Residues of Fluoroquinolone Antibiotics Induce Carbonylation and Reduce In Vitro Digestion of Sarcoplasmic and Myofibrillar Beef Proteins


Abstract

1. Introduction
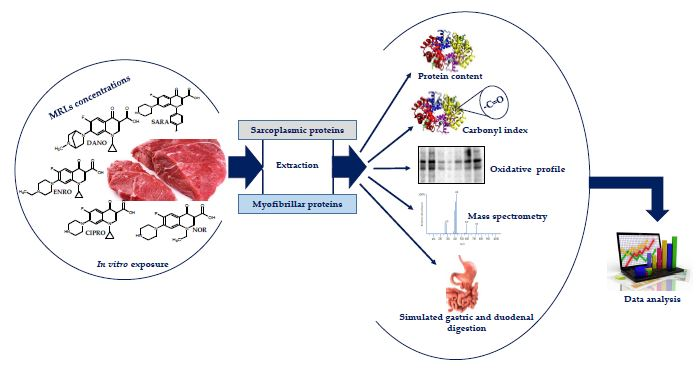
2. Materials and Methods
2.1. Reagents and Materials
2.2. Experimental Design
2.2.1. Meat Samples Obtention
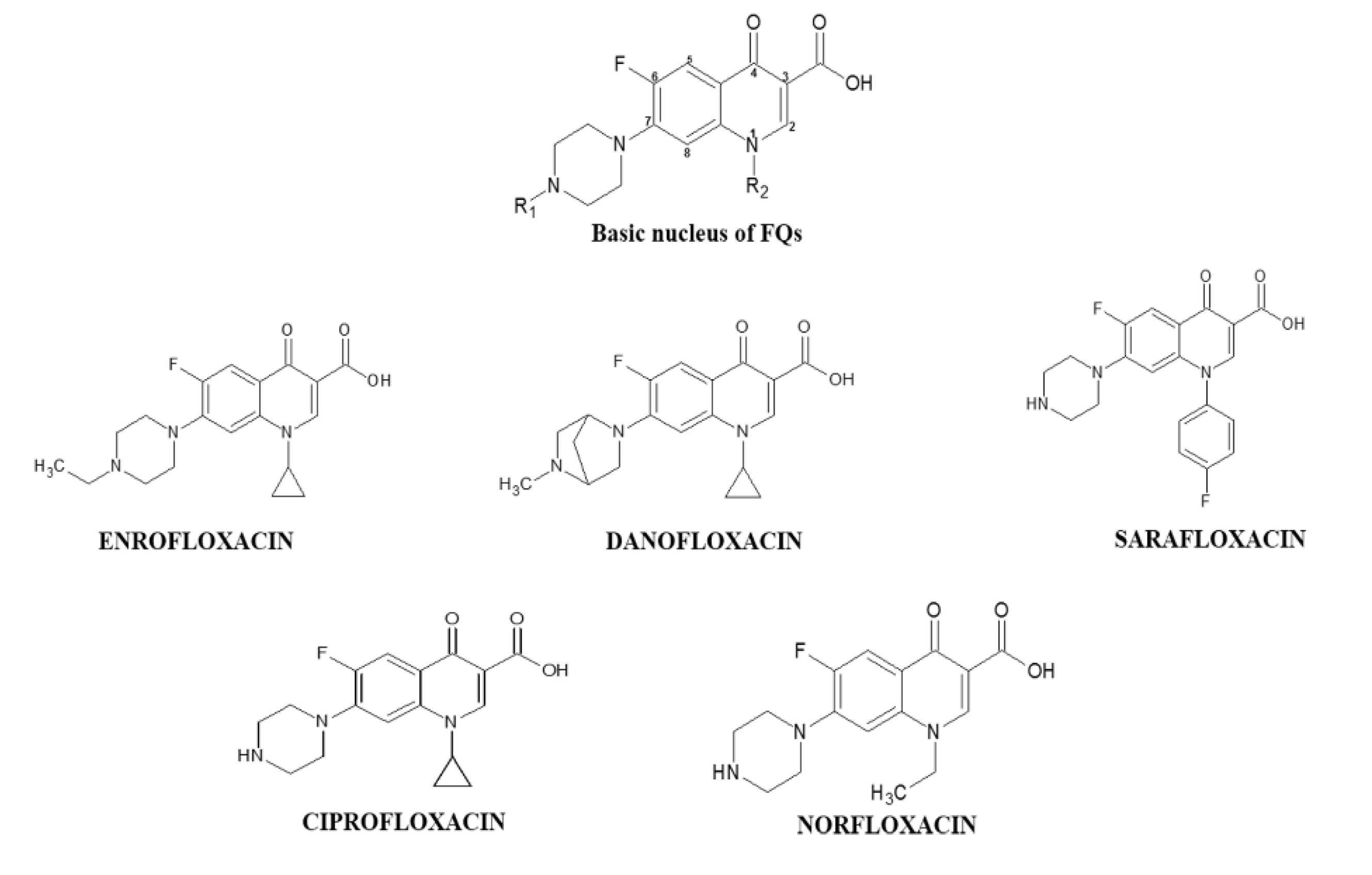
2.2.2. Fluoroquinolones Working Solutions
2.2.3. Preparation of Fluoroquinolones (FQs) Treated Samples
2.3. Protein Extraction and Obtaining Electrophoretic Profiles (Sodium Dodecyl Sulfate–Polyacrylamide Gel Electrophoresis, SDS-PAGE)
2.4. Determination of Carbonyl Index
2.5. Western Blot
2.6. Identification of Proteins in Carbonylated Bands
2.7. In Vitro Digestibility Test
2.8. Statistical Analysis
3. Results and Discussion
3.1. Quantitation of Protein Carbonylation Induced by FQs Antibiotics
3.2. Yielding of Protein Fractions is Affected by Protein Oxidation Induced by Fluoroquinolones
3.3. Identification of Carbonylated Proteins
3.4. Effects of Carbonylation on the In Vitro Digestibility of Proteins
3.4.1. In Vitro Digestibility of Sarcoplasmic and Myofibrillar Proteins Exposed to Danofloxacin (DANO)
3.4.2. In Vitro Digestibility of Sarcoplasmic and Myofibrillar Proteins Exposed to Enrofloxacin (ENRO)
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Lynch, D.; Henihan, A.M.; Bowen, B.; McDonnell, K.; Kwapinski, W.; Leahy, J.J. Utilisation of poultry litter as an energy feedstock. Biomass Bioenergy 2013, 49, 197–204. [Google Scholar] [CrossRef]
- Sneeringer, S.; MacDonald, J.; Key, N.; McBride, W. Economics of Antibiotic Use in U.S. In Livestock Production; ERR-200, U.S.; Department of Agriculture, Economic Research Service: Washington, DC, USA, 2015. [Google Scholar]
- Bacanlı, M.; Başaran, N. Importance of antibiotic residues in animal food. Food Chem. Toxicol. 2019, 125, 462–466. [Google Scholar] [CrossRef] [PubMed]
- Terrado-Campos, D.; Tayeb-Cherif, K.; Peris-Vicente, J.; Carda-Broch, S.; Esteve-Romero, J. Determination of oxolinic acid, danofloxacin, ciprofloxacin, and enrofloxacin in porcine and bovine meat by micellar liquid chromatography with fluorescence detection. Food Chem. 2017, 221, 1277–1284. [Google Scholar] [CrossRef]
- Bousova, K.; Senyuva, H.; Mittendorf, K. Quantitative multi-residue method for determination antibiotics in chicken meat using turbulent flow chromatography coupled to liquid chromatography–tandem mass spectrometry. J. Chromatogr. A 2013, 1274, 19–27. [Google Scholar] [CrossRef] [PubMed]
- Menkem, Z.E.; Ngangom, B.L.; Tamunjoh, S.S.A.; Boyom, F.F. Antibiotic residues in food animals: Public health concern. Acta Ecol. Sin. 2018, 39, 411–415. [Google Scholar] [CrossRef]
- Aslam, B.; Kousar, N.; Javed, I.; Raza, A.; Ali, A.; Khaliq, T.; Muhammad, F.; Khan, J.A. Determination of enrofloxacin residues in commercial broilers using high performance liquid chromatography. Int. J. Food Prop. 2016, 19, 2463–2470. [Google Scholar] [CrossRef]
- Omotoso, A.B.; Omojola, A.B. Fluoroquinolone residues in raw meat from open markets in Ibadan, Southwest, Nigeria. Int. J. Heal. Anim. Sci. Food Saf. 2015, 2, 32–40. [Google Scholar]
- Baghani, A.; Mesdaghinia, A.; Rafieiyan, M.; Soltan Dallal, M.M.; Douraghi, M. Tetracycline and ciprofloxacin multiresidues in beef and chicken meat samples using indirect competitive elisa. J. Immunoass. Immunochem. 2019, 40, 328–342. [Google Scholar] [CrossRef]
- Er, B.; Kaynak Onurdǎ, F.; Demirhan, B.; Özgen Özgacar, S.; Bayhan Öktem, A.; Abbasoǧlu, U. Screening of quinolone antibiotic residues in chicken meat and beef sold in the markets of ankara, Turkey. Poult. Sci. 2013, 92, 2212–2215. [Google Scholar] [CrossRef]
- Mashak, Z.; Mojaddarlangroodi, A.; Mehdizadeh, T.; Fathabad, A.E. Detection of quinolones residues in beef and chicken meat in hypermarkets of urmia, iran using elisa. Iran. Agric. Res. 2017, 36, 73–77. [Google Scholar]
- FAO; OECD. OECD/FAO (2018), OECD-FAO Agricultural Outlook 2018–2027; OECD Publishing: Paris, France, 2018. [Google Scholar]
- European Commission. Commission regulation (EU) Nº 37/2010. Off. J. Eur. Union 2010, L15, 1–72. [Google Scholar]
- Carreras, I.; Castellari, M.; García Regueiro, J.; Guerrero, L.; Esteve-Garcia, E.; Sárraga, C. Influence of enrofloxacin administration and alpha-tocopheryl acetate supplemented diets on oxidative stability of broiler tissues. Poult. Sci. 2004, 83, 796–802. [Google Scholar] [CrossRef] [PubMed]
- Gomes, M.P.; Gonçalves, C.A.; de Brito, J.C.M.; Souza, A.M.; da Silva Cruz, F.V.; Bicalho, E.M.; Figueredo, C.C.; Garcia, Q.S. Diffusion of nitrate and water in pork meat: Effect of the direction of the meat fiber. J. Hazard. Mater. 2017, 328, 140–149. [Google Scholar] [CrossRef] [PubMed]
- Ibrahim, I.G.; Yarsan, E. Enrofloxacin drug induced reactive oxygen species. Res. Opin. Anim. Vet. Sci. 2011, 1, 489–491. [Google Scholar]
- Sehonova, P.; Tokanova, N.; Hodkovicova, N.; Kocour Kroupova, H.; Tumova, J.; Blahova, J.; Marsalek, P.; Plhalova, L.; Doubkova, V.; Dobsikova, R.; et al. Oxidative stress induced by fluoroquinolone enrofloxacin in zebrafish (Danio Rerio) can be ameliorated after a prolonged exposure. Environ. Toxicol. Pharmacol. 2019, 67, 87–93. [Google Scholar] [CrossRef]
- Estévez, M.; Luna, C. Dietary Protein Oxidation: A Silent Threat to Human Health? Crit. Rev. Food Sci. Nutr. 2017, 57, 3781–3793. [Google Scholar] [CrossRef]
- Estévez, M. Protein carbonyls in meat systems: A review. Meat Sci. 2011, 89, 259–279. [Google Scholar] [CrossRef]
- Wang, Z.; He, Z.; Gan, X.; Li, H. Effect of peroxyl radicals on the structure and gel properties of isolated rabbit meat myofibrillar proteins. Int. J. Food Sci. Technol. 2018, 53, 2687–2696. [Google Scholar] [CrossRef]
- Gómez, J.; Sanjuán, N.; Arnau, J.; Bon, J.; Clemente, G. Diffusion of nitrate and water in pork meat: Effect of the direction of the meat fiber. J. Food Eng. 2017, 214, 69–78. [Google Scholar] [CrossRef]
- Lucatello, L.; Cagnardi, P.; Capolongo, F.; Ferraresi, C.; Bernardi, F.; Montesissa, C. Development and validation of an lc-ms/ms/ms method for the quantification of fluoroquinolones in several matrices from treated Turkeys. Food Control. 2015, 48, 2–11. [Google Scholar] [CrossRef]
- Molette, C.; Rémignon, H.; Babilé, R. Maintaining muscles at a high post-mortem temperature induces pse-like meat in Turkey. Meat Sci. 2003, 63, 525–532. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Mesquita, C.S.; Oliveira, R.; Bento, F.; Geraldo, D.; Rodrigues, J.V.; Marcos, J.C. Simplified 2,4-dinitrophenylhydrazine spectrophotometric assay for quantification of carbonyls in oxidized proteins. Anal. Biochem. 2014, 458, 69–71. [Google Scholar] [CrossRef] [PubMed]
- Sechi, S.; Chait, B.T. Modification of cysteine residues by alkylation. a tool in peptide mapping and protein identification. Anal. Chem. 1998, 70, 5150–5158. [Google Scholar] [CrossRef] [PubMed]
- Hu, J.; Zhou, C.; Ma, H.; Yu, X.; Qin, X.; Yagoub, A.E.A. Hydrolysis of rapeseed meal protein under simulated duodenum digestion: Kinetic modeling and antioxidant activity. LWT Food Sci. Technol. 2015, 68, 523–531. [Google Scholar]
- Nunes, B.; Veiga, V.; Frankenbach, S.; Serôdio, J.; Pinto, G. Evaluation of physiological changes induced by the fluoroquinolone antibiotic ciprofloxacin in the freshwater macrophyte species Lemna minor and Lemna gibba. Environ. Toxicol. Pharmacol. 2019, 72, 103242. [Google Scholar] [CrossRef]
- Rodriguez, E.; Moreno-Bondi, M.C.; Marazuela, M.D. Multiresidue determination of fluoroquinolone antimicrobials in baby foods by liquid chromatography. Food Chem. 2011, 127, 1354–1360. [Google Scholar] [CrossRef]
- Guyon, C.; Le Vessel, V.; Meynier, A.; de Lamballerie, M. Modifications of protein-related compounds of beef minced meat treated by high pressure. Meat Sci. 2018, 142, 32–37. [Google Scholar] [CrossRef]
- Holman, B.W.B.; Coombs, C.E.O.; Morris, S.; Kerr, M.J.; Hopkins, D.L. Effect of long term chilled (up to 5 weeks) then frozen (up to 12 months) storage at two different sub-zero holding temperatures on beef: 3. protein structure degradation and a marker of protein oxidation. Meat Sci. 2018, 139, 171–178. [Google Scholar] [CrossRef]
- Turgut, S.S.; Işıkçı, F.; Soyer, A. Antioxidant activity of pomegranate peel extract on lipid and protein oxidation in beef meatballs during frozen storage. Meat Sci. 2017, 129, 111–119. [Google Scholar] [CrossRef]
- Della Malva, A.; Marino, R.; Santillo, A.; Annicchiarico, G.; Caroprese, M.; Sevi, A.; Albenzio, M. Proteomic approach to investigate the impact of different dietary supplementation on lamb meat tenderness. Meat Sci. 2017, 131, 74–81. [Google Scholar] [CrossRef] [PubMed]
- Marcos, B.; Kerry, J.P.; Mullen, A.M. High pressure induced changes on sarcoplasmic protein fraction and quality indicators. Meat Sci. 2010, 85, 115–120. [Google Scholar] [CrossRef] [PubMed]
- Xia, X.; Kong, B.; Liu, Q.; Liu, J. Physicochemical change and protein oxidation in porcine longissimus dorsi as influenced by different freeze-thaw cycles. Meat Sci. 2009, 83, 239–245. [Google Scholar] [CrossRef] [PubMed]
- Blokhina, S.V.; Sharapova, A.V.; Ol’khovich, M.V.; Volkova, T.V.; Perlovich, G.L. Solubility, lipophilicity and membrane permeability of some fluoroquinolone antimicrobials. Eur. J. Pharm. Sci. 2016, 93, 29–37. [Google Scholar] [CrossRef] [PubMed]
- Cramariuc, O.; Rog, T.; Javanainen, M.; Monticelli, L.; Polishchuk, A.V.; Vattulainen, I. Mechanism for translocation of fluoroquinolones across lipid membranes. Biochim. Biophys. Acta Biomembr. 2012, 1818, 2563–2571. [Google Scholar] [CrossRef] [PubMed]
- The UniProt Consortium. UniProt: A worldwide hub of protein knowledge. Nucleic Acids Res. 2018, 47, D506–D515. [Google Scholar]
- Shi, Y.; Mowery, R.A.; Ashley, J.; Hentz, M.; Ramirez, A.J.; Bilgicer, B.; Slunt-Brown, H.; Borchelt, D.R.; Shaw, B.F. Abnormal SDS-PAGE migration of cytosolic proteins can identify domains and mechanisms that control surfactant binding. Protein Sci. 2012, 21, 1197–1209. [Google Scholar] [CrossRef]
- Kowalski, W.; Gizak, A.; Rakus, D. Phosphoglycerate mutase in mammalian striated muscles: Subcellular localization and binding partners. FEBS Lett. 2009, 583, 1841–1845. [Google Scholar] [CrossRef]
- Malheiros, J.M.; Braga, C.P.; Grove, R.A.; Ribeiro, F.A.; Calkins, C.R.; Adamec, J.; Chardulo, L.A.L. Influence of oxidative damage to proteins on meat tenderness using a proteomics approach. Meat Sci. 2019, 148, 64–71. [Google Scholar] [CrossRef]
- Archana, P.R.; Sejian, V.; Ruban, W.; Bagath, M.; Krishnan, G.; Aleena, J.; Manjunathareddy, G.B.; Beena, V.; Bhatta, R. Comparative assessment of heat stress induced changes in carcass traits, plasma leptin profile and skeletal muscle myostatin and HSP70 gene expression patterns between Indigenous Osmanabadi and Salem Black Goat Breeds. Meat Sci. 2018, 141, 66–80. [Google Scholar] [CrossRef]
- Bubb, M.R.; Govindasamy, L.; Yarmola, E.G.; Vorobiev, S.M.; Almo, S.C.; Somasundaram, T.; Chapman, M.S.; Agbandje-McKenna, M.; McKenna, R. Polylysine induces an antiparallel actin dimer that nucleates filament assembly: Crystal structure at 3.5-A resolution. J. Biol. Chem. 2002, 277, 20999–21006. [Google Scholar] [CrossRef] [PubMed]
- Contreras-Castillo, C.J.; Lomiwes, D.; Wu, G.; Frost, D.; Farouk, M.M. The effect of electrical stimulation on post mortem myofibrillar protein degradation and small heat shock protein kinetics in bull beef. Meat Sci. 2016, 113, 65–72. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Arner, A.; Puolanne, E.; Ertbjerg, P. On the water-holding of myofibrils: Effect of sarcoplasmic protein denaturation. Meat Sci. 2016, 119, 32–40. [Google Scholar] [CrossRef] [PubMed]
- Ackermann, M.A.; Kontrogianni-Konstantopoulos, A. Myosin Binding Protein-C: A regulator of actomyosin interaction in striated muscle. J. Biomed. Biotechnol. 2011, 2011, 1–9. [Google Scholar] [CrossRef]
- Nyström, T. Role of oxidative carbonylation in protein quality control and senescence. EMBO J. 2005, 24–27, 1311–1317. [Google Scholar] [CrossRef]
- Tornberg, E. Effects of heat on meat proteins—Implications on structure and quality of meat products. Meat Sci. 2005, 70, 493–508. [Google Scholar] [CrossRef]
- Bao, Y.; Boeren, S.; Ertbjerg, P. Myofibrillar protein oxidation affects filament charges, aggregation and water-holding. Meat Sci. 2018, 135, 102–108. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Assays | Concentration (µg·Kg−1) | SP | MP | Total Protein Loss 3 (%) | ||
|---|---|---|---|---|---|---|
| Amount of Protein 1 | Loss (%) 2 | Amount of Protein 1 | Loss (%) 2 | |||
| BLANK | --- | 58.7 ± 0.17 | --- | 2.6 ± 0.01 | --- | --- |
| Sarafloxacin | 50 | 42.5 ± 0.26 † | 27.6 ± 0.45 a | 2.3 ± 0.04 † | 10.8 ± 1.38 a | 26.9 |
| 100 | 39.1 ± 0.08 † | 33.4 ± 0.14 b | 2.4 ± 0.02 † | 9.4 ± 0.84 a | 32.4 | |
| 150 | 38.4 ± 0.54 † | 34.6 ± 0.92 b | 2.3 ± 0.00 † | 12.2 ± 0.00 a | 33.7 | |
| Danofloxacin | 100 | 40.8 ± 0.24 † | 30.5 ± 0.41 c | 2.3 ± 0.00 † | 11.1 ± 0.04 b | 29.6 |
| 200 | 44.4 ± 0.11 † | 24.3 ± 0.18 d | 2.3 ± 0.00 † | 10.7 ± 0.04 b | 23.8 | |
| 300 | 39.3 ± 0.26 † | 33.0 ± 0.45 e | 2.5 ± 0.01 † | 3.3 ± 0.27 c | 31.8 | |
| Enrofloxacin | 50 | 42.5 ± 0.41 † | 27.6 ± 0.69 f | 2.5 ± 0.01 † | 5.0 ± 0.27 d | 26.7 |
| 100 | 42.0 ± 0.11 † | 28.4 ± 0.18 f | 2.4 ± 0.01 † | 7.3 ± 0.19 e | 27.5 | |
| 150 | 38.9 ± 0.06 † | 33.8 ± 0.11 g | 2.2 ± 0.01 † | 16.5 ± 0.38 f | 33.0 | |
| Ciprofloxacin | 50 | 53.4 ± 1.73 | 0.43 ± 2.07 h | 2.6 ± 0.20 | 2.2 ± 7.62 g | 8.8 |
| 100 | 55.7 ± 0.74 | 9.1 ± 2.94 h | 2.7 ± 0.04 | 1.7 ± 1.36 g | 4.8 | |
| 150 | 38.9 ± 0.06 † | 5.1 ± 1.26 h | 2.7 ± 0.04 | −1.7 ± 1.36 g | 32.3 | |
| Norfloxacin | 50 | 57.7 ± 2.71 | 1.6 ± 4.62 i | 2.2 ± 0.07 | 15.9 ± 2.83 h | 2.2 |
| 100 | 57.1 ± 0.72 | 2.7 ± 1.23 i | 2.4 ± 0.01 | 9.2 ± 0.26 h | 3.0 | |
| 150 | 56.1 ± 1.99 | 4.3 ± 3.39 i | 2.5 ± 0.09 | 5.5 ± 3.46 h | 4.4 | |
| N° Band | Protein Name | Accession Number a | Biological Process b | MW (Da) | Score c | Sequence Coverage (%) | Sample |
|---|---|---|---|---|---|---|---|
| 1 | Pyruvate kinase isozymes M1/M2 | gi|329664500 | Glycolysis | 68,416 | 138 | 22 | dano-enro |
| 2 | Beta-enolase + Fructose-bisphosphate aldolase A | gi|77736349 + gi|156120479 | Glycolysis | 47,409 39,925 | 118 | 43 38 | dano-enro |
| 3 | Fructose-bisphosphate aldolase A | gi|156120479 | Glycolysis | 39,925 | 121 | 45 | all |
| 4 | Creatine kinase M | gi|4838363 | Oxidative stress | 43,172 | 257 | 40 | dano |
| 5 | Fructose-bisphosphate aldolase A | gi|156120479 | Glycolysis | 39,925 | 130 | 53 | all |
| 6 | Phosphoglycerate mutase 2 | gi|84000195 | Glycolysis | 28,838 | 84 | 50 | dano-enro |
| 7 | Triosephosphate isomerase | gi|61888856 | Glycolysis | 26,901 | 246 | 85 | dano-enro |
| N° Band | Protein Name | Accession Number a | Biological Process b | MW (Da) | Score c | Sequence Coverage (%) | Sample |
|---|---|---|---|---|---|---|---|
| 3 | Myosin-binding protein C | gi|160425243 | Muscle contraction | 134,923 | 111 | 20 | dano-enro |
| 4 | Phosphorylase glycogen | gi|154426116 | Glycogen metabolism | 97,683 | 463 | 53 | all |
| 5 | HSP 70 + +mCG5074, isoform CRA_a | gi|261825070 | Oxidative stress | 45,299 35,231 | 146 | 50 40 | all |
| 7 | Pyruvate kinase | gi|329664500 | Glycolysis | 68,416 | 151 | 39 | dano-enro |
| 8 | Actin-Bound Wh2 Domains + Beta-enolase | gi|297343122 + gi|77736349 | Actin filament nucleation + Glycolisis | 40,304 47,409 | 127 | 55 38 | all |
| 9 | Creatine kinase M + Antiparallel Actin Dimer | gi|4838363 + gi|20664362 | Oxidative stress + Actin filament nucleation | 43,172 41,558 | 232 | 58-52 | all |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Márquez-Lázaro, J.; Méndez-Cuadro, D.; Rodríguez-Cavallo, E. Residues of Fluoroquinolone Antibiotics Induce Carbonylation and Reduce In Vitro Digestion of Sarcoplasmic and Myofibrillar Beef Proteins. Foods 2020, 9, 170. https://doi.org/10.3390/foods9020170
Márquez-Lázaro J, Méndez-Cuadro D, Rodríguez-Cavallo E. Residues of Fluoroquinolone Antibiotics Induce Carbonylation and Reduce In Vitro Digestion of Sarcoplasmic and Myofibrillar Beef Proteins. Foods. 2020; 9(2):170. https://doi.org/10.3390/foods9020170
Chicago/Turabian StyleMárquez-Lázaro, Johana, Darío Méndez-Cuadro, and Erika Rodríguez-Cavallo. 2020. "Residues of Fluoroquinolone Antibiotics Induce Carbonylation and Reduce In Vitro Digestion of Sarcoplasmic and Myofibrillar Beef Proteins" Foods 9, no. 2: 170. https://doi.org/10.3390/foods9020170
APA StyleMárquez-Lázaro, J., Méndez-Cuadro, D., & Rodríguez-Cavallo, E. (2020). Residues of Fluoroquinolone Antibiotics Induce Carbonylation and Reduce In Vitro Digestion of Sarcoplasmic and Myofibrillar Beef Proteins. Foods, 9(2), 170. https://doi.org/10.3390/foods9020170