


Preliminary Exploration on the Effects of a Novel Antidepressant Formula Food in a Mouse Model

Abstract
1. Introduction
2. Materials and Methods
2.1. Experimental Animal
2.2. Drugs and Reagents
2.3. Experimental Design
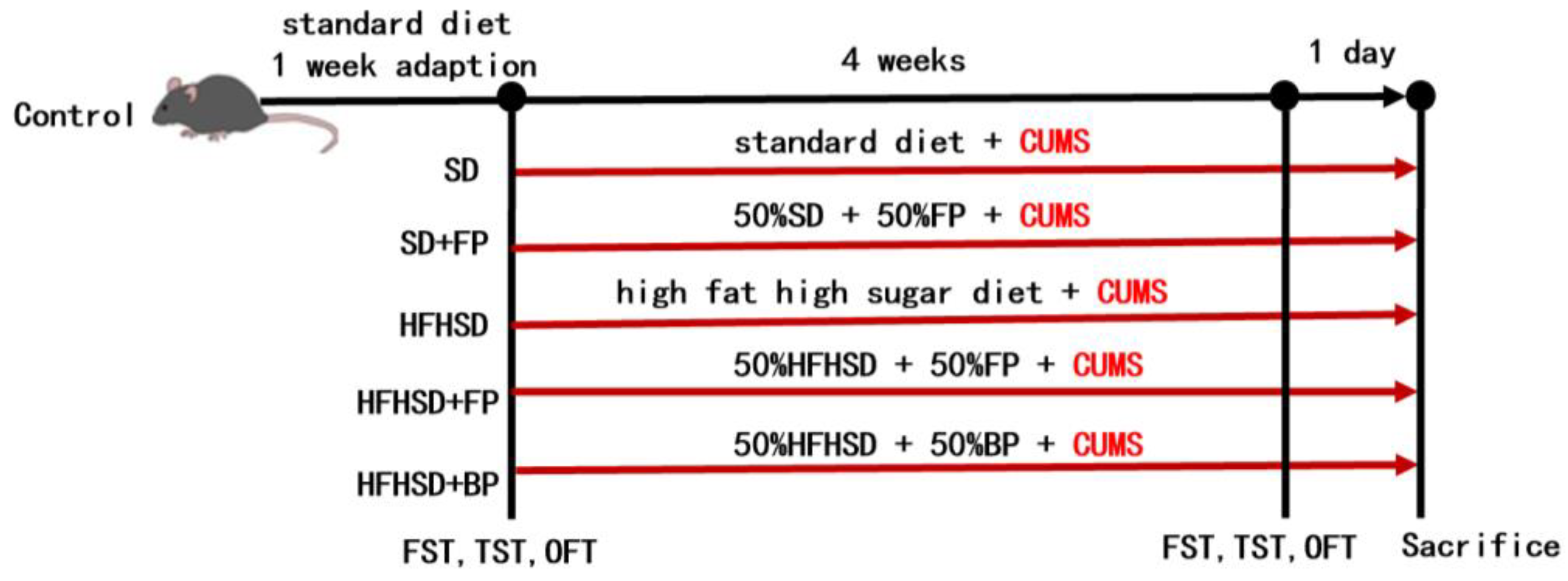
2.3.1. Grouping and Treatment of Mice
2.3.2. Behavioral Experiments
Open Field Test
Tail Suspension Test
Forced Swimming Test
2.3.3. Organ Index
2.3.4. Quantitative Real-Time PCR
2.4. Statistical Analysis
3. Results
3.1. Flow Chart of the Animal Experiment
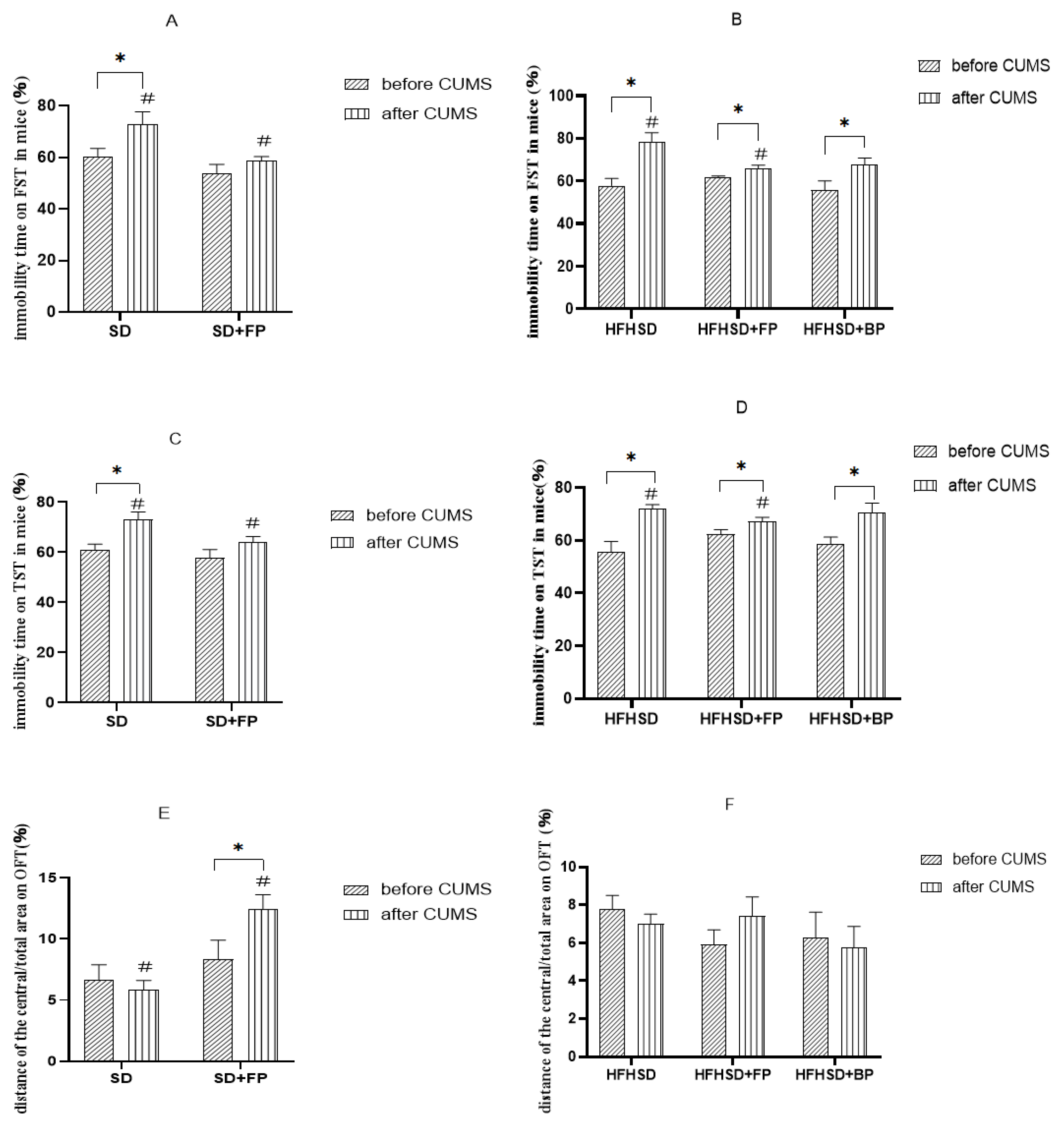
3.2. The Effect of Formula Powder in Treating CUMS Mice
3.3. Effect of Formula Food on Organ Index in CUMS Mice
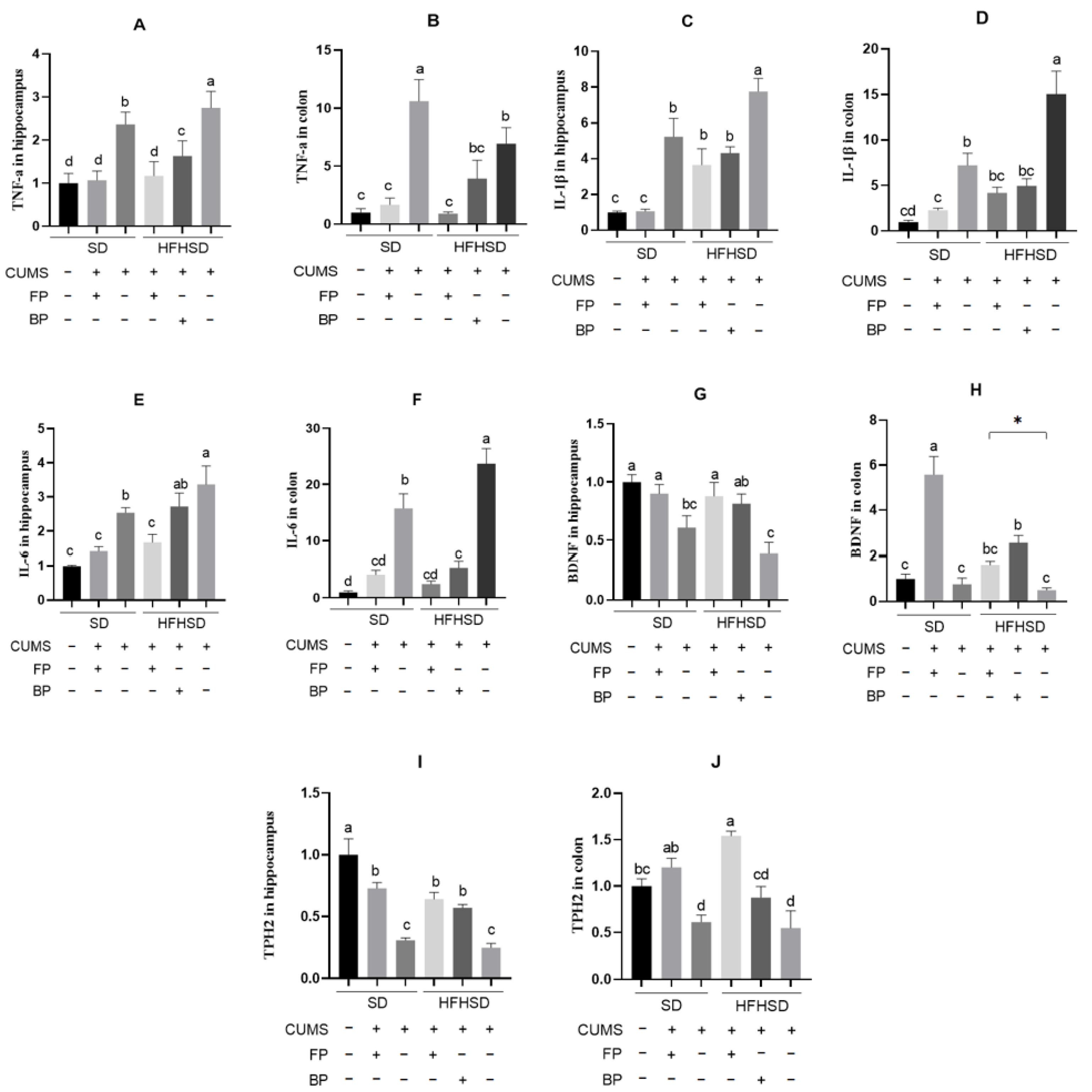
3.4. Effects of the Formula Food on the Expression of Inflammatory Factors in CUMS Mice
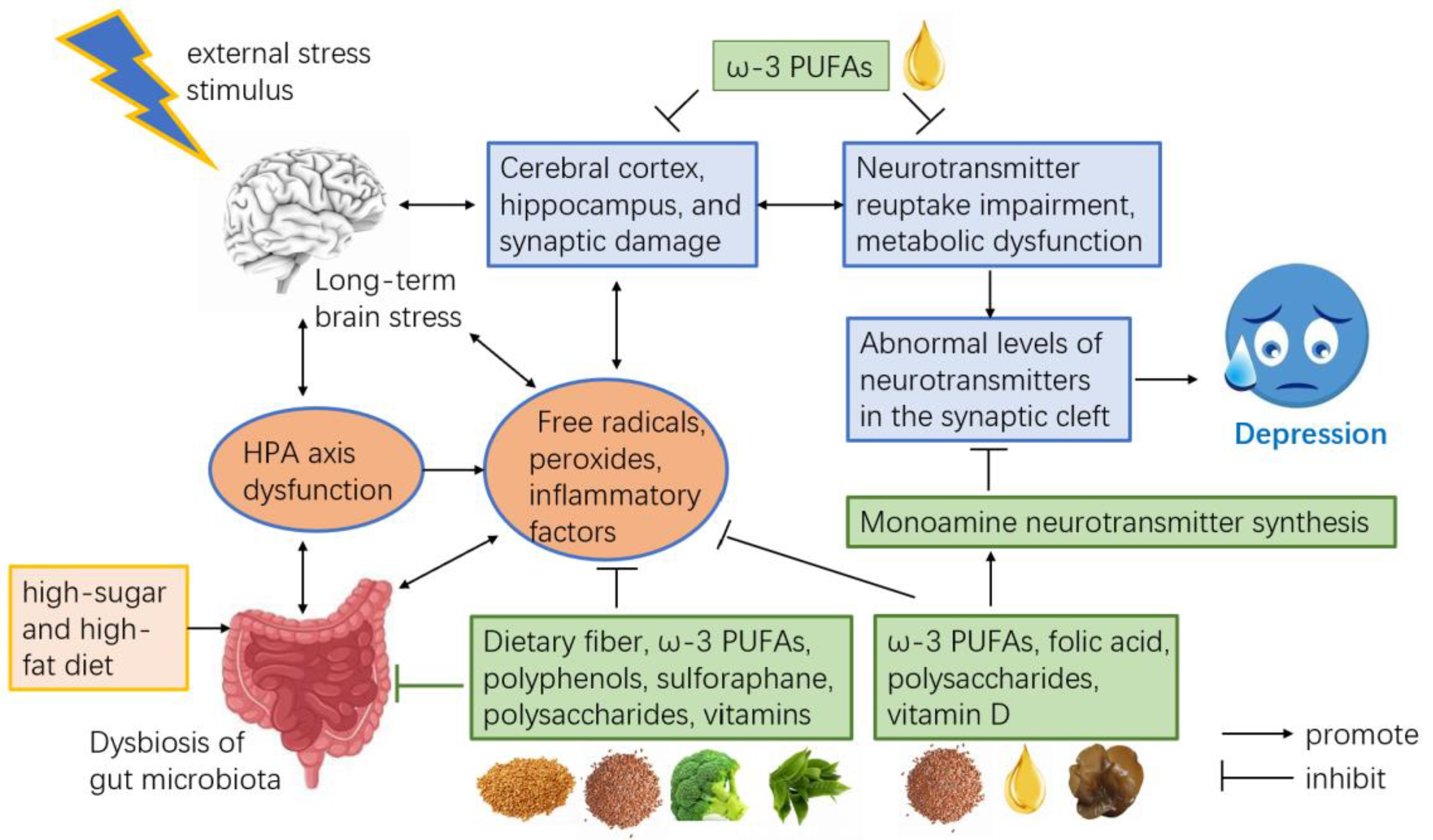
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| SD | Standard diet |
| HFHSD | High-fat high-sugar diet |
| FP | Formula powder |
| BP | Buckwheat powder |
| CUMS | Chronic unpredictable mild stress |
| PCR | Polymerase Chain Reaction |
| FST | Forced swimming test |
| TST | Tail suspension test |
| OFT | Open field test |
| TNF-α | Tumor necrosis factor-α |
| IL-1β | Interleukin-1 beta |
| IL-6 | Interleukin-6 |
| BDNF | Brain-derived neurotrophic factor |
| TPH2 | Tryptophan hydroxylase 2 |
References
- Huang, Y.; Wang, Y.; Wang, H. Prevalence of mental disorders in China: A cross-sectional epidemiological study. Lancet Psychiatry 2019, 6, e11. [Google Scholar] [CrossRef]
- Rong, L.; Zheng, Y.; Duan, X.; Liu, Y.; Zhang, X.; Hu, R.; Zhu, L.; Huang, Y.; Wu, Y.; Mu, F.; et al. Depression and anxiety symptoms and their comorbidity among Chinese residents in 2021 and 2022. Chin. J. Ment. Health 2023, 37, 1023–1030. [Google Scholar]
- Yang, R.; Liu, Z.; Sun, L.; Zhang, J.; Ma, Q.; Yao, G.; Lin, Y. Non-suicidal self-injury detection rate and influencing factors in hospitalized children and adolescents with depressive disorders. Int. J. Psychiatry 2023, 50, 1008–1011. [Google Scholar]
- Hua, Z.; Wang, S.; Yuan, X. Trends in age-standardized incidence rates of depression in adolescents aged 10–24 in 204 countries and regions from 1990 to 2019. J. Affect. Disord. 2024, 350, 831–837. [Google Scholar] [CrossRef] [PubMed]
- Sun, M.; Chen, W. Interpretation of the Chinese Overweight/Obesity Medical Nutrition Therapy Guidelines (2021). Peking. Union. Med. J. 2022, 13, 255–262. [Google Scholar]
- Fitzpatrick, M.; Solberg, W.L. Adenylate cyclase 3: A potential genetic link between obesity and major depressive disorder. Physiol. Genom. 2024, 56, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Frank, P.; Jokela, M.; Batty, G.D.; Lassale, C.; Steptoe, A.; Kivimaki, M. Overweight, obesity, and individual symptoms of depression: A multicohort study with replication in UK Biobank. Brain Behav. Immun. 2022, 105, 192–200. [Google Scholar] [CrossRef]
- Du, Y.; Gao, X.R.; Peng, L.; Ge, J.F. Crosstalk between the microbiota-gut-brain axis and depression. Heliyon 2020, 6, e4097. [Google Scholar] [CrossRef]
- Nanthakumaran, S.; Sridharan, S.; Somagutta, M.R.; Arnold, A.A.; May, V.; Pagad, S.; Malik, B.H. The Gut-Brain Axis and Its Role in Depression. Cureus 2020, 12, e10280. [Google Scholar] [CrossRef]
- Tang, Q. Review on research progress of depression in traditional Chinese and Western medicine. J. Beijing Univ. Chin. Med. 2022, 45, 871–877. [Google Scholar]
- Su, Z.; Ma, T.; Hu, B.; Ma, T.; Zhang, D.; Jing, Y. Research progress on mechanisms of polysaccharides in alleviating depression and anxiety symptoms. Chin. J. Pharmacol. Toxicol. 2023, 37, 498. [Google Scholar]
- Wang, M.; Wang, C.; Zhao, K.; Ma, S.; Sang, J.; Xu, S. Bioactive compounds from Chinese herbal plants for neurological health: Mechanisms, pathways, and functional food applications. Front. Nutr. 2025, 12, 1537363. [Google Scholar]
- Li, L.; Shi, F.; Tan, M.; Lui, F. Research progress on the role of dietary fiber in regulating gut microbiota. Mod. Dr. China 2020, 58, 188–192. [Google Scholar]
- Fatahi, S.; Matin, S.S.; Sohouli, M.H.; Găman, M.A.; Raee, P.; Olang, B.; Kathirgamathamby, V.; Santos, H.; Guimaraes, N.S.; Shidfar, F. Association of dietary fiber and depression symptom: A systematic review and meta-analysis of observational studies. Complement. Ther. Med. 2021, 56, 102621. [Google Scholar] [CrossRef]
- Esnafoglu, E.; Ozturan, D.D. The relationship of severity of depression with homocysteine, folate, vitamin B12, and vitamin D levels in children and adolescents. Child. Adolesc. Ment. Health 2020, 25, 249–255. [Google Scholar] [CrossRef] [PubMed]
- Tan, Y.; Zhou, L.; Huang, J.; Chen, X.; Wu, Y.; Song, X.; Wang, J.; Hu, H.; Yang, Q. Vitamin B12, folate, homocysteine, inflammatory mediators (Interleukin-6, TNF-α, and CRP) levels in adolescents with anxiety or depressive symptoms. Neuropsychiatr. Dis. Treat. 2023, 19, 785–800. [Google Scholar] [CrossRef]
- Somoza-Moncada, M.M.; Turrubiates-Hernández, F.J.; Muñoz-Valle, J.F.; Gutiérrez-Brito, J.A.; Díaz-Pérez, S.A.; Aguayo-Arelis, A.; Hernández-Bello, J. Vitamin D in depression: A potential bioactive agent to reduce suicide and suicide attempt risk. Nutrients 2023, 15, 1765. [Google Scholar] [CrossRef]
- Bagchi, D. Nutraceutical and Functional Food Regulations; Elsevier: New York, NY, USA, 2014. [Google Scholar]
- Nollet, M. Models of Depression: Unpredictable Chronic Mild Stress in Mice. Curr. Protoc. 2021, 1, e208. [Google Scholar] [CrossRef]
- Li, N.; Zhao, X.; Du, C.; Song, Y.; Zhang, S.; Ma, S. Intervention effect of Jiawei Banxia Houpo Decoction on depressed rats. J. Tradit. Chin. Med. 2018, 33, 2379–2382. [Google Scholar]
- Sharma, S.; Chawla, S.; Kumar, P.; Ahmad, R.; Kumar, P. The chronic unpredictable mild stress (CUMS) Paradigm: Bridging the gap in depression research from bench to bedside. Brain Res. 2024, 1843, 149123. [Google Scholar] [CrossRef]
- Wang, Y. Risk Factors for White Matter Damage and Its Interactive Relationship with Cognition and Depression. Ph.D. Thesis, Soochow University Dissertation, Suzhou, China, 2017. [Google Scholar]
- Feng, Y.; Zhang, P.; Cao, M. Study on TNF-α and ADP ratio in patients with depression. Chongqing Med. 2024, 53, 696–699. [Google Scholar]
- Li, S.; Zhang, G.; Zhu, R.; Fan, J.; Yang, S. Changes in serum levels of BDNF and TGF-β1 and their relationship with disease severity and cognitive function in patients with depression. Shandong Med. J. 2024, 64, 77–79. [Google Scholar]
- Zhu, P.; Lu, T.; Wu, J.; Fan, D.; Liu, B.; Zhu, X.; Guo, H.; Du, Y.; Lui, F.; Tian, Y.; et al. Gut microbiota drives macrophage-dependent self-renewal of intestinal stem cells via niche enteric serotonergic neurons. Cell Res. 2022, 32, 555–569. [Google Scholar] [CrossRef]
- Tan, T. Study on the Association Between Abnormal Functional Brain Regions and TPH2 Methylation in Depression. Master’s Thesis, Southeast University Dissertation, Nanjing, China, 2022. [Google Scholar]
- Xu, H.; Chen, X.; Zhao, F.; Mao, R.; Wang, W.; Cheng, Z.; Zhang, Y. Research progress on the effects of dietary structure and nutrients on depression. Food Sci. 2022, 43, 346–355. [Google Scholar]
- Lazarevich, I.; Irigoyen, C.M.; Velazquez-Alva, M.; Zepeda, Z. Relationship among obesity, depression, and emotional eating in young adults. Appetite 2016, 107, 639–644. [Google Scholar] [CrossRef] [PubMed]
- Jitte, S.; Keluth, S.; Bisht, P.; Wal, P.; Singh, S.; Murti, K.; Kumar, N. Obesity and depression: Common link and possible targets. CNS Neurol. Disord. Drug Targets 2024, 23, 1425–1449. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Zhang, J.; Yuan, L.; Hu, H.; Li, T.; Zhao, Y.; Wu, Y.; Wang, M. Obesity and risk of depressive disorder in children and adolescents: A meta-analysis of observational studies. Child. Care Health Dev. 2024, 50, e13237. [Google Scholar] [CrossRef]
- Chen, X.; Zhuo, S.; Zhu, T.; Yao, P.; Yang, M.; Mei, H.; Li, N.; Ma, F.; Wang, J.M.; Chen, S. Fpr2 deficiency alleviates diet-induced insulin resistance by reducing weight gain and inhibiting inflammation mediated by macrophage chemotaxis and M1 polarization. Diabetes 2019, 68, 1130–1142. [Google Scholar] [CrossRef]
- Swann, O.G.; Kilpatrick, M.; Breslin, M.; Oddy, W.H. Dietary fiber and its associations with depression and inflammation. Nutr. Rev. 2020, 78, 394–411. [Google Scholar] [CrossRef]
- Deacon, G.; Kettle, C.; Hayes, D.; Dennis, C.; Tucci, J. Omega 3 polyunsaturated fatty acids and the treatment of depression. Crit. Rev. Food Sci. Nutr. 2017, 57, 212–223. [Google Scholar] [CrossRef]
- Liu, Z.; Zou, Y.; He, M.; Yang, P.; Qu, X. Effect of green tea polyphenols on depressive behavior in social defeat mouse models. Chongqing Med. 2021, 50, 741–745. [Google Scholar]
- Unno, K.; Furushima, D.; Tanaka, Y.; Tominaga, T.; Nakamura, H.; Yamada, H.; Taguchi, K.; Goda, T.; Nakamura, Y. Improvement of depressed mood with green tea intake. Nutrients 2022, 14, 2949. [Google Scholar] [CrossRef] [PubMed]
- Liu, M.; Zhang, Y.; Zhang, Y.; Wang, G.; Zhao, B. Effects and mechanism of Gastrodia polysaccharides on LPS-induced depressive mice. China Pharm. 2021, 24, 2018–2023. [Google Scholar]
- Yang, X. Preliminary Study on Prefrontal microRNA-21 Regulating Anxiety-Like Behavior Via the CRF Signaling Pathway. Master’s Thesis, Zunyi Medical University Dissertation, Zunyi, China, 2022. [Google Scholar]
- Zheng, Y.; Hou, Y.; Li, F.; Shi, X.; Tao, Y.; Zhao, X.; Hu, H.; Wang, L. Near-infrared light exposure significantly improves intestinal dysfunction caused by depression in rats. J. South. Med. Univ. 2023, 43, 1591–1598. [Google Scholar]
- Willner, P. Chronic mild stress (CMS) revisited: Consistency and behavioral-neurobiological concordance in the effects of CMS. Neuropsychobiology 2005, 52, 90–110. [Google Scholar] [CrossRef]
- Albouery, M.; Buteau, B.; Gregoire, S.; Cherbuy, C.; Pais, D.-B.; Martine, L.; Chain, F.; Cabaret, S.; Berdeaux, O.; Bron, A.M.; et al. Age-related changes in the gut microbiota modify brain lipid composition. Front. Cell Infect. Microbiol. 2019, 9, 444. [Google Scholar] [CrossRef] [PubMed]
- Feng, Z.; Fang, C.; Ma, Y.; Chang, J. Obesity-induced blood-brain barrier dysfunction: Phenotypes and mechanisms. J. Neuroinflammation 2024, 21, 110. [Google Scholar] [CrossRef]
- Gomez-Apo, E.; Mondragon-Maya, A.; Ferrari-Diaz, M.; Silva-Pereyra, J. Structural brain changes associated with overweight and obesity. J. Obes. 2021, 2021, 6613385. [Google Scholar] [CrossRef]
- Bruce-Keller, A.J.; Keller, J.N.; Morrison, C.D. Obesity and vulnerability of the CNS. Biochim. Biophys. Acta 2009, 1792, 395–400. [Google Scholar] [CrossRef]
- Henn, R.E.; Elzinga, S.E.; Glass, E.; Parent, R.; Guo, K.; Allouch, A.M.; Mendelson, F.E.; Hayes, J.; Webber-Davis, I.; Murphy, G.G.; et al. Obesity-induced neuroinflammation and cognitive impairment in young adult versus middle-aged mice. Immun. Ageing 2022, 19, 1. [Google Scholar] [CrossRef]
- Shao, J.; Tian, M.; Wang, F.; Sun, Y.; Wang, G.; Ai, L.; Xia, Y. Effects of high sugar and high fat diet on intestinal barrier function and inflammation in mice. J. Chin. Inst. Food Sci. Technol. 2023, 23, 79–88. [Google Scholar]
- Li, K.P.; Yuan, M.; Wu, Y.L.; Pineda, M.; Zhang, C.M.; Chen, Y.F.; Chen, Z.Q.; Rong, X.L.; Turnbull, J.E.; Guo, J. A high-fat high-fructose diet dysregulates the homeostatic crosstalk between gut microbiome, metabolome, and immunity in an experimental model of obesity. Mol. Nutr. Food Res. 2022, 66, e2100950. [Google Scholar] [CrossRef] [PubMed]
- Dalamaga, M.; Tsigalou, C. Diet patterns, gut microbiota and metabolic disorders: Perspectives and challenges. Elsevier 2024, 23, 100310. [Google Scholar] [CrossRef]
- Yao, H.; Yang, J.; Li, S.; Cui, S.; Tan, H.; Nie, S. Effects of different fractions of polysaccharides from Dictyophora indusiata on high-fat diet-induced metabolic syndrome in mice. Int. J. Biol. Macromol. 2024, 272, 132744. [Google Scholar] [CrossRef]
- Zheng, X.; Zhu, Y.; Yuan, Y.; Han, J. Bidirectional relationship between ulcerative colitis and depression and anxiety: Based on ‘disease-induced depression, depression-induced disease’. Shizhen Tradit. Chin. Med. 2022, 33, 1421–1423. [Google Scholar]
- Liu, Z. Study on the Effect and Mechanism of 5-ASA and High-Fat Diet in Ulcerative Colitis. Ph.D. Thesis, Peking Union Medical College Dissertation, Beijing, China, 2023. [Google Scholar]
- Yuan, Q.; Xu, Z.; Wang, W.; Zhu, J.; Wen, C.; Yan, X.; He, J. Effects of chronic unpredictable stress on dynamic weight changes and inflammatory cytokine levels in mice. China Med. Sci. 2021, 11, 25–28. [Google Scholar]
- Kong, X.; Zhang, Z.; Fu, T.; Ji, J.; Yang, J.; Gu, Z. TNF-α regulates microglial activation via the NF-κB signaling pathway in systemic lupus erythematosus with depression. Int. J. Biol. Macromol. 2019, 125, 892–900. [Google Scholar] [CrossRef]
- Yang, Y.; Wang, W.; Li, A.; Han, S.; Liu, J. Effects of high-fat and high-carbohydrate diets on fat metabolism in rats. Chin. J. Anim. Nutr. 2017, 29, 2603–2612. [Google Scholar]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group | Feed | CUMS |
|---|---|---|
| Control Group(CG) | SD | N/A |
| SD | SD | Yes |
| SD + FP | 50%SD + 50%FP | Yes |
| HFHSD | HFHSD | Yes |
| HFHSD + FP | 50%HFHSD + 50%FP | Yes |
| HFHSD + BP | 50%HFHSD + 50%BP | Yes |
| Stressors | Implementation | Date of Implementation |
|---|---|---|
| Damp sawdust | Add 200 mL water to 100 g of bedding for 24 h. | Date 1, 10, 18, 25 |
| Cage tilting | Tilt the cage at 45° and treat for 24 h. | Date 2, 8, 16, 22 |
| Inversion light/dark cycle | Lights were not operated off at night, and rat cages were placed under a black cloth during the day (treated for 24 h). | Date 3, 11, 21, 26 |
| Cage shaking | Shake each cage of mice horizontally at high speed for 5 min. | Date 4, 9, 15, 23 |
| Forced swimming | Mice were placed in an open cylinder with water at 23 ± 1 °C and a depth of 20 cm for 6 min. | Date 5, 13, 19, 28 |
| Restraint | Mice were confined to 50 mL centrifuge tubes with ventilation holes for 4 h. | Date 6, 12, 20, 27 |
| Suspension | Mice were suspended 1 cm from the tail for 6 min. | Date 7, 14, 17, 24 |
| Gene | Primer Sequence (F) | Primer Sequence (R) |
|---|---|---|
| GAPDH | TCTCCTGCGACTTCAACA | TGTAGCCGTATTCATTGTCA |
| TNF-α | ACTGAACTTCGGGGTGATCG | CCACTTGGTGGTTTGTGAGTG |
| IL-1β | CTTCAGGCAGGCAGTATC | CAGCAGGTTATCATCATCATC |
| IL-6 | ACAAAGCCAGAGTCCTTCAGAG | AGGAGAGCATTGGAAATTGGG |
| BDNF | TCTACCCGACTCATGCTTGC | TCACTGTGAAGCCAGATCGC |
| TPH2 | TCTACCCGACTCATGCTTGC | TCACTGTGAAGCCAGATCGC |
| Behavioral Experiment | Group | Cohen’s d |
|---|---|---|
| Immobility time on FST in mice | CUMS + SD vs. CUMS + SD + FP | 1.29 |
| CUMS + HFHSD vs. CUMS + HFHSD + FP | 1.58 | |
| Immobility time on TST in mice | CUMS + SD vs. CUMS + SD + FP | 1.03 |
| CUMS + HFHSD vs. CUMS + HFHSD + FP | 0.96 | |
| Distance of the central/total area on OFT | CUMS + SD vs. CUMS + SD + FP | −2.14 |
| CUMS + HFHSD vs. CUMS + HFHSD + FP | −0.18 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, X.; Mao, R.; Zou, Y.; Yue, W.; Dong, W.; Zhang, Y. Preliminary Exploration on the Effects of a Novel Antidepressant Formula Food in a Mouse Model. Foods 2025, 14, 1640. https://doi.org/10.3390/foods14091640
Chen X, Mao R, Zou Y, Yue W, Dong W, Zhang Y. Preliminary Exploration on the Effects of a Novel Antidepressant Formula Food in a Mouse Model. Foods. 2025; 14(9):1640. https://doi.org/10.3390/foods14091640
Chicago/Turabian StyleChen, Xu, Ruoxi Mao, Yunxia Zou, Wentian Yue, Wenwen Dong, and Yali Zhang. 2025. "Preliminary Exploration on the Effects of a Novel Antidepressant Formula Food in a Mouse Model" Foods 14, no. 9: 1640. https://doi.org/10.3390/foods14091640
APA StyleChen, X., Mao, R., Zou, Y., Yue, W., Dong, W., & Zhang, Y. (2025). Preliminary Exploration on the Effects of a Novel Antidepressant Formula Food in a Mouse Model. Foods, 14(9), 1640. https://doi.org/10.3390/foods14091640