Development and Evaluation of qPCR Detection Method and Zn-MgO/Alginate Active Packaging for Controlling Listeria monocytogenes Contamination in Cold-Smoked Salmon




Abstract
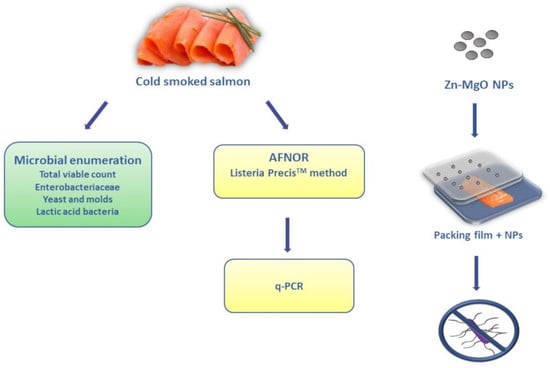
1. Introduction
2. Materials and Methods
2.1. Materials and Reagents
2.2. Primer Design
2.3. PCR and qPCR Analyses
2.4. Plate Count Analyses of Cold-Smoked Salmon Samples
2.5. Artificially Contamination (AC) of Cold-Smoked Samples
2.6. Listeria monocytogenes Inhibition Test Using Zn-MgO Nanoparticles
2.7. Cytotoxicity Test on Mammalian Cell Cultures and FACS Measurements
2.8. Active Film Packaging Formation and Challenge Test Protocol
3. Results and Discussion
3.1. Plate Count Results on Commercial CSS RTE CCS and Artificially Contaminated CSS Samples
3.2. Specificity and Sensitivity of the Primers With PCR and qPCR
3.3. Listeria monocytogenes Plate Inhibition Test
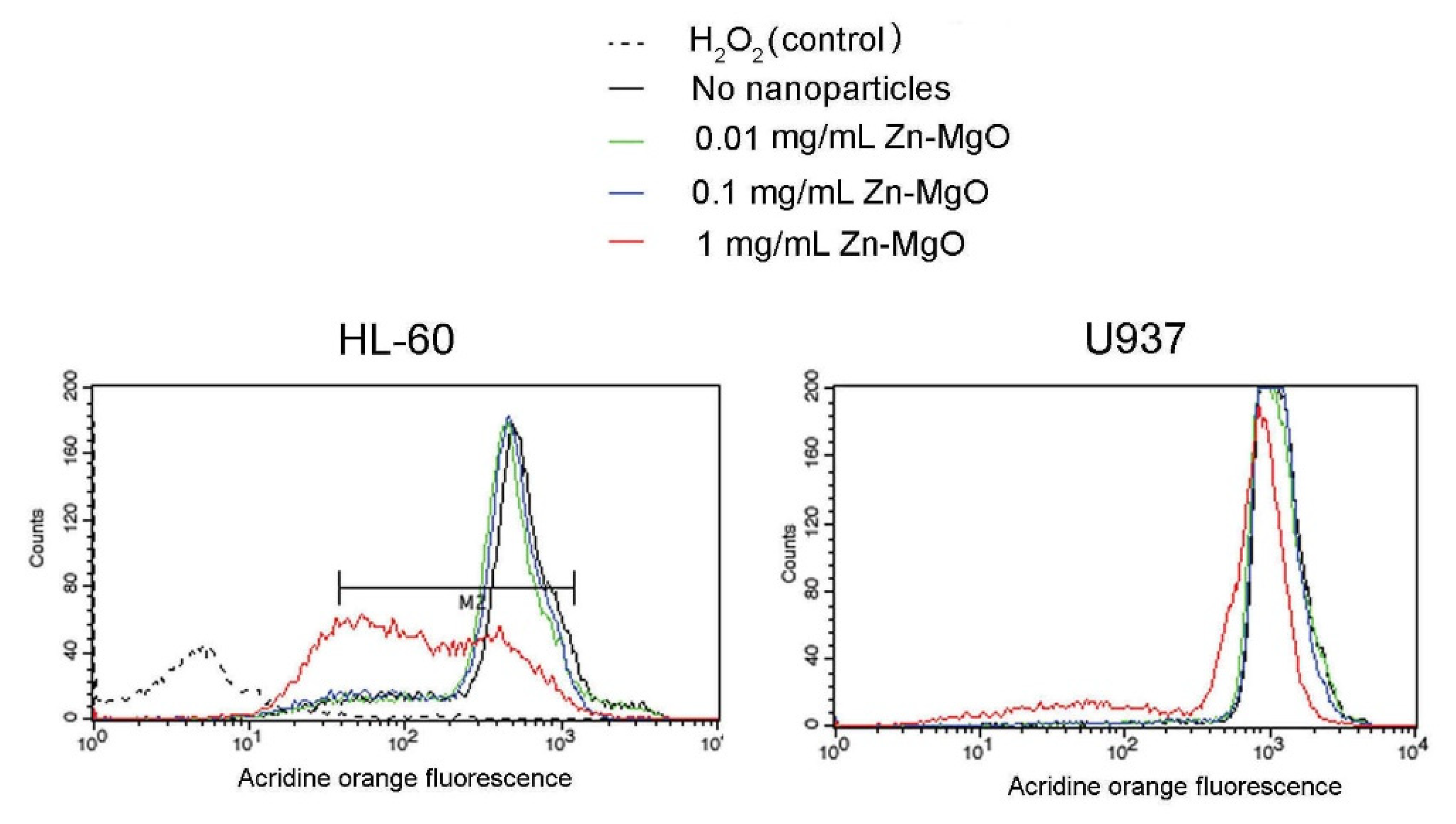
3.4. Zn-MgO Cytotoxicity
3.5. Challenge Test with Zn-MgO Alginate-Nanobiocomposite Active Film
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Auvolat, A.; Besse, N.G. The challenge of enumerating Listeria monocytogenes in food. Food Microbiol. 2016, 53, 135–149. [Google Scholar] [CrossRef]
- Roberts, A.; Williams, S.; Wiedmann, M.; Nightingale, K. Some Listeria monocytogenes outbreak strains demonstrate significantly reduced invasion, inlA transcript levels, and swarming motility in vitro. Appl. Environ. Microbiol. 2009, 75, 5647–5658. [Google Scholar] [CrossRef]
- Swaminathan, B.; Gerner-Smidt, P. The epidemiology of human listeriosis. Microbes Infect. 2007, 9, 1236–1243. [Google Scholar] [CrossRef]
- Gandhi, M.; Chikindas, M.L. Listeria: A foodborne pathogen that knows how to survive. Int. J. Food Microbiol. 2007, 113, 1–15. [Google Scholar] [CrossRef]
- Liu, D.; Lawrence, M.L.; Ainsworth, A.J.; Austin, F.W. Comparative assessment of acid, alkali and salt tolerance in Listeria monocytogenes virulent and avirulent strains. FEMS Microbiol. Lett. 2005, 243, 373–378. [Google Scholar] [CrossRef] [PubMed]
- European Food Safety Authority; European Centre for Disease Prevention and Control. The European Union Summary REPORT on trends and sources of zoonoses, zoonotic agents and food-borne outbreaks in 2012. EFSA J. 2014, 12, 3547. [Google Scholar] [CrossRef]
- European Food Safety Authority; European Centre for Disease Prevention and Control. The European Union summary report on trends and sources of zoonoses, zoonotic agents and food-borne outbreaks in 2014. EFSA J. 2015, 13, 4329. [Google Scholar]
- Jemmi, T.; Stephan, R. Listeria monocytogenes: Food-borne pathogen and hygiene indicator. Rev. Sci. Tech. 2006, 25, 571–580. [Google Scholar] [CrossRef]
- Katzav, M.; Hyvoenen, P.; Muje, P.; Rantala, L.; von Wright, A. Pulsed-field gel electrophoresis typing of Listeria monocytogenes isolated in two Finnish fish farms. J. Food Protect. 2006, 69, 1443–1447. [Google Scholar] [CrossRef]
- Garrido-Maestu, A.; Azinheiro, S.; Carvalho, J.; Prado, M. Rapid and sensitive detection of viable Listeria monocytogenes in food products by a filtration-based protocol and qPCR. Food Microbiol. 2018, 73, 254–263. [Google Scholar]
- Välimaa, A.L.; Tilsala-Timisjärvi, A.; Virtanen, E. Rapid detection and identification methods for Listeria monocytogenes in the food chain—A review. Food Control 2015, 55, 103–114. [Google Scholar]
- ISO. Microbiology of the Food Chain-Horizontal Method for the Detection and Enumeration of Listeria monocytogenes and of Listeria spp.-Part 2: Enumeration Method; ISO: Geneva, Switzerland, 2014. [Google Scholar]
- ISO. Microbiology of the Food Chain-Horizontal Method for the Enumeration of Microorganisms–Part 1: Colony Count at 30 Degrees C by the Pour Plate Technique; ISO: Geneva, Switzerland, 2013. [Google Scholar]
- Vizzini, P.; Braidot, M.; Vidic, J.; Manzano, M. Electrochemical and optical biosensors for the detection of campylobacter and listeria: An update look. Micromachines 2019, 10, 500. [Google Scholar]
- Manzano, M.; Cocolin, L.; Cantoni, C.; Comi, G. Detection and identification ofListeria monocytogenes from milk and cheese by a single-step PCR. Mol. Biotechnol. 1997, 7, 85–88. [Google Scholar] [PubMed]
- Omiccioli, E.; Amagliani, G.; Brandi, G.; Magnani, M. A new platform for Real-Time PCR detection of Salmonella spp., Listeria monocytogenes and Escherichia coli O157 in milk. Food Microbiol. 2009, 26, 615–622. [Google Scholar] [PubMed]
- Vidic, J.; Manzano, M.; Chang, C.M.; Jaffrezic-Renault, N. Advanced biosensors for detection of pathogens related to livestock and poultry. Vet. Res. 2017, 48, 1–22. [Google Scholar]
- Vidic, J.; Vizzini, P.; Manzano, M.; Kavanaugh, D.; Ramarao, N.; Zivkovic, M.; Radonic, V.; Knezevic, N.; Giouroudi, I.; Gadjanski, I. Point-of-need DNA testing for detection of foodborne pathogenic bacteria. Sensors 2019, 19, 1100. [Google Scholar]
- Wang, Y.; Salazar, J.K. Culture-independent rapid detection methods for bacterial pathogens and toxins in food matrices. Compr. Rev. Food Sci. Food Saf. 2016, 15, 183–205. [Google Scholar]
- Quintavalla, S.; Vicini, L. Antimicrobial food packaging in meat industry. Meat Sci. 2002, 62, 373–380. [Google Scholar]
- Stankic, S.; Suman, S.; Haque, F.; Vidic, J. Pure and multi metal oxide nanoparticles: Synthesis, antibacterial and cytotoxic properties. J. Nanobiotechnol. 2016, 14, 1–20. [Google Scholar]
- Vermeiren, L.; Devlieghere, F.; Debevere, J. Effectiveness of some recent antimicrobial packaging concepts. Food Addit. Contam. 2002, 19, 163–171. [Google Scholar]
- Jones, K.E.; Patel, N.G.; Levy, M.A.; Storeygard, A.; Balk, D.; Gittleman, J.L.; Daszak, P. Global trends in emerging infectious diseases. Nature 2008, 451, 990–993. [Google Scholar]
- Jin, T.; Sun, D.; Su, J.; Zhang, H.; Sue, H.J. Antimicrobial efficacy of zinc oxide quantum dots against Listeria monocytogenes, Salmonella enteritidis, and Escherichia coli O157: H7. J. Food Sci. 2009, 74, M46–M52. [Google Scholar] [PubMed]
- Martínez-Carmona, M.; Gun’ko, Y.; Vallet-Regí, M. ZnO nanostructures for drug delivery and theranostic applications. Nanomaterials 2018, 8, 268. [Google Scholar]
- Vidic, J.; Stankic, S.; Haque, F.; Ciric, D.; Le Goffic, R.; Vidy, A.; Jupille, J.; Delmas, B. Selective antibacterial effects of mixed ZnMgO nanoparticles. J. Nanoparticle Res. 2013, 15, 1595. [Google Scholar]
- Auger, S.; Henry, C.; Péchaux, C.; Lejal, N.; Zanet, V.; Nikolic, M.V.; Manzano, M.; Vidic, J. Exploring the impact of Mg-doped ZnO nanoparticles on a model soil microorganism Bacillus subtilis. Ecotoxicol. Environ. Safety 2019, 182, 109421. [Google Scholar]
- Auger, S.; Henry, C.; Péchoux, C.; Suman, S.; Lejal, N.; Bertho, N.; Larcher, T.; Stankic, S.; Vidic, J. Exploring multiple effects of Zn 0.15 Mg 0.85 O nanoparticles on Bacillus subtilis and macrophages. Sci. Rep. 2018, 8, 1–14. [Google Scholar]
- Stankic, S.; Müller, M.; Diwald, O.; Sterrer, M.; Knözinger, E.; Bernardi, J. Size-dependent optical properties of MgO nanocubes. Angew. Chem. Int. Ed. 2005, 44, 4917–4920. [Google Scholar]
- Corpet, F. Multiple sequence alignment with hierarchical clustering. Nucleic Acids Res. 1988, 16, 10881–10890. [Google Scholar]
- Intergated DNA Technologies. Available online: http://eu.idtdna.com/calc/analyzer (accessed on 25 August 2020).
- AmplifX: Manage, Test and Design Your Primers for PCR. Available online: http://crn2m.univ-mrs.fr/pub/amplifx-dist (accessed on 25 August 2020).
- Comi, G.; Cocolin, L.; Cantoni, C.; Manzano, M. A RE-PCR method to distinguish Listeria monocytogenes serovars. FEMS Immunol. Med. Microbiol. 1997, 18, 99–104. [Google Scholar]
- Rohde, A.; Hammerl, J.A.; Boone, I.; Jansen, W.; Fohler, S.; Klein, G.; Dieckmann, R.; Al Dahouk, S. Overview of validated alternative methods for the detection of foodborne bacterial pathogens. Trends Food Sci. Technol. 2017, 62, 113–118. [Google Scholar]
- Cecchini, F.; Manzano, M.; Mandabi, Y.; Perelman, E.; Marks, R.S. Chemiluminescent DNA optical fibre sensor for Brettanomyces bruxellensis detection. J. Biotechnol. 2012, 157, 25–30. [Google Scholar] [CrossRef] [PubMed]
- Balouiri, M.; Sadiki, M.; Ibnsouda, S.K. Methods for in vitro evaluating antimicrobial activity: A review. J. Pharm. Analysis 2016, 6, 71–79. [Google Scholar] [CrossRef] [PubMed]
- Vidic, J.; Richard, C.A.; Péchoux, C.; da Costa, B.; Bertho, N.; Mazerat, S.; Delmas, B.; Chevalier, C. Amyloid assemblies of influenza A virus PB1-F2 protein damage membrane and induce cytotoxicity. J. Biol. Chem. 2016, 291, 739–751. [Google Scholar] [CrossRef] [PubMed]
- Akbar, A.; Anal, A.K. Zinc oxide nanoparticles loaded active packaging, a challenge study against Salmonella typhimurium and Staphylococcus aureus in ready-to-eat poultry meat. Food Control 2014, 38, 88–95. [Google Scholar] [CrossRef]
- Gimenez, B.; Dalgaard, P. Modelling and predicting the simultaneous growth of Listeria monocytogenes and spoilage micro-organisms in cold-smoked salmon. J. Appl. Microbiol. 2004, 96, 96–109. [Google Scholar] [CrossRef]
- Paludan-Müller, C.; Dalgaard, P.; Huss, H.H.; Gram, L. Evaluation of the role of Carnobacterium piscicola in spoilage of vacuum-and modified-atmosphere-packed cold-smoked salmon stored at 5 °C. Int. J. Food Microbiol. 1998, 39, 155–166. [Google Scholar] [CrossRef]
- Commission, E. Commission Regulation (EC) No 2073/2005 of 15 November 2005 on microbiological criteria for foodstuffs. Off. J. Eur. Union L 2005, 338, 1–26. [Google Scholar]
- Pafundo, S.; Gullì, M.; Marmiroli, N. Multiplex real-time PCR using SYBR® GreenER™ for the detection of DNA allergens in food. Anal. Bioanal. Chem. 2010, 396, 1831–1839. [Google Scholar] [CrossRef]
- Vieira, P.M.; Coelho, A.S.G.; Steindorff, A.S.; de Siqueira, S.J.L.; do Nascimento Silva, R.; Ulhoa, C.J. Identification of differentially expressed genes from Trichoderma harzianum during growth on cell wall of Fusarium solanias a tool for biotechnological application. BMC Genom. 2013, 14, 177. [Google Scholar] [CrossRef]
- Bonilla, J.; Atarés, L.; Vargas, M.; Chiralt, A. Edible films and coatings to prevent the detrimental effect of oxygen on food quality: Possibilities and limitations. J. Food Eng. 2012, 110, 208–213. [Google Scholar] [CrossRef]
- Fu, Y.; Sarkar, P.; Bhunia, A.K.; Yao, Y. Delivery systems of antimicrobial compounds to food. Trends Food Sci. Technol. 2016, 57, 165–177. [Google Scholar]
- Ture, H.; Eroglu, E.; Ozen, B.; Soyer, F. Effect of biopolymers containing natamycin against Aspergillus niger and Penicillium roquefortii on fresh kashar cheese. Int. J. Food Sci. Technol. 2011, 46, 154–160. [Google Scholar]
- Bolognesi, C.; Castoldi, A.F.; Crebelli, R.; Barthélémy, E.; Maurici, D.; Wölfle, D.; Volk, K.; Castle, L. Genotoxicity testing approaches for the safety assessment of substances used in food contact materials prior to their authorization in the European Union. Environ. Mol. Mutagen. 2017, 58, 361–374. [Google Scholar] [CrossRef] [PubMed]
- Cierech, M.; Wojnarowicz, J.; Kolenda, A.; Krawczyk-Balska, A.; Prochwicz, E.; Woźniak, B.; Łojkowski, W.; Mierzwińska-Nastalska, E. Zinc oxide nanoparticles cytotoxicity and release from newly formed PMMA–ZnO nanocomposites designed for denture bases. Nanomaterials 2019, 9, 1318. [Google Scholar] [CrossRef]
- Liu, Y.J.; He, L.L.; Mustapha, A.; Li, H.; Hu, Z.; Lin, M.S. Antibacterial activities of zinc oxide nanoparticles against Escherichia coli O157: H7. J. Appl. Microbiol. 2009, 107, 1193–1201. [Google Scholar] [CrossRef] [PubMed]
- Vasiljevic, Z.Z.; Dojcinovic, M.P.; Krstic, J.B.; Ribic, V.; Tadic, N.B.; Ognjanovic, M.; Auger, S.; Vidic, J.; Nikolic, M.V. Synthesis and antibacterial activity of iron manganite (FeMnO3) particles against the environmental bacterium Bacillus subtilis. RSC Adv. 2020, 10, 13879–13888. [Google Scholar] [CrossRef]
- Benabbou, R.; Subirade, M.; Desbiens, M.; Fliss, I. The impact of chitosan-divergicin film on growth of Listeria monocytogenes in cold-smoked salmon. Front. Microbiol. 2018, 9, 2824. [Google Scholar] [CrossRef]
- Marcos, B.; Aymerich, T.; Monfort, J.M.; Garriga, M. Use of antimicrobial biodegradable packaging to control Listeria monocytogenes during storage of cooked ham. Int. J. Food Microbiol. 2007, 120, 152–158. [Google Scholar] [CrossRef]
- Beaufort, A.; Bergis, H.; Lardeux, A.; Lombard, B. EURL Lm TECHNICAL GUIDANCE DOCUMENT for conducting shelf-life studies on Listeria monocytogenes in ready-to-eat foods. Appl. Environ. Microbiol. 2014, 74, 594–604. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Microorganism | Reference Code | |
|---|---|---|
| Positive controls | Listeria monocytogenes 1/2c | * ATCC 7644 |
| Listeria monocytogenes 1/2a | ° DISTAM | |
| Listeria monocytogenes 1/2b | ° DISTAM | |
| Listeria monocytogenes 4b | # DSMZ 15675 | |
| Negative controls | Listeria innocua | # DSMZ 20649 |
| Listeria ivanovii | § DI4A | |
| Salmonella enterica | # DSMZ 9145 | |
| Enterobacter spp. | § DI4A | |
| Escherichia coli | § DI4A | |
| Bacillus cereus | # DSMZ 2301 | |
| Campylobacter jejuni | # DSMZ 49943 | |
| Lactobacillus plantarum | * ATCC BAA-793 | |
| Lactobacillus rhamnosus | * ATCC 53103 | |
| Lactobacillus paracasei | # DSMZ 5622 | |
| Lactobacillus brevis | # DSMZ 20054 | |
| Bacillus subtilis | # DSMZ 4181 |
| Samples | Total Viable | Lactic Acid | Enterobacteriaceae | Yeasts |
|---|---|---|---|---|
| Count | Bacteria | |||
| CSS1t0 | 6.25 | 4.94 | <0.7 | <1.40 |
| CSS2t0 | 5.38 | 4.94 | <4.34 | 4.59 |
| CSS3t0 | 5.78 | <4.34 | 1.65 | 2.89 |
| CSS4t0 | 4.40 | <4.34 | <0.7 | 1.57 |
| CSS5t0 | 6.08 | 4.80 | 3.65 | 3.25 |
| CSS6t0 | 5.23 | 4.87 | <0.7 | 1.40 |
| CSS7t0 | 8.11 | 6.98 | <0.7 | 4.77 |
| CSS8t0 | 5.86 | 5 | <0.7 | 4.50 |
| CSS1t28 | 5.38 | <4.40 | <0.7 | 2.84 |
| CSS2t28 | 7.84 | 7.18 | 4.30 | 5.99 |
| CSS3t28 | 5.91 | 4.87 | <0.7 | 3.84 |
| CSS4t28 | 6.50 | 5.95 | <0.7 | 5.75 |
| CSS5t28 | 6.23 | <4.38 | 1.72 | 2.41 |
| CSS6t28 | 6.11 | 5.96 | 2.36 | <1.40 |
| CSS7t28 | 8.25 | 7.72 | <0.7 | 5.18 |
| CSS8t28 | 6.08 | <4.34 | <0.7 | 4.76 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vizzini, P.; Beltrame, E.; Zanet, V.; Vidic, J.; Manzano, M. Development and Evaluation of qPCR Detection Method and Zn-MgO/Alginate Active Packaging for Controlling Listeria monocytogenes Contamination in Cold-Smoked Salmon. Foods 2020, 9, 1353. https://doi.org/10.3390/foods9101353
Vizzini P, Beltrame E, Zanet V, Vidic J, Manzano M. Development and Evaluation of qPCR Detection Method and Zn-MgO/Alginate Active Packaging for Controlling Listeria monocytogenes Contamination in Cold-Smoked Salmon. Foods. 2020; 9(10):1353. https://doi.org/10.3390/foods9101353
Chicago/Turabian StyleVizzini, Priya, Elena Beltrame, Valentina Zanet, Jasmina Vidic, and Marisa Manzano. 2020. "Development and Evaluation of qPCR Detection Method and Zn-MgO/Alginate Active Packaging for Controlling Listeria monocytogenes Contamination in Cold-Smoked Salmon" Foods 9, no. 10: 1353. https://doi.org/10.3390/foods9101353
APA StyleVizzini, P., Beltrame, E., Zanet, V., Vidic, J., & Manzano, M. (2020). Development and Evaluation of qPCR Detection Method and Zn-MgO/Alginate Active Packaging for Controlling Listeria monocytogenes Contamination in Cold-Smoked Salmon. Foods, 9(10), 1353. https://doi.org/10.3390/foods9101353