An Overview on Cyclic Fatty Acids as Biomarkers of Quality and Authenticity in the Meat Sector


Abstract
1. Introduction
2. Materials and Methods
2.1. Chemicals
2.2. Sampling
- ‒
- Batch 1: commercial meat samples.
- ‒
- Batch 2: certified meat samples for the feeding system.
2.3. Fat Extraction
2.4. GC-MS Analysis
2.5. 1H NMR Analysis
2.6. Data Analysis
3. Results
3.1. CPFAs Determination in Meat Fat
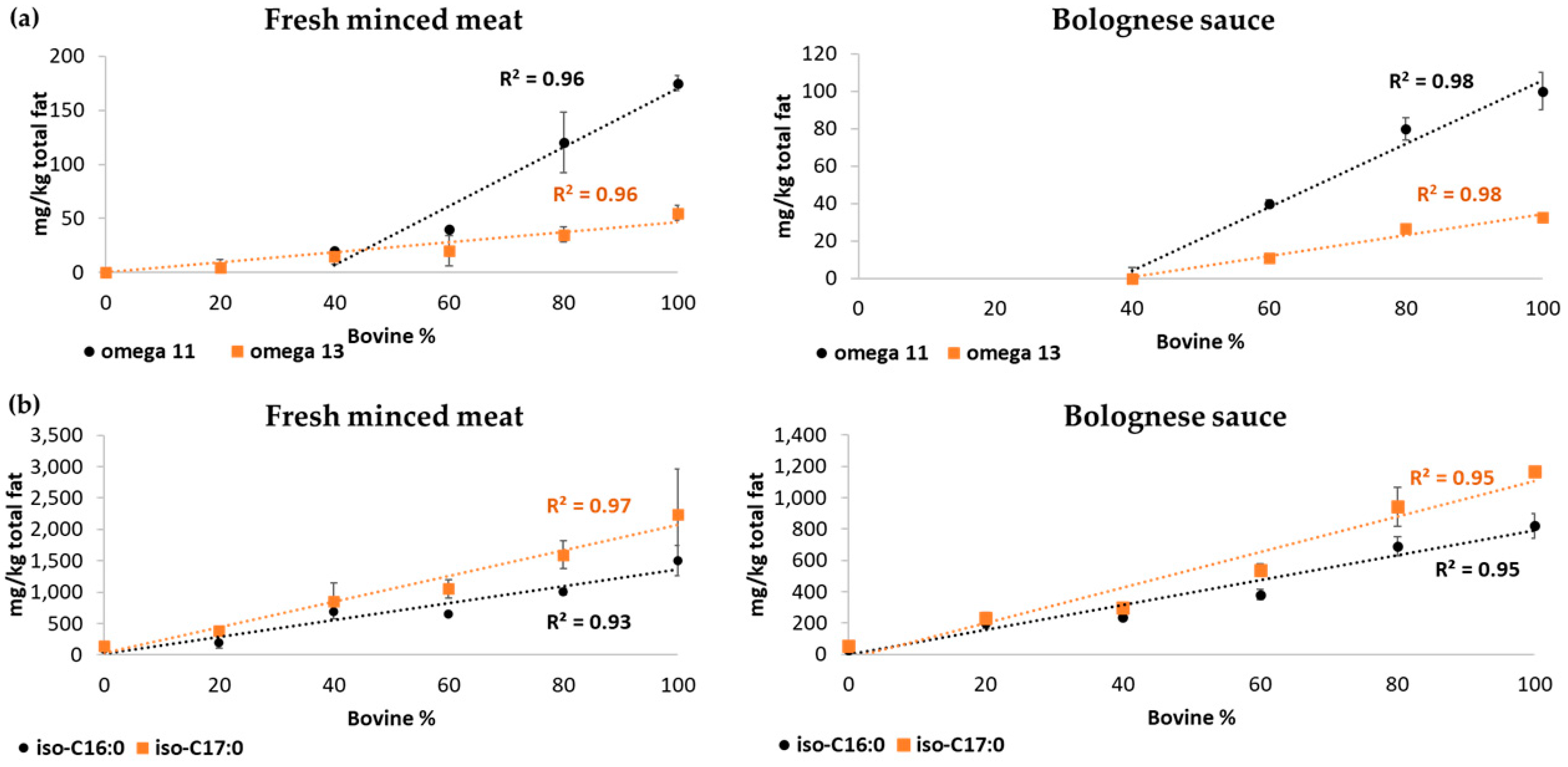
3.2. Omega-Cyclohexyl Fatty Acids Determination in Meat Fat
4. Discussion
4.1. Application of CPFAs in Meat as Markers of Animal Feed Regimen
4.2. Omega-Cyclohexyl Fatty Acids: Species Identification and Quantitation of the Ratio of Beef to Pork in Minced Meat
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Fajardo, V.; González, I.; Martín, I.; Rojas, M.; García, T.; Martín, R. A review of current PCR-based methodologies for the authentication of meats from game animal species. Trends Food Sci. Technol. 2010, 21, 408–421. [Google Scholar] [CrossRef]
- Gvozdanović, K.; Kušec, I.D.; Margeta, P.; Salajpal, K.; Džijan, S.; Bošnjak, Z.; Kušec, G. Multiallelic marker system for traceability of Black Slavonian pig meat. Food Control. 2020, 109, 106917. [Google Scholar] [CrossRef]
- Ballin, N.Z. Authentication of meat and meat products. Meat Sci. 2010, 86, 577–587. [Google Scholar] [CrossRef]
- Mamani-Linares, L.W.; Gallo, C.; Alomar, D. Identification of cattle, llama and horse meat by near infrared reflectance or transflectance spectroscopy. Meat Sci. 2012, 90, 378–385. [Google Scholar] [CrossRef] [PubMed]
- Watson, A.D.; Gunning, Y.; Rigby, N.M.; Philo, M.; Kemsley, E.K. Meat authentication via multiple reaction monitoring mass spectrometry of myoglobin peptides. Anal. Chem. 2015, 87, 10315–10322. [Google Scholar] [CrossRef]
- Kim, J.S.; Hwang, I.M.; Lee, G.H.; Park, Y.M.; Choi, J.Y.; Jamila, N.; Khan, N.; Kim, K.S. Geographical origin authentication of pork using multi-element and multivariate data analyses. Meat Sci. 2017, 123, 13–20. [Google Scholar] [CrossRef]
- Zhao, J.; Chen, A.; You, X.; Xu, Z.; Zhao, Y.; He, W.; Zhao, L.; Yang, S. A panel of SNP markers for meat traceability of Halal beef in the Chinese market. Food Control 2018, 87, 94–99. [Google Scholar] [CrossRef]
- Ballin, N.Z.; Vogensen, F.K.; Karlsson, A.H. Species determination—Can we detect and quantify meat adulteration? Meat Sci. 2009, 83, 165–174. [Google Scholar] [CrossRef]
- Sentandreu, M.A.; Sentandreu, E. Peptide biomarkers as a way to determine meat authenticity. Meat Sci. 2011, 89, 280–285. [Google Scholar] [CrossRef]
- Wood, J.D.; Enser, M.; Fisher, A.V.; Nute, G.R.; Sheard, P.R.; Richardson, R.I.; Hughes, S.I.; Whittington, F.M. Fat deposition, fatty acid composition and meat quality: A review. Meat Sci. 2008, 78, 343–358. [Google Scholar] [CrossRef]
- da Silva Martins, T.; de Lemos, M.V.; Mueller, L.F.; Baldi, F.; de Amorim, T.Y.; Ferinho, A.M.; Muñoz, J.A.; de Souza Fuzikawa, I.H.; de Mouray, G.V.; Gemelli, J.L.; et al. Fat deposition, fatty acid composition, and its relationship with meat quality and human health. In Meat Science and Nutrition; Chapter 2; Arshad, M.S., Ed.; Intechopen: London, UK, 2018; pp. 17–37. [Google Scholar] [CrossRef]
- Kagawa, K.; Matsutaka, H.; Fukuhama, C.; Watanabe, Y.; Fujino, H. Globin digest, acidic protease hydrolysate, inhibits dietary hypertriglyceridemia and Val-Val-Tyr-Pro, one of its constituents, possesses most superior effect. Life Sci. 1996, 58, 1745–1755. [Google Scholar] [CrossRef]
- Szabó, A.; Fébel, H.; Sugár, L.; Romvári, R. Fatty acid regiodistribution analysis of divergent animal triacylglycerol samples—A possible approach for species differentiation. J. Food Lipids 2007, 14, 62–77. [Google Scholar] [CrossRef]
- Warren, H.E.; Scollan, N.D.; Enser, M.; Hughes, S.I.; Richardson, R.I.; Wood, J.D. Effects of breed and a concentrate or grass silage diet on beef quality in cattle of 3 ages. I: Animal performance, carcass quality and muscle fatty acid composition. Meat Sci. 2007, 78, 256–269. [Google Scholar] [CrossRef]
- Monahan, F.J.; Schmidt, O.; Moloney, A.P. Meat provenance: Authentication of geographical origin and dietary background of meat. Meat Sci. 2018, 144, 2–14. [Google Scholar] [CrossRef] [PubMed]
- Moran, L.; Wilson, S.S.; McElhinney, C.K.; Monahan, F.J.; McGee, M.; O’Sullivan, M.G.; O’Riordan, E.G.; Kerry, J.P.; Moloney, A.P. Suckler bulls slaughtered at 15 months of age: Effect of different production systems on the fatty acid profile and selected quality characteristics of Longissimus thoracis. Foods 2019, 8, 264. [Google Scholar] [CrossRef] [PubMed]
- Alfaia, C.P.M.; Alves, S.P.; Martins, S.I.V.; Costa, A.S.H.; Fontes, C.M.G.A.; Lemos, J.P.C.; Bessa, R.J.B.; Prates, J.A.M. Effect of the feeding system on intramuscular fatty acids and conjugated linoleic acid isomers of beef cattle, with emphasis on their nutritional value and discriminatory ability. Food Chem. 2009, 114, 939–946. [Google Scholar] [CrossRef]
- Marseglia, A.; Caligiani, A.; Comino, L.; Righi, F.; Quarantelli, A.; Palla, G. Cyclopropyl and ω-cyclohexyl fatty acids as quality markers of cow milk and cheese. Food Chem. 2013, 140, 711–716. [Google Scholar] [CrossRef] [PubMed]
- Caligiani, A.; Marseglia, A.; Palla, G. An overview of the presence of cyclopropane fatty acids in milk and dairy products. J. Agric. Food Chem. 2014, 62, 7828–7832. [Google Scholar] [CrossRef]
- Lolli, V.; Dall’Asta, M.; Del Rio, D.; Palla, G.; Caligiani, A. Presence of cyclopropane fatty acids in foods and estimation of dietary intake in the Italian population. Int. J. Food Sci. Nutr. 2019, 70, 467–473. [Google Scholar] [CrossRef]
- Caligiani, A.; Nocetti, M.; Lolli, V.; Marseglia, A.; Palla, G. Development of a quantitative GC−MS method for the detection of cyclopropane fatty acids in cheese as new molecular markers for Parmigiano Reggiano authentication. J. Agric. Food Chem. 2016, 64, 4158–4164. [Google Scholar] [CrossRef]
- Lolli, V.; Marseglia, A.; Palla, G.; Zanardi, E.; Caligiani, A. Determination of cyclopropane fatty acids in food of animal origin by 1HNMR. J. Anal. Methods Chem. 2018, 2018, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Caligiani, A.; Lolli, V. Cyclic fatty acids in food: An under investigated class of fatty acids. In Biochemistry and Health Benefits of Fatty Acids; Chapter 3; Waisundara, V., Ed.; IntechOpen: Rijeka, Croatia, 2018; Volume 2018. [Google Scholar] [CrossRef]
- Fievez, V.; Colman, E.; Montoya, J.C.; Stefanov, I.; Vlaemink, B. Milk odd- and branched- chain fatty acids as biomarkers of rumen function—An update. Anim. Feed Sci. Technol. 2012, 172, 51–65. [Google Scholar] [CrossRef]
- Prandi, B.; Lambertini, F.; Faccini, A.; Suman, M.; Leporati, A.; Tedeschi, T.; Sforza, S. Mass spectrometry quantification of beef and pork meat in highly processed food: Application on Bolognese sauce. Food Control 2017, 74, 61–69. [Google Scholar] [CrossRef]
- Folch, J.; Less, M.; Sloane, G.H. A simple method for the isolation and purification of total lipids from animal tissues. J. Biol. Chem. 1957, 226, 497–509. [Google Scholar]
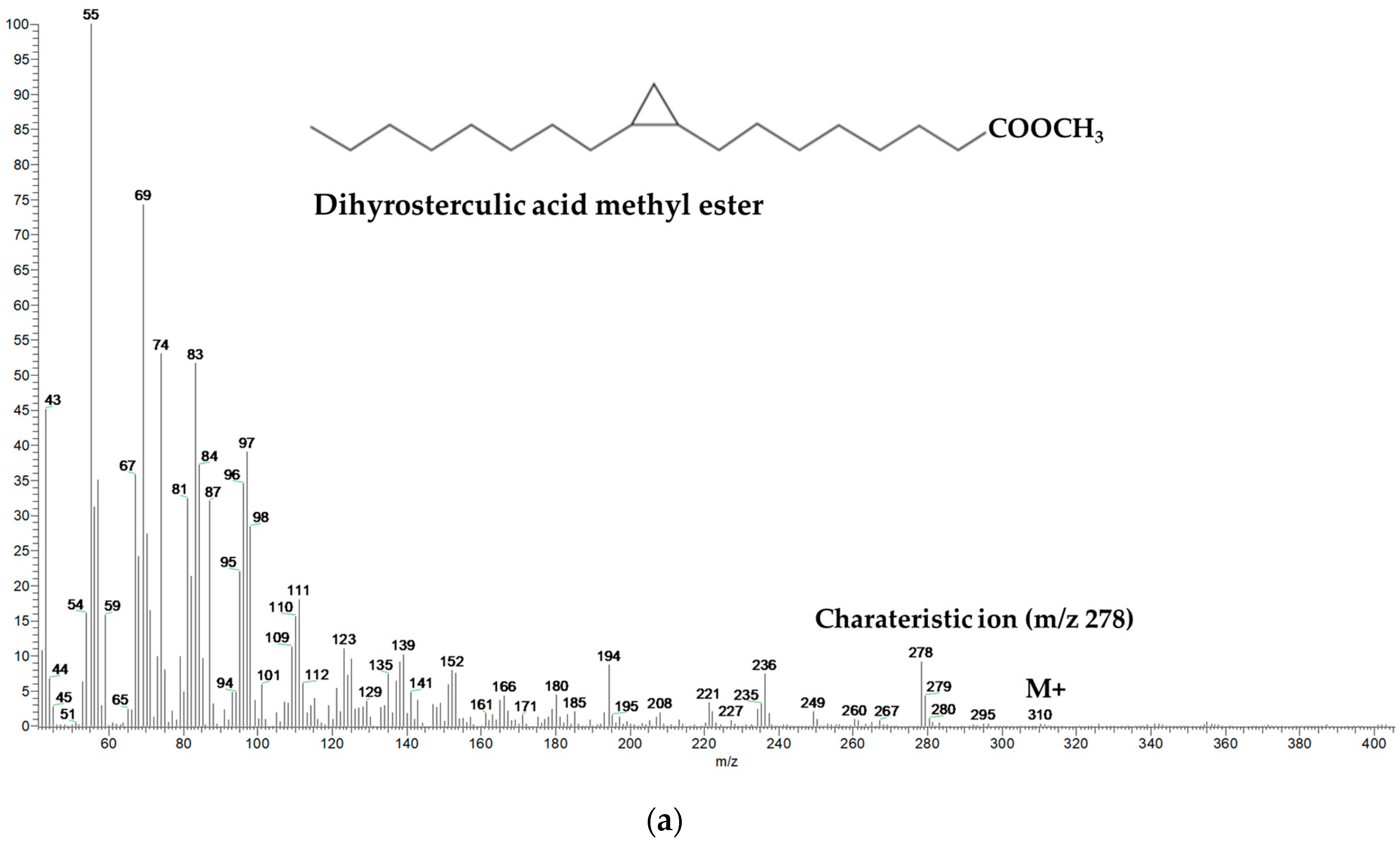
- Knothe, G. NMR characterization of dihydrosterculic acid and its methylesters. Lipids 2006, 41, 393–396. [Google Scholar] [CrossRef]
- Russo, C.; Fronte, B.; Preziuso, G. Meat quality of three different muscles of Chianina beef fed with maize silage. LAR 2012, 18, 241–244. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
| Samples | No. Positive 2 for CPFAs | Mean ± SD (mg/kg fat) | Range (mg/kg fat) |
|---|---|---|---|
| Batch 1 | |||
| Commercial beef meat 3 | 18/18 | 200 ± 100 | 70–410 |
| Other meat products 4 | 0/32 | <LOD 5 | <LOD 5 |
| Batch 2 | |||
| Longissimus dorsi6 | 2/12 | 240 ± 57 | 200–280 |
| G0 7 | 2/15 | 164 ± 23 | 147–180 |
| GC 8 | 7/15 | 158 ± 75 | 79–268 |
| SC 9 | 15/15 | 240 ± 120 | 108–465 |
| Pearson’s r | ||
|---|---|---|
| Fatty Acids | % Bovine (Raw Meat) 1 | % Bovine (Bolognese Sauce) 1 |
| ω-11 CHFA | 0.93 | 0.95 |
| ω-13 CHFA | 0.97 | 0.93 |
| iso-C16:0 | 0.97 | 0.98 |
| iso-C17:0 | 0.98 | 0.98 |
| iso-C16 2 | iso-C17 3 | |||||
|---|---|---|---|---|---|---|
| Test Sample | Real 3 Beef % | Real 3 Pork % | Calculated Beef % | Calculated Pork 4 % | Calculated Beef % | Calculated Pork 4 % |
| A | 35 | 65 | 30.9 ± 3.1 (88%) | 69.1 ± 3.1 (106%) | 30.2 ± 2.3 (86%) | 69.8 ± 2.3 (107%) |
| B | 75 | 25 | 74.1 ± 0.2 (99%) | 25.9 ± 0.2 (104%) | 70.4 ± 2.1 (94%) | 29.6 ± 2.1 (118%) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lolli, V.; Zanardi, E.; Moloney, A.P.; Caligiani, A. An Overview on Cyclic Fatty Acids as Biomarkers of Quality and Authenticity in the Meat Sector. Foods 2020, 9, 1756. https://doi.org/10.3390/foods9121756
Lolli V, Zanardi E, Moloney AP, Caligiani A. An Overview on Cyclic Fatty Acids as Biomarkers of Quality and Authenticity in the Meat Sector. Foods. 2020; 9(12):1756. https://doi.org/10.3390/foods9121756
Chicago/Turabian StyleLolli, Veronica, Emanuela Zanardi, Aidan P. Moloney, and Augusta Caligiani. 2020. "An Overview on Cyclic Fatty Acids as Biomarkers of Quality and Authenticity in the Meat Sector" Foods 9, no. 12: 1756. https://doi.org/10.3390/foods9121756
APA StyleLolli, V., Zanardi, E., Moloney, A. P., & Caligiani, A. (2020). An Overview on Cyclic Fatty Acids as Biomarkers of Quality and Authenticity in the Meat Sector. Foods, 9(12), 1756. https://doi.org/10.3390/foods9121756