
Prebiotic and Health-Promoting Benefits of Dextran-Type Exopolysaccharide Produced by Leuconostoc mesenteroides SJC113


Abstract
1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Bacterial Strains
2.3. Production and Purification of EPS
2.4. Biological Activities of EPS
2.4.1. Cholesterol-Binding
2.4.2. DPPH Scavenging Activity
2.4.3. Hydroxyl Radical Scavenging Activity
2.5. Prebiotic Properties
2.5.1. Mucin Adhesion Assay
2.5.2. Effect of EPS on Probiotic Adhesion to Mucin
2.5.3. Stimulation of LAB Growth
2.6. Effects of EPS on Fecal Microbiome
2.6.1. Gut Nutrient Medium
2.6.2. Fecal Slurry
2.6.3. Microbiological Analysis
2.7. Statistical Analysis
3. Results and Discussion
3.1. Biological Activities of EPS Produced by Ln. Mesenteroides SJC113
3.1.1. Cholesterol-Lowering Ability
3.1.2. Antioxidant Activity
3.2. Prebiotic Properties
3.2.1. Adhesion to Immobilized Mucin
3.2.2. Adhesion to Mucin in the Presence of EPS
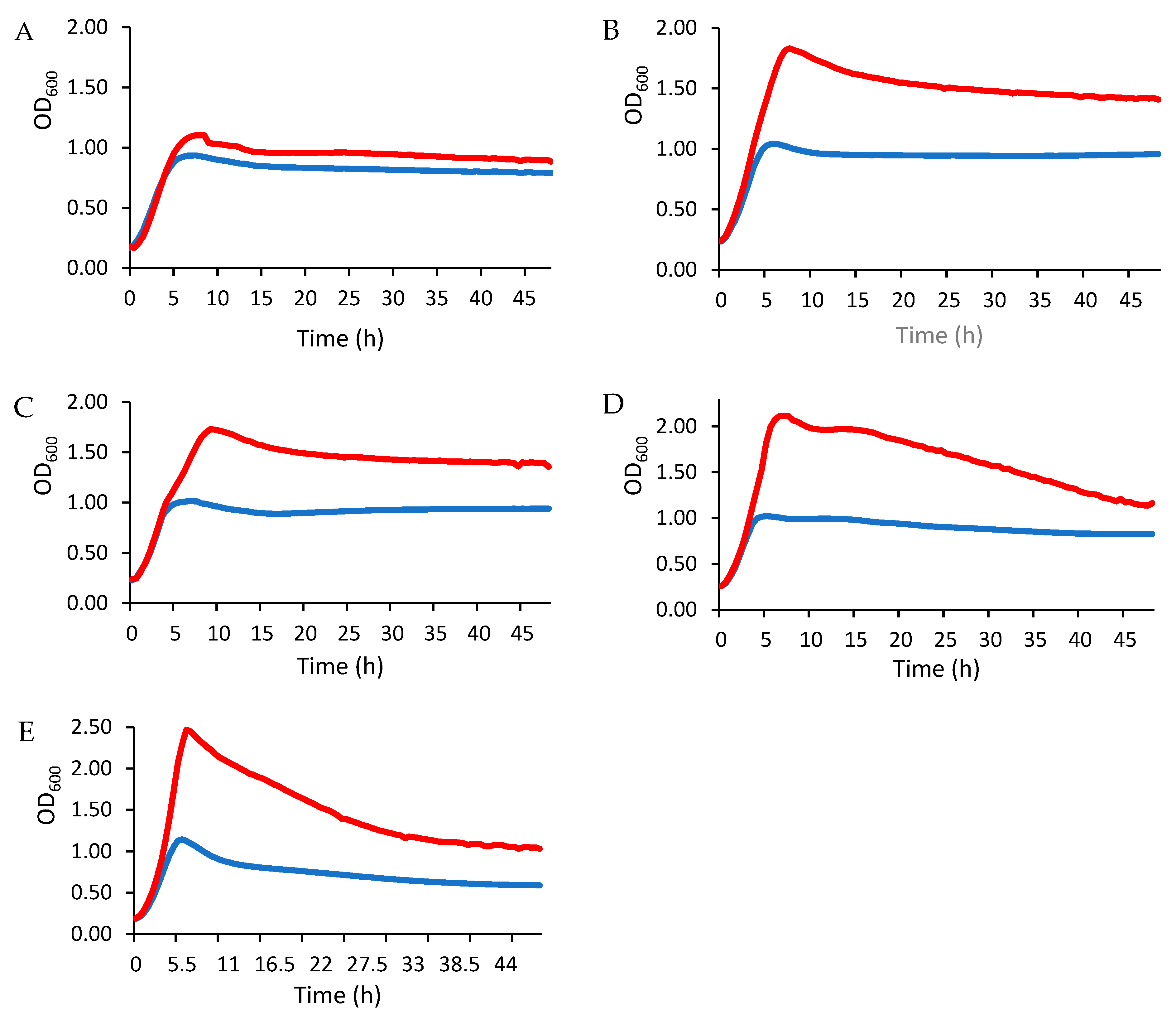
3.2.3. Stimulation of LAB Growth
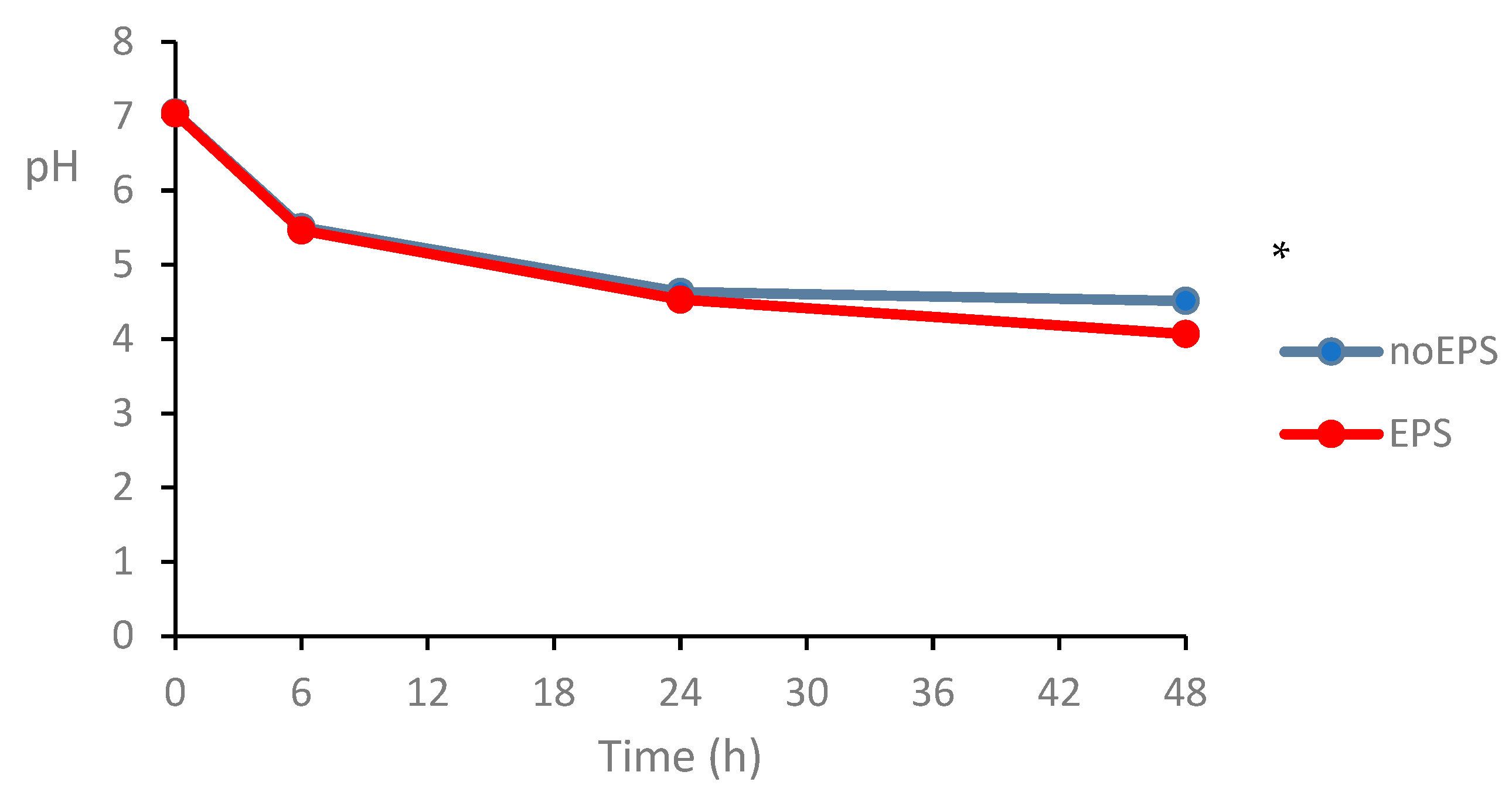
3.2.4. Effects of EPS on Selected Bacterial Groups in Fecal Fermentation
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Rau, S.; Gregg, A.; Yaceczko, S.; Limketkai, B. Prebiotics and probiotics for gastrointestinal disorders. Nutrients 2024, 16, 778. [Google Scholar] [CrossRef] [PubMed]
- Ballini, A.; Charitos, I.A.; Cantore, S.; Topi, S.; Bottalico, L.; Santacroce, L. About functional foods: The probiotics and prebiotics state of art. Antibiotics 2023, 12, 635. [Google Scholar] [CrossRef] [PubMed]
- Chandrasekaran, P.; Weiskirchen, S.; Weiskirchen, R. Effects of probiotics on gut microbiota: An overview. Int. J. Mol. Sci. 2024, 25, 6022. [Google Scholar] [CrossRef] [PubMed]
- Sarita, B.; Samadhan, D.; Hassan, M.Z.; Kovaleva, E.G. A comprehensive review of probiotics and human health-current prospective and applications. Front. Microbiol. 2025, 15, 1487641. [Google Scholar] [CrossRef] [PubMed]
- Jurášková, D.; Ribeiro, S.C.; Silva, C.C. Exopolysaccharides produced by lactic acid bacteria: From biosynthesis to health-promoting properties. Foods 2022, 11, 156. [Google Scholar] [CrossRef] [PubMed]
- Caggianiello, G.; Kleerebezem, M.; Spano, G. Exopolysaccharides produced by lactic acid bacteria: From health-promoting benefits to stress tolerance mechanisms. Appl. Microbiol. Biotechnol. 2016, 100, 3877–3886. [Google Scholar] [CrossRef] [PubMed]
- Davani-Davari, D.; Negahdaripour, M.; Karimzadeh, I.; Seifan, M.; Mohkam, M.; Masoumi, S.J.; Berenjian, A.; Ghasemi, Y. Prebiotics: Definition, types, sources, mechanisms, and clinical applications. Foods 2019, 8, 92. [Google Scholar] [CrossRef] [PubMed]
- Saadat, Y.R.; Khosroushahi, A.Y.; Gargari, B.P. A comprehensive review of anticancer, immunomodulatory and health beneficial effects of the lactic acid bacteria exopolysaccharides. Carbohydr. Polym. 2019, 217, 79–89. [Google Scholar] [CrossRef] [PubMed]
- Kavitake, D.; Veerabhadrappa, B.; Sudharshan, S.; Kandasamy, S.; Devi, P.B.; Dyavaiah, M.; Shetty, P.H. Oxidative stress alleviating potential of galactan exopolysaccharide from Weissella confusa KR780676 in yeast model system. Sci. Rep. 2022, 12, 1089. [Google Scholar] [CrossRef] [PubMed]
- Kusmiati, K.; Nurkanto, A.; Fanani, A.; Nurcahyanto, D.A.; Mamangkey, J.; Marissa, S.; Pasaribu, K.M.; Afiati, F.; Agustini, N.W.S.; Ahmad, R.Z.; et al. Anti-hypercholesterolemia properties of exopolysaccharide from Lactiplantibacillus plantarum MI01: Computational and in vivo approaches. Case Stud. Chem. Environ. Eng. 2025, 11, 101146. [Google Scholar] [CrossRef]
- Korcz, E.; Kerényi, Z.; Varga, L. Dietary fibers, prebiotics, and exopolysaccharides produced by lactic acid bacteria: Potential health benefits with special regard to cholesterol-lowering effects. Food Funct. 2018, 9, 3057–3068. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Han, J.; Ren, Q.; Liu, Z.; Zhang, X.; Wu, Z. The involvement of lactic acid bacteria and their exopolysaccharides in the biosorption and detoxication of heavy metals in the gut. Biol. Trace Elem. Res. 2024, 202, 671–684. [Google Scholar] [CrossRef] [PubMed]
- Senanayake, D.; Ramarao-Milne, P.; Pandey, G.; Hlaing, M.M.; Chandrapala, J.; Torley, P.J.; Terefe, N.S. Genomic insights into exopolysaccharide biosynthesis pathways in novel Lactiplantibacillus plantarum and Leuconostoc mesenteroides strains. LWT 2025, 225, 117863. [Google Scholar] [CrossRef]
- Díaz-Montes, E. Dextran: Sources, structures, and properties. Polysaccharides 2021, 2, 554–565. [Google Scholar] [CrossRef]
- Jurášková, D.; Ribeiro, S.C.; Bastos, R.; Coelho, E.; Coimbra, M.A.; Silva, C.C.G. Exopolysaccharide (EPS) Produced by Leuconostoc mesenteroides SJC113: Characterization of Functional and Technological Properties and Application in Fat-Free Cheese. Macromol 2024, 4, 680–696. [Google Scholar] [CrossRef]
- Domingos-Lopes, M.; Lamosa, P.; Stanton, C.; Ross, R.; Silva, C. Isolation and characterization of an exopolysaccharide-producing Leuconostoc citreum strain from artisanal cheese. Lett. Appl. Microbiol. 2018, 67, 570–578. [Google Scholar] [CrossRef] [PubMed]
- Gawande, K.; Kolhekar, M.; Kumari, M.; Kapila, S.; Sharma, P.; Ali, S.A.; Behare, P.V. Lactic acid bacteria based purified exopolysaccharide showed viscofying and hypercholesterolemic capabilites. Food Hydrocoll. Health 2021, 1, 100042. [Google Scholar] [CrossRef]
- Hu, S.-M.; Zhou, J.-M.; Zhou, Q.-Q.; Li, P.; Xie, Y.-Y.; Zhou, T.; Gu, Q. Purification, characterization and biological activities of exopolysaccharides from Lactobacillus rhamnosus ZFM231 isolated from milk. LWT 2021, 147, 111561. [Google Scholar] [CrossRef]
- Zhang, J.; Liu, L.; Chen, F. Production and characterization of exopolysaccharides from Chlorella zofingiensis and Chlorella vulgaris with anti-colorectal cancer activity. Int. J. Biol. Macromol. 2019, 134, 976–983. [Google Scholar] [CrossRef] [PubMed]
- Dhanani, A.; Bagchi, T. The expression of adhesin EF-Tu in response to mucin and its role in Lactobacillus adhesion and competitive inhibition of enteropathogens to mucin. J. Appl. Microbiol. 2013, 115, 546–554. [Google Scholar] [CrossRef] [PubMed]
- Liu, K.; Zhang, Y.; Yu, Z.; Xu, Q.; Zheng, N.; Zhao, S.; Huang, G.; Wang, J. Ruminal microbiota-host interaction and its effect on nutrient metabolism. Anim. Nutr. 2021, 7, 49–55. [Google Scholar] [CrossRef] [PubMed]
- Molly, K.; Vande Woestyne, M.; Verstraete, W. Development of a 5-step multi-chamber reactor as a simulation of the human intestinal microbial ecosystem. Appl. Microbiol. Biotechnol. 1993, 39, 254–258. [Google Scholar] [CrossRef] [PubMed]
- Tamargo, A.; Cueva, C.; Álvarez, M.D.; Herranz, B.; Bartolomé, B.; Moreno-Arribas, M.V.; Laguna, L. Influence of viscosity on the growth of human gut microbiota. Food Hydrocoll. 2018, 77, 163–167. [Google Scholar] [CrossRef]
- Machado, F.; Gómez-Domínguez, I.; Hurtado-Ribeira, R.; Martin, D.; Coimbra, M.A.; Del Castillo, M.D.; Coreta-Gomes, F. In vitro human colonic fermentation of coffee arabinogalactan and melanoidin-rich fractions. Int. J. Biol. Macromol. 2024, 275, 133740. [Google Scholar] [CrossRef] [PubMed]
- Cueva, C.; Jiménez-Girón, A.; Muñoz-González, I.; Esteban-Fernández, A.; Gil-Sánchez, I.; Dueñas, M.; Martín-Álvarez, P.J.; Pozo-Bayón, M.A.; Bartolomé, B.; Moreno-Arribas, M.V. Application of a new Dynamic Gastrointestinal Simulator (SIMGI) to study the impact of red wine in colonic metabolism. Food Res. Int. 2015, 72, 149–159. [Google Scholar] [CrossRef]
- Barroso, E.; Cueva, C.; Peláez, C.; Martínez-Cuesta, M.C.; Requena, T. Development of human colonic microbiota in the computer-controlled dynamic SIMulator of the GastroIntestinal tract SIMGI. LWT 2015, 61, 283–289. [Google Scholar] [CrossRef]
- Hashemi, S.M.B.; Abedi, E.; Kaveh, S.; Mousavifard, M. Hypocholesterolemic, antidiabetic and bioactive properties of ultrasound-stimulated exopolysaccharide produced by Lactiplantibacillus plantarum strains. Bioact. Carbohydr. Diet. Fibre 2022, 28, 100334. [Google Scholar] [CrossRef]
- Gulcin, A.; Cagatay, G.; Cilak, G.O.; Avci, E. Probable novel probiotics: EPS production, cholesterol removal and glycocholate deconjugation of Lactobacillus plantarum Ga06 and Ga11 isolated from local handmade-cheese. J. Microbiol. Biotechnol. Food Sci. 2020, 10, 83–86. [Google Scholar] [CrossRef]
- Sasikumar, K.; Kozhummal Vaikkath, D.; Devendra, L.P.; Nampoothiri, K.M. An exopolysaccharide (EPS) from a Lactobacillus plantarum BR2 with potential benefits for making functional foods. Bioresour. Technol. 2017, 241, 1152–1156. [Google Scholar] [CrossRef] [PubMed]
- Domingos-Lopes, M.; Stanton, C.; Ross, R.P.; Silva, C. Histamine and cholesterol lowering abilities of lactic acid bacteria isolated from artisanal Pico cheese. J. Appl. Microbiol. 2020, 129, 1428–1440. [Google Scholar] [CrossRef] [PubMed]
- Le, B.; Yang, S.-H. Effect of potential probiotic Leuconostoc mesenteroides FB111 in prevention of cholesterol absorption by modulating NPC1L1/PPARα/SREBP-2 pathways in epithelial Caco-2 cells. Int. Microbiol. 2019, 22, 279–287. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.; Kim, M. Leuconostoc mesenteroides MKSR isolated from kimchi possesses α-glucosidase inhibitory activity, antioxidant activity, and cholesterol-lowering effects. LWT 2019, 116, 108570. [Google Scholar] [CrossRef]
- Park, M.Y.; Kim, J.; Kim, S.; Whang, K.-Y. Lactobacillus curvatus KFP419 and Leuconostoc mesenteroides subsp. mesenteroides KDK411 isolated from kimchi ameliorate hypercholesterolemia in rats. J. Med. Food 2018, 21, 647–653. [Google Scholar] [CrossRef] [PubMed]
- Amiri, S.; Rezaei Mokarram, R.; Sowti Khiabani, M.; Rezazadeh Bari, M.; Alizadeh Khaledabad, M. Exopolysaccharides production by Lactobacillus acidophilus LA5 and Bifidobacterium animalis subsp. lactis BB12: Optimization of fermentation variables and characterization of structure and bioactivities. Int. J. Biol. Macromol. 2019, 123, 752–765. [Google Scholar] [CrossRef] [PubMed]
- Vinoshna, S.; Akula, M.; Mishra, B. In vitro antioxidant efficacy of EPS obtained from Micrococcus luteus snist-CM 02: A brief study. J. Microbiol. Biotechnol. Food Sci. 2017, 6, 1199. [Google Scholar] [CrossRef]
- Zhang, Y.; Hu, P.; Fan, Q.L.M. Study on Effect Elements of Exopolysaccharide Production of Lactobacillus kimchi SR8 and DPPH Radical Scavenging Activity. J. Food Nutr. Res. 2017, 5, 928–934. [Google Scholar] [CrossRef]
- Li, S.; Huang, R.; Shah, N.P.; Tao, X.; Xiong, Y.; Wei, H. Antioxidant and antibacterial activities of exopolysaccharides from Bifidobacterium bifidum WBIN03 and Lactobacillus plantarum R315. J. Dairy Sci. 2014, 97, 7334–7343. [Google Scholar] [CrossRef] [PubMed]
- Huang, Q.-L.; Siu, K.-C.; Wang, W.-Q.; Cheung, Y.-C.; Wu, J.-Y. Fractionation, characterization and antioxidant activity of exopolysaccharides from fermentation broth of a Cordyceps sinensis fungus. Process Biochem. 2013, 48, 380–386. [Google Scholar] [CrossRef]
- Dong, J.; Chi, Z.; Lu, S.; Xie, X.; Gong, P.; Li, H.; Liu, W. Bacterial exopolysaccharides: Characteristics and antioxidant mechanism. Int. J. Biol. Macromol. 2025, 289, 138849. [Google Scholar] [CrossRef] [PubMed]
- Pithva, S.; Shekh, S.; Dave, J.; Vyas, B.R.M. Probiotic attributes of autochthonous Lactobacillus rhamnosus strains of human origin. Appl. Biochem. Biotechnol. 2014, 173, 259–277. [Google Scholar] [CrossRef] [PubMed]
- Tsilia, V.; Van den Abbeele, P.; Van de Wiele, T. Improved in vitro assay for determining the mucin adherence of bacteria sensitive to Triton X-100 treatment. Folia Microbiol. 2015, 60, 435–442. [Google Scholar] [CrossRef] [PubMed]
- Zhao, D.; Jiang, J.; Liu, L.; Wang, S.; Ping, W.; Ge, J. Characterization of exopolysaccharides produced by Weissella confusa XG-3 and their potential biotechnological applications. Int. J. Biol. Macromol. 2021, 178, 306–315. [Google Scholar] [CrossRef] [PubMed]
- Fuhren, J.; Rösch, C.; Ten Napel, M.; Schols, H.A.; Kleerebezem, M. Synbiotic matchmaking in Lactobacillus plantarum: Substrate screening and gene-trait matching to characterize strain-specific carbohydrate utilization. Appl. Environ. Microbiol. 2020, 86, e01081-01020. [Google Scholar] [CrossRef] [PubMed]
- Shalini, R.; Abinaya, G.; Saranya, P.; Antony, U. Growth of selected probiotic bacterial strains with fructans from Nendran banana and garlic. LWT-Food Sci. Technol. 2017, 83, 68–78. [Google Scholar] [CrossRef]
- Stevenson, K.; McVey, A.F.; Clark, I.B.; Swain, P.S.; Pilizota, T. General calibration of microbial growth in microplate readers. Sci. Rep. 2016, 6, 38828. [Google Scholar] [CrossRef] [PubMed]
- Wacogne, B.; Belinger Podevin, M.; Vaccari, N.; Koubevi, C.; Codjiová, C.; Gutierrez, E.; Davoine, L.; Robert-Nicoud, M.; Rouleau, A.; Frelet-Barrand, A. Concentration vs. Optical Density of ESKAPEE Bacteria: A Method to Determine the Optimum Measurement Wavelength. Sensors 2024, 24, 8160. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Tian, J.; Tang, N.; Zhang, X.; Xiao, L.; Xu, M.; Dong, M.; Li, W. In vitro simulated digestion and fecal fermentation of exopolysaccharides from Lacticaseibacillus paracasei GL1. Food Funct. 2023, 14, 5120–5137. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Li, G.; Qin, W.; Cai, J.; Wang, N. Evaluation of in vitro simulated digestion and fermentation characteristics of the crude exopolysaccharide from Levilactobacillus brevis M-14. J. Food Sci. 2024, 89, 9860–9878. [Google Scholar] [CrossRef] [PubMed]
- Tintoré, M.; Cuñé, J.; Vu, L.D.; Poppe, J.; Van den Abbeele, P.; Baudot, A.; de Lecea, C. A Long-Chain Dextran Produced by Weissella cibaria Boosts the Diversity of Health-Related Gut Microbes Ex Vivo. Biology 2024, 13, 51. [Google Scholar] [CrossRef] [PubMed]
- Cinquin, C.; Le Blay, G.; Fliss, I.; Lacroix, C. Comparative effects of exopolysaccharides from lactic acid bacteria and fructo-oligosaccharides on infant gut microbiota tested in an in vitro colonic model with immobilized cells. FEMS Microbiol. Ecol. 2006, 57, 226–238. [Google Scholar] [CrossRef] [PubMed]
- Dal Bello, F.; Walter, J.; Hertel, C.; Hammes, W.P. In vitro study of prebiotic properties of levan-type exopolysaccharides from lactobacilli and non-digestible carbohydrates using denaturing gradient gel electrophoresis. Syst. Appl. Microbiol. 2001, 24, 232–237. [Google Scholar] [CrossRef] [PubMed]
- Olano-Martin, E.; Mountzouris, K.C.; Gibson, G.R.; Rastall, R.A. In vitro fermentability of dextran, oligodextran and maltodextrin by human gut bacteria. Br. J. Nutr. 2000, 83, 247–255. [Google Scholar] [CrossRef] [PubMed]
- Stuivenberg, G.A.; Burton, J.P.; Bron, P.A.; Reid, G. Why are bifidobacteria important for infants? Microorganisms 2022, 10, 278. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strains | Origin | ATCC/GenBank Accession Number | Source |
|---|---|---|---|
| Lacticaseibacillus rhamnosus GG | Human gut | ATCC 53103 | Dicofarm |
| Leuconostoc mesenteroides SJC113 | Cheese | MT742947 | IITAA Collection |
| Lactococcus lactis L3A21M1 | Cheese | KF193424 | IITAA Collection |
| Lacticaseibacillus rhamnosus FB1 | Human feces | OQ713701 | IITAA Collection |
| Lacticaseibacillus paracasei FB2 | Human feces | OQ713702 | IITAA Collection |
| Lacticaseibacillus paracasei FB3 | Human feces | OQ713703 | IITAA Collection |
| Lactobacillus gasseri BM8 | Human milk | OQ713687 | IITAA Collection |
| EPS * | 0.1 mg/mL | 1 mg/mL | 2 mg/mL | 3 mg/mL |
|---|---|---|---|---|
| Cholesterol-binding (%) | 33.01 ± 2.86 a | 40.18 ± 2.42 b | - | - |
| Radical scavenging activity—DPPH (%) | - | 10.94 ± 1.03 a | 13.15 ± 0.81 ab | 17.48 ± 1.74 b |
| Ascorbic acid (%) | - | 89.3 ± 4.8 | - | - |
| Eq. ascorbic acid (mg/mL) | - | 0.092 ± 0.017 a | 0.119 ± 0.011 ab | 0.164 ± 0.015 b |
| Hydroxyl scavenging activity (%) | - | 6.29 ± 1.46 a | 8.57 ± 2.29 a | 9.34 ± 2.06 a |
| Ascorbic acid (%) | - | 65.0 ± 1.2 | - | - |
| Eq. ascorbic acid (mg/mL) | - | 0.065 ± 0.024 a | 0.111 ± 0.039 a | 0.128 ± 0.033 a |
| Strain | Log CFU/mL | |
|---|---|---|
| 0 min * | 90 min | |
| Ln. mesenteroides SJC113 | 11.64 ± 0.07 | 8.62 ± 0.08 |
| L. rhamnosus GG | 10.63 ± 0.06 | 8.03 ± 0.25 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jurášková, D.; Ribeiro, S.C.; Silva, C.C.G. Prebiotic and Health-Promoting Benefits of Dextran-Type Exopolysaccharide Produced by Leuconostoc mesenteroides SJC113. Foods 2025, 14, 2635. https://doi.org/10.3390/foods14152635
Jurášková D, Ribeiro SC, Silva CCG. Prebiotic and Health-Promoting Benefits of Dextran-Type Exopolysaccharide Produced by Leuconostoc mesenteroides SJC113. Foods. 2025; 14(15):2635. https://doi.org/10.3390/foods14152635
Chicago/Turabian StyleJurášková, Dominika, Susana C. Ribeiro, and Célia C. G. Silva. 2025. "Prebiotic and Health-Promoting Benefits of Dextran-Type Exopolysaccharide Produced by Leuconostoc mesenteroides SJC113" Foods 14, no. 15: 2635. https://doi.org/10.3390/foods14152635
APA StyleJurášková, D., Ribeiro, S. C., & Silva, C. C. G. (2025). Prebiotic and Health-Promoting Benefits of Dextran-Type Exopolysaccharide Produced by Leuconostoc mesenteroides SJC113. Foods, 14(15), 2635. https://doi.org/10.3390/foods14152635