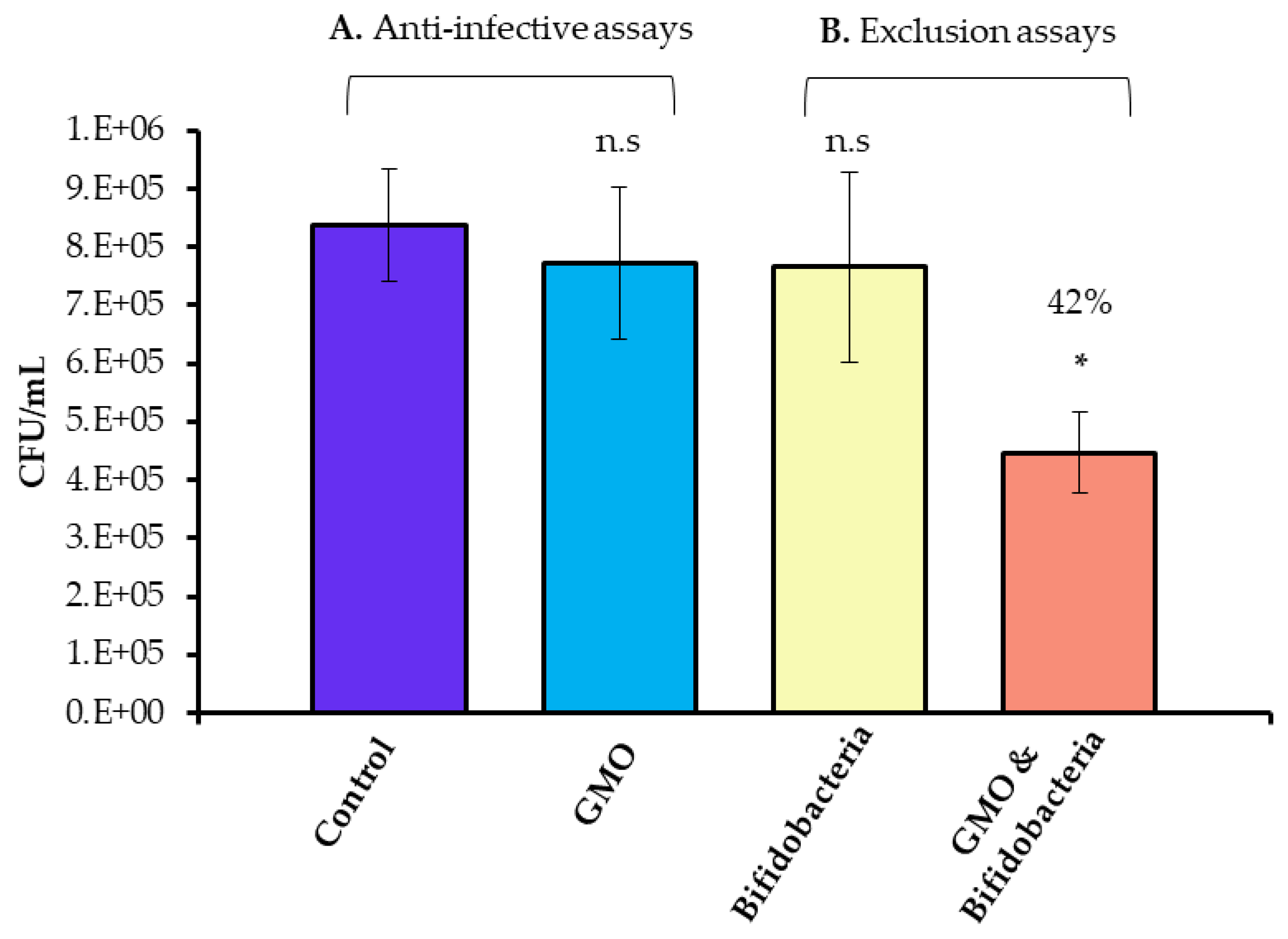
3.2. Combined Effect of GMO and B. infantis on C. jejuni Adhesion
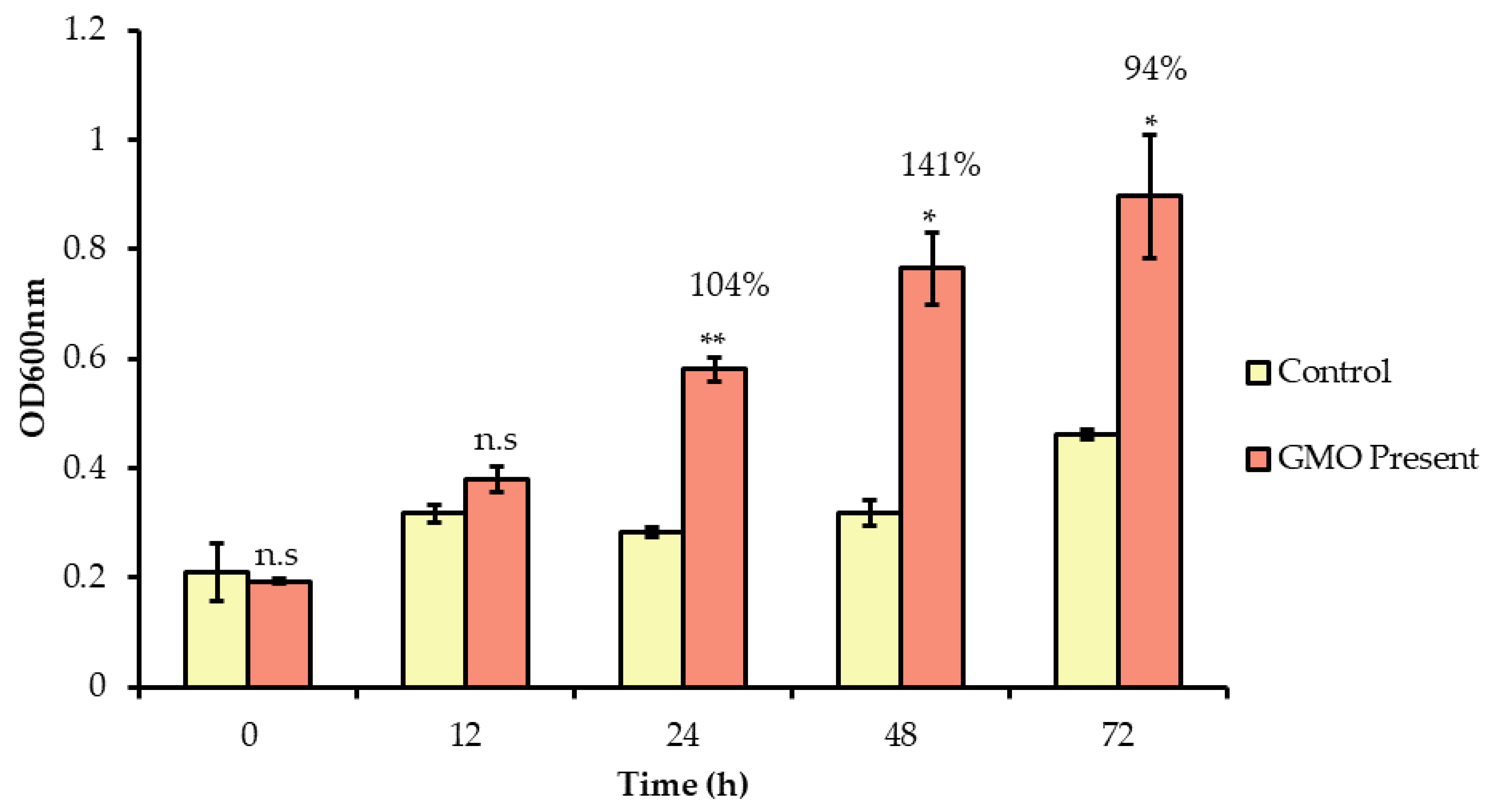
3.3. Prebiotic Effects of GMO
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- O’Callaghan, A.; Bottacini, F.; O’Connell Motherway, M.; van Sinderen, D. Pangenome Analysis of Bifidobacterium longum and Site-Directed Mutagenesis through by-Pass of Restriction-Modification Systems. BMC Genomics 2015, 16, 832. [Google Scholar] [CrossRef]
- Hidalgo-Cantabrana, C.; Delgado, S.; Ruiz, L.; Ruas-Madiedo, P.; Sánchez, B.; Margolles, A. Bifidobacteria and Their Health-Promoting Effects. Microbiol. Spectr. 2017, 5. [Google Scholar] [CrossRef]
- Gueimonde, M.; Margolles, A.G.; de los Reyes-Gavilán, C.; Salminen, S. Competitive Exclusion of Enteropathogens from Human Intestinal Mucus by Bifidobacterium Strains with Acquired Resistance to Bile—A Preliminary Study. Int. J. Food Microbiol. 2007, 113, 228–232. [Google Scholar] [CrossRef]
- Collado, M.C.; Gueimonde, M.; Sanz, Y.; Salminen, S. Adhesion Properties and Competitive Pathogen Exclusion Ability of Bifidobacteria with Acquired Acid Resistance. J. Food Prot. 2006, 69, 1675–1679. [Google Scholar] [CrossRef] [PubMed]
- Muñoz-Quezada, S.; Bermudez-Brito, M.; Chenoll, E.; Genovés, S.; Gomez-Llorente, C.; Plaza-Diaz, J.; Matencio, E.; José Bernal, M.; Romero, F.; Ramón, D.; et al. Competitive Inhibition of Three Novel Bacteria Isolated from Faeces of Breast Milk-Fed Infants against Selected Enteropathogens. Br. J. Nutr. 2013, 109, S63–S69. [Google Scholar] [CrossRef] [PubMed]
- Uraipan, S.; Hongpattarakere, T. Antagonistic Characteristics Against Food-Borne Pathogenic Bacteria of Lactic Acid Bacteria and Bifidobacteria Isolated from Feces of Healthy Thai Infants. Jundishapur J. Microbiol. 2015, 8, e18264. [Google Scholar] [CrossRef]
- Madsen, K.L. Enhancement of Epithelial Barrier Function by Probiotics. J. Epithel. Biol. Pharmacol. 2012, 5, 55–59. [Google Scholar] [CrossRef]
- Westermann, C.; Gleinser, M.; Corr, S.C.; Riedel, C.U. A Critical Evaluation of Bifidobacterial Adhesion to the Host Tissue. Front. Microbiol. 2016, 7, 1220. [Google Scholar] [CrossRef]
- Chichlowski, M.; De Lartigue, G.; German, J.B.; Raybould, H.E.; Mills, D.A. Bifidobacteria Isolated From Infants and Cultured on Human Milk Oligosaccharides Affect Intestinal Epithelial Function. J. Pediatr. Gastroenterol. Nutr. 2012, 55, 321–327. [Google Scholar] [CrossRef]
- Kavanaugh, D.W.; O’Callaghan, J.; Buttó, L.F.; Slattery, H.; Lane, J.; Clyne, M.; Kane, M.; Joshi, L.; Hickey, R.M. Exposure of Bifidobacterium longum subsp. Infantis to Milk Oligosaccharides Increases Adhesion to Epithelial Cells and Induces a Substantial Transcriptional Response. PLoS ONE 2013, 8, e67224. [Google Scholar] [CrossRef]
- Quinn, E.M.; Slattery, H.; Thompson, A.; Kilcoyne, M.; Joshi, L.; Hickey, R.M. Mining Milk for Factors Which Increase the Adherence of Bifidobacterium longum subsp. infantis to Intestinal Cells. Foods 2018, 7, 196. [Google Scholar] [CrossRef]
- Oliveira, D.L.; Costabile, A.; Wilbey, R.A.; Grandison, A.S.; Duarte, L.C.; Roseiro, L.B. In Vitro Evaluation of the Fermentation Properties and Potential Prebiotic Activity of Caprine Cheese Whey Oligosaccharides in Batch Culture Systems. BioFactors 2012, 38, 440–449. [Google Scholar] [CrossRef]
- Leong, A.; Liu, Z.; Almshawit, H.; Zisu, B.; Pillidge, C.; Rochfort, S.; Gill, H. Oligosaccharides in Goats’ Milk-Based Infant Formula and Their Prebiotic and Anti-Infection Properties. Br. J. Nutr. 2019, 122, 441–449. [Google Scholar] [CrossRef]
- Newburg, D.S.; Ruiz-Palacios, G.M.; Morrow, A.L. Human Milk Glycans Protect Infants against Enteric Pathogens. Annu. Rev. Nutr. 2005, 25, 37–58. [Google Scholar] [CrossRef]
- Thum, C.; McNabb, W.C.; Young, W.; Cookson, A.L.; Roy, N.C. Prenatal Caprine Milk Oligosaccharide Consumption Affects the Development of Mice Offspring. Mol. Nutr. Food Res. 2016, 60, 2076–2085. [Google Scholar] [CrossRef]
- Bradford, M.M. A Rapid and Sensitive Method for the Quantitation of Microgram Quantities of Protein Utilizing the Principle of Protein-Dye Binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Alemka, A.; Whelan, S.; Gough, R.; Clyne, M.; Gallagher, M.E.; Carrington, S.D.; Bourke, B. Purified Chicken Intestinal Mucin Attenuates Campylobacter jejuni Pathogenicity In Vitro. J. Med. Microbiol. 2010, 59, 898–903. [Google Scholar] [CrossRef]
- Lane, J.A.; Mariño, K.; Naughton, J.; Kavanaugh, D.; Clyne, M.; Carrington, S.D.; Hickey, R.M. Anti-Infective Bovine Colostrum Oligosaccharides: Campylobacter jejuni as a Case Study. Int. J. Food Microbiol. 2012, 157, 182–188. [Google Scholar] [CrossRef]
- McGuire, M.K.; Meehan, C.L.; McGuire, M.A.; Williams, J.E.; Foster, J.; Sellen, D.W.; Kamau-Mbuthia, E.W.; Kamundia, E.W.; Mbugua, S.; Moore, S.E.; et al. What’s Normal? Oligosaccharide Concentrations and Profiles in Milk Produced by Healthy Women Vary Geographically. Am. J. Clin. Nutr. 2017. [Google Scholar] [CrossRef]
- Coppa, G.V.; Pierani, P.; Zampini, L.; Carloni, I.; Carlucci, A.; Gabrielli, O. Oligosaccharides in Human Milk during Different Phases of Lactation. Acta Paediatr. 1999. [Google Scholar] [CrossRef]
- Kunz, C.; Rodriguez-Palmero, M.; Koletzko, B.; Jensen, R. Nutritional and Biochemical Properties of Human Milk, Part I: General Aspects, Proteins, and Carbohydrates. Clin. Perinatol. 1999, 26, 307–333. [Google Scholar] [CrossRef]
- Shen, Z.; Warren, C.D.; Newburg, D.S. High-Performance Capillary Electrophoresis of Sialylated Oligosaccharides of Human Milk. Anal. Biochem. 2000. [Google Scholar] [CrossRef]
- Chaturvedi, P.; Warren, C.D.; Altaye, M.; Morrow, A.L.; Ruiz-Palacios, G.; Pickering, L.K.; Newburg, D.S. Fucosylated Human Milk Oligosaccharides Vary between Individuals and over the Course of Lactation. Glycobiology 2001, 11, 365–372. [Google Scholar] [CrossRef]
- Thurl, S.; Munzert, M.; Henker, J.; Boehm, G.; Müller-Werner, B.; Jelinek, J.; Stahl, B. Variation of Human Milk Oligosaccharides in Relation to Milk Groups and Lactational Periods. Br. J. Nutr. 2010, 104, 1261–1271. [Google Scholar] [CrossRef]
- Davidson, B.; Meinzen-Derr, J.K.; Wagner, C.L.; Newburg, D.S.; Morrow, A.L. Fucolsylated Oligosaccharides in Human Milk in Relation to Gestational Age and Stage of Lactation. Adv. Ecp Med. Biol. 2004, 554, 427–430. [Google Scholar] [CrossRef]
- Bao, Y.; Zhu, L.; Newburg, D.S. Simultaneous Quantification of Sialyloligosaccharides from Human Milk by Capillary Electrophoresis. Anal. Biochem. 2007, 370, 206–214. [Google Scholar] [CrossRef]
- Urashima, T.; Asakuma, S.; Leo, F.; Fukuda, K.; Messer, M.; Oftedal, O.T. The Predominance of Type I Oligosaccharides Is a Feature Specific to Human Breast Milk. Adv. Nutr. 2012, 3, 473S–482S. [Google Scholar] [CrossRef]
- Lane, J.A.; Mariño, K.; Rudd, P.M.; Carrington, S.D.; Slattery, H.; Hickey, R.M. Methodologies for Screening of Bacteria–Carbohydrate Interactions: Anti-Adhesive Milk Oligosaccharides as a Case Study. J. Microbiol. Methods 2012, 90, 53–59. [Google Scholar] [CrossRef]
- Albrecht, S.; Lane, J.A.; Mariño, K.; Al Busadah, K.A.; Carrington, S.D.; Hickey, R.M.; Rudd, P.M. A Comparative Study of Free Oligosaccharides in the Milk of Domestic Animals. Br. J. Nutr. 2014, 111, 1313–1328. [Google Scholar] [CrossRef]
- Urashima, T.; Saito, T.; Nakamura, T.; Messer, M. Oligosaccharides of Milk and Colostrum in Non-Human Mammals. Glycoconj. J. 2001, 18, 357–371. [Google Scholar] [CrossRef]
- Urashima, T.; Taueik, E.; Fukuda, K.; Asakuma, S. Recent Advances in Studies on Milk Oligosaccharides of Cows and Other Domestic Farm Animals. Biosci. Biotechnol. Biochem. 2013, 77, 455–466. [Google Scholar] [CrossRef]
- Martinez-Ferez, A.; Rudloff, S.; Guadix, A.; Henkel, C.A.; Pohlentz, G.; Boza, J.J.; Guadix, E.M.; Kunz, C. Goats’ Milk as a Natural Source of Lactose-Derived Oligosaccharides: Isolation by Membrane Technology. Int. Dairy J. 2006, 16, 173–181. [Google Scholar] [CrossRef]
- Martinez-Ferez, A.; Zapata, J.E.; Guadix, A.; Almecija, M.C.; Gomez, M.; Guadix, E.M. Obtention of Goat Milk Permeates Enriched in Lactose-Derived Oligosaccharides. Desalination 2009, 245, 730–736. [Google Scholar] [CrossRef]
- Silanikove, N.; Leitner, G.; Merin, U.; Prosser, C.G. Recent Advances in Exploiting Goat’s Milk: Quality, Safety and Production Aspects. Small Rumin. Res. 2010, 89, 110–124. [Google Scholar] [CrossRef]
- Quinn, E.M.; Hickey, R.M.; Lokesh, J. Symposium Review: Dairy-Derived Oligosacchrides-Their Influence on Host-Microbe Interactions in the Gastrointestinal Tract of of Infants. J. Dairy Sci. 2020. [Google Scholar] [CrossRef]
- Aquino, L.F.M.C.; de Moura Bell, J.M.L.N.; Cohen, J.L.; Liu, Y.; Lee, H.; de Melo Silva, V.L.; Domizio, P.; Conte Junior, C.A.; Barile, D. Purification of Caprine Oligosaccharides at Pilot-Scale. J. Food Eng. 2017, 214, 226–235. [Google Scholar] [CrossRef]
- Fernández, M.F.; Boris, S.; Barbés, C. Probiotic Properties of Human Lactobacilli Strains to Be Used in the Gastrointestinal Tract. J. Appl. Microbiol. 2003. [Google Scholar] [CrossRef]
- Servin, A.L. Antagonistic Activities of Lactobacilli and Bifidobacteria against Microbial Pathogens. FEMS Microbiol. Rev. 2004. [Google Scholar] [CrossRef]
- Heimesaat, M.M.; Mrazek, K.; Bereswill, S. Murine Fecal Microbiota Transplantation Alleviates Intestinal and Systemic Immune Responses in Campylobacter jejuni Infected Mice Harboring a Human Gut Microbiota. Front. Immunol. 2019. [Google Scholar] [CrossRef]
- Chen, X.; Xu, J.; Shuai, J.; Chen, J.; Zhang, Z.; Fang, W. The S-Layer Proteins of Lactobacillus crispatus Strain ZJ001 Is Responsible for Competitive Exclusion against Escherichia coli O157:H7 and Salmonella typhimurium. Int. J. Food Microbiol. 2007, 115, 307–312. [Google Scholar] [CrossRef]
- Gagnon, M.; Zihler Berner, A.; Chervet, N.; Chassard, C.; Lacroix, C. Comparison of the Caco-2, HT-29 and the Mucus-Secreting HT29-MTX Intestinal Cell Models to Investigate Salmonella Adhesion and Invasion. J. Microbiol. Methods 2013. [Google Scholar] [CrossRef] [PubMed]
- Cairns, M.T.; Gupta, A.; Naughton, J.A.; Kane, M.; Clyne, M.; Joshi, L. Glycosylation-Related Gene Expression in HT29-MTX-E12 Cells upon Infection by Helicobacter pylori. World J. Gastroenterol. 2017. [Google Scholar] [CrossRef]
- Rousset, M. The Human Colon Carcinoma Cell Lines HT-29 and Caco-2: Two In Vitro Models for the Study of Intestinal Differentiation. Biochimie 1986. [Google Scholar] [CrossRef]
- Morrin, S.T.; Owens, R.A.; Le Berre, M.; Gerlach, J.Q.; Joshi, L.; Bode, L.; Irwin, J.A.; Hickey, R.M. Interrogation of Milk-Driven Changes to the Proteome of Intestinal Epithelial Cells by Integrated Proteomics and Glycomics. J. Agric. Food Chem. 2019. [Google Scholar] [CrossRef]
- Bereswill, S.; Ekmekciu, I.; Escher, U.; Fiebiger, U.; Stingl, K.; Heimesaat, M.M. Lactobacillus johnsonii Ameliorates Intestinal, Extra-Intestinal and Systemic pro-Inflammatory Immune Responses Following Murine Campylobacter jejuni Infection. Sci. Rep. 2017. [Google Scholar] [CrossRef]
- WHO. World Health Organization; Campylobacter. Available online: https://www.who.int/news-room/fact-sheets/detail/campylobacter (accessed on 26 February 2020).
- Janssen, R.; Krogfelt, K.A.; Cawthraw, S.A.; van Pelt, W.; Wagenaar, J.A.; Owen, R.J. Host-Pathogen Interactions in Campylobacter Infections: The Host Perspective. Clin. Microbiol. Rev. 2008, 21, 505–518. [Google Scholar] [CrossRef]
- Callaway, T.R.; Edrington, T.S.; Anderson, R.C.; Harvey, R.B.; Genovese, K.J.; Kennedy, C.N.; Venn, D.W.; Nisbet, D.J. Probiotics, Prebiotics and Competitive Exclusion for Prophylaxis against Bacterial Disease. Anim. Heal. Res. Rev. 2008, 9, 217–225. [Google Scholar] [CrossRef]
- Saint-Cyr, M.J.; Guyard-Nicodème, M.; Messaoudi, S.; Chemaly, M.; Cappelier, J.-M.; Dousset, X.; Haddad, N. Recent Advances in Screening of Anti-Campylobacter Activity in Probiotics for Use in Poultry. Front. Microbiol. 2016, 7, 553. [Google Scholar] [CrossRef]
- Alter, T.; Bereswill, S.; Haag, L.M.; Heimesaat, M.M.; Glünder, G.; Rautenschlein, S.; Weber, R.M.; Hänel, I.; Lugert, R.; Zautner, A.E.; et al. Campylobacteriosis of Man: Livestock as Reservoir for Campylobacter Species. Bundesgesundheitsblatt—Gesundheitsforsch.—Gesundheitsschutz 2011. [Google Scholar] [CrossRef]
- Pielsticker, C.; Glünder, G.; Rautenschlein, S. Colonization Properties of Campylobacter jejuni in Chickens. Eur. J. Microbiol. Immunol. 2012. [Google Scholar] [CrossRef]
- Dec, M.; Nowaczek, A.; Urban-Chmiel, R.; Stępień-Pyśniak, D.; Wernicki, A. Probiotic Potential of Lactobacillus Isolates of Chicken Origin with Anti-Campylobacter Activity. J. Vet. Med. Sci. 2018, 80, 1195–1203. [Google Scholar] [CrossRef]
- Baffoni, L.; Gaggìa, F.; Di Gioia, D.; Santini, C.; Mogna, L.; Biavati, B. A Bifidobacterium-Based Synbiotic Product to Reduce the Transmission of C. jejuni along the Poultry Food Chain. Int. J. Food Microbiol. 2012, 157, 156–161. [Google Scholar] [CrossRef] [PubMed]
- Baffoni, L.; Gaggìa, F.; Garofolo, G.; Di Serafino, G.; Buglione, E.; Di Giannatale, E.; Di Gioia, D. Evidence of Campylobacter jejuni Reduction in Broilers with Early Synbiotic Administration. Int. J. Food Microbiol. 2017, 251, 41–47. [Google Scholar] [CrossRef] [PubMed]
- Arsi, K.; Donoghue, A.M.; Woo-Ming, A.; Blore, P.J.; Donoghue, D.J. The Efficacy of Selected Probiotic and Prebiotic Combinations in Reducing Campylobacter Colonization in Broiler Chickens. J. Appl. Poult. Res. 2015, 24, 327–334. [Google Scholar] [CrossRef]
- Garrido, D.; Dallas, D.C.; Mills, D.A. Consumption of Human Milk Glycoconjugates by Infant-Associated Bifidobacteria: Mechanisms and Implications. Microbiology 2013, 159, 649–664. [Google Scholar] [CrossRef] [PubMed]
- Garrido, D.; Kim, J.H.; German, J.B.; Raybould, H.E.; Mills, D.A. Oligosaccharide Binding Proteins from Bifidobacterium longum subsp. infantis Reveal a Preference for Host Glycans. PLoS ONE 2011, 6, e17315. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.-H.; An, H.J.; Garrido, D.; German, J.B.; Lebrilla, C.B.; Mills, D.A. Proteomic Analysis of Bifidobacterium longum subsp. infantis Reveals the Metabolic Insight on Consumption of Prebiotics and Host Glycans. PLoS ONE 2013, 8, e57535. [Google Scholar] [CrossRef]
- LoCascio, R.G.; Ninonuevo, M.R.; Freeman, S.L.; Sela, D.A.; Grimm, R.; Lebrilla, C.B.; Mills, D.A.; German, J.B. Glycoprofiling of Bifidobacterial Consumption of Human Milk Oligosaccharides Demonstrates Strain Specific, Preferential Consumption of Small Chain Glycans Secreted in Early Human Lactation. J. Agric. Food Chem. 2007, 55, 8914–8919. [Google Scholar] [CrossRef]
- LoCascio, R.G.; Desai, P.; Sela, D.A.; Weimer, B.; Mills, D.A. Broad Conservation of Milk Utilization Genes in Bifidobacterium longum subsp. infantis as Revealed by Comparative Genomic Hybridization. Appl. Environ. Microbiol. 2010, 76, 7373–7381. [Google Scholar] [CrossRef]
- Sela, D.A.; Chapman, J.; Adeuya, A.; Kim, J.H.; Chen, F.; Whitehead, T.R.; Lapidus, A.; Rokhsar, D.S.; Lebrilla, C.B.; German, J.B.; et al. The Genome Sequence of Bifidobacterium longum subsp. infantis Reveals Adaptations for Milk Utilization within the Infant Microbiome. Proc. Natl. Acad. Sci. USA 2008, 105, 18964–18969. [Google Scholar] [CrossRef]
- Sela, D.A.; Garrido, D.; Lerno, L.; Wu, S.; Tan, K.; Eom, H.-J.; Joachimiak, A.; Lebrilla, C.B.; Mills, D.A. Bifidobacterium longum subsp. infantis ATCC 15697 α-Fucosidases Are Active on Fucosylated Human Milk Oligosaccharides. Appl. Environ. Microbiol. 2012, 78, 795–803. [Google Scholar] [CrossRef] [PubMed]
- Sela, D.A.; Li, Y.; Lerno, L.; Wu, S.; Marcobal, A.M.; German, J.B.; Chen, X.; Lebrilla, C.B.; Mills, D.A. An Infant-Associated Bacterial Commensal Utilizes Breast Milk Sialyloligosaccharides. J. Biol. Chem. 2011, 286, 11909–11918. [Google Scholar] [CrossRef]
- Garrido, D.; Ruiz-Moyano, S.; Mills, D.A. Release and Utilization of N-Acetyl-D-glucosamine from Human Milk Oligosaccharides by Bifidobacterium longum subsp. infantis. Anaerobe 2012, 18, 430–435. [Google Scholar] [CrossRef] [PubMed]
- Marcobal, A.; Barboza, M.; Froehlich, J.W.; Block, D.E.; German, J.B.; Lebrilla, C.B.; Mills, D.A. Consumption of Human Milk Oligosaccharides by Gut-Related Microbes. J. Agric. Food Chem. 2010, 58, 5334–5340. [Google Scholar] [CrossRef] [PubMed]
- Ward, R.E.; Niñonuevo, M.; Mills, D.A.; Lebrilla, C.B.; German, J.B. In Vitro Fermentability of Human Milk Oligosaccharides by Several Strains of Bifidobacteria. Mol. Nutr. Food Res. 2007, 51, 1398–1405. [Google Scholar] [CrossRef] [PubMed]
- LoCascio, R.G.; Niñonuevo, M.R.; Kronewitter, S.R.; Freeman, S.L.; German, J.B.; Lebrilla, C.B.; Mills, D.A. A Versatile and Scalable Strategy for Glycoprofiling Bifidobacterial Consumption of Human Milk Oligosaccharides. Microb. Biotechnol. 2009, 2, 333–342. [Google Scholar] [CrossRef] [PubMed]
- Bunesova, V.; Lacroix, C.; Schwab, C. Fucosyllactose and L-Fucose Utilization of Infant Bifidobacterium longum and Bifidobacterium kashiwanohense. BMC Microbiol. 2016, 16, 248. [Google Scholar] [CrossRef]
- Macfarlane, S.; Macfarlane, G.T. Regulation of Short-Chain Fatty Acid Production. Proc. Nutr. Soc. 2003, 62, 67–72. [Google Scholar] [CrossRef] [PubMed]
- Rivière, A.; Selak, M.; Lantin, D.; Leroy, F.; De Vuyst, L. Bifidobacteria and Butyrate-Producing Colon Bacteria: Importance and Strategies for Their Stimulation in the Human Gut. Front. Microbiol. 2016, 7, 979. [Google Scholar] [CrossRef]
- Fooks, L.J.; Gibson, G.R. Probiotics as Modulators of the Gut Flora. Br. J. Nutr. 2002, 88, s39. [Google Scholar] [CrossRef]
- Salminen, S.; Bouley, C.; Boutron-Ruault, M.C.; Cummings, J.H.; Franck, A.; Gibson, G.R.; Isolauri, E.; Moreau, M.C.; Roberfroid, M.; Rowland, I. Functional Food Science and Gastrointestinal Physiology and Function. Br. J. Nutr. 1998, 80, S147–S171. [Google Scholar] [CrossRef]
- Ríos-Covián, D.; Ruas-Madiedo, P.; Margolles, A.; Gueimonde, M.; de Los Reyes-Gavilán, C.G.; Salazar, N. Intestinal Short Chain Fatty Acids and Their Link with Diet and Human Health. Front. Microbiol. 2016, 7, 185. [Google Scholar] [CrossRef]
- Louis, P.; Scott, K.P.; Duncan, S.H.; Flint, H.J. Understanding the Effects of Diet on Bacterial Metabolism in the Large Intestine. J. Appl. Microbiol. 2007, 102, 1197–1208. [Google Scholar] [CrossRef]
- Fukuda, S.; Toh, H.; Hase, K.; Oshima, K.; Nakanishi, Y.; Yoshimura, K.; Tobe, T.; Clarke, J.M.; Topping, D.L.; Suzuki, T.; et al. Bifidobacteria Can Protect from Enteropathogenic Infection through Production of Acetate. Nature 2011, 469, 543–547. [Google Scholar] [CrossRef]
- Özcan, E.; Sela, D.A. Inefficient Metabolism of the Human Milk Oligosaccharides Lacto-N-Tetraose and Lacto-N-Neotetraose Shifts Bifidobacterium longum subsp. infantis Physiology. Front. Nutr. 2018, 5. [Google Scholar] [CrossRef] [PubMed]
- Palframan, R.J.; Gibson, G.R.; Rastall, R.A. Carbohydrate Preferences of Bifidobacterium Species Isolated from the Human Gut. Curr. Issues Intest. Microbiol. 2003, 4, 71–75. [Google Scholar]
- Centanni, M.; Ferguson, S.A.; Sims, I.M.; Biswas, A.; Tannocka, G.W. Bifidobacterium bifidum ATCC 15696 and Bifidobacterium breve 24b Metabolic Interaction Based on 2′-O- Fucosyl-Lactose Studied in Steady-State Cultures in a Freter-Style Chemostat. Appl. Environ. Microbiol. 2019, 85. [Google Scholar] [CrossRef] [PubMed]
- Perrin, S.; Warchol, M.; Grill, J.P.; Schneider, F. Fermentations of Fructo-Oligosaccharides and Their Components by Bifidobacterium infantis ATCC 15697 on Batch Culture in Semi-Synthetic Medium. J. Appl. Microbiol. 2001, 90, 859–865. [Google Scholar] [CrossRef] [PubMed]
- Lauer, E.; Kandler, O. Mechanism of the Variation of the Acetate/Lactate/Ratio during Glucose Fermentation by Bifidobacteria. Arch. Microbiol. 1976, 110, 271–277. [Google Scholar] [CrossRef] [PubMed]
- De Vuyst, L.; Leroy, F. Cross-Feeding between Bifidobacteria and Butyrate-Producing Colon Bacteria Explains Bifdobacterial Competitiveness, Butyrate Production, and Gas Production. Int. J. Food Microbiol. 2011, 149, 73–80. [Google Scholar] [CrossRef] [PubMed]
- De Vuyst, L.; Moens, F.; Selak, M.; Rivière, A.; Leroy, F. Summer Meeting 2013: Growth and Physiology of Bifidobacteria. J. Appl. Microbiol. 2014, 116, 477–491. [Google Scholar] [CrossRef] [PubMed]
- Rivière, A.; Gagnon, M.; Weckx, S.; Roy, D.; De Vuyst, L. Mutual Cross-Feeding Interactions between Bifidobacterium longum subsp. longum NCC2705 and Eubacterium rectale ATCC 33656 Explain the Bifidogenic and Butyrogenic Effects of Arabinoxylan Oligosaccharides. Appl. Environ. Microbiol. 2015, 81, 7767–7781. [Google Scholar] [CrossRef]
- Flint, S.W.; Hudson, J.; Lavallee, D. UK Adults’ Implicit and Explicit Attitudes towards Obesity: A Cross-Sectional Study. BMC Obes. 2015, 2, 31. [Google Scholar] [CrossRef] [PubMed]
- Rinninella, E.; Raoul, P.; Cintoni, M.; Franceschi, F.; Miggiano, G.A.D.; Gasbarrini, A.; Mele, M.C. What Is the Healthy Gut Microbiota Composition? A Changing Ecosystem across Age, Environment, Diet, and Diseases. Microorganisms 2019, 7, 14. [Google Scholar] [CrossRef]
- Mazmanian, S.K.; Lee, Y.K. Interplay between Intestinal Microbiota and Host Immune System. J. Bacteriol. Virol. 2014, 1–9. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
| HPLC Standard | Per 100 mL | Molecular Weights |
|---|---|---|
| 10 mM Lactic acid | 0.09 g | 90.08 |
| 10 mM Acetic acid | 57 μL | 60.05 |
| 10 mM Formic Acid | 38 μL | 46.03 |
| 10 mM Ethanol (99%) | 58 μL | 46.07 |
| Oligosaccharide Structures | μg/mL |
|---|---|
| Lacto-N-neotetraose (LNnT) | 3.4 |
| Lacto-N-neohexaose (LNnH) | 3 |
| 3′-Sialyllactose (3′SL) | 32.83 |
| 6′-Sialyllactose (6′SL) | 33.05 |
| LS-tetrasaccharide c | 0.94 |
| Disialyllactose (DSL) | 33.46 |
| Control | GMO | |||
|---|---|---|---|---|
| Concentration mM | 0 h | 24 h | 0 h | 24 h |
| Acetate | ND | 2.42 a | 0.07 | 30.14 a,b |
| Lactate | ND | 0.62 n.s | ND | 9.04 a,b |
| Formate | 0.81 | 1.13 a | 0.79 b | 8.57 a,b |
| Ethanol | ND | ND | ND | 8.00 a,b |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Quinn, E.M.; Slattery, H.; Walsh, D.; Joshi, L.; Hickey, R.M. Bifidobacterium longum subsp. infantis ATCC 15697 and Goat Milk Oligosaccharides Show Synergism In Vitro as Anti-Infectives against Campylobacter jejuni. Foods 2020, 9, 348. https://doi.org/10.3390/foods9030348
Quinn EM, Slattery H, Walsh D, Joshi L, Hickey RM. Bifidobacterium longum subsp. infantis ATCC 15697 and Goat Milk Oligosaccharides Show Synergism In Vitro as Anti-Infectives against Campylobacter jejuni. Foods. 2020; 9(3):348. https://doi.org/10.3390/foods9030348
Chicago/Turabian StyleQuinn, Erinn M., Helen Slattery, Dan Walsh, Lokesh Joshi, and Rita M. Hickey. 2020. "Bifidobacterium longum subsp. infantis ATCC 15697 and Goat Milk Oligosaccharides Show Synergism In Vitro as Anti-Infectives against Campylobacter jejuni" Foods 9, no. 3: 348. https://doi.org/10.3390/foods9030348
APA StyleQuinn, E. M., Slattery, H., Walsh, D., Joshi, L., & Hickey, R. M. (2020). Bifidobacterium longum subsp. infantis ATCC 15697 and Goat Milk Oligosaccharides Show Synergism In Vitro as Anti-Infectives against Campylobacter jejuni. Foods, 9(3), 348. https://doi.org/10.3390/foods9030348