Changes in Absolute Contents of Compounds Affecting the Taste and Nutritional Properties of the Flesh of Three Plum Species Throughout Development


 ,
,  , ,
, ,  and
and 
Abstract
1. Introduction
2. Materials and Methods
2.1. Chemicals
2.2. Plant Material
2.3. Measurement of Fresh and Dry Weights
2.4. Measurement of Flesh Firmness and Soluble Solids Content
2.5. Preparation of a Lyophilized Powder
2.6. Measurement of Sugar and Malate Content
2.7. Determination of Total Phenol Content (TPC)
2.8. Determination of In Vitro Antioxidant Activity
2.8.1. Ferric Reducing Antioxidant Power (Frap) Assay
2.8.2. Trolox Equivalent Antioxidant Capacity (Teac) Assay-ABTS
2.8.3. DPPH Free Radical-Scavenging Assay
2.9. HPLC Analysis of Phenol Compounds
2.10. Statistical Analysis
3. Results and Discussion
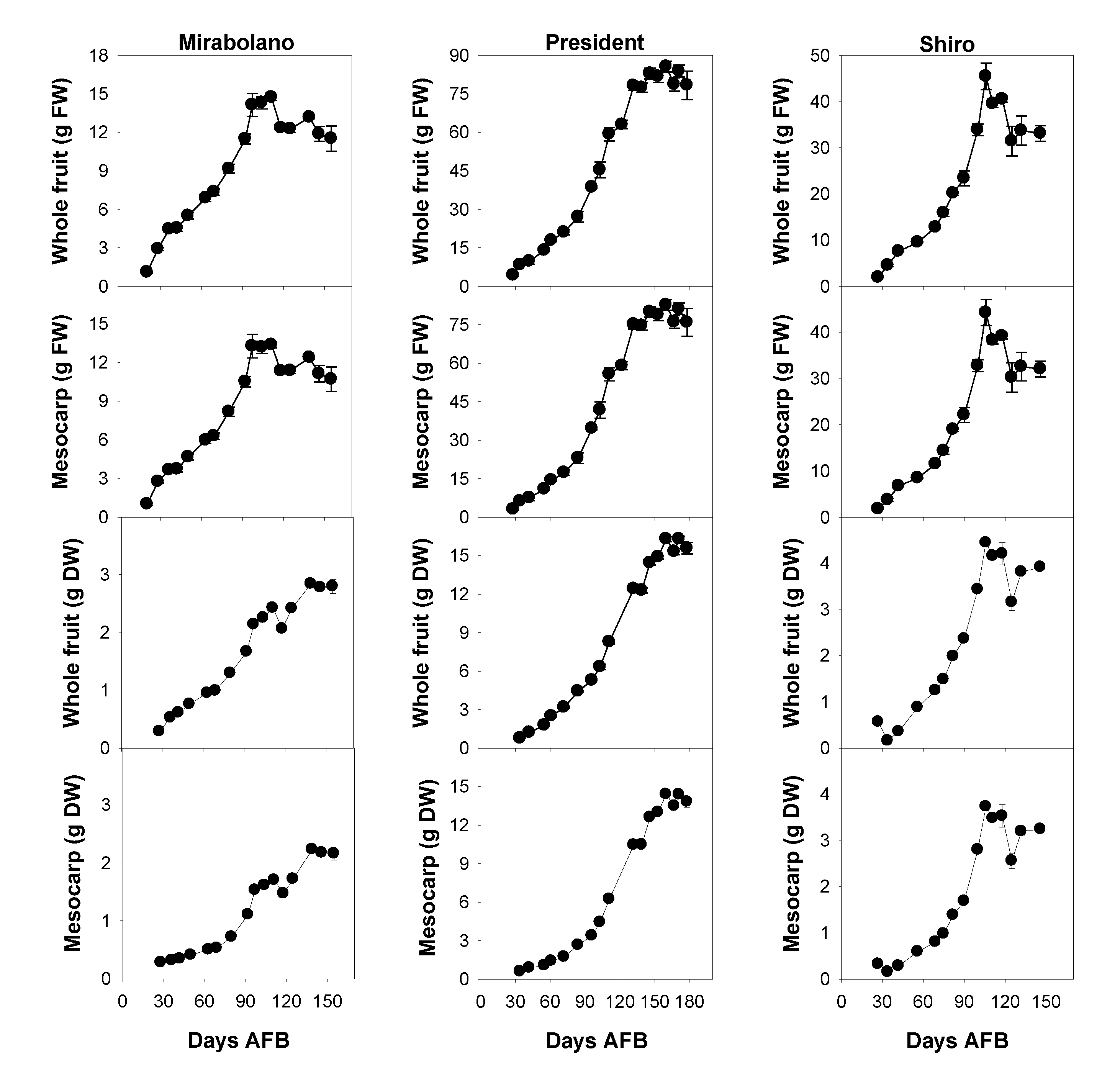
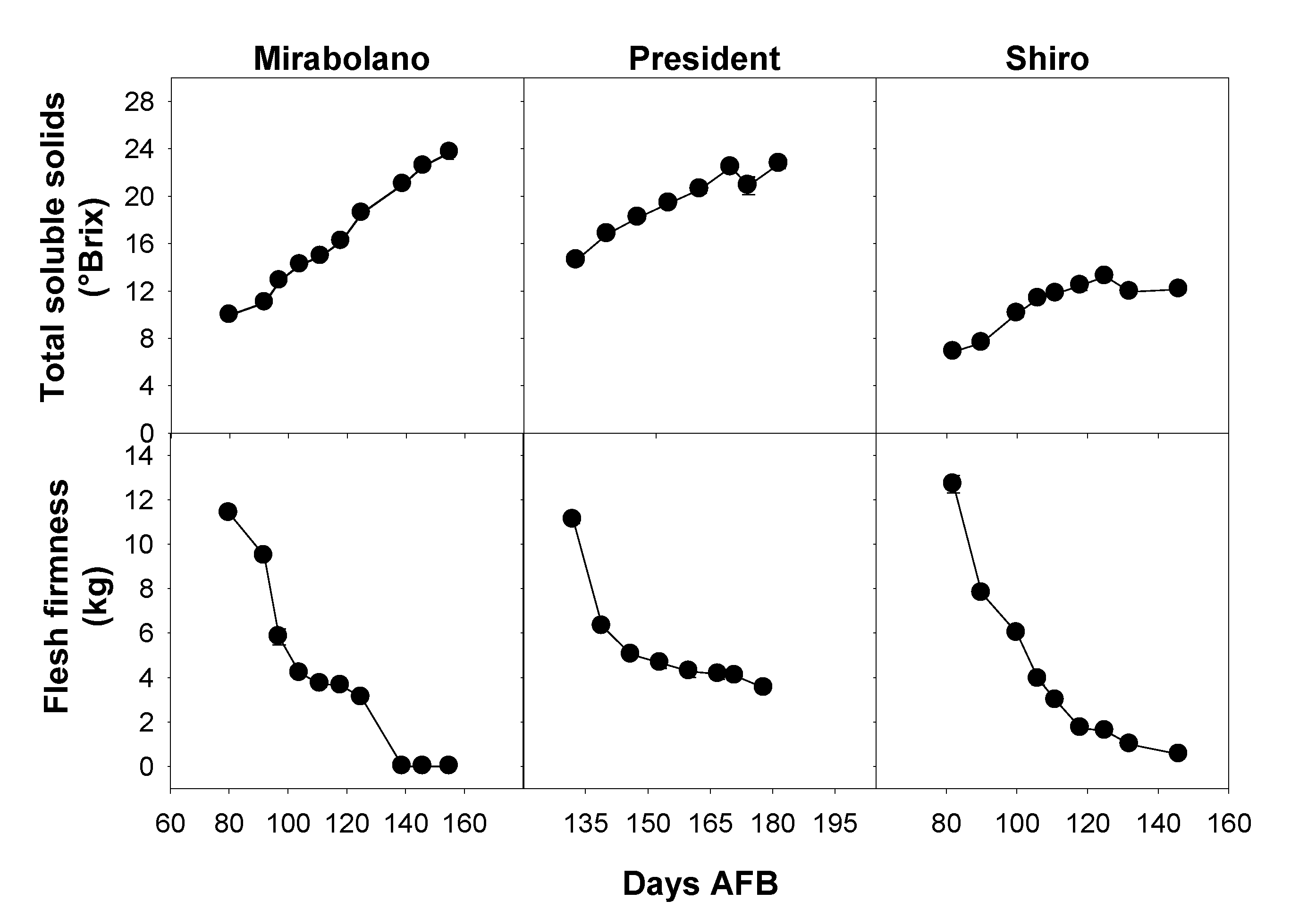
3.1. Physical and Chemical Characteristics of the Flesh during Development
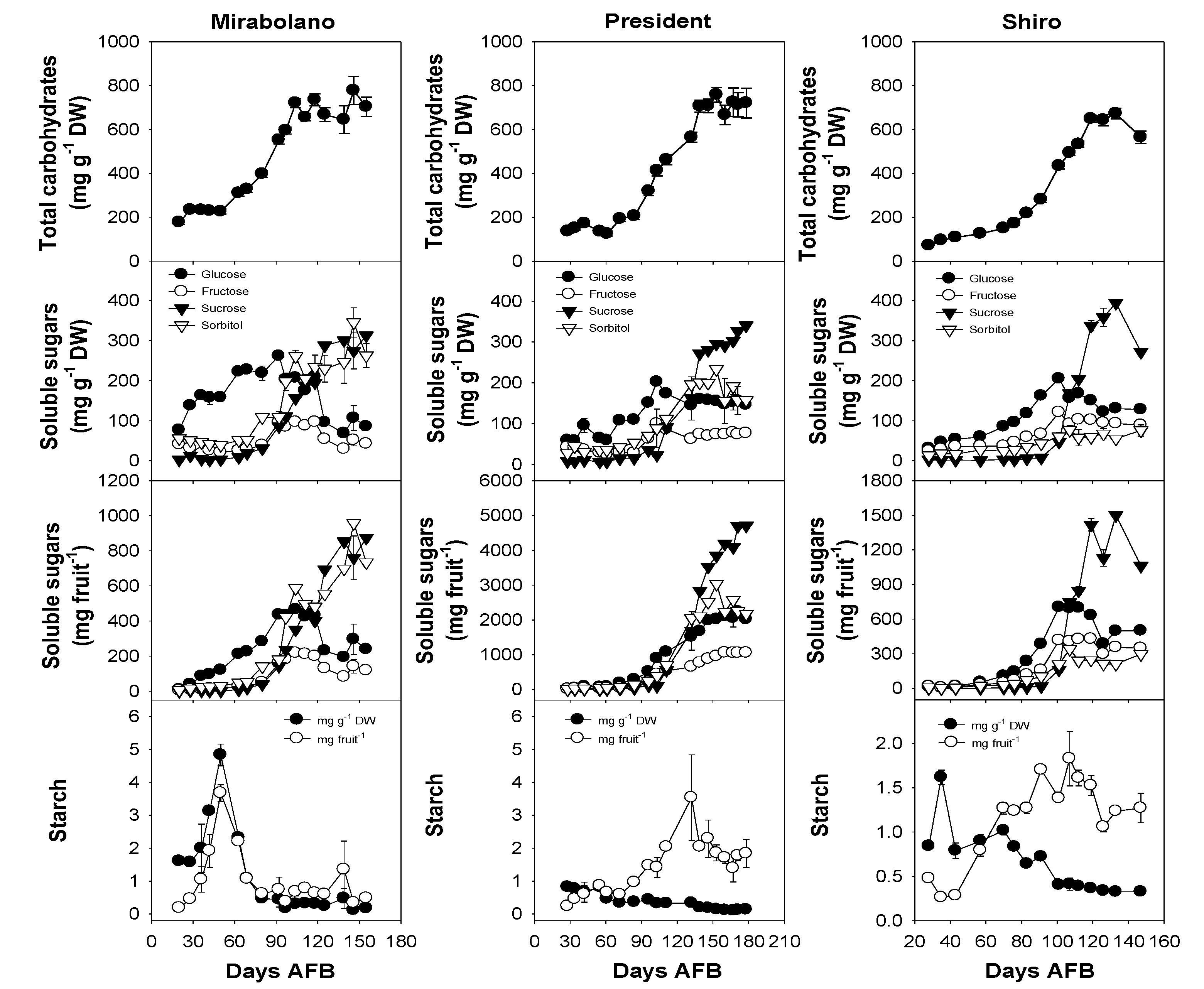
3.2. Carbohydrate Content of the Flesh during Development
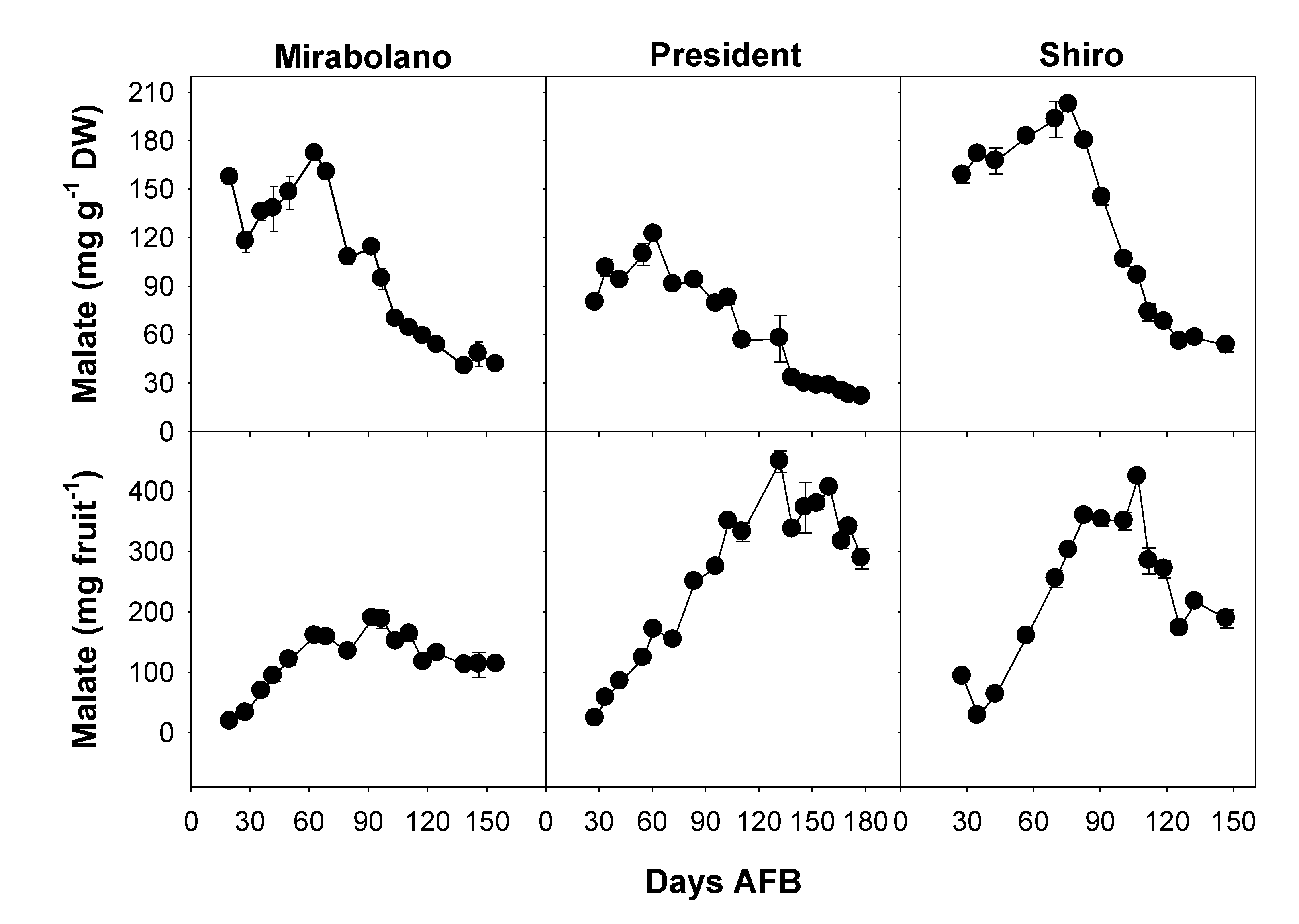
3.3. Malate Content of the Flesh during Development
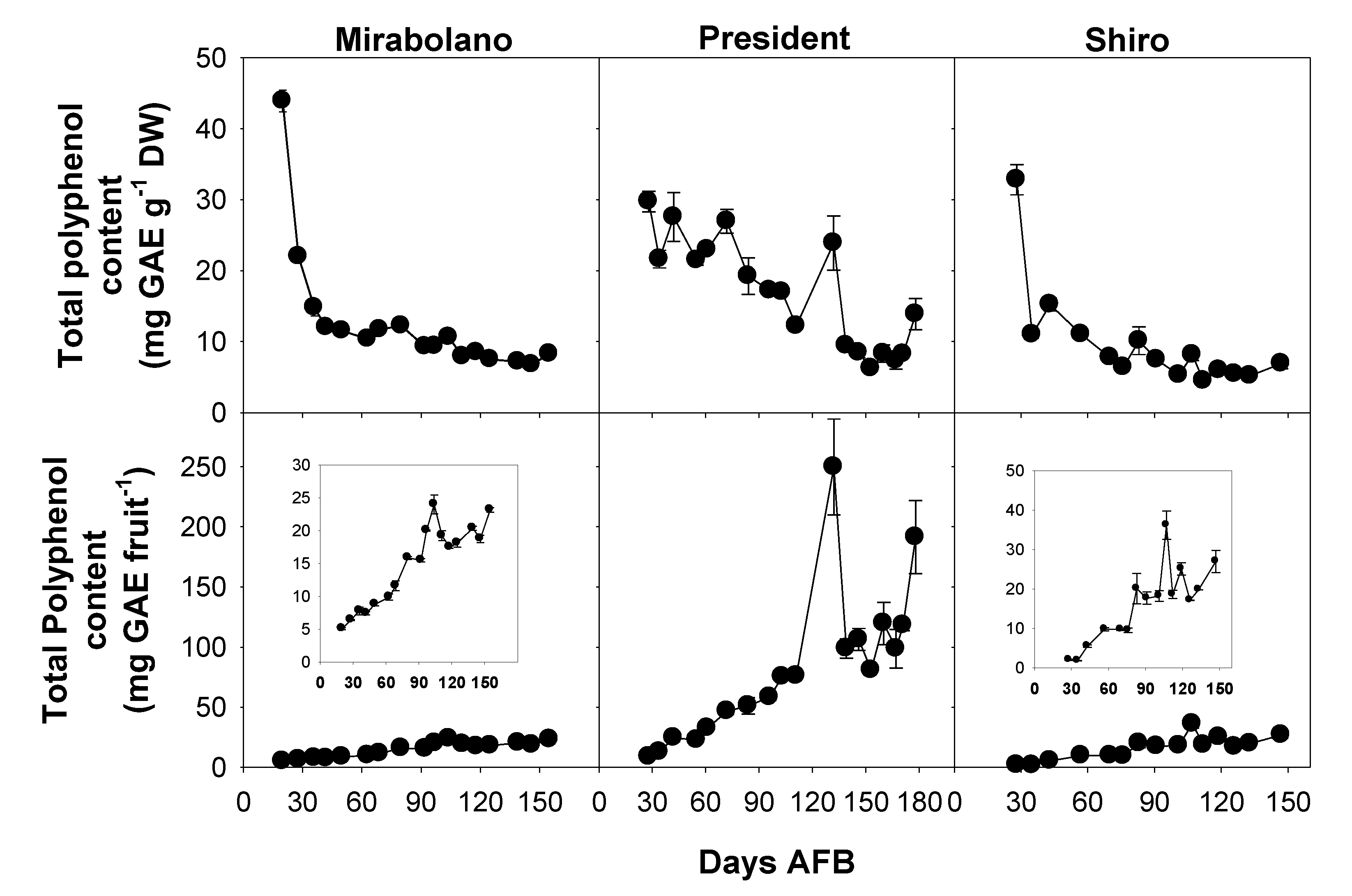
3.4. Determination of Total Phenol Content
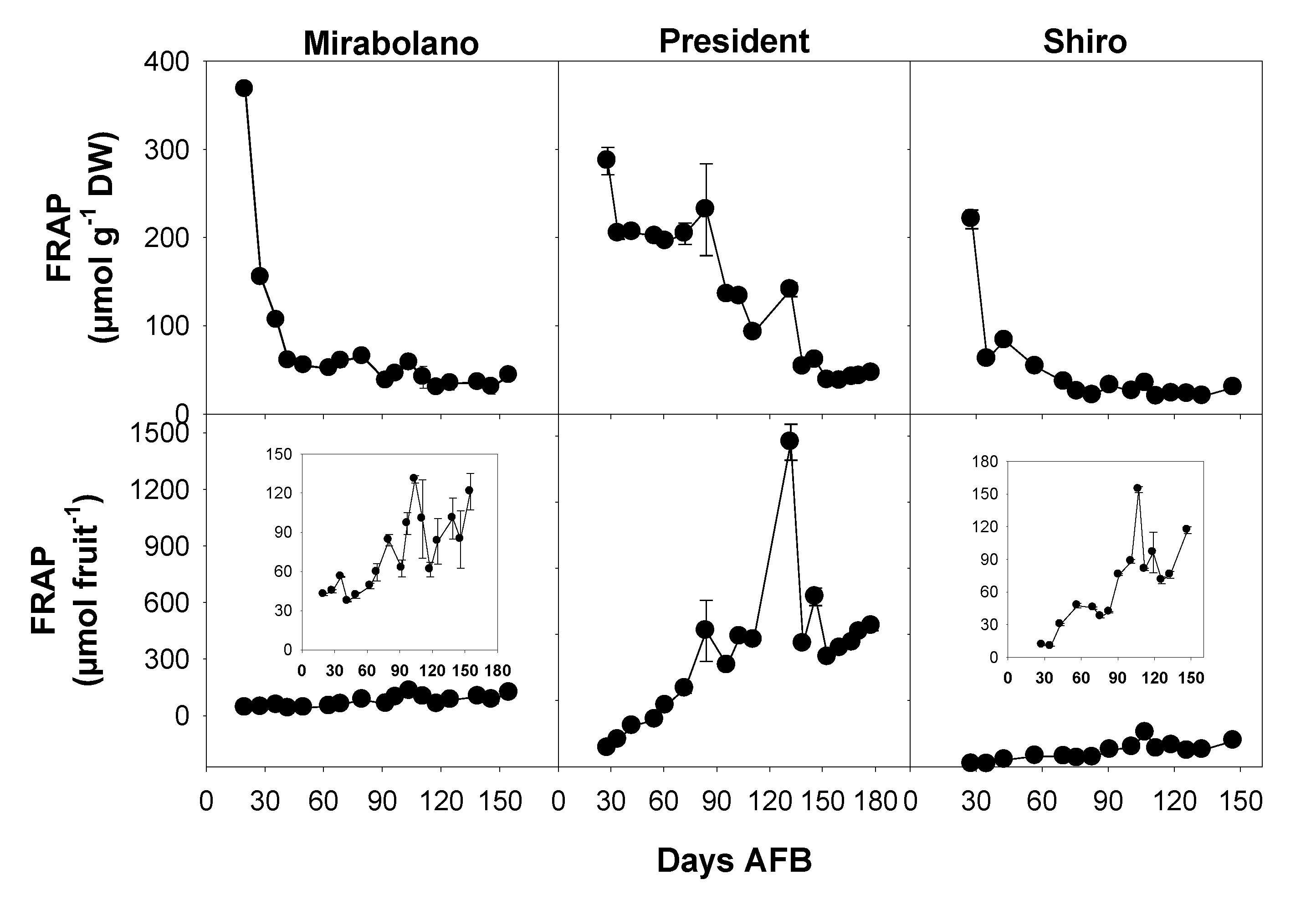
3.5. Evaluation of Antioxidant Activity
3.6. HPLC-DAD Analysis of Phenol Compounds
3.7. Phenolic Compounds and Antioxidant Activity: Correlations
3.8. Changes during Development, Including Over-Ripening
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Famiani, F.; Casulli, V.; Baldicchi, A.; Battistelli, A.; Moscatello, S.; Walker, R.P. Development and metabolism of the fruit and seed of the Japanese plum Ozark Premier (Rosaceae). J. Plant Physiol. 2012, 169, 551–560. [Google Scholar] [CrossRef] [PubMed]
- Lilleland, O. Growth study of the plum fruit-I. The growth and changes in chemical composition of the climax plum. Proc. Am. Soc. Hortic. Sci. 1933, 30, 203–208. [Google Scholar]
- Zuzunaga, M.; Serrano, M.; Martınez-Romero, D.; Valero, D.; Riquelme, F. Comparative study of two plum (Prunus salicina Lindl.) cultivars during growth and ripening. Food Sci. Technol. Int. 2001, 7, 123–130. [Google Scholar] [CrossRef]
- Sterling, C. Developmental anatomy of the fruit of Prunus domestica L. Bull. Torrey Bot. Club 1953, 80, 455–577. [Google Scholar] [CrossRef]
- Brady, C.J. Stone fruit. In Biochemistry of Fruit Ripening; Seymour, G.B., Taylor, J., Tucker, G.A., Eds.; Chapman & Hall: London, UK, 1993; pp. 379–404. [Google Scholar]
- Famiani, F.; Battistelli, A.; Moscatello, S.; Cruz-Castillo, J.G.; Walker, R.P. The organic acids that are accumulated in the flesh of fruits: Occurrence, metabolism and factors affecting their contents. Rev. Chapingo Ser. Hortic. 2015, 21, 97–128. [Google Scholar] [CrossRef]
- Walker, R.P.; Famiani, F. Organic acids in fruits: Metabolism, functions and contents. Hortic. Rev. 2018, 45, 371–430. [Google Scholar]
- Walker, R.P.; Battistelli, A.; Moscatello, S.; Chen, Z.H.; Leegood, R.C.; Famiani, F. Phosphoenolpyruvate carboxykinase in cherry (Prunus avium L.) fruit during development. J. Exp. Bot. 2011, 62, 5357–5365. [Google Scholar] [CrossRef] [PubMed]
- Ballistreri, G.; Continella, A.; Gentile, A.; Amenta, M.; Fabroni, S.; Rapisarda, P. Fruit quality and bioactive compounds relevant to human health of sweet cherry (Prunus avium L.) cultivars grown in Italy. Food Chem. 2013, 140, 630–638. [Google Scholar] [CrossRef]
- Famiani, F.; Farinelli, D.; Moscatello, S.; Battistelli, A.; Leegood, R.C.; Walker, R.P. The contribution of stored malate and citrate to the substrate requirements of metabolism of ripening peach (Prunus persica L. Batsch) flesh is negligible. Implications for the occurrence of phosphoenolpyruvate carboxykinase and gluconeogenesis. Plant Physiol. Biochem. 2016, 101, 33–42. [Google Scholar] [CrossRef] [PubMed]
- Montesano, D.; Cossignani, L.; Giua, L.; Urbani, E.; Simonetti, M.S.; Blasi, F. A simple HPLC-ELSD method for sugar analysis in Goji berry. J. Chem. 2016, 2016, 6271808. [Google Scholar] [CrossRef]
- Singh, S.; Singh, Z.; Swinny, E.W. Sugars and organic acids in Japanese plums (Prunus salicina Lindell) as influenced by maturation, harvest date, storage temperature and period. Int. J. Food Sci. Technol. 2009, 44, 1973–1982. [Google Scholar] [CrossRef]
- Cirilli, M.; Bassi, D.; Ciacciulli, A. Sugars in peach fruit: A breeding perspective. Hortic. Res. 2016, 3, 15067. [Google Scholar] [CrossRef] [PubMed]
- Baldicchi, A.; Farinelli, D.; Micheli, M.; Di Vaio, C.; Moscatello, S.; Battistelli, A.; Walker, R.P.; Famiani, F. Analysis of seed growth, fruit growth and composition and phospoenolpyruvate carboxykinase (PEPCK) occurrence in apricot (Prunus armeniaca L.). Sci. Hortic. 2015, 186, 38–46. [Google Scholar] [CrossRef]
- García-Mariño, N.; De la Torre, F.; Matilla, A.J. Organic acids and soluble sugars in edible and nonedible parts of damson plum (Prunus domestica L. subsp. insititia cv. Syriaca) fruits during development and ripening. Food Sci. Technol. Int. 2008, 14, 187–193. [Google Scholar] [CrossRef]
- Van Gorsel, H.; Li, C.; Kerbel, E.L.; Smits, M.; Kader, A.A. Compositional characterization of prune juice. J. Agric. Food Chem. 1992, 40, 784–789. [Google Scholar] [CrossRef]
- Moing, A.; Svanella, L.; Rolin, D.; Gaudillére, M.; Gaudillére, J.P.; Monet, R. Compositional changes during the fruit development of two peach cultivars differing in juice acidity. J. Am. Soc. Hortic. Sci. 1998, 123, 770–775. [Google Scholar] [CrossRef]
- Di Vaio, C.; Graziani, G.; Marra, L.; Cascone, A.; Ritieni, A. Antioxidant capacities, carotenoids and polyphenols evaluation of fresh and refrigerated peach and nectarine cultivars from Italy. Eur. Food Res. Technol. 2008, 227, 1225–1231. [Google Scholar] [CrossRef]
- Di Vaio, C.; Graziani, G.; Gaspari, A.; Scaglione, G.; Nocerino, S.; Ritieni, A. Essential oils content and antioxidant properties of peel ethanol extract in 18 lemon cultivars. Sci. Hortic. 2010, 126, 50–55. [Google Scholar] [CrossRef]
- Di Vaio, C.; Marallo, N.; Graziani, G.; Ritieni, A.; Di Matteo, A. Evaluation of fruit quality, bioactive compounds and total antioxidant activity of flat peach cultivars. J. Sci. Food Agric. 2015, 95, 2124–2131. [Google Scholar] [CrossRef]
- Lombardi, G.; Vannini, S.; Blasi, F.; Marcotullio, M.C.; Dominici, L.; Villarini, M.; Cossignani, L.; Moretti, M. In vitro safety/protection assessment of resveratrol and pterostilbene in a human hepatoma cell line (HepG2). Nat. Prod. Commun. 2015, 10, 1403–1408. [Google Scholar] [CrossRef]
- Di Matteo, A.; Russo, R.; Graziani, G.; Ritieni, A.; Di Vaio, C. Characterization of autochthonous sweet cherry cultivars (Prunus avium L.) of southern Italy for fruit quality, bioactive compounds and antioxidant activity. J. Sci. Food Agric. 2017, 97, 2782–2794. [Google Scholar] [CrossRef] [PubMed]
- Blasi, F.; Rocchetti, G.; Montesano, D.; Lucini, L.; Chiodelli, G.; Ghisoni, S.; Baccolo, G.; Simonetti, M.S.; Cossignani, L. Changes in extra-virgin olive oil added with Lycium barbarum L. carotenoids during frying: Chemical analyses and metabolomics approach. Food Res. Int. 2018, 105, 507–516. [Google Scholar] [CrossRef] [PubMed]
- Rocchetti, G.; Chiodelli, G.; Giuberti, G.; Ghisoni, S.; Baccolo, G.; Blasi, F.; Montesano, D.; Trevisan, M.; Lucini, L. UHPLC-ESI-QTOF-MS profile of polyphenols in Goji berries (Lycium barbarum L.) and its dynamics during in vitro gastrointestinal digestion and fermentation. J. Funct. Foods 2018, 40, 564–572. [Google Scholar] [CrossRef]
- Bertoldi, D.; Cossignani, L.; Blasi, F.; Perini, M.; Barbero, A.; Pianezze, S.; Montesano, D. Characterisation and geographical traceability of Italian Goji berries. Food Chem. 2019, 275, 585–593. [Google Scholar]
- Tomás-Barberán, F.A.; Espín, J.C. Phenolic compounds and related enzymes as determinants of quality in fruits and vegetables. J. Sci. Food Agric. 2001, 81, 853–876. [Google Scholar] [CrossRef]
- Díaz-Mula, H.M.; Zapata, P.J.; Guillén, F.; Castillo, S.; Martínez-Romero, D.; Valero, D.; Serrano, M. Changes in physicochemical and nutritive parameters and bioactive compounds during development and on-tree ripening of eight plum cultivars: A comparative study. J. Sci. Food Agric. 2008, 88, 2499–2507. [Google Scholar] [CrossRef]
- Huang, D. Dietary antioxidants and health promotion. Antioxidants 2018, 7, 9. [Google Scholar] [CrossRef] [PubMed]
- Jiang, C.C.; Fang, Z.Z.; Zhou, D.R.; Pan, S.L.; Ye, X.F. Changes in secondary metabolites: Organic acids and soluble sugars during the development of plum fruit cv. ‘Furongli’ (Prunus salicina Lindl). J. Sci. Food Agric. 2019, 99, 1010–1019. [Google Scholar] [CrossRef] [PubMed]
- Hummer, K.E.; Janick, J. Rosaceae: Taxonomy, economic importance, genomics. In Genetics and Genomics of Rosaceae; Springer: New York, NY, USA, 2009; pp. 1–17. [Google Scholar]
- Antognozzi, E.; Battistelli, A.; Famiani, F.; Moscatello, S.; Stanica, F.; Tombesi, A. Influence of CPPU on carbohydrate accumulation and metabolism in fruits of Actinidia deliciosa (A Chev). Sci. Hortic. 1996, 65, 37–47. [Google Scholar] [CrossRef]
- Proietti, S.; Moscatello, S.; Villani, F.; Mecucci, F.; Walker, R.P.; Famiani, F.; Battistelli, A. Quality and nutritional compounds of Prunus cerasus L. var. Austera fruit grown in central Italy. HortScience 2019, 54, 1005–1012. [Google Scholar] [CrossRef]
- Usenik, V.; Kastelec, D.; Veberič, R.; Štampar, F. Quality changes during ripening of plums (Prunus domestica L.). Food Chem. 2008, 111, 830–836. [Google Scholar] [CrossRef]
- Pagano, C.; Perioli, L.; Blasi, F.; Bastianini, M.; Chiesi, C.; Cossignani, L. Optimisation of phenol extraction from wine using layered double hydroxides and technological evaluation of the bioactive-rich powder. Int. J. Food Sci. Technol. 2017, 52, 2582–2588. [Google Scholar] [CrossRef]
- Rocchetti, G.; Blasi, F.; Montesano, D.; Ghisoni, S.; Marcotullio, M.C.; Sabatini, S.; Cossignani, L.; Lucini, L. Impact of conventional/non-conventional extraction methods on the untargeted phenolic profile of Moringa oleifera leaves. Food Res. Int. 2019, 115, 319–327. [Google Scholar] [CrossRef] [PubMed]
- Tawaha, K.; Alali, F.Q.; Gharaibeh, M.; Mohammad, M.; El-Elimat, T. Antioxidant activity and total phenolic content of selected Jordanian plant species. Food Chem. 2007, 104, 1372–1378. [Google Scholar] [CrossRef]
- Urbani, E.; Blasi, F.; Simonetti, M.S.; Chiesi, C.; Cossignani, L. Investigation on secondary metabolite content and antioxidant activity of commercial saffron powder. Eur. Food Res. Technol. 2016, 242, 987–993. [Google Scholar] [CrossRef]
- Blasi, F.; Urbani, E.; Simonetti, M.S.; Chiesi, C.; Cossignani, L. Seasonal variations in antioxidant compounds of Olea europaea leaves collected from different Italian cultivars. J. Appl. Bot. Food Qual. 2016, 89, 202–207. [Google Scholar]
- Lombardi, G.; Cossignani, L.; Giua, L.; Simonetti, M.S.; Maurizi, A.; Burini, G.; Coli, R.; Blasi, F. Phenol composition and antioxidant capacity of red wines produced in Central Italy changes after one-year storage. J. Appl. Bot. Food Qual. 2017, 90, 197–204. [Google Scholar]
- Pollini, L.; Rocchi, R.; Cossignani, L.; Mañes, J.; Compagnone, D.; Blasi, F. Phenol profiling and nutraceutical potential of Lycium spp. leaf extracts obtained with ultrasound and microwave assisted techniques. Antioxidants 2019, 8, 260. [Google Scholar] [CrossRef]
- Neri, F.; Pratella, G.C.; Brigati, S. Gli indici di maturazione per ottimizzare la qualità organolettica della frutta. Riv. Fruttic. Ortofloric. 2003, 5, 20–29. [Google Scholar]
- Famiani, F.; Baldicchi, A.; Farinelli, D.; Cruz-Castillo, J.G.; Marocchi, F.; Mastroleo, M.; Moscatello, S.; Proietti, S.; Battistelli, A. Yield affects qualitative kiwifruit characteristics and dry matter content may be an indicator of both quality and storability. Sci. Hortic. 2012, 146, 124–130. [Google Scholar] [CrossRef]
- Lombardi-Boccia, G.; Lucarini, M.; Lanzi, S.; Aguzzi, A.; Cappelloni, M. Nutrients and antioxidant molecules in yellow plums (Prunus domestica L.) from conventional and organic productions: A comparative study. J. Agric. Food Chem. 2004, 52, 90–94. [Google Scholar] [CrossRef]
- Selvaraj, Y.; Kumar, R.; Pal, D.K. Changes in sugars, organic acids, amino acids, lipid constituents and aroma characteristics of ripening mango fruit. J. Food Sci. Technol. 1989, 26, 306–311. [Google Scholar]
- Famiani, F.; Cultrera, N.G.M.; Battistelli, A.; Casulli, V.; Proietti, P.; Standardi, A.; Chen, Z.-H.; Leegood, R.C.; Walker, R.P. Phosphoenolpyruvate carboxykinase and its potential role in the catabolism of organic acids in the flesh of soft fruit during ripening. J. Exp. Bot. 2005, 56, 2959–2969. [Google Scholar] [CrossRef] [PubMed]
- Kaulmann, A.; Jonville, M.C.; Schneider, Y.J.; Hoffmann, L.; Bohn, T. Carotenoids, polyphenols and micronutrient profiles of Brassica oleraceae and plum varieties and their contribution to measures of total antioxidant capacity. Food Chem. 2014, 155, 240–250. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.; Chang, X.X.; Wang, H.; Brennan, C.S.; Guo, X.B.O. Phytochemicals accumulation in Sanhua plum (Prunus salicina L.) during fruit development and their potential use as antioxidants. J. Agric. Food Chem. 2019, 67, 2459–2466. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Chen, X.; Zhang, Y.; Chen, X. Antioxidant activities and major anthocyanins of Myrobalan plum (Prunus cerasifera Ehrh.). J. Food Sci. 2012, 77, C388–C393. [Google Scholar] [CrossRef] [PubMed]
- Venter, A.; Joubert, E.; de Beer, D. Characterisation of phenolic compounds in south African plum fruits (Prunus salicina Lindl.) using HPLC coupled with diode-array, fluorescence, mass spectrometry and on-line antioxidant detection. Molecules 2013, 18, 5072–5090. [Google Scholar] [CrossRef] [PubMed]
- Jang, G.H.; Kim, H.W.; Lee, M.K.; Jeong, S.Y.; Bak, A.R.; Lee, D.J.; Kim, J.B. Characterization and quantification of flavonoid glycosides in the Prunus genus by UPLC-DADQTOF/MS. Saudi J. Biol. Sci. 2018, 25, 1622–1631. [Google Scholar] [CrossRef] [PubMed]
- Ozturk, B.; Kucuker, E.; Karaman, S.; Ozkan, Y. The effects of cold storage and aminoethoxyvinylglycine (AVG) on bioactive compounds of plum fruit (Prunus salicina Lindell cv. ‘Black Amber’). Postharvest Biol. Technol. 2012, 72, 35–41. [Google Scholar] [CrossRef]
- Vangdal, E.; Slimestad, R. Methods to determine antioxidative capacity in fruit. J. Fruit Ornam. Plant Res. 2006, 14, 123–131. [Google Scholar]
- Connor, A.M.; Finn, C.E.; Alspach, P.A. Genotypic and environmental variation in antioxidant activity and total phenolic content among blackberry and hybrid berry cultivars. J. Am. Soc. Hortic. Sci. 2005, 130, 527–533. [Google Scholar] [CrossRef]
- Gil, M.I.; Tomás-Barberán, F.A.; Hess-Pierce, B.; Kader, A.A. Antioxidant capacities, phenolic compounds, carotenoids, and vitamin C contents of nectarine, peach, and plum cultivars from California. J. Agric. Food Chem. 2002, 50, 4976–4982. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Mirabolano | |||||
| M1 | M2 | M3 | M4 | M5 | |
| Days AFB * | 28 | 63 | 80 | 111 | 146 |
| Days AFB | President | ||||
| P1 | P2 | P3 | P4 | P5 | |
| 34 | 72 | 111 | 153 | 178 | |
| Days AFB | Shiro | ||||
| S1 | S2 | S3 | S4 | S5 | |
| 35 | 57 | 91 | 107 | 133 | |
| Plum Species | Samples * | ABTS | ABTS | DPPH |
|---|---|---|---|---|
| mg TE g−1 | mg TE fruit−1 | IC50 | ||
| Mirabolano | M1 | 18.10 ± 0.07 | 5.25 ± 0.02 | 7.49 ± 0.02 |
| M2 | 7.03 ± 0.16 | 6.69 ± 0.16 | 22.78 ± 0.38 | |
| M3 | 6.04 ± 0.52 | 7.82 ± 0.68 | 31.11 ± 1.31 | |
| M4 | 4.69 ± 0.08 | 11.36 ± 0.18 | 76.46 ± 2.52 | |
| M5 | 6.44 ± 0.12 | 17.86 ± 0.34 | 56.22 ± 0.61 | |
| President | P1 | 15.16 ± 0.88 | 2.45 ± 0.14 | 8.54 ± 0.39 |
| P2 | 15.53 ± 0.26 | 13.69 ± 0.23 | 4.22 ± 0.16 | |
| P3 | 7.27 ± 0.14 | 17.15 ± 0.33 | 15.56 ± 1.02 | |
| P4 | 5.54 ± 0.99 | 24.57 ± 4.38 | 30.35 ± 1.71 | |
| P5 | 4.92 ± 0.61 | 18.76 ± 2.31 | 28.63 ± 2.40 | |
| Shiro | S1 | 10.40 ± 0.04 | 6.16 ± 0.03 | 3.35 ± 0.71 |
| S2 | 7.96 ± 0.14 | 13.79 ± 0.24 | 12.48 ± 0.76 | |
| S3 | 3.61 ± 0.07 | 22.51 ± 0.44 | 42.28 ± 1.41 | |
| S4 | 3.57 ± 0.01 | 46.48 ± 0.11 | 75.50 ± 0.12 | |
| S5 | 4.55 ± 0.46 | 62.77 ± 6.35 | 56.51 ± 4.41 |
| μg g−1 | Neochlorogenic Acid | Catechin | Chlorogenic Acid | Chlorogenic Acid Derivatives § | Quercetin-3-O-Glucosides | Kaempferol-3-O-Glucosides | Quercetin | Kaempferol |
|---|---|---|---|---|---|---|---|---|
| M1 | 5432.75 ± 92.53 | 2645.76 ± 53.30 | 2250.68 ± 25.73 | 2076.06 ± 1.03 | nd | 45.99 ± 1.30 | nd | nd |
| M2 | 2188.59 ± 29.66 | 444.40 ± 16.77 | 1354.11 ± 5.45 | 903.20 ± 1.33 | nd | 40.99 ± 0.75 | nd | nd |
| M3 | 1490.47 ± 3.50 | 215.18 ± 8.17 | 949.87 ± 8.19 | 664.35 ± 16.41 | 23.11 ± 0.44 | 33.80 ± 0.66 | nd | nd |
| M4 | 1093.74 ± 3.28 | 161.37 ± 2.71 | 832.94 ± 3.84 | 605.16 ± 14.69 | 143.51 ± 1.83 | 25.38 ± 0.70 | nd | nd |
| M5 | 980.55 ± 1.04 | 67.01 ± 1.03 | 784.78 ± 4.19 | 370.46 ± 2.24 | 184.00 ± 2.82 | 27.22 ± 0.30 | nd | nd |
| P1 | 9643.08 ± 25.77 | 2873.01 ± 16.90 | 4319.22 ± 53.55 | 2955.25 ± 30.51 | 206.04 ± 2.16 | nd | nd | nd |
| P2 | 7751.09 ± 99.35 | 2471.09 ± 14.53 | 2151.47 ± 13.74 | 2558.12 ± 2.35 | 46.89 ± 0.20 | nd | nd | nd |
| P3 | 3129.33 ± 12.37 | 969.02 ± 15.45 | 1323.68 ± 9.68 | 1236.08 ± 20.53 | 45.80 ± 0.67 | nd | nd | nd |
| P4 | 1993.27 ± 21.94 | 469.56 ± 4.03 | 795.62 ± 1.91 | 763.47 ± 18.69 | 41.19 ± 0.39 | nd | nd | nd |
| P5 | 1972.11 ± 2.81 | 439.70 ± 1.41 | 826.88 ± 5.11 | 615.91 ± 4.51 | 74.28 ± 0.41 | nd | nd | nd |
| S1 | 1452.55 ± 9.93 | 987.81 ± 12.32 | 1037.22 ± 7.84 | 523.46 ± 5.13 | 471.65 ± 13.70 | 285.07 ± 7.19 | 101.40 ± 0.20 | 306.81 ± 20.21 |
| S2 | 1281.14 ± 1.76 | 454.64 ± 8.89 | 1017.91 ± 1.44 | 563.34 ± 1.51 | 350.11 ± 30.11 | 186.64 ± 9.95 | 91.82 ± 0.51 | 224.38 ± 1.09 |
| S3 | 773.95 ± 0.80 | 398.96 ± 0.59 | 736.46 ± 3.46 | 197.97 ± 0.29 | 133.17 ± 0.97 | 51.45 ± 5.21 | nd | 126.39 ± 2.15 |
| S4 | 704.74 ± 0.41 | 128.86 ± 1.38 | 684.88 ± 3.36 | 162.41 ± 0.26 | 51.73 ± 0.29 | 9.02 ± 0.06 | nd | 56.79 ± 0.58 |
| S5 | 702.73 ± 2.23 | 439.55 ± 4.29 | 728.38 ± 1.92 | 146.99 ± 1.67 | 141.06 ± 9.33 | 65.63 ± 8.28 | 10.12 ± 1.01 | 47.25 ± 0.11 |
| μg fruit−1 | Neochlorogenic Acid | Catechin | Chlorogenic Acid | Chlorogenic Acid Derivatives § | Quercetin-3-O-Glucosides | Kaempferol-3-O-Glucosides | Quercetin | Kaempferol |
|---|---|---|---|---|---|---|---|---|
| M1 | 1576.04 ± 32.97 | 767.54 ± 18.99 | 652.92 ± 9.17 | 602.26 ± 0.37 | nd | 13.34 ± 0.46 | nd | nd |
| M2 | 2082.00 ± 34.66 | 422.76 ± 19.60 | 1288.17 ± 6.37 | 262.02 ± 0.47 | nd | 38.99 ± 0.88 | nd | nd |
| M3 | 1930.46 ± 5.58 | 278.70 ± 13.01 | 1230.27 ± 13.03 | 192.73 ± 5.85 | 29.93 ± 0.69 | 43.78 ± 1.05 | nd | nd |
| M4 | 2649.69 ± 9.75 | 390.95 ± 8.06 | 2017.87 ± 11.43 | 175.56 ± 5.23 | 347.67 ± 5.45 | 61.48 ± 2.10 | nd | nd |
| M5 | 2720.54 ± 3.55 | 185.92 ± 3.51 | 2188.98 ± 14.27 | 107.47 ± 0.80 | 510.51 ± 9.61 | 75.53 ± 1.01 | nd | nd |
| P1 | 5716.42 ± 18.76 | 1703.12 ± 12.31 | 2560.43 ± 38.99 | 1751.87 ± 22.22 | 122.14 ± 1.57 | nd | nd | nd |
| P2 | 13424.89 ± 211.38 | 4279.93 ± 30.91 | 3726.35 ± 29.24 | 1516.45 ± 1.71 | 81.21 ± 0.43 | nd | nd | nd |
| P3 | 19500.09 ± 94.72 | 6038.34 ± 118.27 | 8248.39 ± 74.07 | 732.75 ± 14.95 | 285.42 ± 5.12 | nd | nd | nd |
| P4 | 25917.90 ± 350.43 | 6105.60 ± 64.32 | 10345.19 ± 30.50 | 452.59 ± 13.61 | 535.55 ± 6.26 | nd | nd | nd |
| P5 | 27236.76 ± 47.66 | 6072.68 ± 23.90 | 11400.52 ± 86.64 | 365.11 ± 3.29 | 1025.85 ± 6.87 | nd | nd | nd |
| S1 | 234.73 ± 1.97 | 159.63 ± 2.45 | 167.62 ± 1.56 | 84.59 ± 1.02 | 76.22 ± 2.72 | 46.07 ± 1.43 | 16.39 ± 0.03 | 49.58 ± 4.01 |
| S2 | 1129.58 ± 1.90 | 400.86 ± 9.63 | 897.49 ± 1.56 | 91.04 ± 0.30 | 308.69 ± 32.61 | 164.56 ± 10.77 | 80.96 ± 0.46 | 197.84 ± 1.18 |
| S3 | 1826.98 ± 2.33 | 941.79 ± 1.73 | 1738.50 ± 10.03 | 31.99 ± 0.06 | 314.35 ± 2.80 | 121.44 ± 15.10 | nd | 298.35 ± 6.24 |
| S4 | 3127.20 ± 2.22 | 571.82 ± 7.51 | 3039.10 ± 18.31 | 26.25 ± 0.05 | 229.56 ± 1.61 | 40.00 ± 0.33 | nd | 252.00 ± 3.15 |
| S5 | 2677.35 ± 10.41 | 1674.63 ± 20.08 | 2775.05 ± 8.96 | 23.75 ± 0.33 | 2442.39 ± 43.65 | 1012.02 ± 38.74 | 419.53 ± 3.86 | 180.01 ± 0.53 |
| Plum Species | TPC/ABTS | TPC/DPPH | TPC/FRAP | ABTS/DPPH | ABTS/FRAP | DPPH/FRAP |
| Mirabolano | 0.88 | 0.91 | 0.99 | 0.81 | 0.92 | 0.66 |
| President | 0.95 | 0.99 | 0.96 | 0.94 | 0.99 | 0.98 |
| Shiro | 0.63 | 0.82 | 0.95 | 0.82 | 0.79 | 0.80 |
| Plum Species | TPH/ABTS | TPH/DPPH | TPH/FRAP | NA/ABTS | NA/DPPH | NA/FRAP |
| Mirabolano | 0.98 | 0.95 | 0.97 | 0.96 | 0.95 | 0.95 |
| President | 0.96 | 0.76 | 0.96 | 0.95 | 0.80 | 0.95 |
| Shiro | 0.99 | 0.98 | 0.91 | 0.95 | 1.00 | 0.91 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Moscatello, S.; Frioni, T.; Blasi, F.; Proietti, S.; Pollini, L.; Verducci, G.; Rosati, A.; Walker, R.P.; Battistelli, A.; Cossignani, L.; et al. Changes in Absolute Contents of Compounds Affecting the Taste and Nutritional Properties of the Flesh of Three Plum Species Throughout Development. Foods 2019, 8, 486. https://doi.org/10.3390/foods8100486
Moscatello S, Frioni T, Blasi F, Proietti S, Pollini L, Verducci G, Rosati A, Walker RP, Battistelli A, Cossignani L, et al. Changes in Absolute Contents of Compounds Affecting the Taste and Nutritional Properties of the Flesh of Three Plum Species Throughout Development. Foods. 2019; 8(10):486. https://doi.org/10.3390/foods8100486
Chicago/Turabian StyleMoscatello, Stefano, Tommaso Frioni, Francesca Blasi, Simona Proietti, Luna Pollini, Giuseppa Verducci, Adolfo Rosati, Robert P. Walker, Alberto Battistelli, Lina Cossignani, and et al. 2019. "Changes in Absolute Contents of Compounds Affecting the Taste and Nutritional Properties of the Flesh of Three Plum Species Throughout Development" Foods 8, no. 10: 486. https://doi.org/10.3390/foods8100486
APA StyleMoscatello, S., Frioni, T., Blasi, F., Proietti, S., Pollini, L., Verducci, G., Rosati, A., Walker, R. P., Battistelli, A., Cossignani, L., & Famiani, F. (2019). Changes in Absolute Contents of Compounds Affecting the Taste and Nutritional Properties of the Flesh of Three Plum Species Throughout Development. Foods, 8(10), 486. https://doi.org/10.3390/foods8100486