Technofunctional and Sensory Properties of Fermented Lupin Protein Isolates



Abstract
1. Introduction
2. Materials and Methods
2.1. Lupin Seeds
2.2. Preparation of Lupin Protein Isolate
2.3. Microbiological Compositions
2.3.1. Nutrient Media
2.3.2. Bacteria Strains and Culture Conditions
2.3.3. Determining Growth Conditions of Microorganisms
2.3.4. Determination of pH and Viable Cell Counts
2.4. Fermentation of Lupin Protein Isolates
2.5. Analysis of d-Glucose
2.6. Chemical Composition
2.7. Sensory Analysis
2.7.1. Panelists
2.7.2. Sensory Evaluation
2.8. Techno-Functional Properties
2.8.1. Protein Solubility
2.8.2. Foam Properties
2.8.3. Emulsifying Capacity
2.9. Analysis of the Molecular Weight Profiles of LPI and Fermented LPI
2.10. Statistical Analysis
3. Results and Discussion
3.1. Chemical Composition
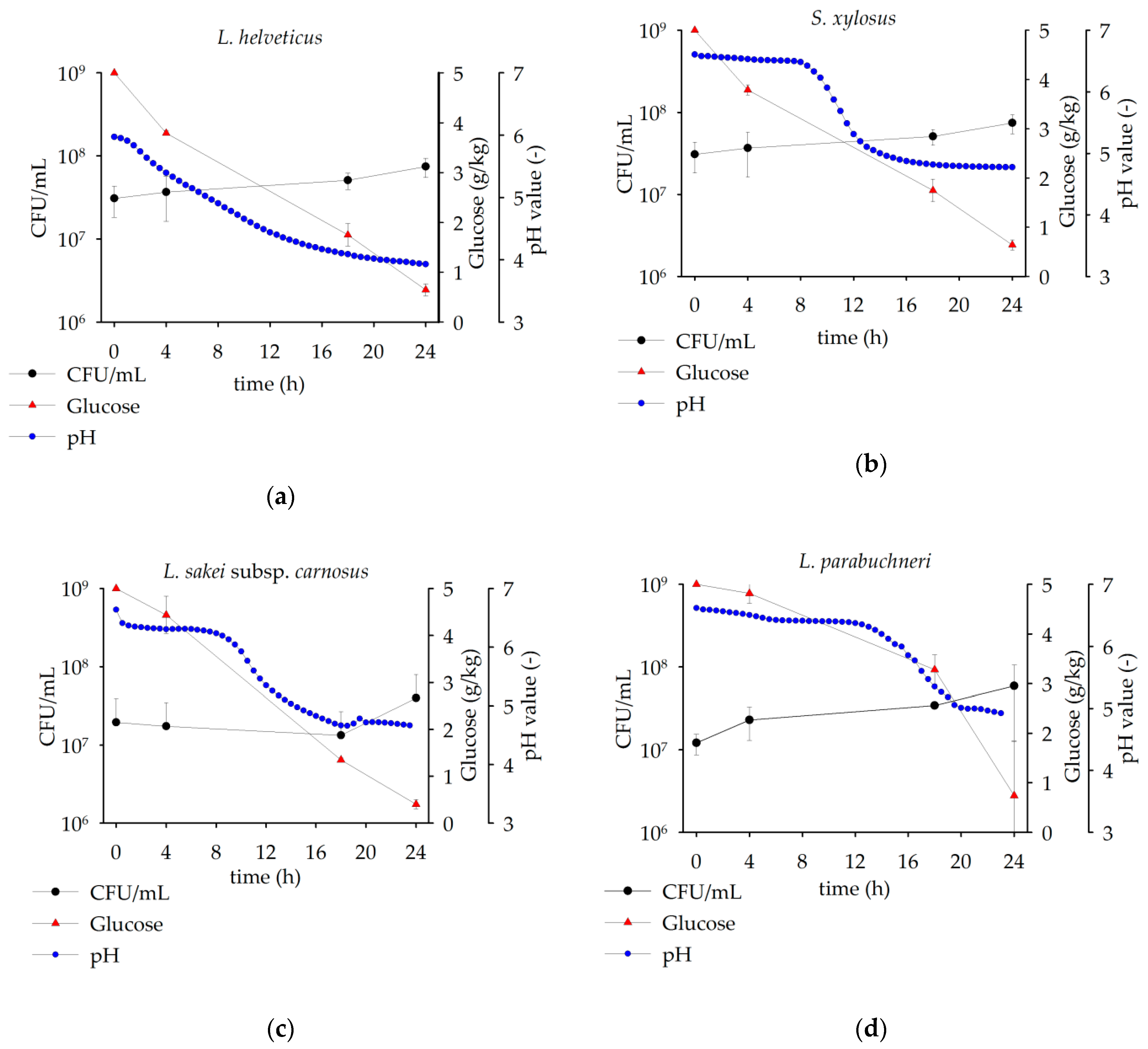
3.2. Comparison of Microbial Growth on Lupin Protein Isolate Solutions
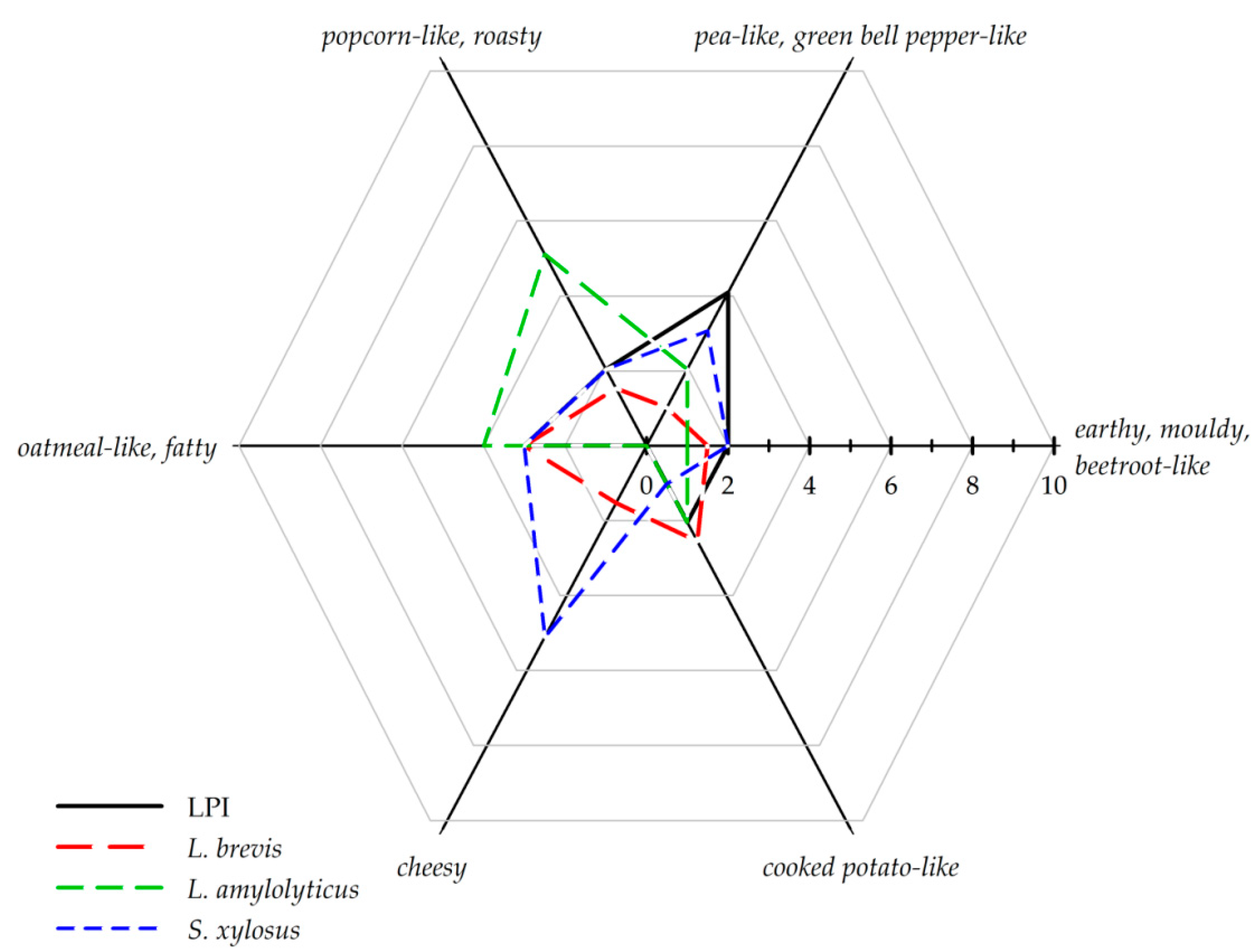
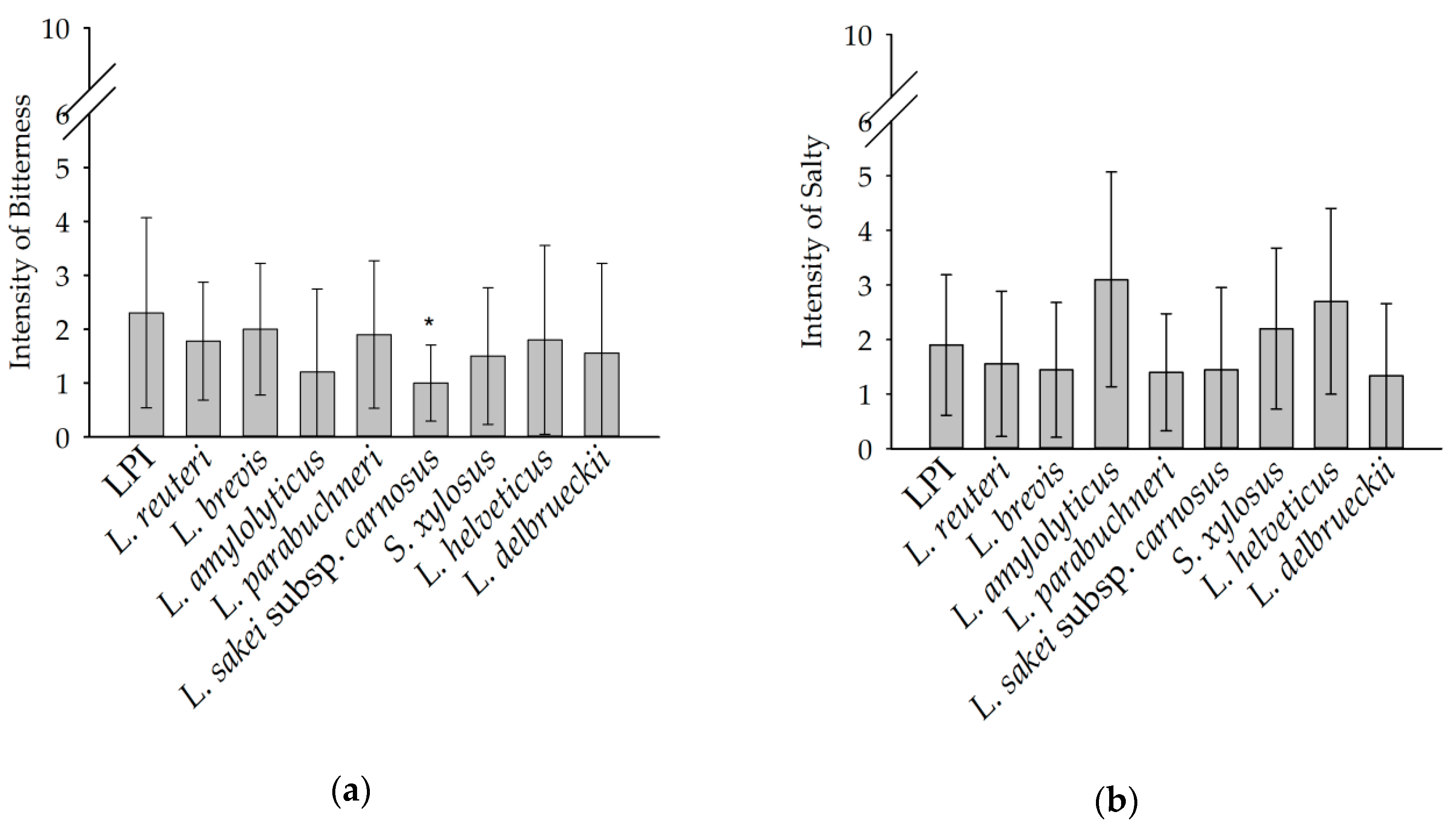
3.3. Sensory Anaylsis
3.4. Techno-Functional Properties
3.4.1. Protein Solubility
3.4.2. Foam Properties
3.4.3. Emulsifying Capacity
3.5. Analysis of the SDS-PAGE Profiles of Unfermented and Fermented (24 h) LPI
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Eshel, G.; Shepon, A.; Makov, T.; Milo, R. Land, irrigation water, greenhouse gas, and reactive nitrogen burdens of meat, eggs, and dairy production in the United States. Proc. Natl. Acad. Sci. USA 2014, 111, 11996–12001. [Google Scholar] [CrossRef] [PubMed]
- Westhoek, H.; Lesschen, J.P.; Rood, T.; Wagner, S.; De Marco, A.; Murphy-Bokern, D.; Leip, A.; van Grinsven, H.; Sutton, M.A.; Oenema, O. Food choices, health and environment: Effects of cutting Europe’s meat and dairy intake. Glob. Environ. Chang. 2014, 26, 196–205. [Google Scholar] [CrossRef]
- Arnoldi, A.; Boschin, G.; Zanoni, C.; Lammi, C. The health benefits of sweet lupin seed flours and isolated proteins. J. Funct. Foods 2015, 18, 550–563. [Google Scholar] [CrossRef]
- Bader, S.; Bez, J.; Eisner, P. Can protein functionalities be enhanced by high-pressure homogenization?—A study on functional properties of lupin proteins. Procedia Food Sci. 2011, 1, 1359–1366. [Google Scholar] [CrossRef]
- Schlegel, K.; Sontheimer, K.; Hickisch, A.; Wani, A.A.; Eisner, P.; Schweiggert-Weisz, U. Enzymatic hydrolysis of lupin protein isolates—Changes in the molecular weight distribution, technofunctional characteristics and sensory attributes. Food Sci. Nutr. 2019, 7, 2747–2759. [Google Scholar] [CrossRef] [PubMed]
- Schindler, S.; Zelena, K.; Krings, U.; Bez, J.; Eisner, P.; Berger, R.G. Improvement of the Aroma of Pea (Pisum sativum) Protein Extracts by Lactic Acid Fermentation. Food Biotechnol. 2012, 26, 58–74. [Google Scholar] [CrossRef]
- Schindler, S.; Wittig, M.; Zelena, K.; Krings, U.; Bez, J.; Eisner, P.; Berger, R.G. Lactic fermentation to improve the aroma of protein extracts of sweet lupin (Lupinus angustifolius). Food Chem. 2011, 128, 330–337. [Google Scholar] [CrossRef]
- Blagden, T.D.; Gilliland, S.E. Reduction of Levels of Volatile Components Associated with the “Beany” Flavor in Soymilk by Lactobacilli and Streptococci. J. Food Sci. 2005, 70, M186–M189. [Google Scholar] [CrossRef]
- Lampart-Szczapa, E.; Konieczny, P.; Nogala-Kałucka, M.; Walczak, S.; Kossowska, I.; Malinowska, M. Some functional properties of lupin proteins modified by lactic fermentation and extrusion. Food Chem. 2006, 96, 290–296. [Google Scholar] [CrossRef]
- Meinlschmidt, P.; Ueberham, E.; Lehmann, J.; Schweiggert-Weisz, U.; Eisner, P. Immunoreactivity, sensory and physicochemical properties of fermented soy protein isolate. Food Chem. 2016, 205, 229–238. [Google Scholar] [CrossRef]
- Song, Y.-S.; Frías, J.; Martinez-Villaluenga, C.; Vidal-Valdeverde, C.; de Mejia, E.G. Immunoreactivity reduction of soybean meal by fermentation, effect on amino acid composition and antigenicity of commercial soy products. Food Chem. 2008, 108, 571–581. [Google Scholar] [CrossRef] [PubMed]
- Song, Y.S.; Pérez, V.G.; Pettigrew, J.E.; Martinez-Villaluenga, C.; de Mejia, E.G. Fermentation of soybean meal and its inclusion in diets for newly weaned pigs reduced diarrhea and measures of immunoreactivity in the plasma. Anim. Feed Sci. Technol. 2010, 159, 41–49. [Google Scholar] [CrossRef]
- Frias, J.; Song, Y.S.; Martínez-Villaluenga, C.; De Mejia, E.G.; Vidal-Valverde, C. Immunoreactivity and Amino Acid Content of Fermented Soybean Products. J. Agric. Food Chem. 2008, 56, 99–105. [Google Scholar] [CrossRef] [PubMed]
- Kobayashi, M. Immunological functions of soy sauce: Hypoallergenicity and antiallergic activity of soy sauce. J. Biosci. Bioeng. 2005, 100, 144–151. [Google Scholar] [CrossRef] [PubMed]
- Mosse, J.; Huet, J.-C.; Baudet, J. Relationships between nitrogen, amino acids and storage proteins in Lupinus albus seeds. Phytochemistry 1987, 26, 2453–2458. [Google Scholar] [CrossRef]
- Morr, C.V.; German, B.; Kinsella, J.E.; Regenstein, J.M.; Vanburen, J.P.; Kilara, A.; Lewis, B.A.; Mangino, M.E. A Collaborative Study to Develop a Standardized Food Protein Solubility Procedure. J. Food Sci. 1985, 50, 1715–1718. [Google Scholar] [CrossRef]
- Lowry, O.H.; Rosebrough, N.J.; Farr, A.L.; Randall, R.J. Protein measurement with the Folin phenol reagent. J. Biol. Chem. 1951, 193, 265–275. [Google Scholar]
- Phillips, L.G.; Haque, Z.; Kinsella, J.E. A Method for the Measurement of Foam Formation and Stability. J. Food Sci. 1987, 52, 1074–1077. [Google Scholar] [CrossRef]
- Wang, C.Y.; Johnson, L.A. Functional properties of hydrothermally cooked soy protein products. J. Am. Oil Chem. Soc. 2001, 78, 189–195. [Google Scholar] [CrossRef]
- Laemmli, U.K. Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature 1970, 227, 680–685. [Google Scholar] [CrossRef]
- Fritsch, C.; Vogel, R.F.; Toelstede, S. Fermentation performance of lactic acid bacteria in different lupin substrates—Influence and degradation ability of antinutritives and secondary plant metabolites. J. Appl. Microbiol. 2015, 119, 1075–1088. [Google Scholar] [CrossRef] [PubMed]
- Lqari, H.; Pedroche, J.; Girón-Calle, J.; Vioque, J.; Millán, F. Production of Lupinus angustifolius protein hydrolysates with improved functional properties. Grasas Aceites 2005, 56, 135–140. [Google Scholar] [CrossRef]
- Piornos, J.A.; Burgos-Díaz, C.; Ogura, T.; Morales, E.; Rubilar, M.; Maureira-Butler, I.; Salvo-Garrido, H. Functional and physicochemical properties of a protein isolate from AluProt-CGNA: A novel protein-rich lupin variety (Lupinus luteus). Food Res. Int. 2015, 76, 719–724. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez-Ambriz, S.L.; Martínez-Ayala, A.L.; Millán, F.; Dávila-Ortíz, G. Composition and Functional Properties of Lupinus campestris Protein Isolates. Plant Foods Hum. Nutr. 2005, 60, 99–107. [Google Scholar] [CrossRef]
- Weng, T.M.; Chen, M.T. Changes of Protein in Natto (a fermented soybean food) Affected by Fermenting Time. Food Sci. Technol. Res. 2010, 16, 537–542. [Google Scholar] [CrossRef]
- Murekatete, N.; Hua, Y.; Kong, X.; Zhang, C. Effects of Fermentation on Nutritional and Functional Properties of Soybean, Maize, and Germinated Sorghum Composite Flour. Int. J. Food Eng. 2012, 8, 1–15. [Google Scholar] [CrossRef]
- Klupsaite, D.; Juodeikiene, G.; Zadeike, D.; Bartkiene, E.; Maknickiene, Z.; Liutkute, G. The influence of lactic acid fermentation on functional properties of narrow-leaved lupine protein as functional additive for higher value wheat bread. LWT-Food Sci. Technol. 2017, 75, 180–186. [Google Scholar] [CrossRef]
- El-Adawy, T.A.; Rahma, E.H.; El-Bedawey, A.A.; Gafar, A.F. Nutritional potential and functional properties of sweet and bitter lupin seed protein isolates. Food Chem. 2001, 74, 455–462. [Google Scholar] [CrossRef]
- Qi, M.; Hettiarachchy, N.S.; Kalapathy, U. Solubility and Emulsifying Properties of Soy Protein Isolates Modified by Pancreatin. J. Food Sci. 1997, 62, 1110–1115. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Bacteria Strain | |
|---|---|
| Lactobacillus reuteri | DSM 20016 |
| Lactobacillus brevis | TMW 1.1326 |
| Lactobacillus amylolyticus | TL 5 |
| Lactobacillus parabuchneri | DSM 5987 |
| Lactobacillus sakei subsp. carnosus | DSM 15831 |
| Staphylococcus xylosus | DSM 20266 |
| Lactobacillus helveticus | DSM 20075 |
| Lactobacillus delbrueckii | DSM 20081 |
| Dry Matter | Protein Content | |
|---|---|---|
| LPI (unfermented) | 95.4 ± 0.0% | 89.6 ± 0.0% |
| L. reuteri | 94.6 ± 0.0% * | 82.4 ± 1.3% |
| L. brevis | 94.9 ± 0.9% | 80.1 ± 0.5% * |
| L. amylolyticus | 94.7 ± 0.2% | 79.7 ±0.7% * |
| L. parabuchneri | 94.7 ± 0.0% * | 81.7 ± 0.2% * |
| L. sakei subsp. carnosus | 94.2 ± 0.4% | 80.6 ± 1.5% |
| S. xylosus | 95.3 ± 1.1% | 80.1 ± 1.3% |
| L. helveticus | 94.7 ± 0.2% | 78.5 ± 0.4% * |
| L. delbrueckii | 92.8 ± 1.4% | 78.5 ± 1.7% |
| (a) | |||||||||
| CFU (CFU/mL) | |||||||||
| 0 h | 4 h | 18 h | 24 h | ΔECFU 1 | |||||
| L. reuteri | 2.68 × 106 ± 2.40 × 105 a | 6.52 × 106 ± 2.02 × 106 a | 5.77 × 107 ± 5.16 × 106 a | 1.63 × 107 ± 2.83 × 106 a | 1.36 × 107 ± 3.07 × 106 a | ||||
| L. brevis | 1.83 × 107 ± 2.33 × 106 a | 4.20 × 107 ± 8.91 × 106 a | 1.24 × 108 ± 3.75 × 107 a | 1.31 × 108 ± 1.16 × 108 a | 1.13 × 108 ± 1.19 × 108 a | ||||
| L. amylolyticus | 1.38 × 107 ± 7.38 × 106 a | 4.39 × 106 ± 1.11 × 106 a | 5.77 × 107 ± 2.33 × 106 a | 5.88 × 107 ± 4.60 × 106 a | 4.50 × 107 ± 1.20 × 107 a | ||||
| L. parabuchneri | 1.21 × 107 ± 3.39 × 106 a | 2.30 × 107 ± 1.00 × 107 a | 3.43 × 107 ± 7.07 × 105 a | 5.91 × 107 ± 4.62 × 107 a | 4.70 × 107 ± 4.29 × 107 a | ||||
| L. sakei subsp. carnosus | 1.95 × 107 ± 9.19 × 106 a | 1.74 × 107 ± 5.09 × 106 a | 1.33 × 107 ± 3.32 × 106 a | 3.99 × 107 ± 1.56 × 106 a | 2.04 × 107 ± 7.64 × 106 a | ||||
| S. xylosus | 1.24 × 107 ± 1.54 × 107 a | 3.13 × 107 ± 1.27 × 107 a | 1.86 × 108 ± 1.91 × 108 a | 6.13 × 108 ± 7.59 × 108 a | 6.01 × 108 ± 7.75 × 108 a | ||||
| L. helveticus | 3.08 × 107 ± 1.25 × 107 a | 3.67 × 107 ± 2.03 × 107 a | 5.09 × 107 ± 1.12 × 107 a | 7.44 × 107 ± 1.94 × 107 a | 4.37 × 107 ± 3.19 × 107 a | ||||
| L. delbrueckii | 3.10 × 106 ± 9.97 × 105 a | 3.01 × 106 ± 1.77 × 105 a | 4.34 × 107 ± 1.51 × 107 a | 1.20 × 108 ± 1.84 × 107 a | 1.17 × 108 ± 1.74 × 107 a | ||||
| (b) | |||||||||
| pH | Glucose (g/kg) | ||||||||
| 0 h | 4 h | 18 h | 24 h | 0 h | 4 h | 18 h | 24 h | ΔEGlucose 2 | |
| L. reuteri | 6.5 ± 0.0 b | 6.4 ± 0.0 c,d,e | 4.9 ± 0.0 b,c,d | 4.8 ± 0.0 b,c | 5.0 ± 0.0 a | 4.1 ± 0.2 a,b,c | 0.9 ± 0.1 a | 0.2 ± 0.2 a | 4.8 ± 0.0 d |
| L. brevis | 6.6 ± 0.0 b,c | 6.5 ± 0.1 d,e | 5.3 ± 0.4 c,d,e | 5.0 ± 0.3 b,c | 5.0 ± 0.0 a | 4.7 ± 0.3 d | 3.7 ± 1.1 e | 0.9 ± 0.3 d | 4.1 ± 0.2 b |
| L. amylolyticus | 6.6 ± 0.0 c | 6.2 ± 0.0 c | 5.6 ± 0.3 e | 5.2 ± 0.1 c | 5.0 ± 0.0 a | 4.1 ± 0.1 a,b,c | 2.5 ± 0.0 c,d | 1.7 ± 0.3 e | 3.4 ± 0.1 a |
| L. parabuchneri | 6.6 ± 0.0 c | 6.5 ± 0.0 d,e | 5.5 ± 0.0 d,e | 4.9 ± 0.0 b,c | 5.0 ± 0.0 a | 4.3 ± 0.2 b,c,d | 3.3 ± 0.3 d,e | 0.7 ± 0.1 c,d | 4.3 ± 0.1 b,c |
| L. sakei subsp. carnosus | 6.6 ± 0.0 c | 6.3 ± 0.0 c,d | 4.7 ± 0.0 a,b,c | 4.7 ± 0.0 b | 5.0 ± 0.0 a | 4.4 ± 0.4 c,d | 1.4 ± 0.0 a,b | 0.4 ± 0.1 a,b | 4.6 ± 0.1 c,d |
| S. xylosus | 6.6 ± 0.0 c | 6.5 ± 0.0 e | 4.8 ± 0.1 b,c | 4.8 ± 0.0 b,c | 5.0 ± 0.0 a | 4.3 ± 0.2 b,c,d | 1.4 ± 0.3 a,b | 0.5 ± 0.4 a,b,c | 4.5 ± 0.1 c,d |
| L. helveticus | 6.0 ± 0.0 a | 5.4 ± 0.1 a | 4.1 ± 0.2 a | 3.9 ± 0.2 a | 5.0 ± 0.0 a | 3.8 ± 0.1 a | 1.8 ± 0.2 b | 0.6 ± 0.1 b,c,d | 4.4 ± 0.0 b,c |
| L. delbrueckii | 6.0 ± 0.1 a | 6.0 ± 0.0 b | 4.2 ± 0.1 a,b | 4.4 ± 0.0 a | 5.0 ± 0.0 a | 4.0 ± 0.4 a,b | 2.1 ± 0.2 b,c | 0.8 ± 0.2 c,d | 4.2 ± 0.1 b,c |
| Samples | Protein Solubility | |
|---|---|---|
| pH 4 | pH 7 | |
| LPI (unfermented) | 7.31 ± 0.26 | 63.59 ± 3.04 |
| L. reuteri | 7.40 ± 0.69 | 42.35 ± 3.76 * |
| L. brevis | 7.41 ± 0.97 | 38.45 ± 2.87 * |
| L. amylolyticus | 7.13 ± 0.31 | 35.47 ± 3.16 * |
| L. parabuchneri | 8.01 ± 0.90 | 27.37 ± 4.00 * |
| L. sakei subsp. carnosus | 7.17 ± 1.01 | 37.40 ± 4.53 * |
| S. xylosus | 8.11 ± 0.77 * | 28.04 ± 3.01 * |
| L. helveticus | 5.92 ± 0.92 * | 23.57 ± 2.99 * |
| L. delbrueckii | 5.55 ± 0.41 * | 27.48 ± 2.51 * |
| Samples | Foam Activity (%) | Foam Stability (%) |
|---|---|---|
| LPI (unfermented) | 1613 ± 11 | 89 ± 3 |
| L. reuteri | 1646 ± 20 * | 87 ± 3 |
| L. brevis | 1683 ± 57 | 86 ± 6 |
| L. amylolyticus | 1688 ± 52 * | 94 ± 2 |
| L. parabuchneri | 1703 ± 25 * | 16 ± 5 * |
| L. sakei subsp. carnosus | 1670 ± 32 * | 83 ± 3 |
| S. xylosus | 1678 ± 23 * | 91 ± 1 |
| L. helveticus | 1698 ± 17 * | 20 ± 0 * |
| L. delbrueckii | 1652 ± 36 | 80 ± 0 * |
| Samples | Emulsifying Capacity (mg/mL) |
|---|---|
| LPI (unfermented) | 552.9 ± 9.8 |
| L. reuteri | 471.5 ± 10.7 * |
| L. brevis | 447.2 ± 27.4 * |
| L. amylolyticus | 564.1 ± 18.7 |
| L. parabuchneri | 347.7 ± 5.2 * |
| L. sakei subsp. carnosus | 407.2 ± 7.6 * |
| S. xylosus | 595.6 ± 55.6 |
| L. helveticus | 455.2 ± 51.9 |
| L. delbrueckii | 370.3 ± 6.8 * |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Schlegel, K.; Leidigkeit, A.; Eisner, P.; Schweiggert-Weisz, U. Technofunctional and Sensory Properties of Fermented Lupin Protein Isolates. Foods 2019, 8, 678. https://doi.org/10.3390/foods8120678
Schlegel K, Leidigkeit A, Eisner P, Schweiggert-Weisz U. Technofunctional and Sensory Properties of Fermented Lupin Protein Isolates. Foods. 2019; 8(12):678. https://doi.org/10.3390/foods8120678
Chicago/Turabian StyleSchlegel, Katharina, Anika Leidigkeit, Peter Eisner, and Ute Schweiggert-Weisz. 2019. "Technofunctional and Sensory Properties of Fermented Lupin Protein Isolates" Foods 8, no. 12: 678. https://doi.org/10.3390/foods8120678
APA StyleSchlegel, K., Leidigkeit, A., Eisner, P., & Schweiggert-Weisz, U. (2019). Technofunctional and Sensory Properties of Fermented Lupin Protein Isolates. Foods, 8(12), 678. https://doi.org/10.3390/foods8120678