
Synergistic Neuroprotection Through Epigenetic Modulation by Combined Curcumin-Enriched Turmeric Extract and L-Ascorbic Acid in Oxidative Stress-Induced SH-SY5Y Cell Damage

 , , and
, , and
Abstract
1. Introduction
2. Materials and Methods
2.1. Test Substances Used in the Study
2.2. Evaluation of Biological Activities
2.2.1. DPPH Assay for Free Radical Scavenging
2.2.2. FRAP Assay: Ferric Reducing Antioxidant Power
2.2.3. Radical Scavenging Assay for ABTS
2.2.4. Assay for Cyclooxygenase-2 (COX-2) Inhibition
2.3. Evaluation of the Combination Index (CI) and Dose Reduction Index (DRI)
2.3.1. Calculating the Combination Index
2.3.2. Calculating the Dose Reduction Index
2.4. Cell Culture
2.5. Cell Viability Assay
2.6. Determination of Oxidative Stress Markers
2.6.1. Malondialdehyde (MDA)
2.6.2. Intracellular Reactive Oxygen Species (ROS)
2.7. Determination of Antioxidant Enzyme Activities
2.7.1. Catalase (CAT) Activity
2.7.2. Superoxide Dismutase (SOD) Activity
2.7.3. Glutathione Peroxidase (GSH-Px) Activity
2.8. Western Blotting Analysis
2.9. Statistical Analysis
3. Results
3.1. Synergistic Effects of Curcumin-Enriched Turmeric Extract and L-Ascorbic Acid
3.2. Hydrogen Peroxide (H2O2)-Induced Oxidative Stress Model
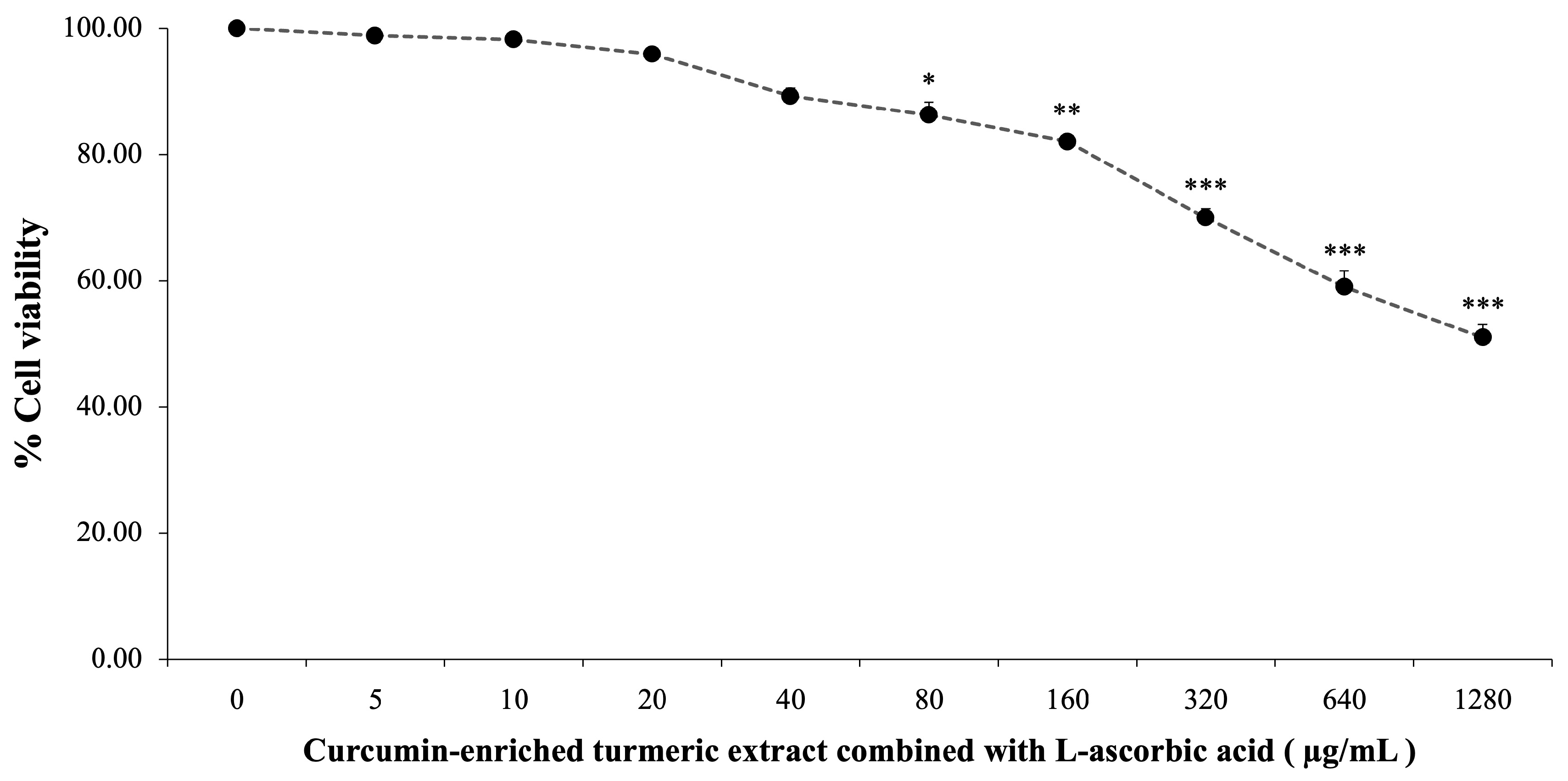
3.3. Cytotoxicity of Curcumin-Enriched Turmeric Extract Combined with L-Ascorbic Acid
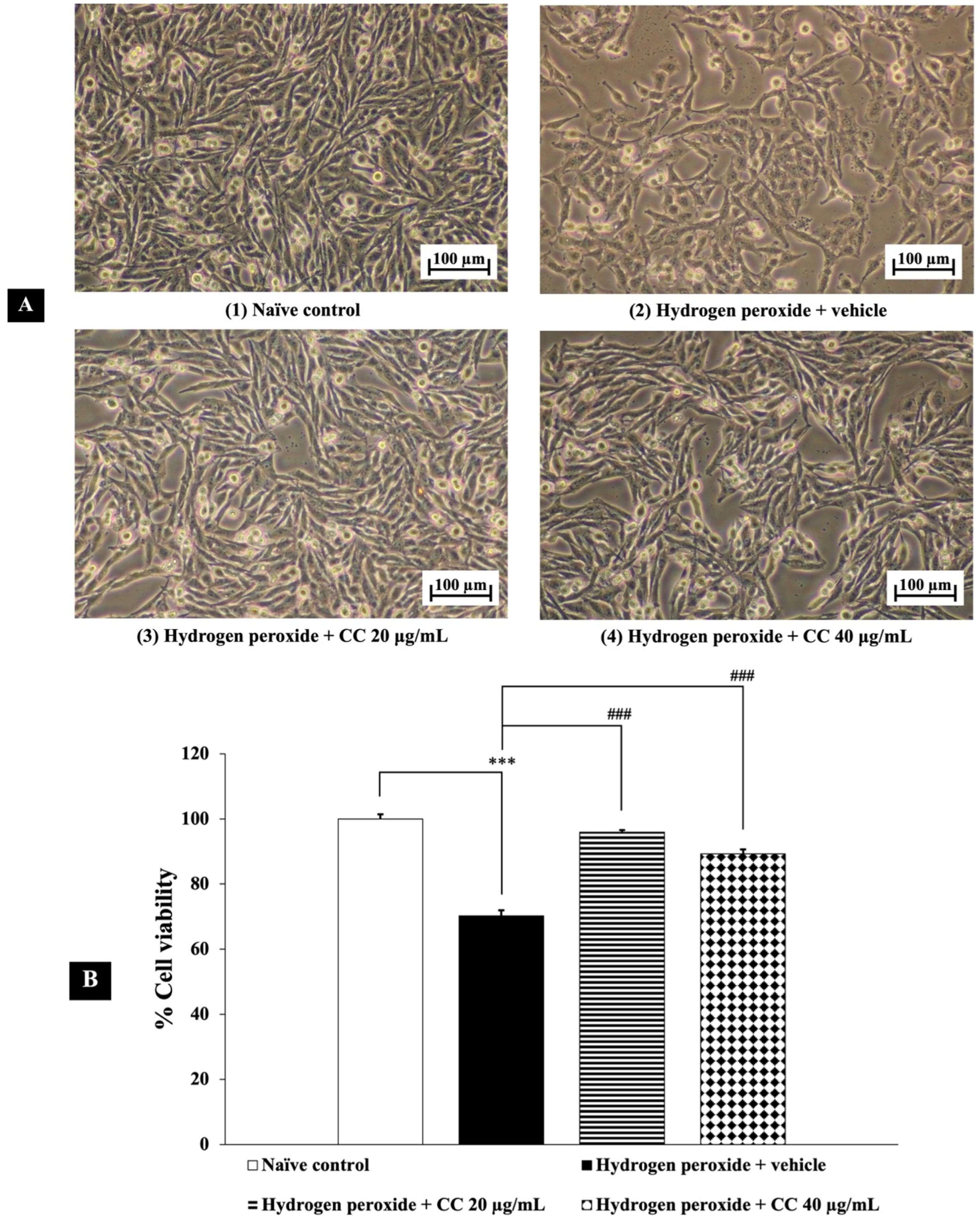
3.4. Neuroprotective Effects of Curcumin-Enriched Turmeric Extract Combined with L-Ascorbic Acid on H2O2-Induced Oxidative Damage in SH-SY5Y Cell
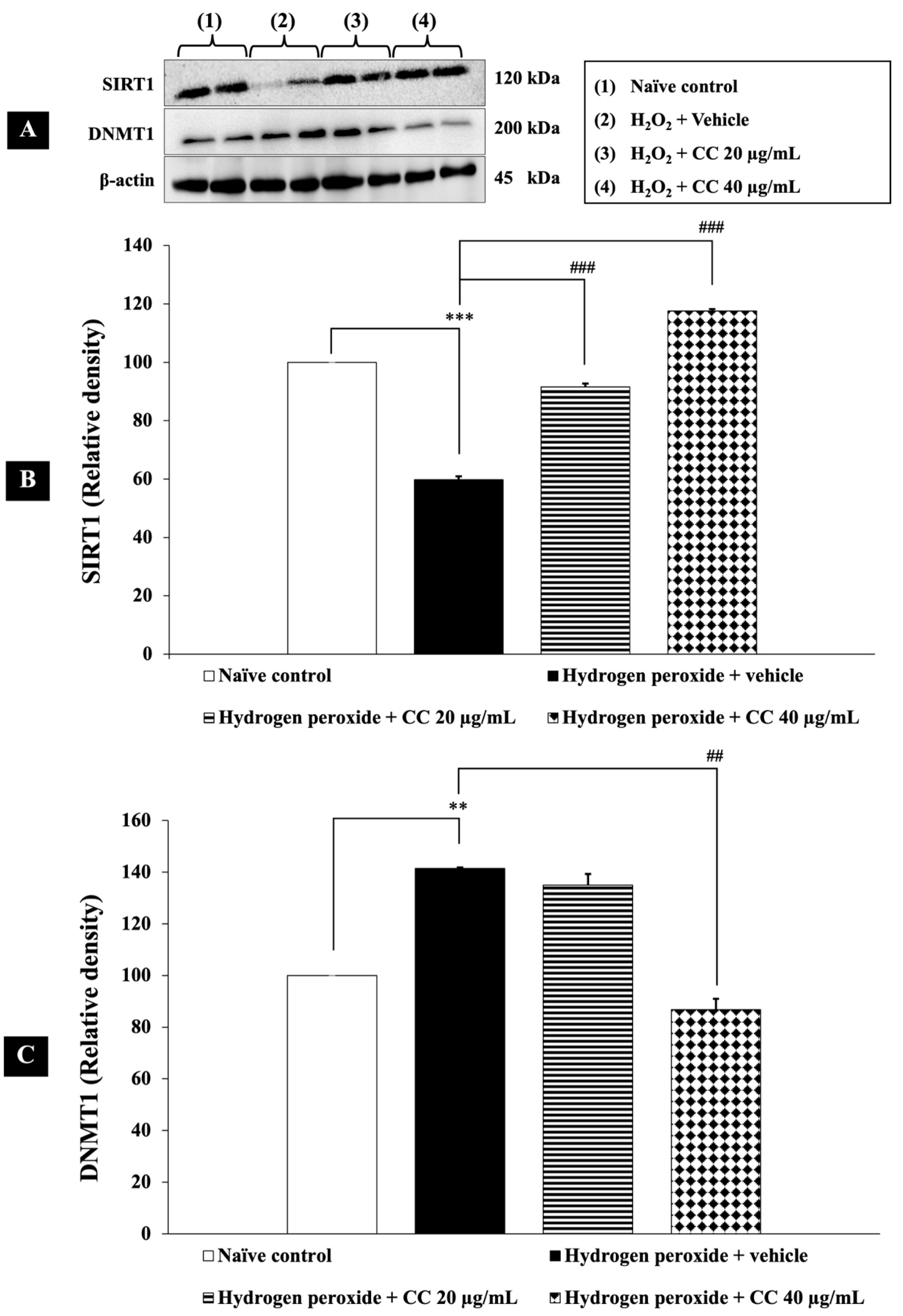
3.5. Epigenetic Modulation by Curcumin-Enriched Turmeric Extract Combined with L-Ascorbic Acid
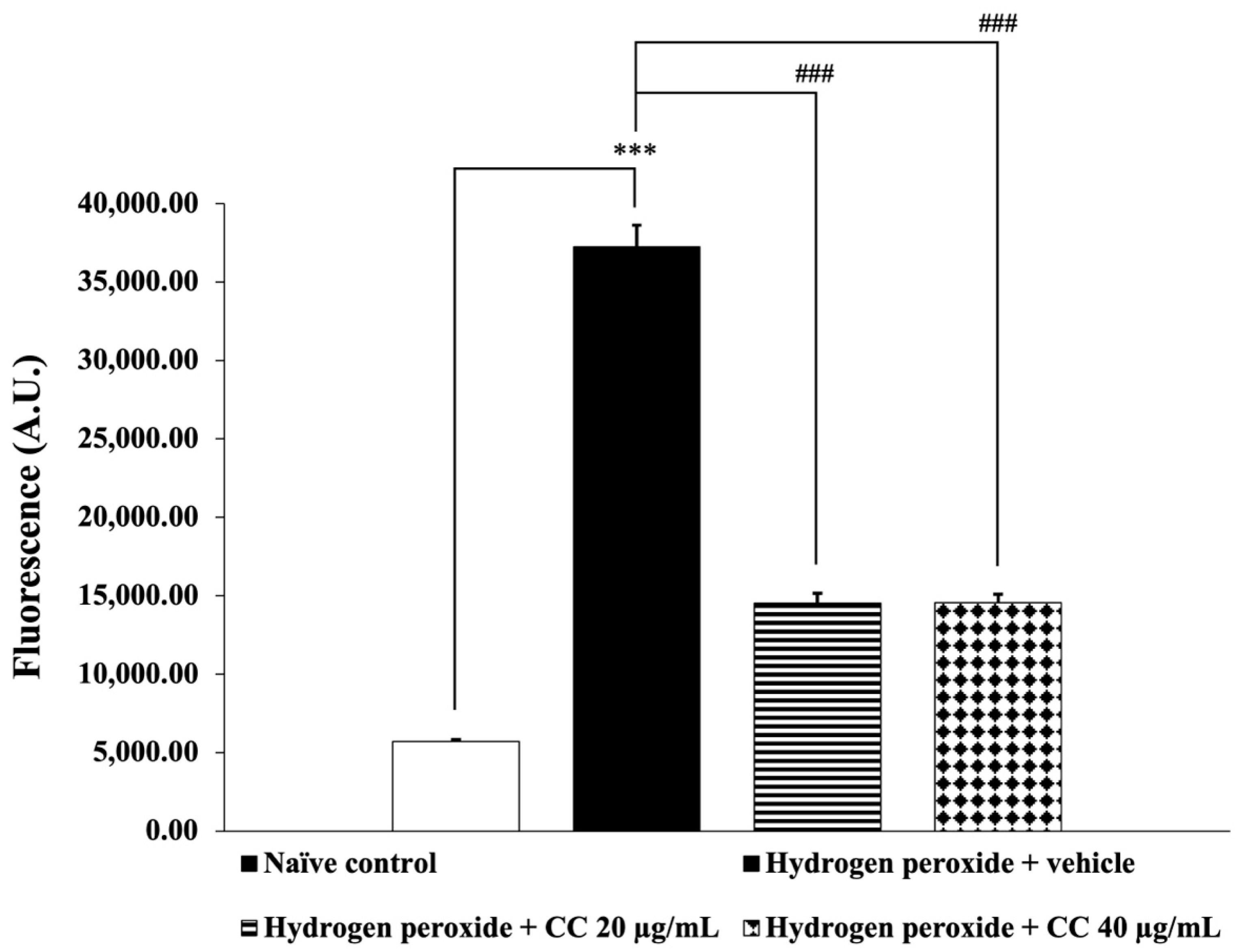
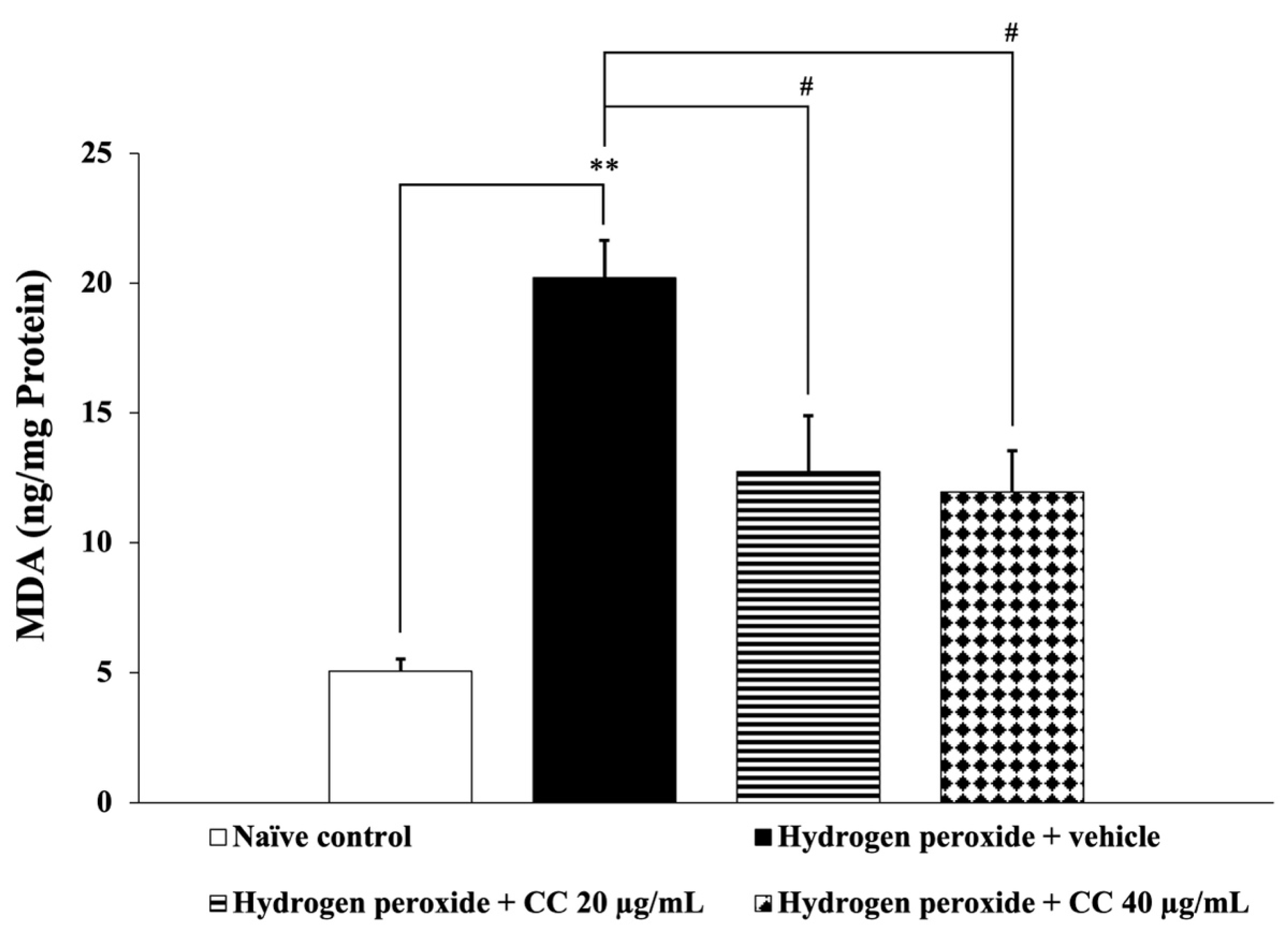
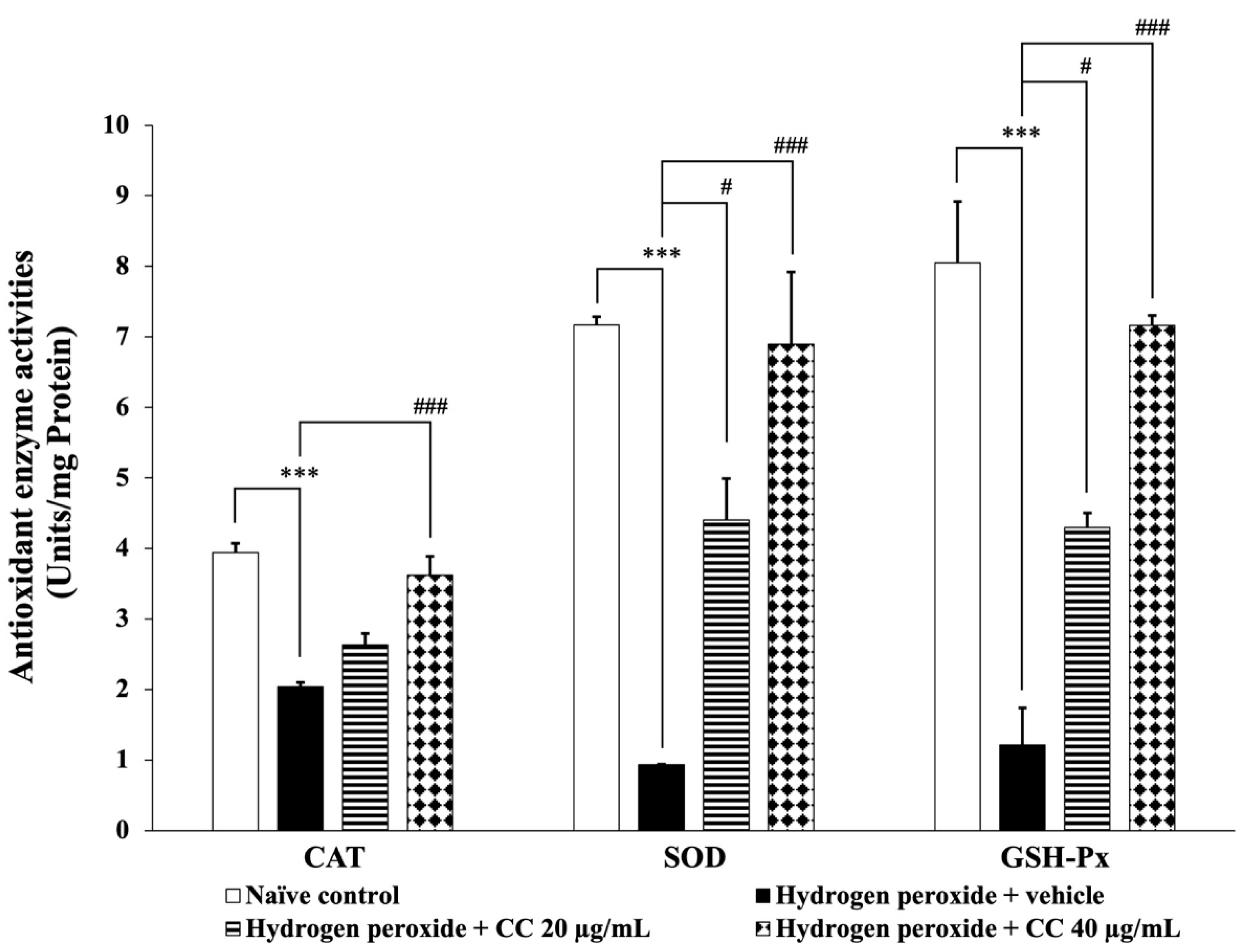
3.6. Effects of Curcumin-Enriched Turmeric Extract Combined with L-Ascorbic Acid on Oxidative Stress Markers
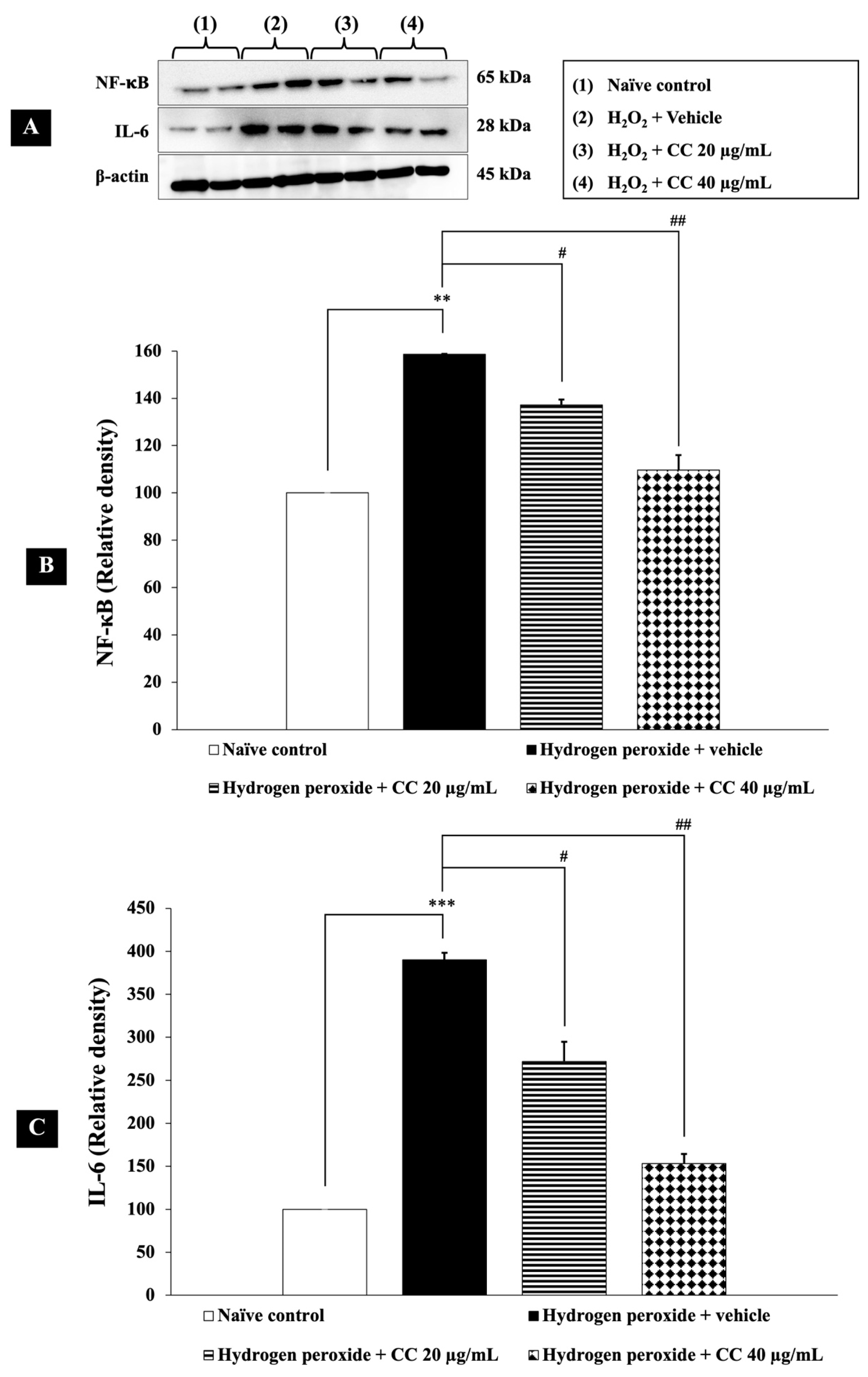
3.7. Effects of Curcumin-Enriched Turmeric Extract Combined with L-Ascorbic Acid on Inflammatory Markers
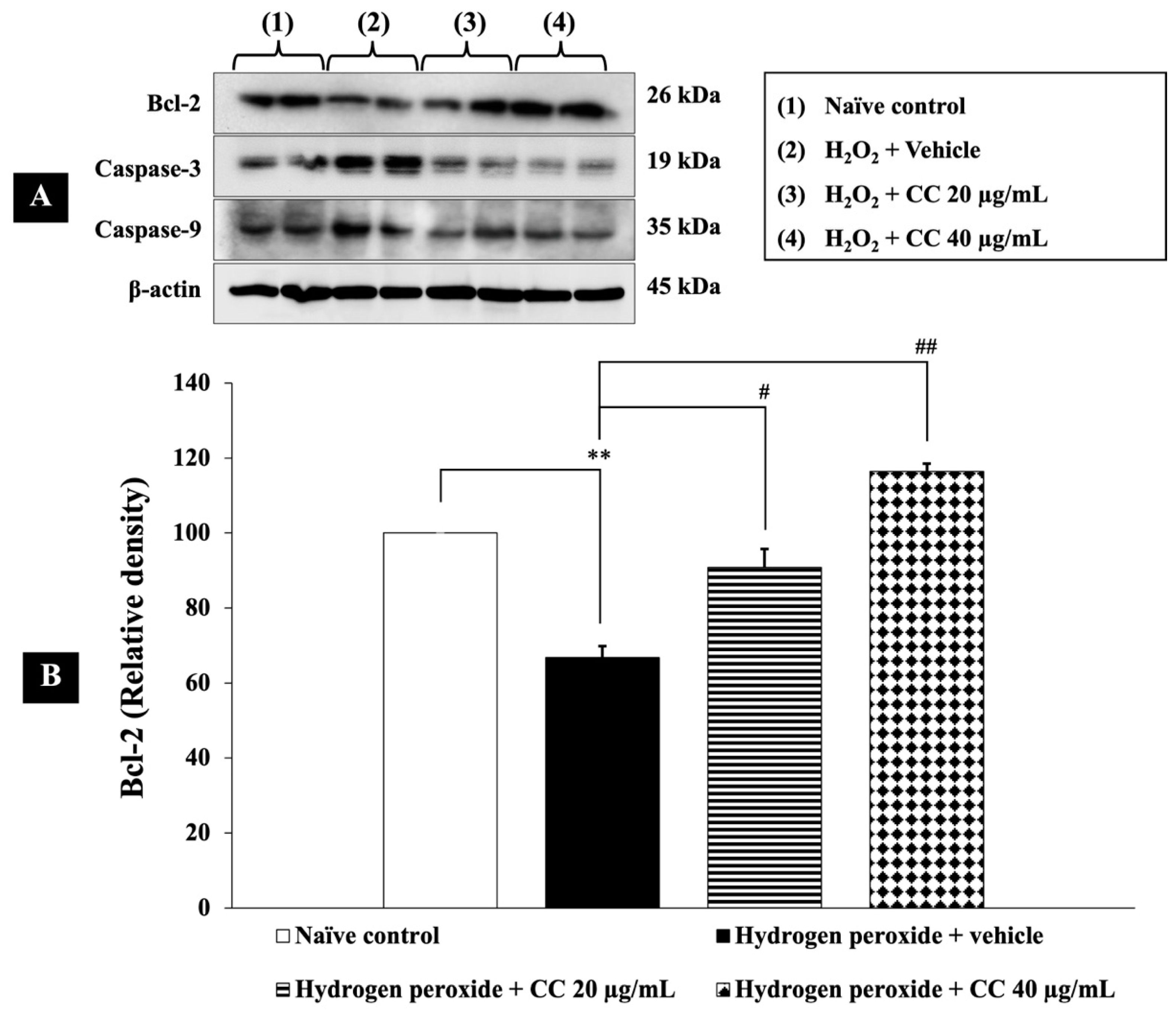
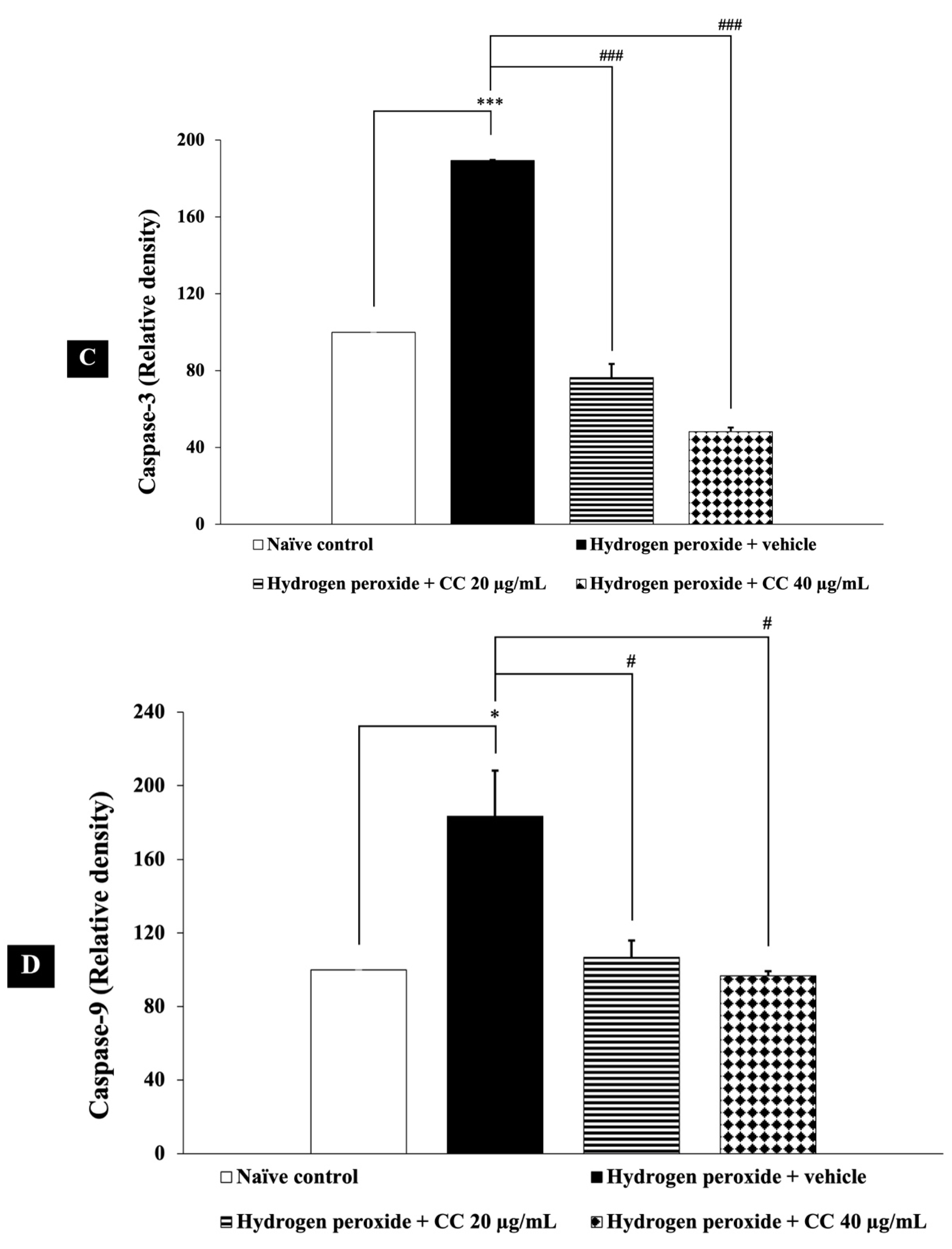
3.8. Effects of Curcumin-Enriched Turmeric Extract Combined with L-Ascorbic Acid on Apoptotic Markers
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Erkkinen, M.G.; Kim, M.O.; Geschwind, M.D. Clinical Neurology and Epidemiology of the Major Neurodegenerative Diseases. Cold Spring Harb. Perspect. Biol. 2018, 10, a033118. [Google Scholar] [CrossRef] [PubMed]
- Lamptey, R.N.L.; Chaulagain, B.; Trivedi, R.; Gothwal, A.; Layek, B.; Singh, J. A Review of the Common Neurodegenerative Disorders: Current Therapeutic Approaches and the Potential Role of Nanotherapeutics. Int. J. Mol. Sci. 2022, 23, 1851. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Wang, X.; Yang, Z.; Lin, X. Epigenetic Modifications and Antioxidant Defense in Neurological Disorders. Antioxidants 2020, 9, 444. [Google Scholar] [CrossRef]
- Wen, K.-X.; Miliç, J.; El-Khodor, B.; Dhana, K.; Nano, J.; Pulido, T.; Kraja, B.; Zaciragic, A.; Bramer, W.M.; Troup, J.; et al. The Role of DNA Methylation and Histone Modifications in Neurodegenerative Diseases: A Systematic Review. PLoS ONE 2016, 11, e0167201. [Google Scholar] [CrossRef]
- Berson, A.; Nativio, R.; Berger, S.L.; Bonini, N.M. Epigenetic Regulation in Neurodegenerative Diseases. Trends Neurosci. 2018, 41, 587–598. [Google Scholar] [CrossRef]
- Jiao, F.; Gong, Z. The Beneficial Roles of SIRT1 in Neuroinflammation-Related Diseases. Oxidative Med. Cell Longev. 2020, 2020, 6782872. [Google Scholar] [CrossRef]
- Yang, Y.; Liu, Y.; Wang, Y.; Chao, Y.; Zhang, J.; Jia, Y.; Tie, J.; Hu, D. Regulation of SIRT1 and Its Roles in Inflammation. Front. Immunol. 2022, 13, 831168. [Google Scholar] [CrossRef]
- Xylaki, M.; Atzler, B.; Outeiro, T.F. Epigenetics of the Synapse in Neurodegeneration. Curr. Neurol. Neurosci. Rep. 2019, 19, 72. [Google Scholar] [CrossRef]
- Klokkaris, A.; Migdalska-Richards, A. An Overview of Epigenetic Changes in the Parkinson’s Disease Brain. Int. J. Mol. Sci. 2024, 25, 6168. [Google Scholar] [CrossRef]
- Zhang, Y.; Kutateladze, T.G. Diet and the Epigenome. Nat. Commun. 2018, 9, 3375. [Google Scholar] [CrossRef]
- Tiffon, C. The Impact of Nutrition and Environmental Epigenetics on Human Health and Disease. Int. J. Mol. Sci. 2018, 19, 3425. [Google Scholar] [CrossRef] [PubMed]
- Hassan, F.U.; Rehman, M.S.; Khan, M.S.; Ali, M.A.; Javed, A.; Nawaz, A.; Yang, C. Curcumin as an Alternative Epigenetic Modulator: Mechanism of Action and Potential Effects. Front. Genet. 2019, 10, 514. [Google Scholar] [CrossRef] [PubMed]
- Abdul-Rahman, T.; Awuah, W.A.; Mikhailova, T.; Kalmanovich, J.; Mehta, A.; Ng, J.C.; Coghlan, M.A.; Zivcevska, M.; Tedeschi, A.J.; de Oliveira, E.C.; et al. Antioxidant, Anti-Inflammatory, and Epigenetic Potential of Curcumin in Alzheimer’s Disease. Biofactors 2024, 50, 693–708. [Google Scholar] [CrossRef] [PubMed]
- Mudgal, R.; Sharma, S.; Singh, S.; Ravichandiran, V. The Neuroprotective Effect of Ascorbic Acid Against Imidacloprid-Induced Neurotoxicity and the Role of HO-1 in Mice. Front. Neurol. 2023, 14, 1130575. [Google Scholar] [CrossRef]
- Jiaranaikulwanitch, J.; Pandith, H.; Tadtong, S.; Thammarat, P.; Jiranusornkul, S.; Chauthong, N.; Nilkosol, S.; Vajragupta, O. Novel Multifunctional Ascorbic Triazole Derivatives for Amyloidogenic Pathway Inhibition, Anti-Inflammation, and Neuroprotection. Molecules 2021, 26, 1562. [Google Scholar] [CrossRef]
- Hussain, S.A.; Hameed, A.; Nazir, Y.; Naz, T.; Wu, Y.; Suleria, H.A.R.; Song, Y. Microencapsulation and the Characterization of Polyherbal Formulation (PHF) Rich in Natural Polyphenolic Compounds. Nutrients 2018, 10, 843. [Google Scholar] [CrossRef]
- Karole, S.; Shrivastava, S.; Thomas, S.; Soni, B.; Khan, S.; Dubey, J.; Dubey, S.P.; Khan, N.; Jain, D.K. Polyherbal Formulation Concept for Synergic Action. J. Drug Deliv. Ther. 2019, 9, 453–466. [Google Scholar] [CrossRef]
- Che, C.-T.; Wang, Z.J.; Chow, M.S.S.; Lam, C.W.K. Herb-Herb Combination for Therapeutic Enhancement and Advancement: Theory, Practice and Future Perspectives. Molecules 2013, 18, 5125–5141. [Google Scholar] [CrossRef]
- Mairuae, N.; Palachai, N.; Noisa, P. The neuroprotective effects of the combined extract of mulberry fruit and mulberry leaf against hydrogen peroxide-induced cytotoxicity in SH-SY5Y Cells. BMC Complement. Med. Ther. 2023, 23, 117. [Google Scholar] [CrossRef]
- Mairuae, N.; Noisa, P.; Palachai, N. Phytosome-Encapsulated 6-Gingerol- and 6-Shogaol-Enriched Extracts from Zingiber officinale Roscoe Protect Against Oxidative Stress-Induced Neurotoxicity. Molecules 2024, 29, 6046. [Google Scholar] [CrossRef]
- Mairuae, N.; Palachai, N.; Noisa, P. An anthocyanin-rich extract from Zea mays L. var. ceratina alleviates neuronal cell death caused by hydrogen peroxide-induced cytotoxicity in SH-SY5Y cells. BMC Complement. Med. Ther. 2024, 24, 162. [Google Scholar] [CrossRef] [PubMed]
- Palachai, N.; Wattanathorn, J.; Muchimapura, S.; Thukham-Mee, W. Antimetabolic Syndrome Effect of Phytosome Containing the Combined Extracts of Mulberry and Ginger in an Animal Model of Metabolic Syndrome. Oxidative Med. Cell Longev. 2019, 2019, 5972575. [Google Scholar] [CrossRef] [PubMed]
- Palachai, N.; Wattanathorn, J.; Muchimapura, S.; Thukham-Mee, W. Phytosome Loading the Combined Extract of Mulberry Fruit and Ginger Protects against Cerebral Ischemia in Metabolic Syndrome Rats. Oxidative Med. Cell Longev. 2020, 2020, 5305437. [Google Scholar] [CrossRef] [PubMed]
- Wattanathorn, J.; Palachai, N.; Thukham-Mee, W.; Muchimapura, S. Memory-Enhancing Effect of a Phytosome Containing the Combined Extract of Mulberry Fruit and Ginger in an Animal Model of Ischemic Stroke with Metabolic Syndrome. Oxidative Med. Cell Longev. 2020, 2020, 3096826. [Google Scholar] [CrossRef]
- Palachai, N.; Thukham-mee, W.; Wattanathorn, J. The Protective Effect against Lung Injury of Phytosome Containing the Extract of Purple Waxy Corn Tassel in an Animal Model of PM2.5-Induced Lung Inflammation. Foods 2024, 13, 3258. [Google Scholar] [CrossRef]
- Tallarida, R.J. Quantitative Methods for Assessing Drug Synergism. Genes Cancer 2011, 2, 1003–1008. [Google Scholar] [CrossRef]
- Pezzani, R.; Salehi, B.; Vitalini, S.; Iriti, M.; Zuñiga, F.A.; Sharifi-Rad, J.; Martorell, M.; Martins, N. Synergistic Effects of Plant Derivatives and Conventional Chemotherapeutic Agents: An Update on the Cancer Perspective. Medicina 2019, 55, 110. [Google Scholar] [CrossRef]
- Dei Cas, M.; Ghidoni, R. Dietary Curcumin: Correlation between Bioavailability and Health Potential. Nutrients 2019, 11, 2147. [Google Scholar] [CrossRef]
- Huang, M.; Wu, Q.; Jiang, Z.H. Epigenetic Alterations Under Oxidative Stress in Stem Cells. Oxidative Med. Cell Longev. 2022, 2022, 6439097. [Google Scholar] [CrossRef]
- Manjula, R.; Anuja, K.; Alcain, F.J. SIRT1 and SIRT2 Activity Control in Neurodegenerative Diseases. Front. Pharmacol. 2021, 11, 585821. [Google Scholar] [CrossRef]
- Wang, W.; Zhao, X.; Shao, Y.; Duan, X.; Wang, Y.; Li, J.; Li, J.; Li, D.; Li, X.; Wong, J. Mutation-Induced DNMT1 Cleavage Drives Neurodegenerative Disease. Sci. Adv. 2021, 7, eabe8511. [Google Scholar] [CrossRef] [PubMed]
- Aramouni, K.; Assaf, R.; Shaito, A.; Fardoun, M.; Al-Asmakh, M.; Sahebkar, A.; Eid, A.H. Biochemical and Cellular Basis of Oxidative Stress: Implications for Disease Onset. J. Cell Physiol. 2023, 238, 1951–1963. [Google Scholar] [CrossRef] [PubMed]
- Olmos, Y.; Sánchez-Gómez, F.J.; Wild, B.; García-Quintans, N.; Cabezudo, S.; Lamas, S.; Monsalve, M. SirT1 Regulation of Antioxidant Genes Is Dependent on the Formation of a FoxO3a/PGC-1α Complex. Antioxid. Redox Signal. 2013, 19, 1507–1521. [Google Scholar] [CrossRef]
- Maresca, A.; Del Dotto, V.; Capristo, M.; Scimonelli, E.; Tagliavini, F.; Morandi, L.; Tropeano, C.V.; Caporali, L.; Mohamed, S.; Roberti, M.; et al. DNMT1 Mutations Leading to Neurodegeneration Paradoxically Reflect on Mitochondrial Metabolism. Hum. Mol. Genet. 2020, 29, 1864–1881. [Google Scholar] [CrossRef] [PubMed]
- Singh, M.; Saxena, S.; Mohan, K.N. DNMT1 Downregulation as Well as Its Overexpression Distinctly Affect Mostly Overlapping Genes Implicated in Schizophrenia, Autism Spectrum, Epilepsy, and Bipolar Disorders. Front. Mol. Neurosci. 2023, 16, 1275697. [Google Scholar] [CrossRef]
- Gonçalves, A.C.; Alves, R.; Baldeiras, I.; Marques, B.; Oliveiros, B.; Pereira, A.; Nascimento Costa, J.M.; Cortesão, E.; Mota Vieira, L.; Sarmento Ribeiro, A.B. DNA Methylation Is Correlated with Oxidative Stress in Myelodysplastic Syndrome—Relevance as Complementary Prognostic Biomarkers. Cancers 2021, 13, 3138. [Google Scholar] [CrossRef]
- Lingappan, K. NF-κB in Oxidative Stress. Curr. Opin. Toxicol. 2018, 7, 81–86. [Google Scholar] [CrossRef]
- Guo, Q.; Jin, Y.; Chen, X.; Ye, X.; Shen, X.; Lin, M.; Zeng, C.; Zhou, T.; Zhang, J. NF-κB in Biology and Targeted Therapy: New Insights and Translational Implications. Signal Transduct. Target. Ther. 2024, 9, 53. [Google Scholar] [CrossRef]
- Anilkumar, S.; Wright-Jin, E. NF-κB as an Inducible Regulator of Inflammation in the Central Nervous System. Cells 2024, 13, 485. [Google Scholar] [CrossRef]
- Chen, M.; Chen, Z.; Huang, D.; Sun, C.; Xie, J.; Chen, T.; Zhao, X.; Huang, Y.; Li, D.; Wu, B.; et al. Myricetin Inhibits TNF-α-Induced Inflammation in A549 Cells via the SIRT1/NF-κB Pathway. Pulm. Pharmacol. Ther. 2020, 65, 102000. [Google Scholar] [CrossRef]
- Hussar, P. Apoptosis Regulators Bcl-2 and Caspase-3. Encyclopedia 2022, 2, 1624–1636. [Google Scholar] [CrossRef]
- Hollville, E.; Romero, S.E.; Deshmukh, M. Apoptotic Cell Death Regulation in Neurons. FEBS J. 2019, 286, 3276–3298. [Google Scholar] [CrossRef] [PubMed]
- Xia, X.; Wang, S.; Wu, L.; Li, G.; Hou, K.; Yu, A.; Yang, Z. TIPE2 Attenuates Neuroinflammation and Brain Injury through Bcl-2/Bax/Cleaved Caspase-3 Apoptotic Pathways after Intracerebral Hemorrhage in Mice. Brain Res. Bull. 2022, 191, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.T.; Gu, W. SIRT1: Regulator of p53 Deacetylation. Genes Cancer 2013, 4, 112–117. [Google Scholar] [CrossRef] [PubMed]
- Hori, Y.S.; Kuno, A.; Hosoda, R.; Horio, Y. Regulation of FOXOs and p53 by SIRT1 modulators under oxidative stress. PLoS ONE 2013, 8, e73875. [Google Scholar] [CrossRef]
- Hervouet, E.; Cheray, M.; Vallette, F.M.; Cartron, P.-F. DNA Methylation and Apoptosis Resistance in Cancer Cells. Cells 2013, 2, 545–573. [Google Scholar] [CrossRef]
- Zhang, X.; Bustos, M.A.; Shoji, Y.; Ramos, R.I.; Iida, Y.; Gentry, R.; Takeshima, T.-L.; Hoon, D.S.B. Acetylated DNMT1 Downregulation and Related Regulatory Factors Influence Metastatic Melanoma Patients Survival. Cancers 2021, 13, 4691. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameter | Unit | Curcumin-Enriched Turmeric Extract | L-Ascorbic Acid | Curcumin-Enriched Turmeric Extract Combined with L-Ascorbic Acid |
|---|---|---|---|---|
| Antioxidant activities | ||||
| DPPH | EC50 (μg/mL) | 67.36 ± 4.25 | 13.77 ± 0.18 | 4.70 ± 1.05 ***, ### |
| FRAP | EC50 (μg/mL) | 26.55 ± 0.94 | 12.40 ± 2.35 | 4.64 ± 0.88 ***, # |
| ABTS | EC50 (μg/mL) | 75.24 ± 0.73 | 19.03 ± 2.62 | 8.06 ± 1.35 ***, ### |
| Anti-inflammatory activity | ||||
| COX-II | EC50 (μg/mL) | 77.63 ± 0.04 | 121.18 ± 0.12 | 33.82 ± 0.05 ***, ### |
| Parameter | Combination Index (Type of Interaction) | Dose Reduction Index | |
|---|---|---|---|
| Curcumin-Enriched Turmeric Extract | L-Ascorbic Acid | ||
| Antioxidant activities | |||
| DPPH | 0.41 ± 0.05 (synergism) | 15.59 ± 1.68 | 3.20 ± 0.35 |
| FRAP | 0.55 ± 0.02 (synergism) | 6.20 ± 0.73 | 2.67 ± 0.00 |
| ABTS | 0.54 ± 0.06 (synergism) | 9.88 ± 0.94 | 2.46 ± 0.25 |
| Anti-inflammatory activity | |||
| COX-II | 0.71 ± 0.00 (synergism) | 2.30 ± 0.00 | 3.58 ± 0.00 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Khongrum, J.; Mairuae, N.; Thanchomnang, T.; Zhang, M.; Bai, G.; Palachai, N. Synergistic Neuroprotection Through Epigenetic Modulation by Combined Curcumin-Enriched Turmeric Extract and L-Ascorbic Acid in Oxidative Stress-Induced SH-SY5Y Cell Damage. Foods 2025, 14, 892. https://doi.org/10.3390/foods14050892
Khongrum J, Mairuae N, Thanchomnang T, Zhang M, Bai G, Palachai N. Synergistic Neuroprotection Through Epigenetic Modulation by Combined Curcumin-Enriched Turmeric Extract and L-Ascorbic Acid in Oxidative Stress-Induced SH-SY5Y Cell Damage. Foods. 2025; 14(5):892. https://doi.org/10.3390/foods14050892
Chicago/Turabian StyleKhongrum, Jurairat, Nootchanat Mairuae, Tongjit Thanchomnang, Man Zhang, Gang Bai, and Nut Palachai. 2025. "Synergistic Neuroprotection Through Epigenetic Modulation by Combined Curcumin-Enriched Turmeric Extract and L-Ascorbic Acid in Oxidative Stress-Induced SH-SY5Y Cell Damage" Foods 14, no. 5: 892. https://doi.org/10.3390/foods14050892
APA StyleKhongrum, J., Mairuae, N., Thanchomnang, T., Zhang, M., Bai, G., & Palachai, N. (2025). Synergistic Neuroprotection Through Epigenetic Modulation by Combined Curcumin-Enriched Turmeric Extract and L-Ascorbic Acid in Oxidative Stress-Induced SH-SY5Y Cell Damage. Foods, 14(5), 892. https://doi.org/10.3390/foods14050892