Development of a DNA-Based Detection Method for Cocos Nucifera Using TaqMan™ Real-Time PCR


Abstract
1. Introduction
2. Materials and Methods
2.1. Sample Material
2.2. Sample Preparation and DNA Extraction
2.3. Determination of DNA Concentration and Purity
2.4. PCR Primers and Probes
2.5. Conventional PCR
2.6. Gel Electrophoresis
2.7. DNA Sequencing
2.8. Real-Time PCR
2.8.1. Specificity of the Real-Time PCR Assay
3. Results
3.1. Extraction of DNA from Palm Leaves
3.2. Verification of the Botanical Origin of the Palm Leaves
3.3. Design and Specificity of Coconut-Specific Primers
3.4. PCR Amplification of DNA from Coconut, Coconut Chips, and Coconut Blossom Sugar
3.5. Development of a TaqMan™ Real-Time PCR Assay
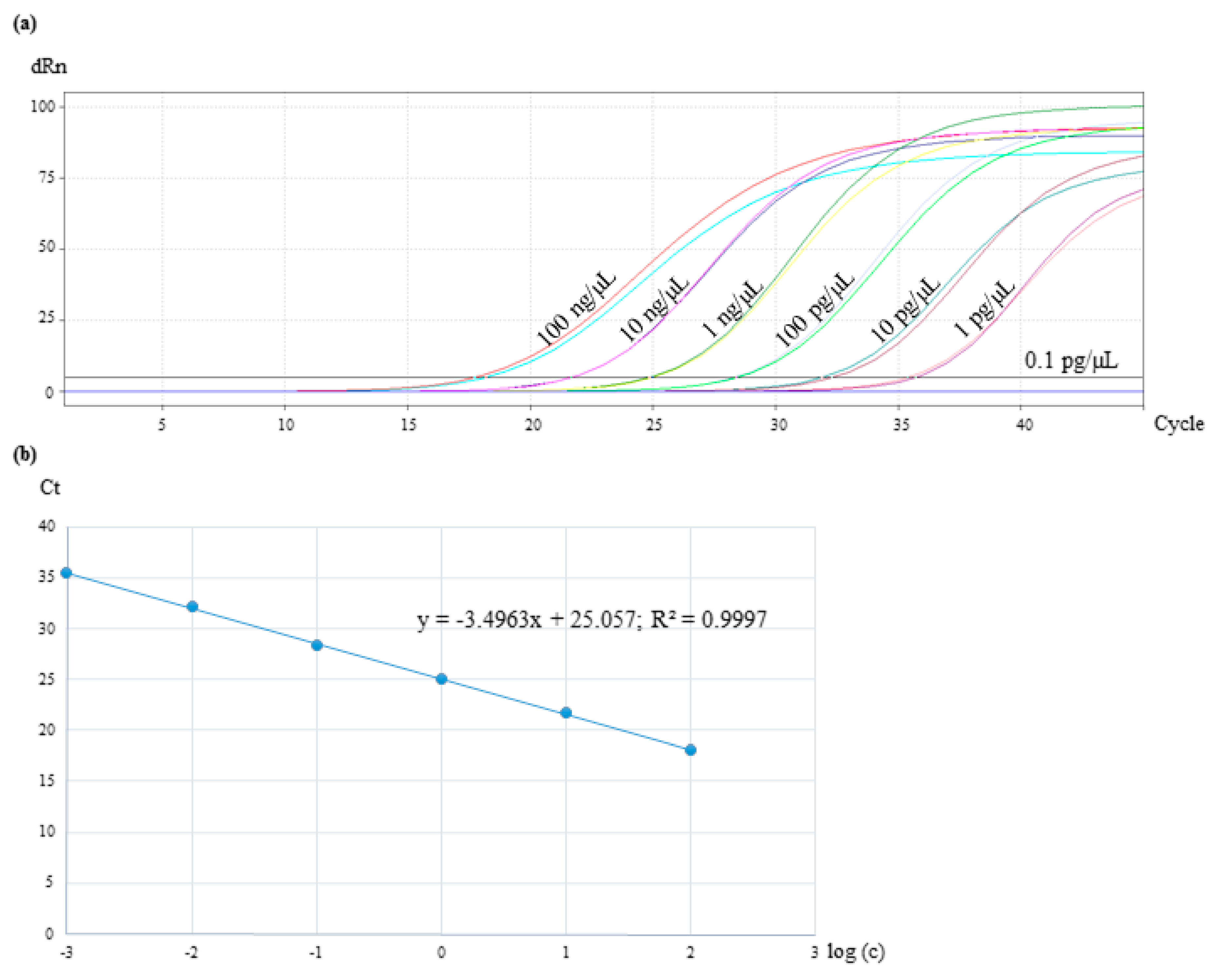
3.6. Assay Specificity and Sensitivity
4. Discussion
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Lieberei, R.; Reisdorff, C. Nutzpflanzen; Georg Thieme Verlag: New York, NY, USA, 2012. [Google Scholar]
- FAOSTAT. Crops Statistics-Production Quantities of Coconuts by Country 2017. Available online: http://www.fao.org/faostat/en/#data/QC/visualize (accessed on 20 May 2019).
- Anagnostou, K. Coconut allergy revisited. Children 2017, 4, 85. [Google Scholar] [CrossRef] [PubMed]
- Benito, C.; González-Mancebo, E.; de Durana, M.D.A.D.; Tolón, R.M.; Fernández-Rivas, M. Identification of a 7S globulin as a novel coconut allergen. Ann. Allergy Asthma Immunol. 2007, 98, 580–584. [Google Scholar] [CrossRef]
- Carr, H.; Plumb, G.; Parker, M.; Lambert, N. Characterisation and crystallisation of an 11S seed storage globulin from coconut (Cocos nucifera). Food Chem. 1990, 38, 11–20. [Google Scholar] [CrossRef]
- Garcia, R.N.; Arocena, R.V.; Laurena, A.C.; Tecson-Mendoza, E.M. 11S and 7S globulins of coconut (Cocos nucifera L.): Purification and characterization. J. Agric. Food. Chem. 2005, 53, 1734–1739. [Google Scholar] [CrossRef]
- EU. Regulation (EU) No 1169/2011 of the European Parliament and of the Council of 25 October 2011 on the provision of food information to consumers, amending Regulations (EC) No 1924/2006 and (EC) No 1925/2006 of the European Parliament and of the Council, and repealing Commission Directive 87/250/EEC, Council Directive 90/496/EEC, Commission Directive 1999/10/EC, Directive 2000/13/EC of the European Parliament and of the Council, Commission Directives 2002/67/EC and 2008/5/EC and Commission Regulation (EC) No 608/2004; European Union: Brussels, Belgium, 2011. [Google Scholar]
- FDA. A Food Labeling Guide; Center for Food Safety and Applied Nutrition-Office of Nutrition, Labeling, and Dietary Supplements; FDA: College Park, MD, USA, 2013.
- Galan, A.M.G.; Brohée, M.; de Andrade Silva, E.; van Hengel, A.J.; Chassaigne, H. Development of a real-time PCR method for the simultaneous detection of soya and lupin mitochondrial DNA as markers for the presence of allergens in processed food. Food Chem. 2011, 127, 834–841. [Google Scholar] [CrossRef]
- Demmel, A.; Hupfer, C.; Ilg Hampe, E.; Busch, U.; Engel, K.-H. Development of a real-time PCR for the detection of lupine DNA (Lupinus species) in foods. J. Agric. Food Chem. 2008, 56, 4328–4332. [Google Scholar] [CrossRef]
- Houhoula, D.; Koussissis, S.; Lougovois, V.; Tsaknis, J.; Kassavita, D.; Papatheodorou, S.; Akkermans, S.; Van Impe, J.; Tsakali, E. Detection of peanut allergen traces with a Real Time PCR assay-the challenge to protect food-allergic consumers. J. Food Res. 2018, 7, 32–37. [Google Scholar] [CrossRef][Green Version]
- Houhoula, D.; Papatheodorou, S.A.; Moschou, D.; Pappa, S.; Tsaatazoglou, N.; Koussissis, S.; Tsaknis, J.; Lougovois, V.; Impe, J.; Tsakali, E. Evaluation of a Real Time PCR Assay and a ELISA Method for the Detection of Walnuts and Almonds Allergen Traces in Food Products. J. Food Res. 2019, 8, 71–76. [Google Scholar] [CrossRef][Green Version]
- BVL. Amtliche Sammlung von Untersuchungsverfahren nach § 64 LFGB, Vefahren zum Nachweis von gentechnisch modifizierten Organismen und ihren Produkten in Lebensmitteln L 00.00-119:2014-02; Bundesamt für Verbraucherschutz und Lebensmittelsicherheit: Braunschweig, Germany, 2014.
- Miziorko, H.M. Phosphoribulokinase: Current perspectives on the structure/function basis for regulation and catalysis. Adv. Enzymol. Relat. Areas Mol. Biol. 2000, 74, 95–127. [Google Scholar]
- Baker, W.J.; Norup, M.V.; Clarkson, J.J.; Couvreur, T.L.; Dowe, J.L.; Lewis, C.E.; Pintaud, J.-C.; Savolainen, V.; Wilmot, T.; Chase, M.W. Phylogenetic relationships among arecoid palms (Arecaceae: Arecoideae). Ann. Bot. 2011, 108, 1417–1432. [Google Scholar] [CrossRef]
- Gunn, B.F. The phylogeny of the Cocoeae (Arecaceae) with emphasis on Cocos nucifera. Ann. Mo. Bot. Gard. 2004, 91, 505–522. [Google Scholar]
- Porter, C.H.; Collins, F.H. Species-diagnostic differences in a ribosomal DNA internal transcribed spacer from the sibling species Anopheles freeborni and Anopheles hermsi (Diptera: Culicidae). Am. J. Trop. Med. Hyg. 1991, 45, 271–279. [Google Scholar] [CrossRef] [PubMed]
- Schultz, J.; Müller, T.; Achtziger, M.; Seibel, P.N.; Dandekar, T.; Wolf, M. The internal transcribed spacer 2 database—A web server for (not only) low level phylogenetic analyses. Nucleic Acids Res. 2006, 34, 704–707. [Google Scholar] [CrossRef] [PubMed]
- Yao, H.; Song, J.; Liu, C.; Luo, K.; Han, J.; Li, Y.; Pang, X.; Xu, H.; Zhu, Y.; Xiao, P. Use of ITS2 region as the universal DNA barcode for plants and animals. PLoS ONE 2010, 5, e13102. [Google Scholar] [CrossRef] [PubMed]
- Zeltner, D.; Glomb, M.A.; Maede, D. Real-time PCR systems for the detection of the gluten-containing cereals wheat, spelt, kamut, rye, barley and oat. Eur. Food Res. Technol. 2009, 228, 321–330. [Google Scholar] [CrossRef]
- Allmann, M.; Candrian, U.; Höfelein, C.; Lüthy, J. Polymerase chain reaction (PCR): A possible alternative to immunochemical methods assuring safety and quality of food Detection of wheat contamination in non-wheat food products. Unters. Forsch. 1993, 196, 248–251. [Google Scholar] [CrossRef]
- Atputharajah, J.; Widanapathirana, S.; Samarajeewa, U. Microbiology and biochemistry of natural fermentation of coconut palm sap. Food Microbiol. 1986, 3, 273–280. [Google Scholar] [CrossRef]
- Hebbar, K.; Arivalagan, M.; Manikantan, M.; Mathew, A.; Thamban, C.; Thomas, G.V.; Chowdappa, P. Coconut inflorescence sap and its value addition as sugar–collection techniques, yield, properties and market perspective. Curr. Sci. 2015, 1411–1417. [Google Scholar] [CrossRef]
- Lee, L.G.; Connell, C.R.; Bloch, W. Allelic discrimination by nick-translation PCR with fluorgenic probes. Nucleic Acids Res. 1993, 21, 3761–3766. [Google Scholar] [CrossRef]
- Holland, P.M.; Abramson, R.D.; Watson, R.; Gelfand, D.H. Detection of specific polymerase chain reaction product by utilizing the 5′----3′exonuclease activity of Thermus aquaticus DNA polymerase. Proc. Natl. Acad. Sci. USA 1991, 88, 7276–7280. [Google Scholar] [CrossRef]
- Livak, K.J.; Flood, S.; Marmaro, J.; Giusti, W.; Deetz, K. Oligonucleotides with fluorescent dyes at opposite ends provide a quenched probe system useful for detecting PCR product and nucleic acid hybridization. Genome Res. 1995, 4, 357–362. [Google Scholar] [CrossRef] [PubMed]
- Ju, J.; Ruan, C.; Fuller, C.W.; Glazer, A.N.; Mathies, R.A. Fluorescence energy transfer dye-labeled primers for DNA sequencing and analysis. Proc. Natl. Acad. Sci. USA 1995, 92, 4347–4351. [Google Scholar] [CrossRef] [PubMed]
- Johansson, M.K.; Cook, R.M. Intramolecular dimers: A new design strategy for fluorescence-quenched probes. Chem. Eur. J. 2003, 9, 3466–3471. [Google Scholar] [CrossRef]
- Biosearchtech. Black Hole Quencher® Dyes. Available online: https://www.biosearchtech.com/support/education/fluorophores-and-quenchers/black-hole-quencher-dyes (accessed on 16 May 2019).
- García, A.; Madrid, R.; García, T.; Martín, R.; González, I. Detection of food allergens by Taqman real-time PCR methodology. In Food Allergens: Methods and Protocols; Lin, J., Alcocer, M., Eds.; Springer: New York, NY, USA, 2017; pp. 95–108. [Google Scholar] [CrossRef]
- Busch, U. Molekularbiologische Methoden in der Lebensmittelanalytik: Grundlegende Methoden und Anwendungen; Springer: Berlin, Germany, 2010. [Google Scholar]
- WHO/IUIS. Allergen Nomenclature. World Health Organization and International Union of Immunological Societies. Available online: http://www.allergen.org/ (accessed on 17 May 2019).
- Magi, M.; Garcia, L.; Vandenbranden, M.; Palmantier, R.; Jacquet, A. Heat denaturation affects the ProDer p 1 IgE reactivity and downregulates the development of the specific allergic response. J. Allergy Clin. Immunol. 2004, 114, 545–552. [Google Scholar] [CrossRef] [PubMed]
- Johansen, P.; Senti, G.; Martínez Gómez, J.M.; Wüthrich, B.; Bot, A.; Kündig, T.M. Heat denaturation, a simple method to improve the immunotherapeutic potential of allergens. Eur. J. Immunol. 2005, 35, 3591–3598. [Google Scholar] [CrossRef] [PubMed]
- EU. Directive 2010/63/EU of the European Parliament and of the Council of 22 September 2010 on the Protection of Animals Used for Scientific Purposes; European Union: Brussels, Belgium, 2010 22 September. [Google Scholar]
- Stephan, O.; Vieths, S. Development of a real-time PCR and a sandwich ELISA for detection of potentially allergenic trace amounts of peanut (Arachis hypogaea) in processed foods. J. Agric. Food Chem. 2004, 52, 3754–3760. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer/Probe Name | Description | Target Gene | Sequence (5′ → 3′) |
|---|---|---|---|
| euk18S-for. | Eukaryotic-specific for. primer | 18SrRNA | TCT GCC CTA TCA ACT TTC GAT GGT A |
| euk18S-rev. | Eukaryotic-specific rev. primer | 18SrRNA | AAT TTG CGC GCC TGC TGC CTT CCT T |
| cocosITS109-for. | Coconut-specific for. primer | ITS | GGC CTC CTG AGG TAT ATC CG |
| cocosITS109-rev. | Coconut-specific rev. primer | ITS | CAT CCA CCA TCC ACC GTG TC |
| cocosITS197-for. | Coconut-specific for. primer | ITS | TAT CCG GAT GTG GAT GCT GG |
| cocosITS197-rev. | Coconut-specific rev. primer | ITS | CAT CCG ATG GCT GGG GTG |
| cocosPRK-for. | Coconut-specific for. primer | prk | ACA AGA CCT ACT GGA CTG G |
| cocos-PRK-rev. | Coconut-specific rev. primer | prk | TCT GAT ATG TAT AAG ACT CAG CA |
| cocosPRK | Coconut probe | prk | FAM-AAT TGT CTC ATT ATC TCA ATG AAC CGG GTG-BHQ1 |
| palmPRK-for. | Palmae-specific for. primer | prk | CTA GCA AAG AAT CTG ATC GAT AAG T |
| palmPRK-rev. | Palmae-specific rev. primer | prk | CAT ATT GCT TCT GTG GGT CTG |
| Palmea Species | Sample | DNA Concentration [µg/µL] | A260/A280 | Amplifiability |
|---|---|---|---|---|
| Chilean wine palm (Jubaea chilensis) | A B C D | 1.64 2.21 1.60 1.47 | 2.00 2.07 2.01 2.01 | + |
| Coconut palm (Cocos nucifera) | A B C B | 1.88 1.86 1.86 1.77 | 1.98 1.92 1.92 1.97 | + |
| Jelly palm (Butia capitata) | A B C D | 2.71 2.80 2.35 2.49 | 1.94 1.94 1.93 1.93 | + |
| Oil palm (Elaeis guineensis) | A B C D | 2.73 2.98 2.60 1.91 | 1.97 1.97 1.98 2.01 | + |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wrage, J.; Kleyner, O.; Rohn, S.; Kuballa, J. Development of a DNA-Based Detection Method for Cocos Nucifera Using TaqMan™ Real-Time PCR. Foods 2020, 9, 332. https://doi.org/10.3390/foods9030332
Wrage J, Kleyner O, Rohn S, Kuballa J. Development of a DNA-Based Detection Method for Cocos Nucifera Using TaqMan™ Real-Time PCR. Foods. 2020; 9(3):332. https://doi.org/10.3390/foods9030332
Chicago/Turabian StyleWrage, Jasmin, Oxana Kleyner, Sascha Rohn, and Jürgen Kuballa. 2020. "Development of a DNA-Based Detection Method for Cocos Nucifera Using TaqMan™ Real-Time PCR" Foods 9, no. 3: 332. https://doi.org/10.3390/foods9030332
APA StyleWrage, J., Kleyner, O., Rohn, S., & Kuballa, J. (2020). Development of a DNA-Based Detection Method for Cocos Nucifera Using TaqMan™ Real-Time PCR. Foods, 9(3), 332. https://doi.org/10.3390/foods9030332