Abstract
Sourdough fermentation, a traditional biotechnology for making leavened baked goods, was almost completely replaced by the use of baker’s yeast and chemical leavening agents in the last century. Recently, it has been rediscovered by the scientific community, consumers, and producers, thanks to several effects on organoleptic, technological, nutritional, and functional features of cereal-based products. Acidification, proteolysis, and activation of endogenous enzymes cause several changes during sourdough fermentation, carried out by lactic acid bacteria and yeasts, which positively affect the overall quality of the baked goods. In particular, the hydrolysis of native proteins of the cereal flours may improve the functional features of baked goods. The wheat flour processed with fungal proteases and selected lactic acid bacteria was demonstrated to be safe for coeliac patients. This review article focuses on the biotechnologies that use selected sourdough lactic acid bacteria to potentially counteract the adverse reactions to gluten, and the risk of gluten contamination.
1. The Sourdough Fermentation
The use of sourdough as a natural starter for leavening goods is considered one of the oldest biotechnological processes in food fermentation [1]. Sourdough is a mixture of flour (e.g., wheat, rye), water, and other ingredients (e.g., NaCl) that is fermented by naturally occurring lactic acid bacteria (LAB) and yeasts. These microorganisms originate mainly from flours and processing equipment, but the resulting composition of the sourdough microbiota is determined by endogenous (e.g., chemical and enzyme composition of the flour) and exogenous (e.g., temperature, redox potential, water content, and duration of the fermentation process) factors [2]. In mature sourdoughs, LAB dominate, occurring at concentrations of >108 cfu/g, whereas the number of yeasts is commonly one/two logarithmic cycles lower [3]. Sourdough fermentation positively influences all aspects of baked goods’ quality such as texture, aroma, nutritional properties, and shelf life. Recently, sourdough has been successfully applied for the improvement of the quality of naturally gluten-free (GF) bread due to the complex metabolic activity of LAB. Moreover, novel biotechnologies based on sourdough fermentation have been proposed for the complete degradation of gluten in cereal flours, rendering them suitable for the production of innovative GF products. Both these aspects, which are rapidly evolving thanks to the scientific community and the food industry, are taken into account and described in this review.
2. The Sourdough Lactic Acid Bacteria
Microbiological studies have revealed that more than 50 species of LAB and more than 25 species of yeasts, especially belonging to the genera Saccharomyces and Candida, occur in mature sourdoughs. Sourdough is considered a unique food ecosystem in that it (i) selects for LAB strains that are adapted to its environment and (ii) harbours LAB communities specific to each sourdough [4,5,6,7]. Representative genera of sourdough LAB are Lactobacillus, Leuconostoc, Pediococcus, and Weissella [8]. The largest biodiversity was found within the genus Lactobacillus and a relatively high number of species was discovered recently [2,4,9,10,11,12,13,14]. Depending on the protocols used for sourdough propagation, various microbial consortia of mainly obligate and facultative hetero-fermentative LAB are found. Lactobacillus brevis, Lactobacillus fermentum, Lactobacillus paralimentarius, Lactobacillus plantarum, Lactobacillus pontis, and, especially, Lactobacillus sanfranciscensis, considered to be a key sourdough bacterium [14], are commonly isolated from traditional sourdoughs.
3. Sourdough Properties and Functions
Beyond its natural and additive-free image, it is generally accepted that sourdough has various positive effects when used for manufacturing baked goods. Compared to other leavening agents (e.g., baker’s yeast), it improves the texture, flavour, nutritional value, and shelf life of bread [15]. Notwithstanding the role of sourdough yeasts, the main metabolic properties of LAB determining the above effects are briefly described.
3.1. Texture and Structure
Depending on the level of lactic acidification, sourdough fermentation leads to an increase in bread extensibility, softness, and volume [16,17,18,19,20]. Overall, sourdough fermentation improves the gas retention in bread dough [2,18]. Acidification affects the solubility of structure-forming components like gluten, starch, and arabinoxylans, and positively interferes with the activity of endogenous enzymes [21]. Acidification affects the mixing behaviour of the dough: at low pH, a shorter mixing time and less stability than normal dough are achieved [21].
3.2. Flavour
The fermentation of soluble carbohydrates (e.g., maltose, glucose, and fructose), metabolism of nitrogenous compounds, and generation of volatile compounds by sourdough LAB directly or indirectly influence the flavour of baked goods. Beyond the Embden–Meyerhof–Parnas (EMP, facultative hetero-fermentative strains) and phosphogluconate (obligate hetero-fermentative strains) pathways, the use of external acceptors of electrons [22,23,24] or alternative energy sources [25], and the interactions with endogenous and exogenous enzymes [26] lead to different quotients of fermentation (molar ratio between lactic and acetic acids) that differently affect the flavour of baked goods. Overall, sourdough fermentation results in a large increase of free amino acids (FAAs), compared to the baker’s yeast process [7]. Proteolysis during sourdough fermentation includes the hydrolysis of proteins to intermediate-sized polypeptides and subsequent release of FAAs from polypeptides allowed by the LAB peptidase system [5,27]. Once liberated, FAAs contribute directly to flavour or are further subjected to chemical conversion during baking or enzymatic catabolism [28], thus leading to the synthesis of flavour volatile compounds. Within the catabolism of FAAs, the expression of the arginine deiminase (ADI) pathway in sourdough LAB [29] enhances the growth and tolerance to acid stress, and, especially, increases the synthesis of ornithine, which is the precursor of the 2-acetyl-pyrroline, responsible for the roasted note of the wheat bread crust [5].
Among the compounds having a key role in baked goods’ flavour formation, homo-fermentative LAB mainly synthesize diacetyl, acetaldehyde, and hexanal, while hetero-fermentative strains mainly produce ethyl acetate, alcohols, and aldehydes. Iso-alcohols with their respective aldehydes and ethyl-acetate are characteristic volatile compounds of yeast fermentation [30,31].
3.3. Nutrition
Sourdough fermentation modifies nutritional features of cereals by (i) improving texture and palatability of wholegrain and fibre-rich bread; (ii) stabilizing or increasing levels of bioactive compounds; (iii) decreasing starch bioavailability (low glycaemic index products); and (iv) improving mineral bioavailability [32]. Lactic acidification increases the levels of bioactive compounds (e.g., phenolic compounds) [33] and causes the degradation of phytate, increasing mineral bioavailability [34,35]. Furthermore, lactic acidification also increases the magnesium and phosphorus solubility [32] and has been found to be a protective factor for β-glucan in bread. Organic acids such as those produced during sourdough fermentation have also been shown to play a role in the postprandial glycaemic responses. The presence of lactic acid during heat treatment promotes interactions between starch and gluten, reducing starch bioavailability and, consequently, the glycaemic index of baked goods [36,37].
3.4. Shelf Life
The improvement of the loaf’s specific volume and crumb softness by sourdough fermentation have been associated with the decrease of the rate of bread going stale [15,16]. Starch molecules may be affected by enzymes synthesized by LAB, causing a variation in the retrogradation properties of the starch and decreasing the rate of going stale. Besides going stale, microbial spoilage by bacteria, and especially moulds, remain responsible for huge economic losses in the bakery industries. Acidification through sourdough fermentation has been shown to inhibit the endospore germination and growth of Bacillus sp. responsible for rope spoilage [38]. Besides various compounds (e.g., organic acids, hydrogen peroxide, diacetyl), sourdough LAB may inhibit the growth of other related micro-organisms by synthesizing bacteriocins, bacteriocin-like inhibitory substances (BLIS) [5,39], and low-molecular mass antibiotics such as the reutericyclin of L. reuteri LTH2584 [40]. A number of antifungal metabolites, e.g., a mixture of organic acids (acetic, caproic, and formic acids), cyclic dipeptides, phenyllactic acid, proteinaceous compounds, and 3-hydroxylated fatty acids, are potentially synthesized by LAB [41,42,43,44,45] acting against moulds responsible for bread spoilage. Different peptides with antifungal activity were identified in the water-soluble extracts of wheat flour fermented with LAB, as the results of the proteolytic activity on the native wheat proteins show [45,46]. Overall, all the peptides produced by LAB in wheat-based matrices were characterized by a large inhibitory spectrum against species that commonly contaminate baked goods and bakeries [45,46,47], allowed a long storage of bread (at least 21–28 days).
3.5. Functional Properties
During sourdough fermentation, LAB may also produce bioactive compounds such as peptides and amino acid derivatives (e.g., γ-amino butyric acid) with various functionalities, and potentially prebiotic exo-polysaccharides.
The potential of sourdough lactic acid bacteria to release lunasin, a strong anti-tumoural peptide, during fermentation of cereal flours, was recently exploited [48]. Recently, flours obtained from different legume species were subjected to fermentation with selected LAB strains, showing the release of lunasin-like polypeptides, as the consequence of the proteolysis of native proteins [49]. A marked inhibitory effect on the proliferation of human adenocarcinoma Caco-2 cells was observed using extracts from fermented legume doughs (up to 70%) [49].
The capacity of selected lactic acid bacteria to release antioxidant peptides was shown during the fermentation of various cereal flours. Purified peptides showed ex vivo antioxidant activity on mouse fibroblasts artificially subjected to oxidative stress [50].
Lactic acid bacteria selected for proteolytic activity were used for wheat and rye fermentation with the aim of producing anti-hypertensive peptides. A strong ACE-inhibitory activity was found when fermenting flours under semi-liquid conditions and, especially, when using whole wheat flour [51].
4. Adverse Reactions to Gluten
Coeliac disease (CD), also known as coeliac sprue and gluten-sensitive enteropathy, is a food hypersensitivity disorder caused by an inflammatory response to wheat gluten and similar proteins of barley and rye [52]. In recent years, the view of CD has undergone a profound revision. Nowadays, CD is considered more than a just a gluten-sensitive enteropathy but a systemic immune-mediated disorder elicited by gluten and related prolamines in genetically susceptible individuals. The common denominator for all subjects with CD is the presence of a variable combination of gluten-dependent clinical manifestations, specific autoantibodies (anti-tissue transglutaminase [TG]2, anti-endomysium [EMA] antibodies), HLA-DQ2, and/or DQ8 haplotypes, and different degrees of enteropathy, ranging from lymphocytic infiltration of the epithelium to complete villous atrophy [53]. Reports of CD date back to the first century AD [54], but it was not until 1888 that Samuel Gee gave the classical description of the disease [55], and it was only in the 1930s that Willem-Karel Dicke observed that removal of wheat from the diet alleviated the symptoms and signs of CD [56]. Nowadays, the prevalence of CD worldwide is increasing; it is estimated to be 0.5%–2.0% in most of the European countries and the United States. Such a rate establishes CD as one of the most common food intolerances [57]. Gluten may also induce other pathological conditions, such as wheat allergy (WA) [58], which is an immunoglobulin (Ig)E-mediated disease also well characterized from the immunological and clinical point of view but completely unrelated to CD. More recently, attention was given to another entity, gluten sensitivity (GS), for which the limits and possible overlap with CD are still poorly defined [59]. GS subjects are unable to tolerate gluten and develop an adverse reaction when eating gluten that usually, and differently from CD, does not lead to damage in the small intestine. A number of morphological, functional, and immunological disorders have been considered under the definition of GS that miss one or more of the key CD criteria (enteropathy, associated HLA haplotypes, and presence of anti-TG2 antibodies), but respond to gluten exclusion. Nowadays, the only effective treatment for CD consists of a lifelong gluten-free diet (GFD). The regression of symptoms in response to the GFD was also shown in WA and GS subjects. Nevertheless, gluten is a common, and in many countries unlabelled, ingredient in the human diet, presenting a big challenge for CD and WA patients, and GS subjects; therefore, there is an increasing need to develop safe and effective alternatives. Beyond genetic predisposition, several environmental factors influence adverse reactions to gluten. Recent epidemiological studies show that the introduction of gluten-containing grains, which occurred about 10,000 years ago with the advent of agriculture, represented an evolutionary challenge that created the conditions for human diseases related to gluten exposure [60]. More recently, cereal food technology has changed dramatically by influencing the daily diet of entire populations previously not exposed to high concentrations of gluten. Cereal baked goods are currently manufactured by a very accelerated process where long fermentations by sourdough, a cocktail of acidifying and proteolytic lactic acid bacteria with or without Saccharomyces cerevisiae, were almost totally replaced by the indiscriminate use of chemical and/or baker’s yeast leavening agents. Under these technological circumstances, cereal components (e.g., proteins) are subjected to very mild or absent degradation during manufacture, resulting, probably, in less digestible foods compared to traditional and ancient sourdough baked goods [7].
5. Applications of Sourdough in Gluten-Free Products
Gluten is one of the most important structure-building protein complexes responsible for the quality and structure of wheat-based products. Its viscoelastic properties render the development of GF dough having similar quality and structural properties a highly challenging task, and form a major industrial hurdle [61]. The development of GF products to date remains a technologically intriguing area for researchers as well as the food industry [61]. Indeed, dough produced from GF formulations lacks a cohesive and elastic nature due to the absence of gluten, which makes industrial handling of dough a greater challenge. However, in the recent past, various alternative approaches (e.g., high-pressure, extrusion, enzymatic treatments) have been adopted to modify functional attributes necessary for GF dough development. These approaches could provide potential solutions in the development of GF products for the coeliac community globally [61].
The sourdough biotechnology has been applied to the manufacture of gluten-free products with the aim of improving their sensory and nutritional features. Overall, the gluten-free products available on the market are of low quality, exhibiting poor mouth-feel and flavour [62].
Since they do not contain gluten, and are mainly starch-based, GF bread products go stale more rapidly than gluten-containing bread [63]. In addition, when limiting the use of gluten-free flours to the most common sources (e.g., rice, corn, and starches), nutrient deficiencies may occur due to the very low dietary fibre content and excess calories [64]. Nevertheless, the current literature indicates a limited number of papers dealing with the use of sourdough in gluten-free goods. The few available results indicate that sourdough has a positive effect on the baking quality, particularly on volume, texture, and flavour. The influence of sourdoughs fermented by different LAB strains on the textural quality of gluten-free bread was evaluated during storage and compared to that of chemically acidified or non-acidified doughs [17,18]. The growth of selected LAB in gluten-free batters was similar to that reported for wheat sourdoughs [18]. Sourdough fermentation caused an increase in the dough elasticity and delayed the process of going stale [65]. These effects were mainly attributed to the breakdown of non-gluten proteins and starch components by sourdough LAB. Based on triangle tests, gluten-free sourdough bread was discriminated from the control bread and clearly preferred. In a recent patent [66] L. sanfranciscensis LS40 and LS41, and L. plantarum CF1, previously isolated from traditional sourdoughs, were selected. This microbial mixture was used to ferment gluten-free ingredients (e.g., corn starch, rice, buckwheat, and millet flours) and compared to baker’s yeast fermentation. The sourdough fermentation allowed us to: (i) completely degrade about 300 ppm of gluten, eventually present as contaminant; (ii) increase by about 10-fold the concentration of FAAs; (iii) increase by about 10-fold the phytase activity during fermentation; and (iv) improve the sensory characteristics of the resulting bread as evaluated by descriptive analysis.
A Type-I GF sourdough was obtained using only naturally GF flours, through the typical backslopping procedure [67]. After few refreshments in controlled conditions, the sourdough presented a stable association between L. sanfranciscensis and Candida humilis, constant fermentation times, and technological properties (in terms of dough consistency, dough maximum height, CO2 production, and retention) [67]. The results showed that a traditional sourdough biotechnology can also be used to improve the overall quality of GF baked products [67].
The positive contribution of sourdough could be exploited for the design of high-quality GF bread from various GF cereals and pseudocereals [68]. For example, the effect of adding fresh and freeze-dried amaranth and buckwheat into the GF bread formula has recently been investigated [68,69], showing several advantages in taste and aroma [68]. Sourdoughs obtained with teff and buckwheat (through the use of a selected L. helveticus strain) were used for the making of experimental GF breads that were characterized by sensory analysis and sensory tesst showing enhanced bread aroma and increased fruity, cereal, and toasty notes [68]. Chestnut flour was subjected to a spontaneous fermentation and a typical backslopping procedure [70], obtaining a sourdough that was included in the formulation of corn-based GF bread, along with a volume increase, a decrease of the crust hardness, and a longer shelf life [70]. A Lactobacillus amylovorus strain was employed as a starter culture for gluten-free quinoa sourdough bread under pilot-plant conditions to extend the microbial shelf life [71]. It was demonstrated that the use of quinoa sourdough extended the mould-free shelf-life up to four days compared to the non-acidified control, thanks to the high concentration of 4-hydroxyphenyllactic acid, phloretic acid, 3-phenyllactic acid, and hydroferulic acid [71]. Evaluation of bread characteristics such as specific volume or crumb hardness revealed that the addition of L. amylovorus-fermented sourdough also improved bread quality [71].
Breads based on gluten-free buckwheat, quinoa, sorghum, and teff flours were produced with the addition of sourdough fermented with exopolysaccharide (EPS) producing Weissella cibaria MG1, showing that the acidification increased crumb porosity and decreased hardness [72]. Moreover, the authors reported that the staling rate was significantly reduced [72]. In any case, the use of sourdough decreased the degree of in vitro starch hydrolysis (and, consequently, the predicted glycaemic index) [73].
The increasing demand for high-quality gluten-free baked goods, clean labels, and natural products points to the need for new approaches in GF bread making [74]. Overall, the positive effects of sourdough, extensively studied for traditional baking, overlap those found using sourdough in GF baking. The microbiological and qualitative characterization suggests that the metabolic activities of the sourdough microbiota are still retained during fermentation of GF matrices [74]. Thus, due to the sensory, texture, and nutritional improvements, the large-scale industrial use of sourdough for the manufacture of gluten-free goods can be recommended.
6. Sourdough Lactic Acid Bacteria as a Tool for Detoxifying Gluten in Wheat-Based Foods
Beyond genetic predisposition, several environmental factors influence CD prevalence. Recent epidemiological studies show that, besides being frequently found in countries where individuals are mostly of European origin, CD is a common disorder in many areas of the developing world (the Middle East, North Africa, South and East Asia, and Latin America). As mentioned before, the modern food industry has replaced sourdough biotechnology with the large-scale use of chemical and/or baker’s yeast leavening agents [7]. Nevertheless, the traditional biotechnology of sourdough bread making has been recently exploited for the capacity to degrade toxic epitopes during food processing. This “food technological approach,” together with other methods aiming at the hydrolysis of toxic gluten peptides prior to ingestion, has been proposed and developed as an alternative to the hydrolysis of gluten peptides after ingestion in the gastrointestinal tract (the “medical approach”) [75]. Extensive research in this field is ongoing at the authors’ laboratory in a joint project with medicine specialists to show the potential of proteolytic enzymes of sourdough LAB as it has been widely shown for prolyl endopeptidases (PePs) of Flavobacterium meningosepticum [76], Myxococcus xanthus [77], and Aspergillus niger [78,79].
6.1. Use of Selected Lactic Acid Bacteria for Gluten Degradation
Since the last decade, several studies [80,81,82,83] have been carried out aiming at showing the capacity of proteolytic enzymes, mainly peptidases, of selected sourdough lactobacilli to degrade gluten during food processing. The use of pooled cell suspensions and cell-free extracts obtained by different lactic acid bacteria strains was investigated [84]. Although a number of in vitro (e.g., agglutination and Caco-2/TC assays), ex vivo (biopsy-derived T cells), and acute in vivo (intestinal permeability) tests were carried out, the above results [80,81,82,83] only showed a marked decrease of the gliadin fraction, but not a complete degradation. Recently, it was shown that a traditional sourdough fermentation, leading to a partial gluten hydrolysis of wheat flour proteins, is not able to prevent the interaction of transglutaminase 2 with α2-gliadin or gluten; thus it cannot be considered safe for making GF products [85]. This route might be helpful to eliminate the risk of cross-contamination of gluten-free products but not to completely eliminate the toxicity of wheat flour.
Nevertheless, further efforts were made to increase the hydrolysing capacity of sourdough LAB. Together with two fungal proteases (obtained from Aspergillus niger and A. oryzae), routinely used in bread making, other lactobacilli strains, characterized by a marked peptidase activity towards Pro-rich peptides [86], were used during long-time fermentation of semi-liquid wheat flour doughs. As determined by R5-sandwich and competitive ELISA, the residual concentration of gluten in the fermented sourdough was <20 ppm, as required by the standard of the Codex Alimentarius Commission for gluten-free products. Nevertheless, due to the limitation of the R5-ELISA methods, further investigations based on in vitro, ex vivo, and in vivo assays were performed in order to assess the complete degradation of all the protein epitopes involved in the pathology, including those of glutenins. Two-dimensional electrophoresis and MALDI-TOF mass spectrometry analyses showed the complete hydrolysis of albumins/globulins and gliadins [87]. After hydrolysis, the spray-dried flour from fermented sourdough was mainly a mixture of water/salt-soluble low molecular weight peptides and amino acids. Many chemical analyses and ex vivo tests on human cell cultures confirmed the complete detoxification of gluten [87,88]. After sourdough fermentation, the water was removed and the pre-treated wheat flour was used for bread making by using baker’s yeast and structuring agents (e.g., gums). Structuring ingredients are necessary since the resulting gluten network was completely disrupted [89]. This sourdough bread was compared to baker’s yeast bread made with non-treated flour and without structuring agents. The specific loaf volume of sourdough bread was similar to that of baker’s yeast bread and showed the typical flavour of the sourdough wheat bread, as judged by an internal panel test [87,88]. Recently, a bread made with semolina rendered GF by the protocol here described was compared to commercial GF breads made with naturally GF ingredients [90]. Beyond the huge potential of market expansion, the main advantages of wheat flour rendered GF are the high availability of FAAs, the high protein digestibility, the low starch hydrolysis index, and the better technological properties of bread compared to the commercial GF products currently on the market [90]. Vitamins, minerals, and dietary fibre profiles are comparable to those of gluten-containing wheat bread. The sensory profile, determined by a panel test, can be considered the most similar to those of conventional baked goods, showing all the classic attributes of sourdough bread [90].
6.2. Mechanisms of Detoxification and in Vivo Tests
Activity of fungal proteases was responsible for the primary proteolysis, liberating various sizes of polypeptides. The large proportion of proline residues in the amino acid sequences characterizing the toxic peptides make them extremely resistant to further hydrolysis [91,92,93]. The specific cyclic structure of the proline imposes many restrictions on the structural aspects of peptides. To adequately deal with such peptides, a group of specific peptidases is necessary to hydrolyse peptide bonds. Prolyl endopeptidases (PEPs) of microbial origin are endoproteolytic enzymes, which, in contrast to human gastrointestinal proteases, may readily cleave Pro-rich immune-stimulatory gluten peptides [93]. Through a complex system of ABC and ATP transporters, gluten peptides are moved across the cytoplasmic membrane of sourdough lactobacilli. Already a few minutes after entry, the concentration of polypeptides markedly decreases, to about 100 times lower than that of the environment [94]. A pool of intracellular peptidases of the selected sourdough lactobacilli was used to simulate hydrolysis towards the 33-mer epitope. The combined activity of peptidases was responsible for the complete degradation of the 33-mer or other synthetic immunogenic peptides, which occurred within 14 h of incubation [94]. Lactic acid bacteria possess a very complex peptidase system [95], although not a unique strain that may possess the entire pattern of peptidases needed for hydrolysing all the potential peptides where Pro is involved. Nevertheless, the hydrolysing capacity was lost when individual strains were tested, confirming that no single strain contains the entire portfolio of peptidases necessary to degrade Pro-rich polypeptides (Figure 1). Sweet baked goods were made using the complete hydrolysed wheat flour. Two clinical challenges were carried out on coeliac patients, who ingested the equivalent of about 8 or 10 g of native gluten per day for 60 days [96,97]. Haematology, serology, and intestinal permeability analyses showed complete tolerance by all coeliac patients during the whole time. None of the CD patients had clinical complaints and none produced anti-TG2 antibodies or had modification of the small intestinal mucosa compared to the pre-challenge situation [96]. No increase of CD3 and gamma delta cells was found, and the Marsh grade was unchanged after the challenge.
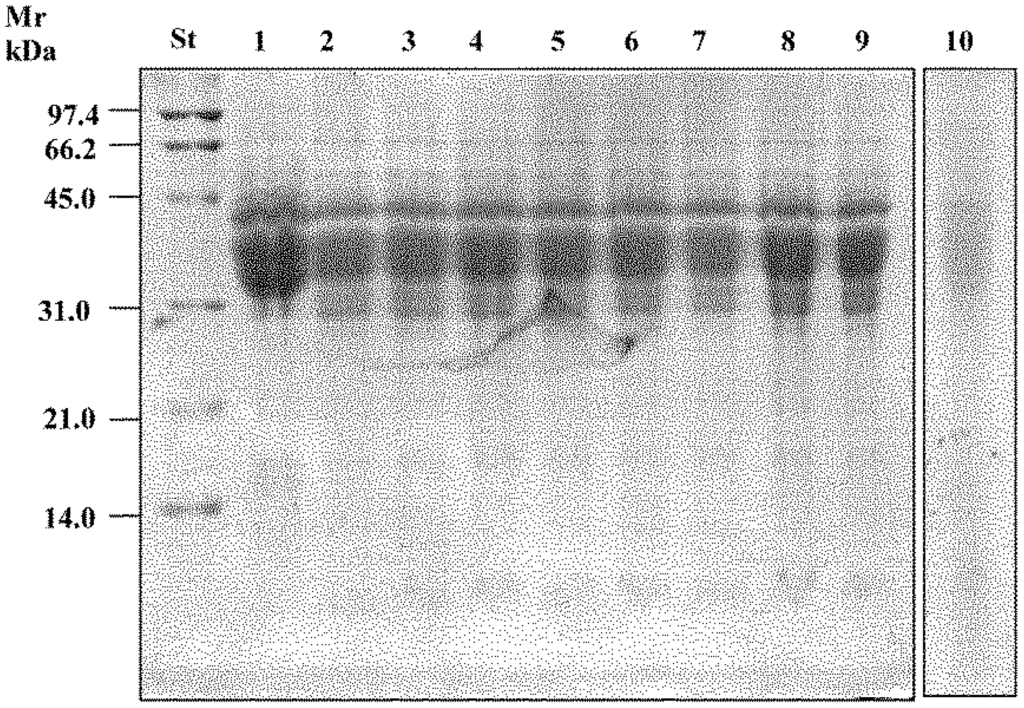
Figure 1.
SDS-PAGE analysis of gliadins polypeptides from wheat flour doughs incubated for 24 h with the different cell preparations (109 cfu/mL) that composed the VSL#3 preparation. Protein standard (St). Chemically acidified dough (1); doughs incubated with cells of Bifidobacterium longum (2); Lactobacillus delbrueckii subsp. bulgaricus (3); L. plantarum (4); L. casei (5); B. infantis (6); L. acidophilus (7); Streptococcus thermophilus (8); B. breve (9); and VSL#3 preparation (10). Adapted from De Angelis et al. [96].
6.3. Use of Detoxified Wheat Flour for Pasta Making
Wheat flour, which was rendered gluten-free by sourdough LAB fermentation and fungal proteases, was used for manufacturing experimental gluten-free pasta (E-GFp), according to a traditional process with a low-temperature drying cycle [98]. Chemical, technology, structural, nutritional, and sensory features were characterized and compared with those of commercial gluten-free (C-GFp) and durum wheat pasta (C-DWp). E-GFp showed rapid water uptake and shorter optimal cooking time compared to the other pastas. Despite the absence of the gluten network, the supplementation with pre-gelatinized rice flour allowed structural properties of E-GFp, which were comparable to those of C-GFp [98]. The in vitro protein digestibility of E-GFp had the best results. Probably due to proteolysis during sourdough fermentation, the chemical scores, essential amino acids profile, biological value, and nutritional index of E-GFp were higher than those of C-DWp. The hydrolysis index (HI) of E-GFp was about 30% lower than that found for C-GFp (Figure 2). As shown by sensory analysis, the characteristics of E-GFp were acceptable. This novel pasta has rather good structural and sensory properties, enhanced digestibility, low HI, and high nutritional quality [98]. The manufacture of E-GFp should be promising to expand the choice of gluten-free foods, which combine sensory and nutritional properties [98].
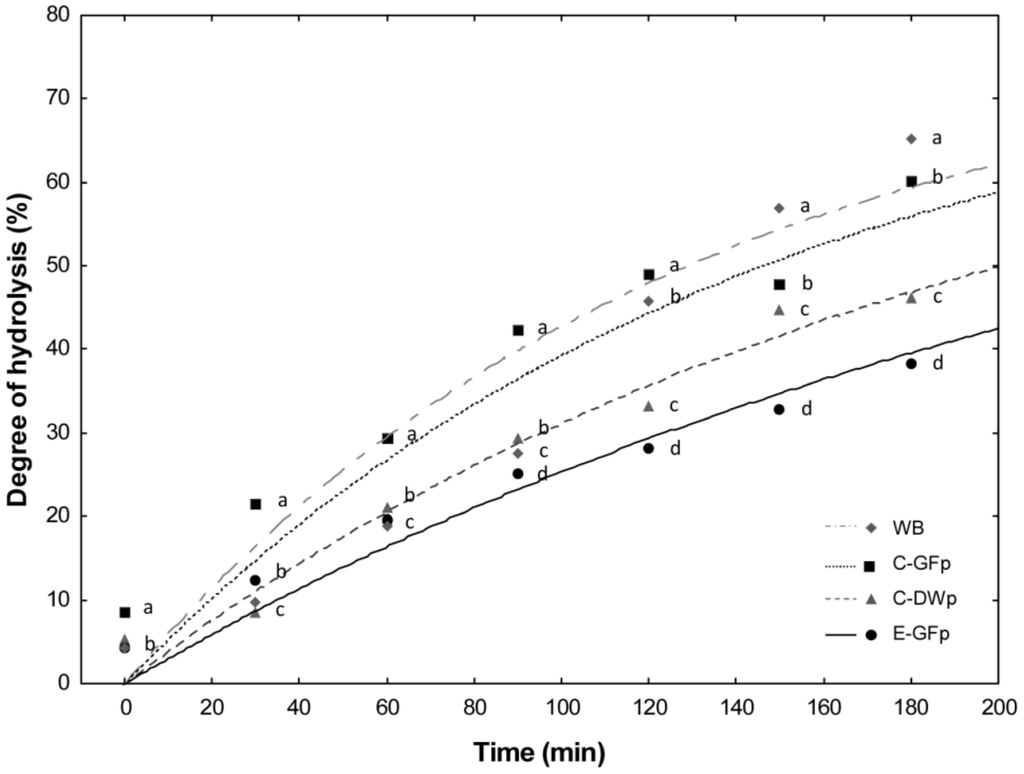
Figure 2.
Rate of starch hydrolysis of pasta following chewing, incubation with pepsin, and further incubation with pancreatic α-amilase in dialysis tubing. E-GFp: experimental gluten-free pasta; C-GFp: commercial gluten-free pasta; C-DWp: commercial durum wheat pasta; WB: white wheat bread (reference). a–d Values obtained at the same time with different superscript letters differ significantly (p < 0.05). Adapted from Curiel et al. [93].
7. Conclusions
Compliance with a gluten-free diet is an extremely challenging task, given a number of problems related to cross-contamination, lack of clear food labelling policies, and poor quality of gluten-free products compared to their gluten-rich counterpart. Even if the exploitation of sourdough in gluten-free systems is still in its infancy, available data indicate that sourdough may be considered a technological tool for improving the texture and flavour of gluten-free products. Indeed, it was demonstrated that the application of sourdough biotechnology to naturally GF ingredients, including GF cereals, pseudocereals, and legumes, may improve the sensory, technological, nutritional, and functional features of final products, similarly to the positive effects described for the gluten-containing matrices.
Besides the application to naturally GF flours, the setup of alternative biotechnologies based on sourdough fermentation is an active area of research that may provide novel possibilities to GF product development in the near future [61,94]. Indeed, the use of sourdough LAB was first proposed with the aim of eliminating traces of gluten epitopes in processed foods and will minimize the long-term risks of a multitude of individuals affected by CD worldwide. Recently, a novel biotechnology process to completely hydrolyse gluten in wheat flour [87] was optimized, patented [99], and industrialized, and GF baked goods made with rendered GF wheat flour are now available on the global market, providing new options for the food industry and consumers [88,90].
Together with recent medical and technological advances, sourdough-based biotechnology could contribute to improve the quality of life of coeliac patients in the near future.
Author Contributions
Luana Nionelli, bibliographic research and data evaluation; Carlo G. Rizzello, article conception and writing.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Röcken, W.; Voysey, P.A. Sourdough fermentation in bread making. J. Appl. Microbiol. 1995, 79, 38S–48S. [Google Scholar]
- Hammes, W.P.; Gänzle, M.G. Sourdough breads and related products. In Microbiology of Fermented Foods; Woods, B.J.B., Ed.; Blackie Academic/Professional: London, UK, 1998; Volume 1, pp. 199–216. [Google Scholar]
- Ehrmann, M.A.; Vogel, R.F. Molecular taxonomy and genetics of sourdough lactic acid bacteria. Trends Food Sci. Tech. 2005, 16, 31–42. [Google Scholar] [CrossRef]
- De Vuyst, L.; Neysens, P. The sourdough microflora: Biodiversity and metabolic interactions. Trends Food Sci. Tech. 2005, 16, 43–56. [Google Scholar] [CrossRef]
- Gobbetti, M.; De Angelis, M.; Corsetti, A.; Di Cagno, R. Biochemistry and physiology of sourdough lactic acid bacteria. Trends Food Sci. Tech. 2005, 16, 57–69. [Google Scholar] [CrossRef]
- De Vuyst, L.; Schrijvers, V.; Paramithiotis, B.; Hoste, M.; Vancanneyt, M.; Swings, J.; Kalantzopoulos, G.; Tsakalidou, E.; Messens, W. The biodiversity of lactic acid bacteria in Greek traditional wheat sourdoughs is reflected in both composition and metabolite formation. Appl. Environ. Microbiol. 2002, 68, 6059–6069. [Google Scholar] [CrossRef] [PubMed]
- Gobbetti, M. The sourdough microflora: Interactions of lactic acid bacteria and yeasts. Trends Food Sci. Tech. 1998, 9, 267–274. [Google Scholar] [CrossRef]
- De Vuyst, L.; Vancanneyt, M. Biodiversity and identification of sourdough lactic acid bacteria. Food Microbiol. 2007, 24, 120–127. [Google Scholar] [CrossRef] [PubMed]
- Aslam, Z.; Im, W.T.; Ten, L.N.; Lee, M.J.; Kim, K.H.; Lee, S.T. Lactobacillus siliginis sp. nov., isolated from wheat sourdough in South Korea. Int. J. Syst. Evol. Microbiol. 2006, 56, 2209–2213. [Google Scholar] [CrossRef] [PubMed]
- Scheirlink, I.; Van der Meulen, R.; Van Schoor, A.; Cleenwerck, I.; Huys, G.; Vandamme, P.; De Vuyst, L.; Vancanneyt, M. Lactobacillus namurensis sp. nov., isolated from a traditional belgian sourdough. Int. J. Syst. Evol. Microbiol. 2007, 57, 223–227. [Google Scholar] [CrossRef] [PubMed]
- Valcheva, R.; Korakli, M.; Onno, B.; Prévost, H.; Ivanova, H.; Ehrmann, I.; Dousset, M.A.; Gänzle, M.G.; Vogel, R.F. Lactobacillus hammesii sp. nov., isolated from French sourdough. Int. J. Syst. Evol. Microbiol. 2005, 55, 763–767. [Google Scholar] [CrossRef] [PubMed]
- Valcheva, R.; Ferchichi, M.; Korakli, M.; Ivanova, I.; Gänzle, M.G.; Vogel, R.F.; Prévost, F.; Onno, B.; Dousset, X. Lactobacillus nantentis sp. nov. isolated from French wheat sourdough. Int. J. Syst. Evol. Microbiol. 2006, 56, 587–591. [Google Scholar] [PubMed]
- Vancanneyt, M.; Neysens, P.; Dewachter, M.; Engelbeen, K.; Snauwaert, C.; Cleenwerck, I.; van der Meulen, R.; Hoste, B.; Tsakalidou, E.; De Vuyst, L.; et al. Lactobacillus acidifarinae sp. nov., from wheat sourdoughs. Int. J. Syst. Evol. Microbiol. 2005, 55, 615–620. [Google Scholar] [CrossRef] [PubMed]
- Gobbetti, M.; Corsetti, A. Lactobacillus sanfrancisco a key sourdough lactic acid bacterium: A review. Food Microbiol. 1997, 14, 175–187. [Google Scholar] [CrossRef]
- Chavan, R.S.; Chavan, S.R. Sourdough technology - A traditional way for wholesome foods: A review. Compr. Rev. Food Sci. Food Saf. 2011, 10, 169–182. [Google Scholar] [CrossRef]
- Corsetti, A.; Gobbetti, M.; De Marco, B.; Balestrieri, F.; Paletti, F.; Rossi, J. Combined effect of sourdough lactic acid bacteria and additives on bread firmness and staling. J. Agric. Food Chem. 2000, 48, 3044–3051. [Google Scholar] [CrossRef] [PubMed]
- Crowley, P.; Schober, T.; Clarke, C.; Arendt, E.K. The effect of storage time on textural and crumb grain characteristics of sourdough wheat bread. Eur. Food Res. Technol. 2002, 214, 489–496. [Google Scholar] [CrossRef]
- Clarke, C.I.; Schober, T.J.; Arendt, E.K. The effect of single strain and traditional mixed strain starter cultures on rheological properties of wheat dough and bread quality. Cereal Chem. 2002, 79, 640–647. [Google Scholar] [CrossRef]
- Di Cagno, R.; De Angelis, M.; Lavermicocca, P.; De Vincenzi, M.; Giovannini, C.; Faccia, M.; Gobbetti, M. Proteolysis by sourdough lactic acid bacteria: Effects on wheat flour protein fractions and gliadin peptides involved in human cereal intolerance. Appl. Environ. Microbiol. 2002, 68, 623–633. [Google Scholar] [CrossRef] [PubMed]
- Korakli, M.; Rossmann, A.; Gänzle, G.; Vogel, R.F. Sucrose metabolism and exopolysaccharide production in wheat and rye sourdoughs by Lactobacillus sanfranciscensis. J. Agric. Food Chem. 2001, 49, 5194–5200. [Google Scholar] [CrossRef] [PubMed]
- Hoseney, C. Principles of Cereals Science and Technology, 2nd ed.; American Association of Cereal Chemists: St. Paul, MN, USA, 1994. [Google Scholar]
- Gänzle, M.G.; Vermeulen, N.; Vogel, R.F. Carbohydrate, peptide and lipid metabolism of lactic acid bacteria in sourdough. Food Microbiol. 2007, 24, 128–138. [Google Scholar] [CrossRef] [PubMed]
- Vermeulen, N.; Kretzer, J.; Machalitza, H.; Vogel, R.F.; Gänzle, M.G. Influence of redox-reactions catalysed by homo-and hetero-fermentative lactobacilli on gluten in wheat sourdoughs. J. Cereal Sci. 2006, 43, 137–143. [Google Scholar] [CrossRef]
- Gobbetti, M.; Lavermicocca, P.; Minervini, F.; De Angelis, M.; Corsetti, A. Arabinose fermentation by Lactobacillus plantarum in sourdough added of pentosans and α-L-arabinofuranosidase: A tool to increase the production of acetic acid. J. Appl. Microbiol. 2000, 88, 317–324. [Google Scholar] [CrossRef] [PubMed]
- Gobbetti, M.; Corsetti, A. Co-metabolism of citrate and maltose by Lactobacillus brevis subsp. lindneri CB1 citrate-negative strains: Effect on growth, end-products and sourdough fermentation. Z. Lebensm. Unters. Forsch. 1996, 203, 82–87. [Google Scholar]
- Di Cagno, R.; De Angelis, M.; Corsetti, A.; Lavermicocca, P.; Arnoult, P.; Tossut, P.; Gallo, G.; Gobbetti, M. Interaction between sourdough lactic acid bacteria and exogenous enzymes: Effects on the microbial kinetics of acidification and dough textural properties. Food Microbiol. 2003, 20, 67–75. [Google Scholar] [CrossRef]
- Thiele, C.; Gänzle, M.G.; Vogel, R.F. Contribution of sourdough lactobacilli, yeast and cereal enzymes to the generation of amino acids in dough relevant for bread flavour. Cereal Chem. 2002, 79, 45–51. [Google Scholar] [CrossRef]
- Kieronczyk, A.; Skeie, S.; Olsen, K.; Langsrud, T. Metabolism of amino acids by resting cells of non-starter lactobacilli in relation to flavour development in cheese. Int. Dairy J. 2001, 11, 217–224. [Google Scholar] [CrossRef]
- De Angelis, M.; Mariotti, L.; Rossi, J.; Servili, M.; Fox, P.F.; Rollàn, G.; Gobbetti, M. Arginine catabolism by sourdough lactic acid bacteria: Purification and characterization of the arginine deiminase pathway enzymes from Lactobacillus sanfranciscensis CB1. Appl. Environ. Microbiol. 2002, 68, 6193–6201. [Google Scholar] [CrossRef] [PubMed]
- Schieberle, P. Intense aroma compounds-useful tools to monitor the influence of processing and storage on bread aroma. Adv. Food Sci. 1996, 18, 237–244. [Google Scholar]
- Damiani, P.; Gobbetti, M.; Cossignani, L.; Corsetti, A.; Simonetti, M.S.; Rossi, J. The sourdough microflora. Characterization of hetero- and homofermentative lactic acid bacteria, yeasts and their interactions on the basis of the volatile compounds produced. Lebensm. Wiss. Technol. 1996, 29, 63–70. [Google Scholar] [CrossRef]
- Katina, K.; Arendt, E.K.; Liukkonen, K.H.; Autio, K.; Flander, L.; Poutanen, K. Potential of sourdough for cereal products. Trends Food Sci. Technol. 2005, 16, 104–112. [Google Scholar] [CrossRef]
- Liukkonen, K.H.; Katina, K.; Wilhelmson, A.; Myllymäki, O.; Lampi, A.M.; Kariluoto, S.; Piironen, V.; Heinonen, S.M.; Nurmi, T.; Adlercreutz, H.; et al. Process-induced changes on bioactive compounds in whole grain rye. Proc. Nutr. Soc. 2003, 62, 117–122. [Google Scholar] [CrossRef] [PubMed]
- De Angelis, M.; Gallo, G.; Corbo, M.R.; McSweeney, P.L.H.; Faccia, M.; Giovine, M.; Gobbetti, M. Phytase activity in sourdough lactic acid bacteria: Purification and characterization of a phytase from Lactobacillus sanfranciscensis CB1. Int. J. Food Microbiol. 2003, 87, 259–270. [Google Scholar] [CrossRef]
- Lopez, H.; Krspine, V.; Guy, C.; Messager, A.; Demigne, C.; Remesy, C. Prolonged fermentation of whole wheat magnesium. J. Agric. Food Chem. 2001, 49, 2657–2662. [Google Scholar] [CrossRef] [PubMed]
- Östman, E.; Nilsson, M.; Liljeberg-Elmståhl, H.; Molin, G.; Björck, I. On the effect of lactic acid on blood glucose and insulin responses to cereal products: Mechanistic studies in healthy subjects and in vitro. J. Cereal Sci. 2002, 36, 339–346. [Google Scholar] [CrossRef]
- De Angelis, M.; Rizzello, C.G.; Alfonsi, G.; Arnault, P.; Cappelle, S.; Tossut, P.; Di Cagno, R.; Gobbetti, M. Use of sourdough lactobacilli and oat fibre to decrease the glycemic index of white wheat bread. Br. J. Nutr. 2007, 98, 1196–1205. [Google Scholar] [CrossRef] [PubMed]
- Kirschner, L.; Von Holy, A. Rope spoilage of bread. S. Afr. J. Sci. 1989, 85, 425–427. [Google Scholar]
- Corsetti, A.; Settanni, L.; Van Sinderen, D. Characterization of bacteriocin-like inhibitory substances (BLIS) from sourdough lactic acid bacteria and evaluation of their in vitro and in situ activity. J. Appl. Microbiol. 2004, 96, 521–534. [Google Scholar] [CrossRef] [PubMed]
- Höltzel, A.; Gänzle, M.G.; Nicholson, G.J.; Hammes, W.P.; Jung, G. The first low-molecular-weight antibiotic from lactic acid bacteria: Reutericyclin, a new tetramic acid. Angew. Chem. Int. Ed. 2000, 39, 2766–2768. [Google Scholar] [CrossRef]
- Schnürer, J.; Magnusson, J. Antifungal lactic acid bacteria as bio-preservatives. Trends Food Sci. Technol. 2005, 16, 70–78. [Google Scholar] [CrossRef]
- Dal Bello, F.; Clarke, C.I.; Ryan, L.A.; Ulmer, H.; Schober, T.J.; Stroem, K.; Sjoergen, J.; van Sinderen, D.; Schnuerer, J.; Arendt, E.K. Improvement of the quality and shelf life of wheat bread by fermentation with the antifungal strain Lactobacillus plantarum FST 1.7. J. Cereal Sci. 2007, 45, 309–318. [Google Scholar] [CrossRef]
- Corsetti, A.; Gobbetti, M.; Rossi, J.; Damiani, P. Antimould activity of sourdough lactic acid bacteria: Identification of a mixture of organic acids produced by Lactobacillus sanfrancisco CB1. Appl. Microbiol. Biotechnol. 1998, 50, 253–256. [Google Scholar] [CrossRef] [PubMed]
- Lavermicocca, P.; Valerio, F.; Visconti, A. Antifungal activity of phenyllactic acid against moulds isolated from bakery products. Appl. Environ. Microbiol. 2003, 69, 634–640. [Google Scholar] [CrossRef] [PubMed]
- Coda, R.; Rizzello, C.G.; Nigro, F.; De Angelis, M.; Arnault, P.; Gobbetti, M. Long-term fungi inhibitory activity of water-soluble extract from Phaseolus vulgaris cv Pinto and sourdough lactic acid bacteria during bread storage. Appl. Environ. Microbiol. 2008, 74, 7391–7398. [Google Scholar] [CrossRef] [PubMed]
- Coda, R.; Cassone, A.; Rizzello, C.G.; Nionelli, L.; Cardinali, G.; Gobbetti, M. Antifungal activity of Wickerhamomyces anomalu and Lactobacillus plantarum during sourdough fermentation: Identification of novel compounds and long-term effect during storage of wheat bread. Appl. Environ. Microbiol. 2011, 77, 3484–3492. [Google Scholar] [CrossRef] [PubMed]
- Rizzello, C.G.; Cassone, A.; Coda, R.; Gobbetti, M. Antifungal activity of sourdough fermented wheat germ used as an ingredient for bread making. Food Chem. 2011, 127, 952–959. [Google Scholar] [CrossRef] [PubMed]
- Rizzello, C.G.; Nionelli, L.; Coda, R.; Gobbetti, M. Synthesis of the cancer preventive peptide lunasin by lactic acid bacteria during sourdough fermentation. Nutr. Cancer 2012, 64, 111–120. [Google Scholar] [CrossRef] [PubMed]
- Rizzello, C.G.; Hernández-Ledesma, B.; Fernández-Tomé, S.; Curiel, J.A.; Pinto, D.; Marzani, B.; Coda, R.; Gobbetti, M. Italian legumes: Effect of sourdough fermentation on lunasin-like polypeptides. Microb. Cell Fact. 2015, 14, 168. [Google Scholar] [CrossRef] [PubMed]
- Coda, R.; Rizzello, C.G.; Pinto, D.; Gobbetti, M. Selected lactic acid bacteria synthesize antioxidant peptides during sourdough fermentation of cereal flours. Appl. Environ. Microbiol. 2012, 78, 1087–1096. [Google Scholar] [CrossRef] [PubMed]
- Rizzello, C.G.; Cassone, A.; Di Cagno, R.; Gobbetti, M. Synthesis of angiotensin I-converting enzyme (ACE)-inhibitory peptides and gamma-aminobutyric acid (GABA) during sourdough fermentation by selected lactic acid bacteria. J. Agric. Food Chem. 2008, 56, 6936–6943. [Google Scholar] [CrossRef] [PubMed]
- Maki, M.; Mustalahti, K.; Korhonen, J.; Kulmala, P.; Haapalahti, M.; Karttunen, T. Prevalence of celiac disease among children in Finland. N. Engl. J. Med. 2003, 348, 2517–2524. [Google Scholar] [CrossRef] [PubMed]
- Di Sabatino, A.; Corazza, G.R. Coeliac disease. Lancet 2009, 373, 1480–1493. [Google Scholar] [CrossRef]
- Adams, F. On the Coeliac Affection. The Extant Works of Aretaeus, The Cappadocian; Sydenham Society: London, UK, 1856; pp. 350–351. [Google Scholar]
- Gee, S. On the Celiac Desease; Saint Bartholomew’s Hospital: London, UK, 1888; pp. 17–20. [Google Scholar]
- Van Berge-Henegouwen, G.P.; Mulder, C.J.J. Pioneer in the gluten free diet: Willem- Karel Dicke 1905–1962, over 50 years of gluten free diet. Gut 1993, 34, 1473–1475. [Google Scholar] [CrossRef] [PubMed]
- Fasano, A.; Catassi, C. Current approaches to diagnosis and treatment of celiac desease. An evolving sprectrum. Gastroenterology 2001, 120, 636–651. [Google Scholar] [CrossRef] [PubMed]
- Battais, F.; Richard, C.; Jacquenet, S.; Denery-Papini, S.; Moneret-Vautrin, D.A. Wheat grain allergies: An update on wheat allergens. Eur. Ann. Allergy Clin. Immunol. 2008, 40, 67–76. [Google Scholar] [PubMed]
- Verdu, E.F.; Armstrong, D.; Murray, J.A. Between celiac desease and irritable bowel syndrome: The “no man’s land” of gluten sensitivity. Am. J. Gastroenterol. 2009, 104, 1587–1594. [Google Scholar] [CrossRef] [PubMed]
- Fasano, A. Prevalence and genetics. In AGA Clinical Symposium- Celiac Disease Clinical Symposium, Program and Abstract of Digestive Disease Week; Lousiana: New Orleans, LA, USA, 2004. [Google Scholar]
- Deora, N.S.; Deswal, A.; Mishra, H.N. Alternative approaches towards gluten-free dough development: Recent trends. Food Eng. Rev. 2014, 6, 89–104. [Google Scholar] [CrossRef]
- Gallagher, E.; Gormley, T.R.; Arendt, E.K. Recent advances in the formulation of gluten-free cereal-based products. Trends Food Sci. Technol. 2004, 15, 143–152. [Google Scholar] [CrossRef]
- Moore, M.M. Novel Approaches in the Structural Development of Gluten Free Bread. Doctoral Dissertation, University College, Cork, Ireland, 2005. [Google Scholar]
- Diowksz, A.; Kozioł, G.; Kordialik-Bogacka, E.; Ambroziak, W.; Sucharzewska, D. β-glucan Content in Gluten Free Sourdough Breads Supplemented with Soya Sprouts. In Proceedings of the 3th International Symposium on Sourdough, Bari, Italy, 25–28 October 2006.
- Ryan, L.A.; Dal Bello, F.; Renzetti, S.; Arendt, E.K. The use of selected lactic acid bacteria to improve the baking and rheological quality of gluten-free batter and bread. In Proceedings of the World Grains Summit: Food and Beverages, San Francisco, CA, USA, 17–21 September 2006.
- Giuliani, G.M.; Benedusi, A.; Di Cagno, R.; De Angelis, M.; Luisi, A.; Gobbetti, M. Mixture of Lactic Bacteria for the Preparation of Gluten Free Baked Products. Patent U.S. 9237753 B2, 19 January 2016. [Google Scholar]
- Picozzi, C.; Mariotti, M.; Cappa, C.; Tedesco, B.; Vigentini, I.; Foschino, R.; Lucisano, M. Development of Type I gluten-free sourdough. Lett. Appl. Microbiol. 2015, 62, 119–125. [Google Scholar] [CrossRef] [PubMed]
- Campo, E.; del Arco, L.; Urtasun, L.; Oria, R.; Ferre-Mairal, A. Impact of sourdough on sensory properties and consumer’ preference of gluten-.free breads eriche with teff flour. J. Cereal Sci. 2016, 67, 75–82. [Google Scholar] [CrossRef]
- Różylo, R.; Rudy, S.; Krzykowski, A.; Dziki, D.; Gawlik-Dziki, U.; Różylo, k.; Skonecki, S. Effect of adding fresh and freese-dried buckwheat sourdough on gluten-free bread quality. Int. J. Food Sci. Technol. 2015, 50, 313–322. [Google Scholar] [CrossRef]
- Aguilar, N.; Albanell, E.; Miñarro, B.; Capellas, M. Chestnut flour sourdough for glutn-free bread making. Eur. Food Res. Technol. 2016, 242, 1–8. [Google Scholar] [CrossRef]
- Axel, C.; Röcker, B.; Brosnon, B.; Zannini, E.; Furey, A.; Coffey, A.; Arendt, E.K. Application of Lactobacillus amylovorus DSM19280 in gluten-free sourdough bread to improve the microbial shelf life. Food Microbiol. 2015, 47, 36–44. [Google Scholar] [CrossRef] [PubMed]
- Wolter, A.; Hager, A.-S.; Zannini, E.; Czeny, M.; Arendt, E.K. Influence of dextran- producing Weissella cibaria on baking properties and sensory profile of gluten-free and wheat breads. Int. J. Food Microbiol. 2014, 172, 83–91. [Google Scholar] [CrossRef] [PubMed]
- Wolter, A.; Hager, A.-S.; Zannini, E.; Arendt, E.K. Influence of sourdough on in vitro starch digestibility and predicted glycemin indices of gluten-free bread. Food Funct. 2014, 5, 564–572. [Google Scholar] [CrossRef] [PubMed]
- Moroni, A.V.; Dal Bello, F.; Arendt, E.K. Sourdough in gluten-free bread-making: An ancient technology to solve a novel issue? Food Microbiol. 2009, 26, 676–684. [Google Scholar] [CrossRef] [PubMed]
- M’hir, S.; Ziadi, M.; Chammem, N.; Hamdi, M. Gluten proteolysis as alternative therapy for celiac patients: A mini-review. Afr. J. Biotechnol. 2012, 11, 7323–7330. [Google Scholar]
- Pyle, G.G.; Paaso, B.; Anderson, B.E.; Allen, D.A.; Marti, T.; Li, Q.; Siegel, M.; Khosla, C.; Gray, G.M. Effect of pretreatment of food gluten with prolyl endopeptidase on gluten-induced malabsorption in celiac sprue. Clin. Gastroenterol. Hepatol. 2005, 3, 687–694. [Google Scholar] [CrossRef]
- Shan, L.; Marti, T.; Sollid, L.M.; Gray, G.M.; Khosla, C. Comparative biochemical analysis of three bacterial prolyl endopeptidases: Implications for celiac sprue. Biochem J. 2004, 383, 311–318. [Google Scholar] [CrossRef] [PubMed]
- Stepniak, D.; Spaenij-Dekking, L.; Mitea, C.; Moester, M.; de Ru, A.; Baak-Pablo, R.; van Veelen, P.; Edens, L.; Koning, F. Highly efficient gluten degradation with a newly identified prolyl endoprotease: implications for celiac disease. Am. J. Physiol. Gastrointest. Liver Physiol. 2006, 291, G621–G629. [Google Scholar] [CrossRef] [PubMed]
- Walter, T.; Wieser, H.; Koehler, P. Degradation of gluten in rye sourdough products by means of a proline-specific peptidase. Eur. Food Res. Technol. 2015, 240, 517–524. [Google Scholar] [CrossRef]
- Di Cagno, R.; De Angelis, M.; Auricchio, S.; Greco, L.; Clarke, C.; De Vincenzi, M.; Giovannini, C.; D’Archivio, M.; Landolfo, F.; Parrilli, G.; et al. A sourdough bread made from wheat and non-toxic flours and started with selected lactobacilli is tolerated in celiac sprue. Appl. Environ. Microbiol. 2004, 70, 1088–1096. [Google Scholar] [CrossRef] [PubMed]
- De Angelis, M.; Rizzello, C.G.; Scala, E.; De Simone, C.; Farris, G.A.; Turrini, F.; Gobbetti, M. VSL#3 probiotic preparation has the capacity to hydrolyze gliadin polypeptides responsible for celiac sprue. Biochim. Biophys. Acta 2006, 1762, 80–93. [Google Scholar] [PubMed]
- Di Cagno, R.; De Angelis, M.; Alfonsi, G.; De Vincenzi, M.; Silano, M.; Vincentini, O.; Gobbetti, M. Pasta made from durum wheat semolina fermented with selected lactobacilli as a tool for a potential decrease of the gluten intolerance. J. Agric. Food Chem. 2005, 53, 4379–4402. [Google Scholar] [CrossRef] [PubMed]
- De Angelis, M.; Coda, R.; Silano, M.; Minervini, F.; Rizzello, C.G.; Di Cagno, R.; Vicentini, O.; De Vincenzi, M.; Gobbetti, M. Fermentation by selected sourdough lactic acid bacteria to decrease the intolerance to rye and barley flours. J. Cereal Sci. 2006, 43, 301–314. [Google Scholar] [CrossRef]
- Gerez, C.L.; Dallagnol, A.; Rollán, G.; Font de Valdez, G. A combination of two lactic acid bacteria improves the hydrolysis of gliadin during wheat dough fermentation. Food Microbiol. 2012, 32, 427–430. [Google Scholar] [CrossRef] [PubMed]
- Engström, N.; Sandberg, A.-S.; Scheers, N. Sourdough fermentation of wheat flour does not prevent the interaction of transglutaminase 2 with α2-gliadin or gluten. Nutrients 2015, 7, 2134–2144. [Google Scholar] [CrossRef] [PubMed]
- De Angelis, M.; Di Cagno, R.; Gallo, G.; Curci, M.; Siragusa, S.; Crecchio, C.; Parente, E.; Gobbetti, M. Molecular and functional characterization of Lactobacillus sanfranciscensis strains isolated from sourdoughs. Int. J. Food Microbiol. 2007, 114, 69–82. [Google Scholar] [CrossRef] [PubMed]
- Rizzello, C.G.; De Angelis, M.; Di Cagno, R.; Camarca, A.; Silano, M.; Losito, I.; De Vincenzi, M.; De Bari, M.D.; Palmisano, F.; Maurano, F.; et al. Highly efficient gluten degradation by lactobacilli and fungal proteases during food processing: New perspectives for celiac disease. Appl. Environ. Microbiol. 2007, 73, 4499–4507. [Google Scholar] [CrossRef] [PubMed]
- Di Cagno, R.; Rizzello, C.G.; Gobbetti, M. Adverse reactions to gluten: Exploitation of sourdough fermentation. In Wheat and Rye in Disease Prevention and Health; Watson, R.R., Preedy, V., Ziboli, S., Eds.; Academic Press: Cambridge, MA, USA, 2014; pp. 171–177. [Google Scholar]
- Cabrera-Chavez, F.; Calderon de la Barca, A.M. Trends in wheat technology and modification of gluten proteins for dietary treatment of coeliac disease patients. J. Cereal Sci. 2010, 52, 337–341. [Google Scholar] [CrossRef]
- Rizzello, C.G.; Montemurro, M.; Gobbetti, M. Characterization of the bread made with durum wheat semolina rendered gluten-free by sourdough biotechnology in comparison with commercial gluten-free products. J. Food Sci. 2016. [Google Scholar] [CrossRef] [PubMed]
- Auricchio, S.; Greco, L.; De Vizia, B.; Buonocore, V. Dipeptidyalaminopeptidase and carboxypeptidases activities of the brush border of rabbit small intestine. Gastroenterology 1978, 75, 1073–1079. [Google Scholar] [PubMed]
- Andria, G.; Cucchiara, S.; De Vizia, B.; De Ritis, G.; Mazzacca, G.; Auricchio, S. Brush border and cytosol peptidase activities of human small intestine in normal subjects and celiac patients. Pediatr. Res. 1980, 14, 812–818. [Google Scholar] [CrossRef] [PubMed]
- Hausch, F.; Shan, L.; Santiago, N.A.; Gray, G.M.; Khosla, C. Intestinal digestive resistance of immunodominant gliadin peptides. Am. J. Physiol. Gastrointest. Liver Physiol. 2002, 283, 996–1003. [Google Scholar] [CrossRef] [PubMed]
- De Angelis, M.; Cassone, A.; Rizzello, C.G.; Gagliardi, F.; Minervini, F.; Calasso, M.; Di Cagno, R.; Francavilla, R.; Gobbetti, M. Mechanism of degradation of immunogenic gluten epitopes from Triticum turgidum L. var. durum by sourdough lactobacilli and fungal proteases. Appl. Environ. Microbiol. 2010, 76, 508–518. [Google Scholar] [PubMed]
- Kunji, E.R.S.; Mierau, I.; Hagting, A.; Poolman, B.; Konings, W.N. The proteolytic systems of lactic acid bacteria. Antonie Van Leeuw. J. Microb. 1996, 70, 187–221. [Google Scholar] [CrossRef]
- Di Cagno, R.; Barbato, M.; Di Camillo, C.; Rizzello, C.G.; De Angelis, M.; Giuliani, G.; De Vincenzi, M.; Gobbetti, M.; Cucchiara, S. Gluten-free sourdough wheat baked goods appear safe for young celiac patients: A pilot study. J. Pediatr. Gastroenterol. Nutr. 2010, 51, 777–783. [Google Scholar] [CrossRef] [PubMed]
- Greco, L.; Gobbetti, M.; Auricchio, R.; Di Mase, R.; Landolfi, F.; Paparo, F.; Di Cagno, R.; De Angelis, M.; Rizzello, C.G.; Cassone, A.; et al. Safety for celiac patients of baked goods made of wheat flour 533 hydrolyzed during food processing. Clin. Gastroenterol. Hepatol. 2011, 9, 24–29. [Google Scholar] [CrossRef] [PubMed]
- Curiel, J.A.; Coda, R.; Limitone, A.; Katina, K.; Raulio, M.; Giuliani, G.; Rizzello, C.G.; Gobbetti, M. Manufacture and characterization of pasta made with wheat flour rendered gluten-free using fungal proteases and selected sourdough lactic acid bacteria. J. Cereal Sci. 2014, 59, 79–87. [Google Scholar] [CrossRef]
- Giuliani, G.; Benedusi, A.; Di Cagno, R.; Rizzello, C.G.; De Angelis, M.; Gobbetti, M.; Cassone, A. Process of Microbic Biotechnology for Completely Degrading Gluten in Flours. W.O. Patent 2010073283-A2, 1 July 2010. [Google Scholar]
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).