Structurally Different Pectic Oligosaccharides Produced from Apple Pomace and Their Biological Activity In Vitro

 ,
,  ,
,
Abstract
1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Mild Acid Hydrolysis Combined with Enzymatic Hydrolysis
2.3. Enzymatic Hydrolysis
2.4. High-Performance Anion-Exchange Chromatography
2.5. Strains of Lactic Acid Bacteria
2.6. Pathogens and Fecal Isolates
2.7. Caco-2 Cell Culture
2.8. Adhesion Assay
2.9. Microscopic Observations
2.10. In Vitro Fermentability of Pectin-Derived Oligosaccharides (POS)
2.11. Short-Chain Fatty Acid (SCFA) and Lactic Acid Analysis
2.12. Statistical Analysis
3. Results
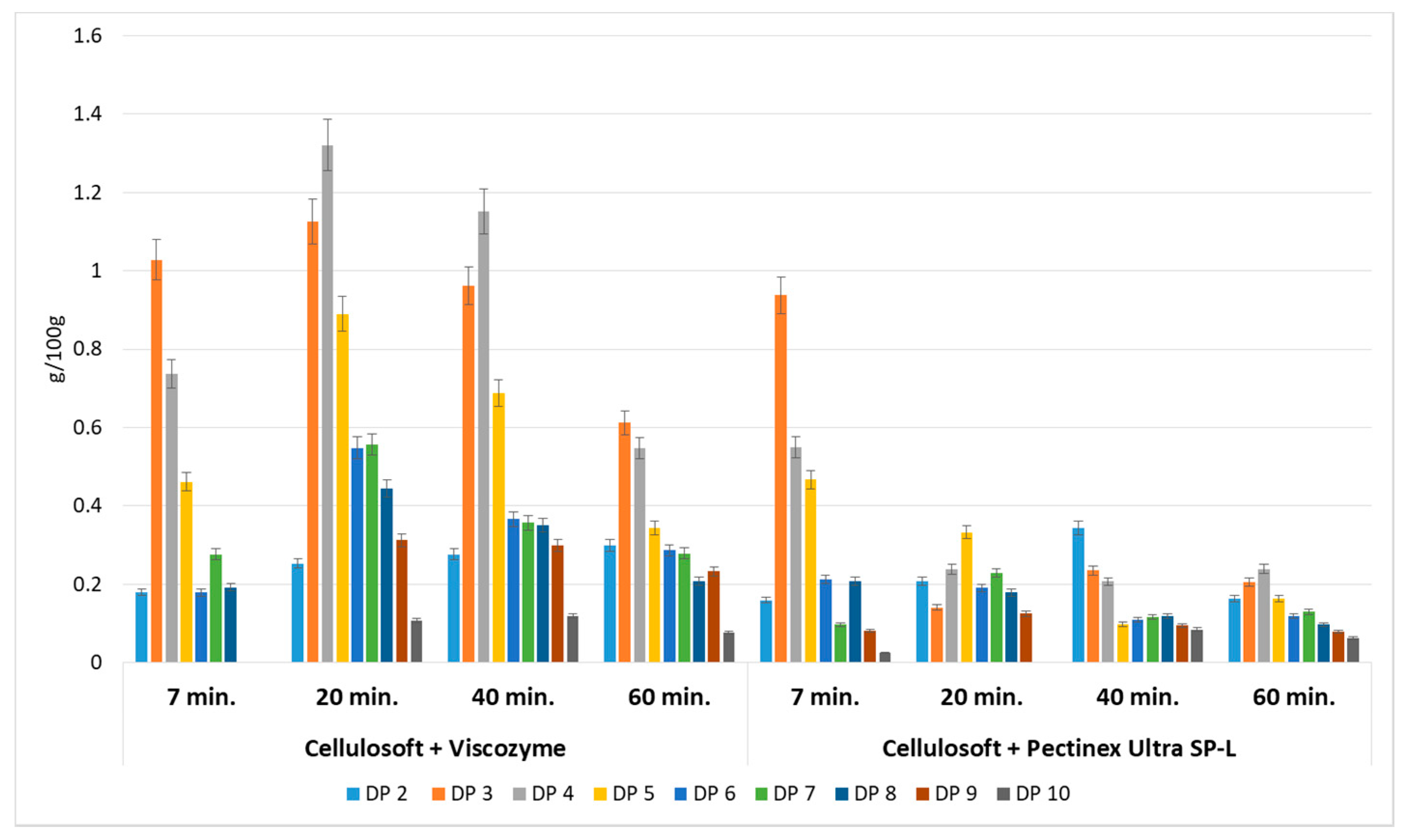
3.1. Enzymatic Hydrolysis of Apple Pomace
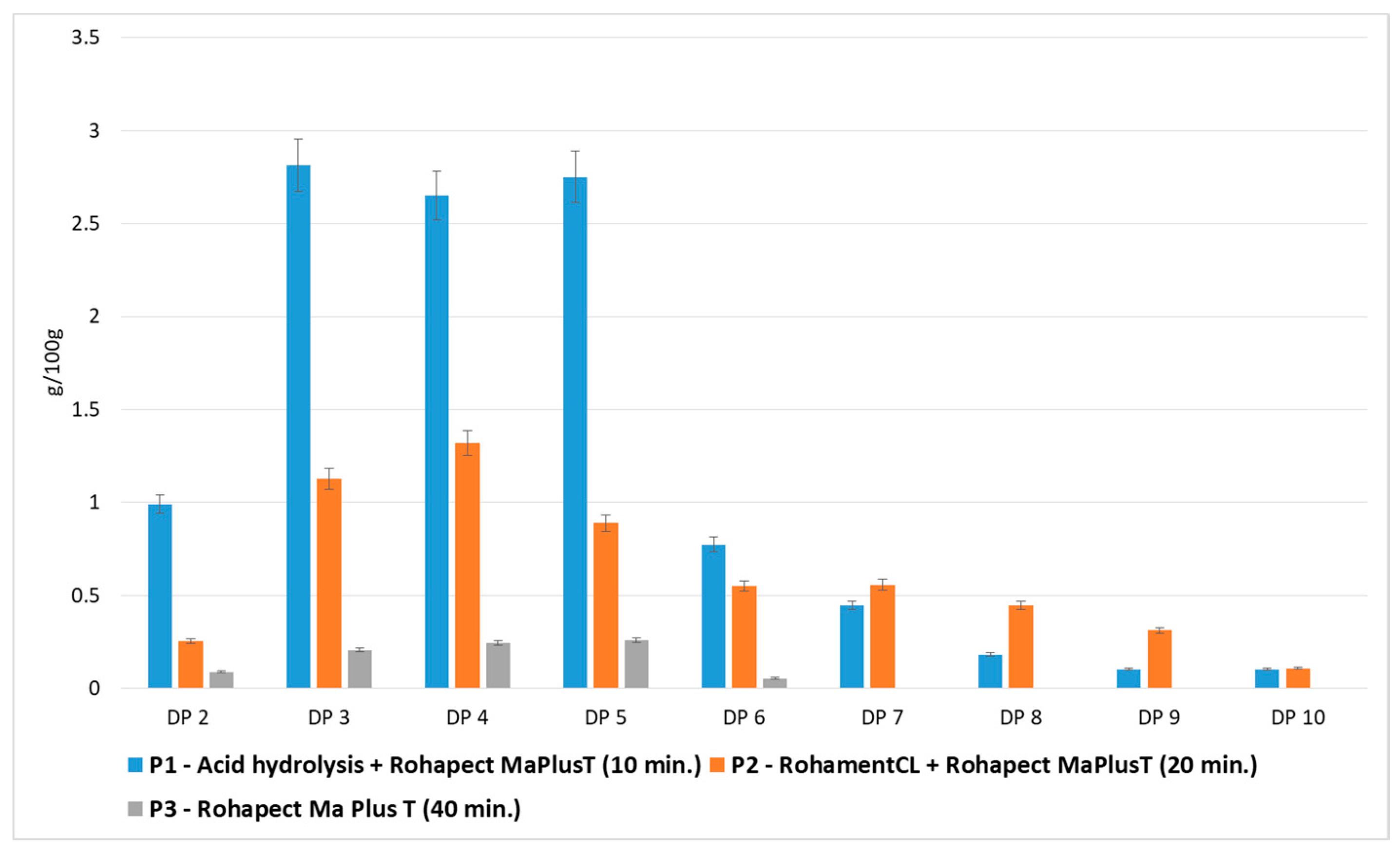
3.2. Mild Acid Hydrolysis Followed by Enzymatic Hydrolysis of Apple Pomace
3.3. Comparison of Applied Hydrolytic Methods
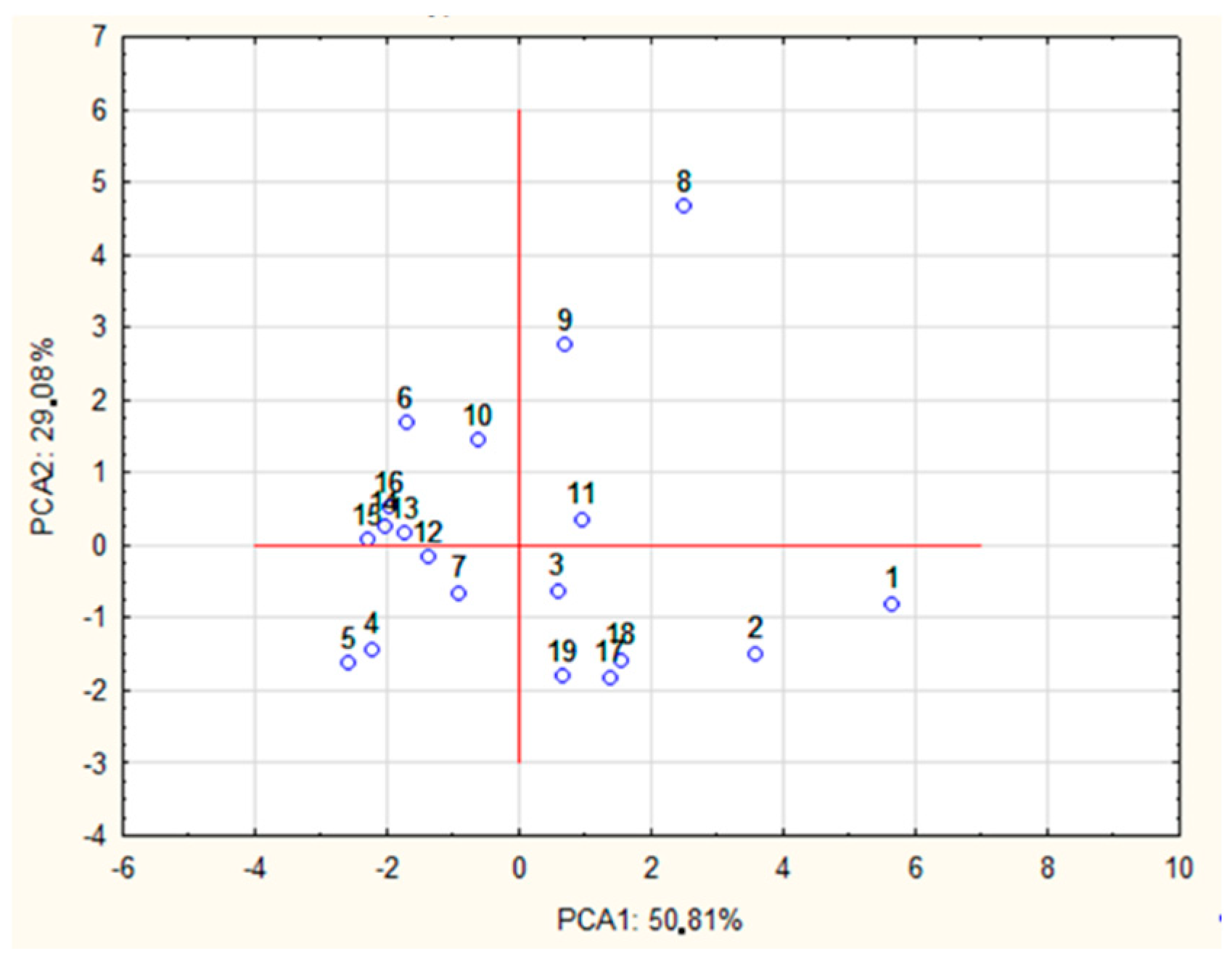
3.4. Relationship Between the Type of POS Obtained and the Hydrolytic Method
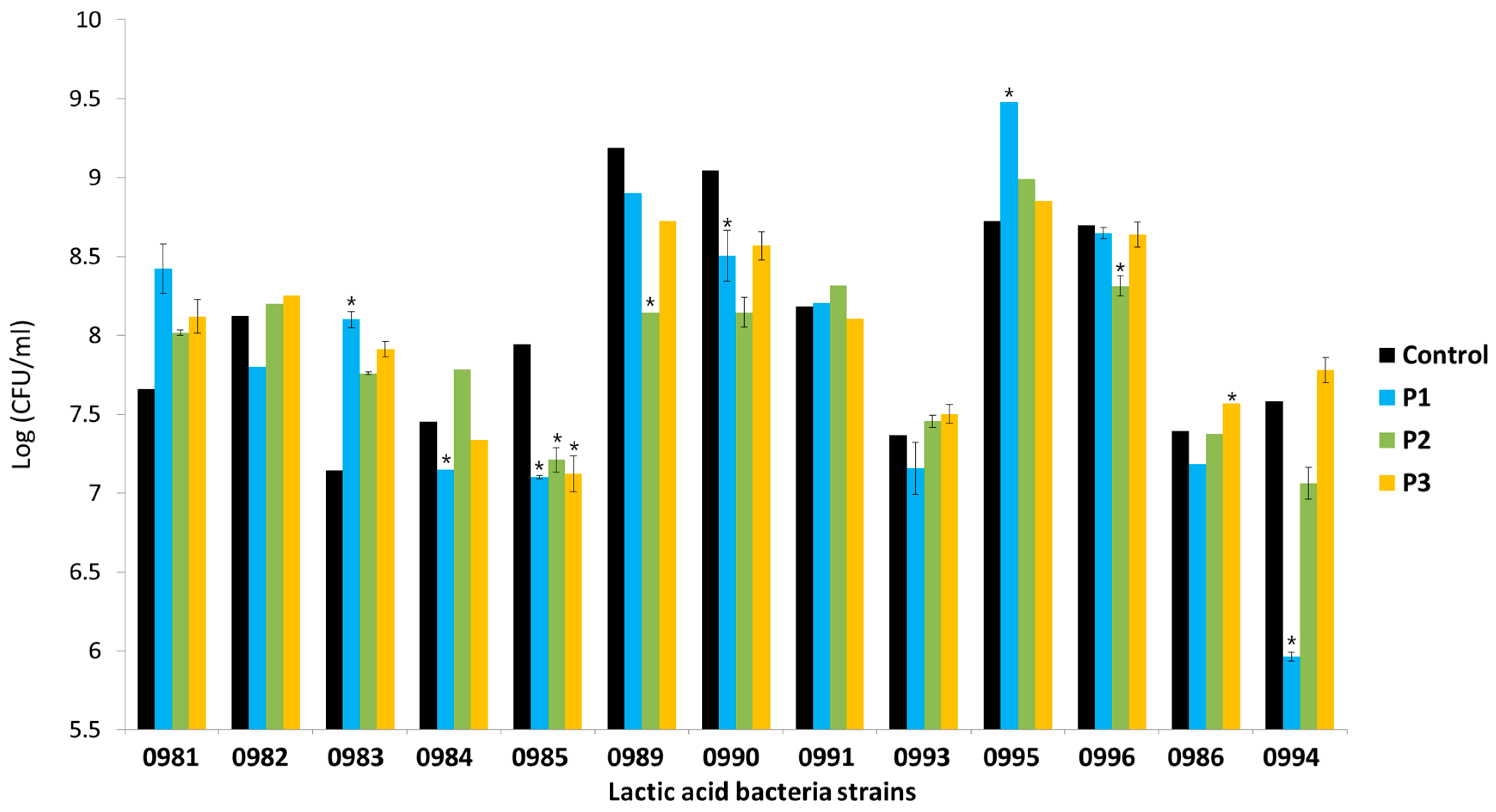
3.5. Adhesion of Lactic Acid Bacteria in the Presence of Apple Pomace Hydrolysates
3.6. Adhesion of Pathogens and Fecal Bacteria in the Presence of Apple Pomace Hydrolysates
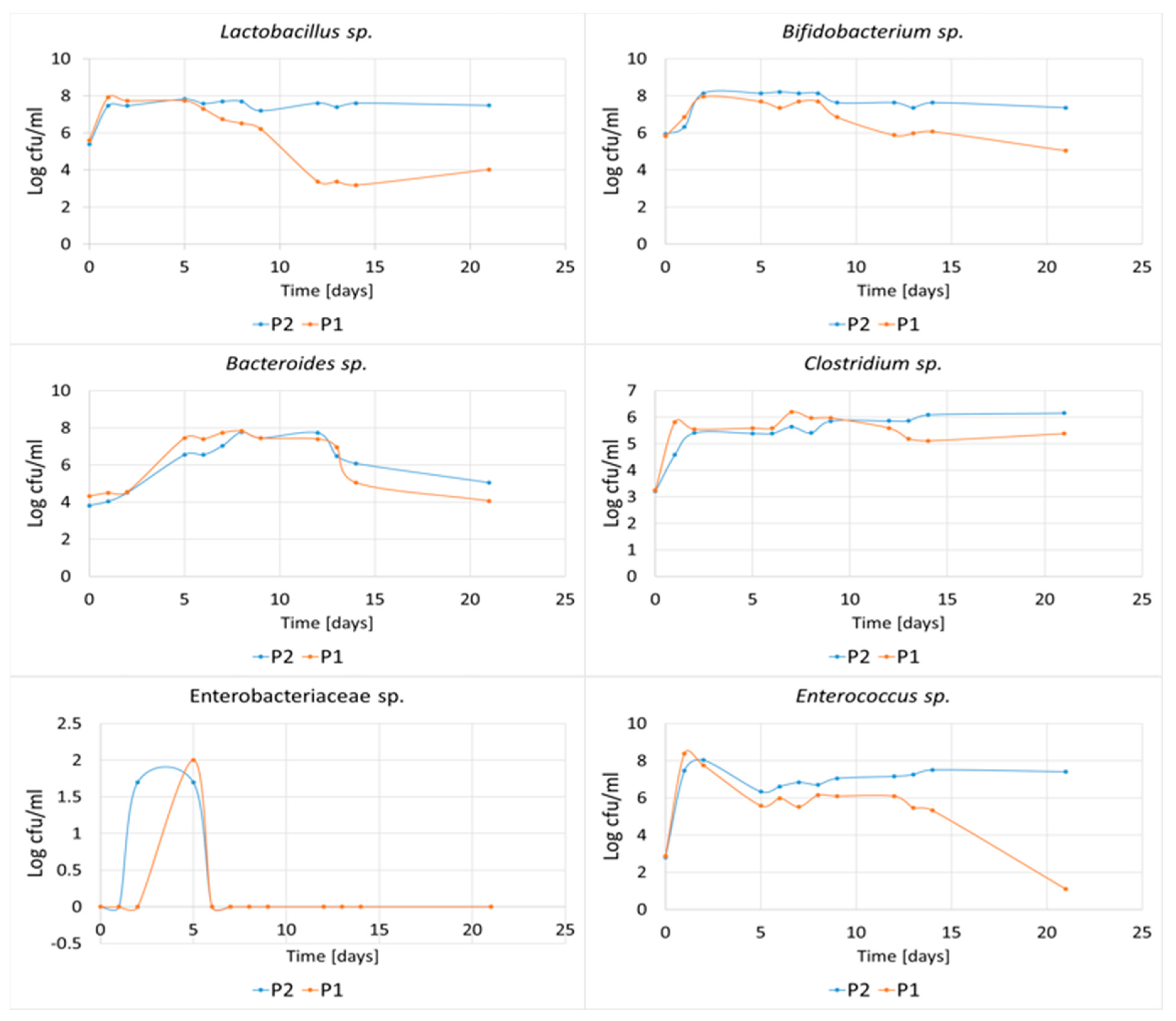
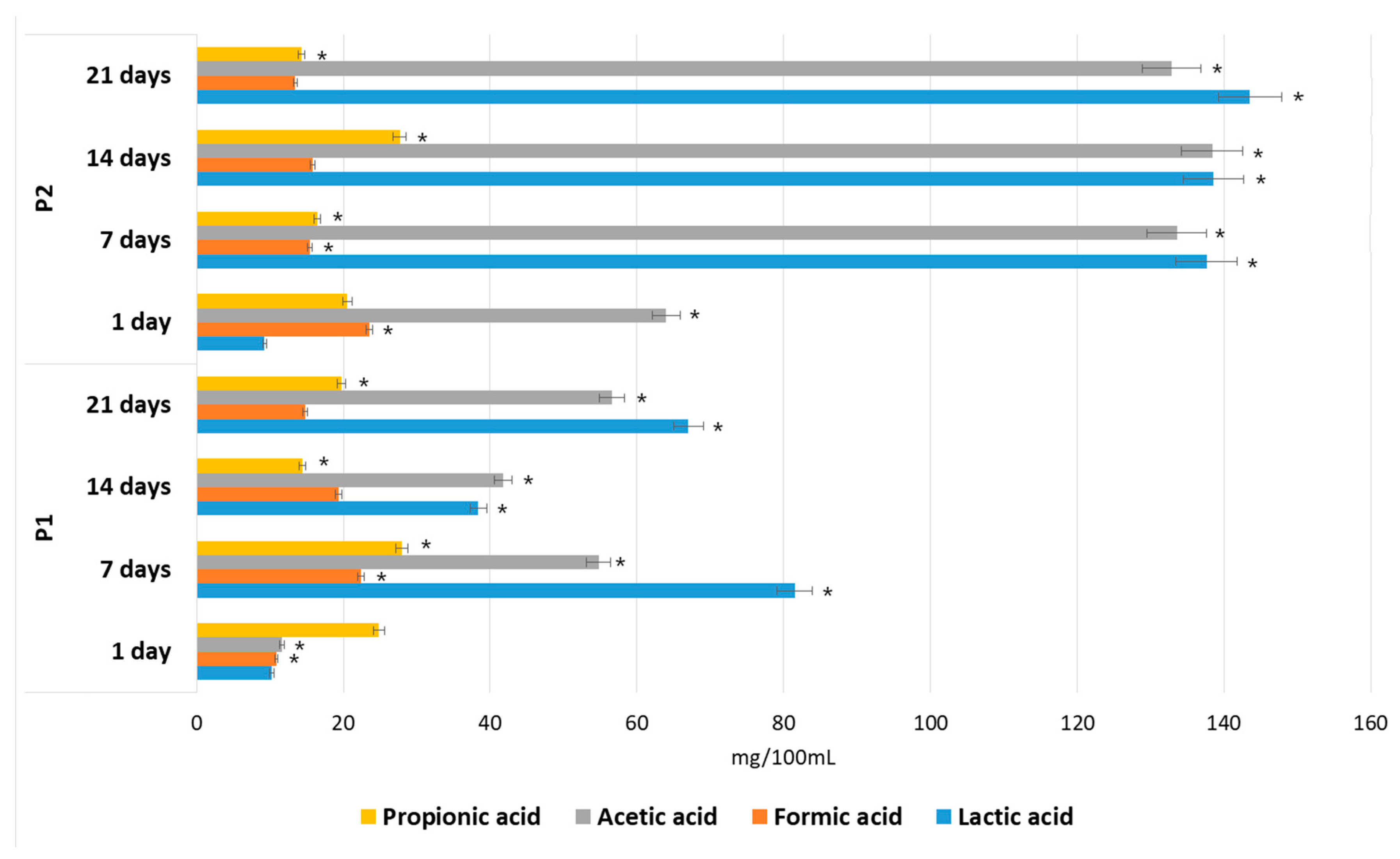
3.7. Effect of POS Composition on the Growth and Metabolism of Microbiota from the Human Gastrointestinal Tract In Vitro
4. Discussion
4.1. The Effect of the Polymerization of POS Contained in the AP Hydrolysate on the Growth and Metabolism of Microbiota from the Human Gastrointestinal Tract
4.2. The Effect of the Polymerization of POS Contained in the AP Hydrolysate on the Adhesion of Lactic Acid Bacteria (LAB) or Pathogens to Human Gut Epithelial Cells
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- FAOSTAT, Value of Agricultural Production. 2017. Available online: http://www.fao.org/faostat/en/#data/QC (accessed on 5 May 2019).
- Sato, M.F.; Vieira, R.G.; Zardo, D.M.; Falcão, L.D.; Nogueira, A.; Wociacki, G. Apple pomace from eleven cultivars: An approach to identify sources of bioactive compounds. Acta Sci. Agron. 2010, 32, 33–34. [Google Scholar]
- Rabetafika, H.N.; Bchir, B.; Blecker, C.; Richel, A. Fractionation of apple by-products as source of new ingredients: Current situation and perspectives. Trends Food Sci. Technol. 2014, 40, 99–114. [Google Scholar] [CrossRef]
- FAO Technical Meeting on Prebiotics. Food Quality and Standards Service (AGNS); FAO Technical Meeting Report; Food and Agriculture Organization of the United Nations (FAO): Rome, Italy, 2007. [Google Scholar]
- Crittenden, R.; Playne, M.J. Prebiotics. In Handbook of Probiotics and Prebiotics; Lee, Y.K., Salminen, S., Eds.; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2009; pp. 535–561. [Google Scholar]
- Ohkuma, K.; Matsuda, I.; Katta, Y.; Hanno, Y. Pyrolysis of starch and its digestibility by enzymes—Characterization of indegestible dextrin. J. Jpn. Soc. Starch Sci. 1999, 37, 107–114. [Google Scholar] [CrossRef]
- Canteri-Schemin, M.H.; Fertonani, H.C.R.; Waszczynskyj, N.; Wosiacki, G. Extraction of pectin from apple pomace. Braz. Arch. Biol. Technol. 2005, 48, 259–266. [Google Scholar] [CrossRef]
- Gómez, B.; Gullón, B.; Yáñez, R.; Schols, H.; Alonso, J.L. Prebiotic potential of pectins and pectic oligosaccharides derived from lemon peel wastes and sugar beet pulp: A comparative evaluation. J. Funct. Foods 2016, 20, 108–121. [Google Scholar] [CrossRef]
- Khodaei, N.; Karboune, S. Optimization of enzymatic production of prebiotic galacto/galacto(arabino)- oligosaccharides and oligomers from potato rhamnogalacturonan I. Carbohydr. Polym. 2018, 181, 1153–1159. [Google Scholar] [CrossRef]
- Hotchkiss, A.T.; Manderson, K.; Tuohy, K.M.; Widmer, W.W.; Nunez, A.; Gibson, G.R.; Rastall, R.A. Bioactive properties of pectic oligosaccharides from sugar beet and Valencia oranges. In Proceedings of the 233rd ACS National Meeting, Chicago, IL, USA, 25–29 March 2007. [Google Scholar]
- Mandalari, G.; Nueno Palop, C.; Tuohy, K.; Gibson, G.R.; Bennett, R.N.; Waldron, K.W. In vitro evaluation of the prebiotic activity of a pectic oligosaccharide-rich extract enzymatically derived from bergamot peel. Appl. Microbiol. Biotechnol. 2007, 73, 1173–1179. [Google Scholar] [CrossRef]
- Gullón, B.; Garrote, G.; Alonso, J.L.; Parajo, J.C. Production of L-lactic acid and oligomeric compounds of apple pomace by simultaneous saccharification and fermentation: A response surface methodology assessment. J. Agric. Food Chem. 2007, 55, 5580–5587. [Google Scholar] [CrossRef]
- Gullón, B.; Yáñez, R.; Alonso, J.L.; Parajó, J.C. l-Lactic acid production from apple pomace by sequential hydrolysis and fermentation. Bioresour. Technol. 2008, 99, 308–319. [Google Scholar] [CrossRef]
- Gullón, B.; Alonso, J.L.; Parajo, J.C. Ion-exchange processing of fermentation media containing lactic acid and oligomeric saccharides. Ind. Eng. Chem. Res. 2010, 49, 3741–3750. [Google Scholar] [CrossRef]
- Gullon, B.; Gullon, P.; Sanz, Y.; Alonso, J.L.; Parajo, J.C. Prebiotic potential of a refined product containing pectic oligosaccharides. LWT-Food Sci. Technol. 2011, 44, 1687–1696. [Google Scholar] [CrossRef]
- Pizarro-Cerdá, J.; Cossart, P. Bacterial Adhesion and Entry into Host Cells. Cell 2006, 124, 715–727. [Google Scholar] [CrossRef] [PubMed]
- Bonnin, E.; Clavurier, K.; Daniel, S.; Kauppinen, S.; Mikkelsen, J.D.M.; Thibault, J.-F. Pectin acetylesterases from Aspergillus are able to deacetylate homogalacturonan as well as rhamnogalacturonan. Carbohydr. Polym. 2008, 74, 411–418. [Google Scholar] [CrossRef]
- Nowak, A.; Motyl, I.; Śliżewska, K.; Libudzisz, Z.; Klewicka, E. Adherence of probiotic bacteria to human colon epithelial cells and inhibitory effect against enteric pathogens. Int. J. Dairy Technol. 2016, 69, 532–539. [Google Scholar]
- Czyżowska, A.; Kucharska, A.Z.; Nowak, A.; Sokół-Łętowska, A.; Motyl, I.; Piórecki, N. Suitability of the probiotic lactic acid bacteria strains as the starter cultures in unripe cornelian cherry (Cornus mas L.) fermentation. J. Food Sci. Technol. 2017, 54, 29–36. [Google Scholar] [CrossRef] [PubMed]
- Mollea, C.; Chiampo, F.; Conti, R. Extraction and characterization of pectins from cocoa husks: A preliminary study. Food Chem. 2008, 107, 1353–1356. [Google Scholar] [CrossRef]
- Yapo, B.M. Pectin quantity, composition and physiochemical behavior as influenced by the purification process. Food Res. Int. 2009, 42, 1197–1202. [Google Scholar] [CrossRef]
- Yapo, B.M.; Robert, C.; Etienne, I.; Wathelet, B.; Paquot, M. Effect of extraction conditions on the yield, purity and surface properties of sugar beet pulp pectin extracts. Food Chem. 2007, 100, 1356–1364. [Google Scholar] [CrossRef]
- Yapo, B.M.; Lerouge, P.; Thibault, J.-F.; Ralet, M.-C. Pectins from citrus peel cell walls contain homogalacturonans homogenous with respect to molar mass, hamnogalacturonan I and rhamnogalacturonan II. Carbohydr. Polym. 2007, 69, 426–435. [Google Scholar] [CrossRef]
- Dahiya, D.K.; Renuka KumarDangi, A.; Shandilya, U.K.; KumarPuniya, A.; Shukla, P. New-Generation Probiotics. In Microbiome and Metabolome in Diagnosis, Therapy, and Other Strategic Applications; Academic Press: New York, NY, USA, 2019; pp. 417–424. [Google Scholar]
- Topping, D.L.; Clifton, P.M. Short chain fatty acids and human colonic function: Roles of resistant starch and non starch polysaccharides. Physiol. Rev. 2001, 81, 1031–1064. [Google Scholar] [CrossRef]
- Muegge, B.D. Diet drives convergence in gut microbiome functions across mammalian phylogeny and within humans. Science 2011, 332, 970–974. [Google Scholar] [CrossRef] [PubMed]
- Duncan, S.H.; Belenguer, A.; Holtrop, G.; Johnstone, A.M.; Flint, H.J.; Lobley, G.E. Reduced dietary intake of carbohydrates by obese subjects results in decreased concentrations of butyrate and butyrate-producing bacteria in feces. Appl. Environ. Microbiol. 2007, 73, 1073–1078. [Google Scholar] [CrossRef] [PubMed]
- Turnbaugh, P.J.; Ridaura, V.K.; Faith, J.J.; Rey, F.E.; Knight, R.; Gordon, J.I. The effect of diet on the human gut microbiome: A metagenomic analysis in humanized gnotobiotic mice. Sci. Transl. Med. 2009, 1, 6–14. [Google Scholar] [CrossRef] [PubMed]
- De Filippo, C.; Cavalieri, D.; Di Paola, M.; Ramazzotti, M.; Poullet, J.B.; Massart, S.; Collini, S.; Pieraccini, G.; Lionetti, P. Impact of diet in shaping gut microbiota revealed by a comparative study in children from Europe and rural Africa. Proc. Natl. Acad. Sci. USA 2010, 107, 14691–14696. [Google Scholar] [CrossRef] [PubMed]
- Macfarlane, G.T.; Macfarlane, S. Fermentation in the human large intestine: Its physiologic consequences and the potential contribution of prebiotics. J. Clin. Gastroenterol. 2011, 45, 120–127. [Google Scholar] [CrossRef] [PubMed]
- Brownawell, A.M.; Caers, W.; Gibson, G.R.; Kendall, C.W.; Lewis, K.D.; Ringel, Y.; Slavin, J.L. Prebiotics and the health benefits of fiber: Current regulatory status, future research, and goals. J. Nutr. 2012, 142, 962–974. [Google Scholar] [CrossRef] [PubMed]
- Meyer, D. Health benefits of prebiotic fibers. Adv. Food Nutr. Res. 2015, 74, 47–91. [Google Scholar] [PubMed]
- Tan, J.; McKenzie, C.; Potamitis, M.; Thorburn, A.N.; Mackay, C.R.; Macia, L. The role of short-chain fatty acids in health and disease. Adv. Immunol. 2014, 121, 91–119. [Google Scholar]
- Wilson, B.; Whelan, K. Prebiotic inulin-type fructans and galacto-oligosaccharides: Definition, specificity, function, and application in gastrointestinal disorders. J. Gastroenterol. Hepatol. 2017, 32, 64–68. [Google Scholar] [CrossRef]
- Moon, J.S.; Shin, S.Y.; Choi, H.S.; Joo, W.; Cho, S.K.; Li, L.; Kanga, J.H.; Kim, T.J.; Han, N.S. In vitro digestion and fermentation properties of linear sugar-beet arabinan and its oligosaccharides. Carbohydr. Polym. 2015, 131, 50–56. [Google Scholar] [CrossRef]
- Gibson, G.R.; Roberfroid, M.B. Dietary modulation of the colonic microbiota: Introducing the concept of prebiotics. J. Nutr. 1995, 125, 1401–1412. [Google Scholar] [CrossRef]
- Kadlec, R.; Jakubec, M. The effect of prebiotics on adherence of probiotics. J. Dairy Sci. 2014, 97, 1983–1990. [Google Scholar] [CrossRef]
- Laparra, J.M.; Sanz, Y. Comparison of in vitro models to study bacterial adhesion to the intestinal epithelium. Lett. Appl. Microbiol. 2009, 49, 695–701. [Google Scholar] [CrossRef]
- Tallon, R.; Arias, S.; Bressollier, P.; Urdaci, M.C. Strain- and matrix-dependent adhesion of Lactobacillus plantarum is mediated by proteinaceous bacterial compounds. J. Appl. Microbiol. 2007, 102, 442–451. [Google Scholar] [CrossRef]
- Quigley, E.M.M. Prebiotics and probiotics; modifying and mining the microbiota. Pharmacol. Res. 2010, 61, 213–218. [Google Scholar] [CrossRef]
- Koh, J.H.; Choi, S.H.; Park, S.W.; Choi, N.J.; Kim, Y.; Kim, S.H. Synbiotic impact of tagatose on viability of Lactobacillus rhamnosus strain GG mediated by the phosphotransferase system (PTS). Food Microbiol. 2013, 36, 7–13. [Google Scholar] [CrossRef]
- Kavanaugh, D.W.; O’Callaghan, J.; Butto, L.F.; Slattery, H.; Lane, J.; Clyne, M.; Kane, M.; Joshi, L.; Hickey, R.M. Exposure of Bifidobacterium longum subsp. infantis to Milk Oligosaccharides Increases Adhesion to Epithelial Cells and Induces a Substantial Transcriptional Response. PLoS ONE 2013, 8, 1–14. [Google Scholar] [CrossRef]
- Krausova, G.; Hyrsolva, I.; Jakubec, M.; Hynstova, I. In Vitro Evaluation of Prebiotics on Adherence of Lactobacilli. J. Microb. Biochem. Technol. 2016, 8, 6–8. [Google Scholar]
- Russo, P.; López, P.; Capozzi, V.; Palencia, P.F.; Dueñas, M.T.; Spano, G.; Fiocco, D. Beta-Glucans Improve Growth, Viability and Colonization of Probiotic Microorganisms. Int. J. Mol. Sci. 2012, 13, 6026–6039. [Google Scholar] [CrossRef]
- Tamminen, M.; Salminen, S.; Ouwehand, A.C. Fermentation of carrot juice by probiotics: Viability and preservation of adhesion. Int. J. Biotechnol. Wellness Ind. 2013, 2, 10–15. [Google Scholar] [CrossRef]
- Ouwehand, A.C.; Tuomola, E.M.; Tölkkö, S.; Salminen, S. Assessment of adhesion properties of novel probiotic strains to human intestinal mucus. Int. J. Food Microbiol. 2001, 64, 119–126. [Google Scholar] [CrossRef]
- Al-Ghazzewi, F.H.; Tester, R.F. Inhibition of the adhesion of Escherichia coli to human epithelial cells by carbohydrates. Bioact. Carbohydr. Diet. Fibre 2014, 4, 1–5. [Google Scholar] [CrossRef]
- Antão, E.-M.; Wieler, L.H.; Ewers, C. Adhesive threads of extra intestinal pathogenic Escherichia coli. Gut Pathog. 2009, 1, 22. [Google Scholar] [CrossRef]
- Shoaf-Sweeney, K.D.; Hutkins, R.W. Adherence, anti-adherence, and oligosaccharides preventing pathogens from sticking to the host. Adv. Food Nutr. Res. 2009, 55, 101–161. [Google Scholar]
- Ebersbach, T.; Andersen, J.B.; Bergström, A.; Hutkins, R.W.; Licht, T.R. Xylo-oligosaccharides inhibit pathogen adhesion to enterocytes in vitro. Res. Microbiol. 2012, 163, 22–27. [Google Scholar] [CrossRef]
- Gupta, K.; Chou, M.Y.; Howell, A.; Wobbe, C.; Grady, R.; Stapleton, A.E. Cranberry products inhibit adherence of p-fimbriated Escherichia coli to primary cultured bladder and vaginal epithelial cells. J. Urol. 2007, 177, 2357–2360. [Google Scholar] [CrossRef]
- Hickey, R.M. The role of oligosaccharides from human milk and other sources in prevention of pathogen adhesion. Int. Dairy J. 2012, 22, 141–146. [Google Scholar] [CrossRef]
- Parkar, S.G.; Redgate, E.L.; Wibisono, R.; Luo, X.; Koh, E.T.H.; Schröder, R. Gut health benefits of kiwifruit pectins: Comparison with commercial functional polysaccharides. J. Funct. Foods 2010, 2, 210–218. [Google Scholar] [CrossRef]
- Rhoades, J.; Manderson, K.; Wells, A.; Hotchkiss, A.T.; Gibson, G.R.; Formentin, K.; Beer, M.; Rastall, R.A. Oligosaccharide-Mediated Inhibition of the Adhesion of Pathogenic Escherichia coli Strains to Human Gut Epithelial Cells in Vitro. J. Food Prot. 2008, 71, 2272–2277. [Google Scholar] [CrossRef]
- Buddington, K.; Donahoo, J.; Buddington, R.K. Dietary oligofructose and inulin protect mice from enteric and systemic pathogens and tumor inducers. J. Nutr. 2002, 132, 472–477. [Google Scholar] [CrossRef]
- Altamimi, M.; Abdelhay, O.; Rastall, R.A. Effect of oligosaccharides on the adhesion of gut bacteria to human HT-29 cells. Anaerobe 2016, 39, 136–142. [Google Scholar] [CrossRef]
- Petersen, A.; Bergström, A.; Andersen, J.B.; Hansen, M.; Lahtinen, S.J.; Wilcks, A.; Licht, T.R. Analysis of the intestinal microbiota of oligosaccharides fed mice exhibiting reduced resistance to Salmonella infection. Benef. Microbes 2010, 1, 271–281. [Google Scholar] [CrossRef]
- Petersen, A.; Heegaard, P.M.H.; Pedersen, A.L.; Andersen, J.B.; Sorensen, R.B.; Frokiar, H.; Lahtinen, S.J.; Ouwehand, A.C.; Poulsen, M.; Licht, T.R.; et al. Some putative prebiotics increase the severity of Salmonella enterica serovar Typhimurium infection in mice. BMC Microbiol. 2009, 9, 245–255. [Google Scholar] [CrossRef]













{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Microorganism | Cultivation Media | Cultivation Conditions |
|---|---|---|
| Lactobacillus | Rogosa Agar (MERCK) | 37 °C, 6 days, anaerobic |
| Bifidobacterium | Bifidobacterium medium | 37 °C, 6 days, anaerobic |
| Clostridium | Tryptose Sulfite Cycloserine Agar (MERCK) | 37 °C, 6 days, anaerobic |
| Enterococcus | Bile Aesculin Agar (MERCK) | 37 °C, 24 h, aerobic |
| Bacteroides | Agar SCHAEDLERA-KV (MERCK) | 37 °C, 6 days, anaerobic |
| Enterobacteriaceae | Mac Conkey Agar (MERCK) | 37 °C, 24 h, aerobic |
| PCA1 | PCA2 | |
|---|---|---|
| DP 2 | 0.918 | −0.189 |
| DP 3 | 0.934 | 0.031 |
| DP 4 | 0.950 | −0.007 |
| DP 5 | 0.967 | −0.035 |
| DP 6 | 0.946 | 0.246 |
| DP 7 | 0.678 | 0.593 |
| DP 8 | 0.109 | 0.886 |
| DP 9 | −0.109 | 0.955 |
| DP 10 | −0.025 | 0.800 |
| Strain | Stimulation (+)/Inhibition Rate (−) (%) | ||
|---|---|---|---|
| P1 | P2 | P3 | |
| Lb. plantarum 0981 | +10.0 | +4.7 | +6.0 |
| Lb. plantarum 0982 | −4.0 | +0.9 | +1.5 |
| Lb. brevis 0983 | +13.4 | +8.6 | +10.7 |
| Lb. brevis 0984 | −4.1 | +4.5 | −1.5 |
| Lb. paracasei 0985 | −10.6 | −9.2 | −10.4 |
| Lb. plantarum 0989 | −3.1 | −11.3 | −5.0 |
| Lb. plantarum 0990 | −6.0 | −9.9 | −5.3 |
| Lb. plantarum 0991 | +0.3 | +1.7 | −0.9 |
| Lb. paracasei 0993 | −2.9 | +1.2 | +1.9 |
| Lb. plantarum 0995 | +8.7 | +3.1 | +1.5 |
| Lb. plantarum 0996 | −0.6 | −4.4 | −1.8 |
| Leu. mesenteroides 0986 | −2.9 | −0.2 | +2.4 |
| Leu. mesenteroides 0994 | −21.3 | −6.8 | +2.6 |
| Strain | Inhibition (−)/Stimulation (+) Rate (%) | ||
|---|---|---|---|
| P1 | P2 | P3 | |
| Escherichia coli ATCC 10536 | −13.9 | −12.9 | −9.3 |
| Salmonella Typhimurium ATCC 14028 | −12.0 | −13.6 | −14.2 |
| Listeria monocytogenes ATCC 19115 | −27.9 | −27.9 | −27.9 |
| Aerobic fecal bacteria | −9.4 | −7.7 | +2.9 |
| Anaerobic fecal bacteria | −2.9 | −2.1 | −3.1 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wilkowska, A.; Nowak, A.; Antczak-Chrobot, A.; Motyl, I.; Czyżowska, A.; Paliwoda, A. Structurally Different Pectic Oligosaccharides Produced from Apple Pomace and Their Biological Activity In Vitro. Foods 2019, 8, 365. https://doi.org/10.3390/foods8090365
Wilkowska A, Nowak A, Antczak-Chrobot A, Motyl I, Czyżowska A, Paliwoda A. Structurally Different Pectic Oligosaccharides Produced from Apple Pomace and Their Biological Activity In Vitro. Foods. 2019; 8(9):365. https://doi.org/10.3390/foods8090365
Chicago/Turabian StyleWilkowska, Agnieszka, Adriana Nowak, Aneta Antczak-Chrobot, Ilona Motyl, Agata Czyżowska, and Anna Paliwoda. 2019. "Structurally Different Pectic Oligosaccharides Produced from Apple Pomace and Their Biological Activity In Vitro" Foods 8, no. 9: 365. https://doi.org/10.3390/foods8090365
APA StyleWilkowska, A., Nowak, A., Antczak-Chrobot, A., Motyl, I., Czyżowska, A., & Paliwoda, A. (2019). Structurally Different Pectic Oligosaccharides Produced from Apple Pomace and Their Biological Activity In Vitro. Foods, 8(9), 365. https://doi.org/10.3390/foods8090365