

Arthrospira platensis var. toliarensis: A Local Sustainable Microalga for Food System Resilience

 , , ,
, , ,  ,
,  and
and
Abstract
1. Introduction
2. Materials and Methods
2.1. Presentation of the Strain and Sampling
2.2. Experimental Cultivation Procedures
2.2.1. Experimental Design and Culture Conditions
2.2.2. Growth Monitoring and Biomass Estimation
2.3. Sample Collection and Biochemical Analyses
2.3.1. Sampling and Preparation
2.3.2. Physicochemical and Nutritional Composition
2.3.3. Amino Acid Analysis by HPLC-UV
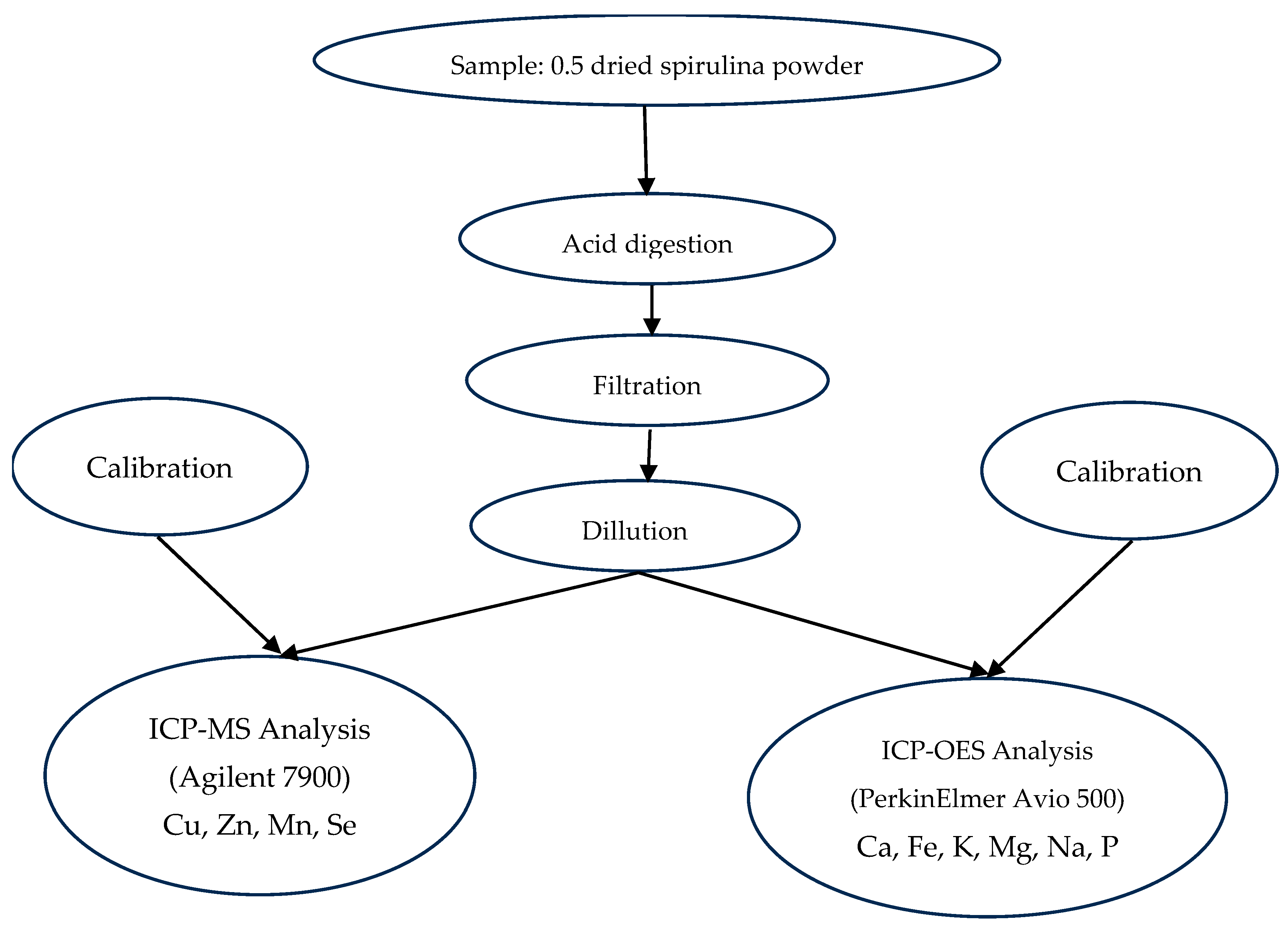
2.3.4. Mineral Element Quantification by ICP-OES and ICP-MS
2.3.5. Procedures for the Analysis of Bioactive Compounds
Extraction Procedure
Total Phenolic Content (TPC)
Total Flavonoid Content (TFC)
Antioxidant Activity (DPPH)
2.3.6. Toxicological Analysis: Aflatoxin Quantification (B1, B2, G1, and G2)
2.4. Statistical Analysis
3. Results
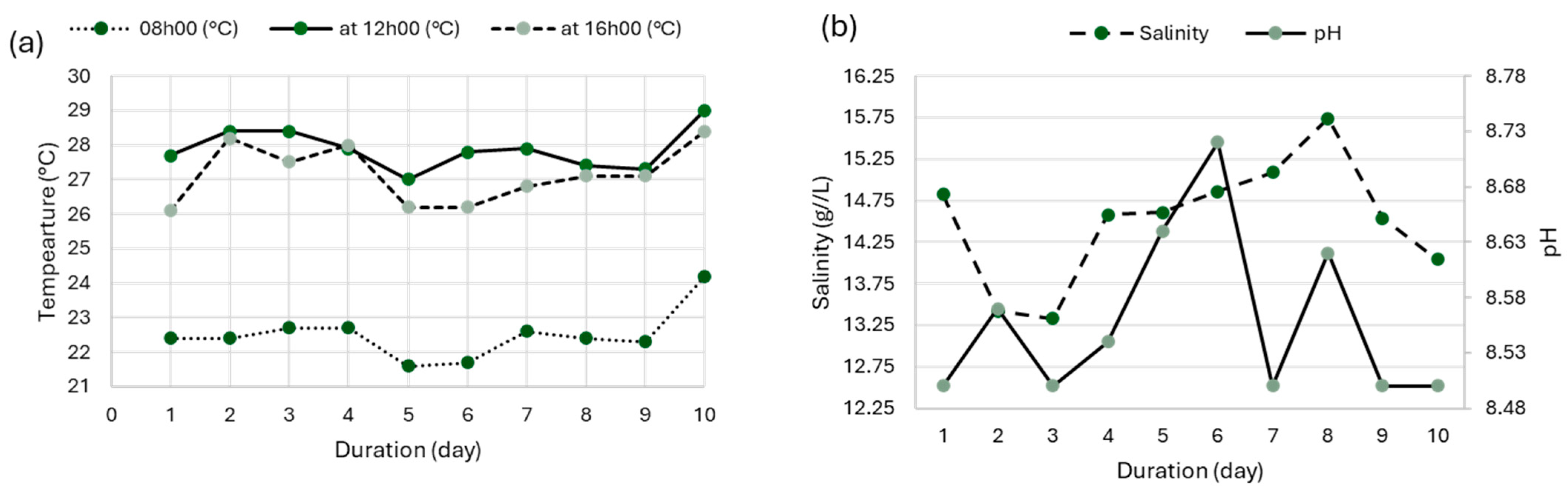
3.1. Experimental Cultivation
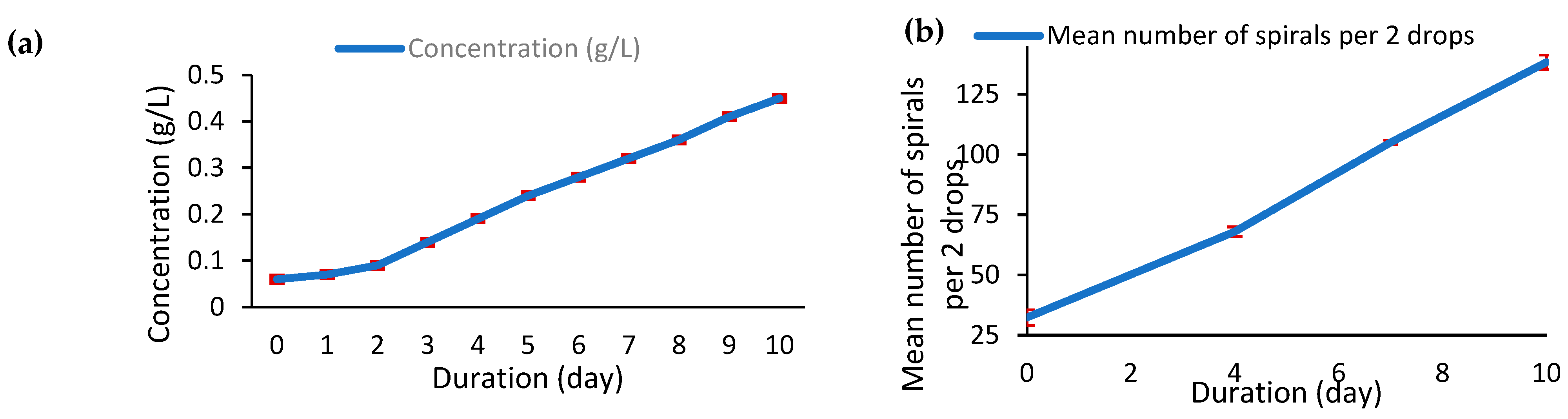
3.1.1. Growth Kinetics of Arthrospira platensis var. toliarensis
3.1.2. Biomass and Protein Yield
3.2. Proximate Composition of Arthrospira platensis var. toliarensis Powder
3.3. Micronutrient Composition: Amino Acid and Mineral Profiles
3.4. Analysis of Bioactive Compounds
3.4.1. Total Polyphenol Content in Dry Biomass
3.4.2. Polyphenols, Flavonoids, and Antioxidant Activity in Solvent Extracts
3.5. Toxicological Analysis: Aflatoxin Quantification
4. Discussion
4.1. Cultivation Performance of Arthrospira platensis var. toliarensis
4.2. Biochemical Composition and Nutritional Value
4.3. Phytochemical Richness and Antioxidant Activity
4.4. Toxicological Safety
4.5. Nutritional, Industrial, and Functional Perspectives: Arthrospira platensis var. toliarensis as a Strategic Food-Grade Super-Ingredient
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- FAO; IFAD; UNICEF; WFP; WHO. The State of Food Security and Nutrition in the World 2024—Financing to End Hunger, Food Insecurity and Malnutrition in All Its Forms; FAO: Rome, Italy, 2024; ISBN 978-92-5-138882-2. [Google Scholar]
- ONN. Plan National d’Action Multisectorielle Pour La Nutrition 2022–2026; Office National de Nutrition: Antananarivo, Madagascar, 2022; 240p.
- PAM. Plan Stratégique de Pays—Madagascar (2024–2028); Programme Alimentaire Mondial (PAM): Rome, Italy, 2024; p. 52. [Google Scholar]
- Li, X.; Siddique, K.H.M. (Eds.) Future Smart Food: Rediscovering Hidden Treasures of Neglected and Underutilized Species for Zero Hunger in Asia; Food and Agriculture Organization of the United Nations: Bangkok, Thailand, 2018; ISBN 978-92-5-130495-2. [Google Scholar]
- Stewart, B.A.; Lal, R. Increasing World Average Yields of Cereal Crops. In Advances in Agronomy; Elsevier: Amsterdam, The Netherlands, 2018; Volume 151, pp. 1–44. ISBN 978-0-12-815173-0. [Google Scholar]
- Henchion, M.; Hayes, M.; Mullen, A.; Fenelon, M.; Tiwari, B. Future Protein Supply and Demand: Strategies and Factors Influencing a Sustainable Equilibrium. Foods 2017, 6, 53. [Google Scholar] [CrossRef]
- Smith, K.; Watson, A.W.; Lonnie, M.; Peeters, W.M.; Oonincx, D.; Tsoutsoura, N.; Simon-Miquel, G.; Szepe, K.; Cochetel, N.; Pearson, A.G.; et al. Meeting the Global Protein Supply Requirements of a Growing and Ageing Population. Eur. J. Nutr. 2024, 63, 1425–1433. [Google Scholar] [CrossRef]
- Nicholson, C.C.; Emery, B.F.; Niles, M.T. Global Relationships between Crop Diversity and Nutritional Stability. Nat. Commun. 2021, 12, 5310. [Google Scholar] [CrossRef]
- Gavilan-Figari, I.M.; Peña-Urdániga, G.V.; Naka, A.; Castro-Rosas, M.A. The Role of Spirulina in Food Security: Nutritional and Ecological Implications in the Context of Climate Change. CABI Rev. 2024, 19, 1–10. [Google Scholar] [CrossRef]
- Ramos, S.D.P.; Bürck, M.; Costa, S.F.F.D.; Assis, M.; Braga, A.R.C. Spirulina as a Key Ingredient in the Evolution of Eco-Friendly Cosmetics. BioTech 2025, 14, 41. [Google Scholar] [CrossRef]
- Sharma, R.; Singh, G.P.; Sharma, V.K. Effects of Culture Conditions on Growth and Biochemical Profile of Chlorella vulgaris. J. Plant Pathol. Microb. 2012, 3, 1–6. [Google Scholar] [CrossRef]
- Fox, R.D. Spirulina: Production and Potential; Edisud: Aix-en-Provence, France, 1996; ISBN 978-2-85744-883-9. [Google Scholar]
- Ramilavonjy Ramiandrisoa, B.C.; Andrianatoandro, V.N.; Rasoanarivo, R.; Ranarijaona, H.L.T. Adaptation de Culture de Spiruline Arthrospira platensis Pour Une Meilleure Économie de l’ouest: Cas Du Centre Du Développement d’aquaculture de Mahajanga à Madagascar. Sci. Hum. Et Soc. 2023, 3, 1–18. [Google Scholar] [CrossRef]
- Ravelonandro, P.H.; Ratianarivo, D.H.; Joannis-Cassan, C.; Isambert, A.; Raherimandimby, M. Improvement of the Growth of Arthrospira (Spirulina) Platensis from Toliara (Madagascar): Effect of Agitation, Salinity and CO2 Addition. Food Bioprod. Process. 2011, 89, 209–216. [Google Scholar] [CrossRef]
- Jourdan, J.P. Cultivez Votre Spiruline: Manuel de Culture Artisanale; Antenna Technologies: Genève, Switzerland, 2012. [Google Scholar]
- Gilles, P.; Charito, F. La spiruline Pour Tous: Culture Familiale; Passerelle Éco: La Chapelle-sous-Uchon, France, 2014; ISBN 978-2-9533448-8-2. [Google Scholar]
- AOAC (Ed.) Official Methods of Analysis of AOAC International, 21st ed.; AOAC International: Gaithersburg, MD, USA, 2019; ISBN 978-0-935584-89-9. [Google Scholar]
- ISO 13903:2005; Animal Feeding Stuffs—Determination of Amino Acids Content. International Organization for Standardization (ISO): Geneva, Switzerland, 2005.
- MSZ EN 13805:2015; Foodstuffs—Determination of Trace Elements by Graphite Furnace Atomic Absorption Spectrometry (GFAAS). Hungarian Standards Institution (MSZ): Budapest, Hungary, 2015.
- EPA Method 6020B; Method 6020B (SW-846): Inductively Coupled Plasma—Mass Spectrometry. United States Environmental Protection Agency U.S. EPA: Washington, DC, USA, 2014.
- EPA Method 6010D; Method 6010D (SW-846): Inductively Coupled Plasma—Atomic Emission Spectrometry. United States Environmental Protection Agency U.S. EPA: Washington, DC, USA, 2018.
- Martí-Quijal, F.J.; Ramon-Mascarell, F.; Pallarés, N.; Ferrer, E.; Berrada, H.; Phimolsiripol, Y.; Barba, F.J. Extraction of Antioxidant Compounds and Pigments from Spirulina (Arthrospira platensis) Assisted by Pulsed Electric Fields and the Binary Mixture of Organic Solvents and Water. Appl. Sci. 2021, 11, 7629. [Google Scholar] [CrossRef]
- Balanescu, F.; Mihaila, M.; Cârâc, G.; Furdui, B.; Vînătoru, C.; Avramescu, S.; Lisa, E.; Cudalbeanu, M.; Dinica, R. Flavonoid Profiles of Two New Approved Romanian Ocimum Hybrids. Molecules 2020, 25, 4573. [Google Scholar] [CrossRef]
- Schincaglia, A.; Aspromonte, J.; Franchina, F.A.; Chenet, T.; Pasti, L.; Cavazzini, A.; Purcaro, G.; Beccaria, M. Current Developments of Analytical Methodologies for Aflatoxins’ Determination in Food during the Last Decade (2013–2022), with a Particular Focus on Nuts and Nut Products. Foods 2023, 12, 527. [Google Scholar] [CrossRef]
- Yu, L.; Ma, F.; Zhang, L.; Li, P. Determination of Aflatoxin B1 and B2 in Vegetable Oils Using Fe3O4/rGO Magnetic Solid Phase Extraction Coupled with High-Performance Liquid Chromatography Fluorescence with Post-Column Photochemical Derivatization. Toxins 2019, 11, 621. [Google Scholar] [CrossRef]
- FAO. FAOSTAT Statistical Database: Soybean Yield (Kg/Ha), 2023; Food and Agriculture Organization of the United Nations: Rome, Italy, 2023. [Google Scholar]
- Mekonnen, M.M.; Hoekstra, A.Y. The Green, Blue and Grey Water Footprint of Crops and Derived Crop Products. Hydrol. Earth Syst. Sci. 2011, 15, 1577–1600. [Google Scholar] [CrossRef]
- Singh, G. The Soybean: Botany, Production and Uses; CABI: Wallingford, CT, USA, 2010; ISBN 978-1-84593-644-0. [Google Scholar]
- Sobko, O.; Zikeli, S.; Claupein, W.; Gruber, S. Seed Yield, Seed Protein, Oil Content, and Agronomic Characteristics of Soybean (Glycine max L. Merrill) Depending on Different Seeding Systems and Cultivars in Germany. Agronomy 2020, 10, 1020. [Google Scholar] [CrossRef]
- Poore, J.; Nemecek, T. Reducing Food’s Environmental Impacts through Producers and Consumers. Science 2018, 360, 987–992. [Google Scholar] [CrossRef]
- Broom, D.M. Land and Water Usage in Beef Production Systems. Animals 2019, 9, 286. [Google Scholar] [CrossRef]
- Murphy, E.; Curran, T.P.; Holden, N.M.; O’Brien, D.; Upton, J. Water Footprinting of Pasture-Based Farms; Beef and Sheep. Animal 2018, 12, 1068–1076. [Google Scholar] [CrossRef]
- Delrue, F.; Alaux, E.; Moudjaoui, L.; Gaignard, C.; Fleury, G.; Perilhou, A.; Richaud, P.; Petitjean, M.; Sassi, J.-F. Optimization of Arthrospira Platensis (Spirulina) Growth: From Laboratory Scale to Pilot Scale. Fermentation 2017, 3, 59. [Google Scholar] [CrossRef]
- Du, X.; Zhu, C.; Hu, C.; Hao, H.; Xiao, Y.; Chi, Z. Techno-Economic Analysis of Spirulina Biomass Production in the Floating Photobioreactors for Fishmeal Replacements. J. Clean. Prod. 2025, 519, 146000. [Google Scholar] [CrossRef]
- Zhu, C.; Zhai, X.; Wang, J.; Han, D.; Li, Y.; Xi, Y.; Tang, Y.; Chi, Z. Large-Scale Cultivation of Spirulina in a Floating Horizontal Photobioreactor without Aeration or an Agitation Device. Appl. Microbiol. Biotechnol. 2018, 102, 8979–8987. [Google Scholar] [CrossRef]
- Soni, R.A.; Sudhakar, K.; Rana, R.S. Comparative Study on the Growth Performance of Spirulina Platensis on Modifying Culture Media. Energy Rep. 2019, 5, 327–336. [Google Scholar] [CrossRef]
- Vonshak, A. (Ed.) Spirulina Platensis Arthrospira: Physiology, Cell-Biology and Biotechnology; CRC Press: Boca Raton, FL, USA, 1997; ISBN 978-1-4822-7297-0. [Google Scholar]
- Podgórska-Kryszczuk, I. Spirulina an Invaluable Source of Macro- and Micronutrients with Broad Biological Activity and Application Potential. Molecules 2024, 29, 5387. [Google Scholar] [CrossRef]
- Reijnders, L.; Soret, S. Quantification of the Environmental Impact of Different Dietary Protein Choices. Am. J. Clin. Nutr. 2003, 78, 664S–668S. [Google Scholar] [CrossRef]
- Assaye, H.; Belay, A.; Desse, G.; Gray, D. Seasonal Variation in the Nutrient Profile of Arthrospira Fusiformis Biomass Harvested from an Ethiopian Soda Lake, Lake Chitu. J. Appl. Phycol. 2018, 30, 1597–1606. [Google Scholar] [CrossRef]
- Pimentel, J.D.S.M.; Nunes, Á.C.; Pereira, D.A.; Giani, A. Cyanobacteria Gene Expression in Response to Environmental Stress and Seasonal Changes. World J. Microbiol. Biotechnol. 2025, 41, 163. [Google Scholar] [CrossRef]
- Kharchuk, I.A.; Rylkova, O.A.; Beregovaya, N.M. State of Cyanobacteria Arthrospira Platensis and of Associated Microflora during Long-Term Storage in the State of Anhydrobiosis. Microbiology 2022, 91, 704–712. [Google Scholar] [CrossRef]
- Begum, N.; Qi, F.; Yang, F.; Khan, Q.U.; Faizan; Fu, Q.; Li, J.; Wang, X.; Wang, X.; Wang, J.; et al. Nutritional Composition and Functional Properties of A. Platensis-Derived Peptides: A Green and Sustainable Protein-Rich Supplement. Processes 2024, 12, 2608. [Google Scholar] [CrossRef]
- Habib, M.A.B. (Ed.) A Review on Culture, Production and Use of Spirulina as Food for Humans and Feeds for Domestic Animals and Fish; FAO Fisheries and Aquaculture Circular; Food and Agriculture Organization of the United Nations: Rome, Italy, 2008; ISBN 978-92-5-106106-0. [Google Scholar]
- Ogbonda, K.H.; Aminigo, R.E.; Abu, G.O. Influence of Temperature and pH on Biomass Production and Protein Biosynthesis in a Putative Spirulina sp. Bioresour. Technol. 2007, 98, 2207–2211. [Google Scholar] [CrossRef]
- Tian, X.; Lin, X.; Xie, Q.; Liu, J.; Luo, L. Effects of Temperature and Light on Microalgal Growth and Nutrient Removal in Turtle Aquaculture Wastewater. Biology 2024, 13, 901. [Google Scholar] [CrossRef]
- Akbarbaglu, Z.; Ayaseh, A.; Ghanbarzadeh, B.; Sarabandi, K. Biological Stabilization of Arthrospira Bioactive-Peptides within Biopolymers: Functional Food Formulation; Bitterness-Masking and Nutritional Aspects. LWT 2024, 191, 115653. [Google Scholar] [CrossRef]
- Bortolini, D.G.; Maciel, G.M.; Fernandes, I.D.A.A.; Pedro, A.C.; Rubio, F.T.V.; Branco, I.G.; Haminiuk, C.W.I. Functional Properties of Bioactive Compounds from Spirulina spp.: Current Status and Future Trends. Food Chem. Mol. Sci. 2022, 5, 100134. [Google Scholar] [CrossRef]
- Kudełka, W.; Kowalska, M.; Popis, M. Quality of Soybean Products in Terms of Essential Amino Acids Composition. Molecules 2021, 26, 5071. [Google Scholar] [CrossRef]
- Kumar, N.; Hong, S.; Zhu, Y.; Garay, A.; Yang, J.; Henderson, D.; Zhang, X.; Xu, Y.; Li, Y. Comprehensive Review of Chickpea (Cicer arietinum): Nutritional Significance, Health Benefits, Techno-functionalities, and Food Applications. Comp. Rev. Food Sci. Food Saf. 2025, 24, e70152. [Google Scholar] [CrossRef]
- Neag, E.; Stupar, Z.; Varaticeanu, C.; Senila, M.; Roman, C. Optimization of Lipid Extraction from Spirulina Spp. by Ultrasound Application and Mechanical Stirring Using the Taguchi Method of Experimental Design. Molecules 2022, 27, 6794. [Google Scholar] [CrossRef]
- Moreno-Garcia, L.; Gariépy, Y.; Barnabé, S.; Raghavan, G.S.V. Effect of Environmental Factors on the Biomass and Lipid Production of Microalgae Grown in Wastewaters. Algal Res. 2019, 41, 101521. [Google Scholar] [CrossRef]
- Hamad, G.M.; Abd El-Baky, N.; Sharaf, M.M.; Amara, A.A. Volatile Compounds, Fatty Acids Constituents, and Antimicrobial Activity of Cultured Spirulina (Arthrospira fusiformis) Isolated from Lake Mariout in Egypt. Sci. World J. 2023, 2023, 9919814. [Google Scholar] [CrossRef]
- Wang, Y.-Y.; Xu, B.-L.; Dong, C.-M.; Sun, Y.-Y. The Nutritional Value of Spirulina and Utilization Research. Life Res. 2023, 6, 15. [Google Scholar] [CrossRef]
- Panyakampol, J.; Cheevadhanarak, S.; Sutheeworapong, S.; Chaijaruwanich, J.; Senachak, J.; Siangdung, W.; Jeamton, W.; Tanticharoen, M.; Paithoonrangsarid, K. Physiological and Transcriptional Responses to High Temperature in Arthrospira (Spirulina) Platensis C1. Plant Cell Physiol. 2015, 56, 481–496. [Google Scholar] [CrossRef]
- Torzillo, G.; Vonshak, A. Effect of Light and Temperature on the Photosynthetic Activity of the Cyanobacterium Spirulina Platensis. Biomass Bioenergy 1994, 6, 399–403. [Google Scholar] [CrossRef]
- Lafarga, T.; Fernández-Sevilla, J.M.; González-López, C.; Acién-Fernández, F.G. Spirulina for the Food and Functional Food Industries. Food Res. Int. 2020, 137, 109356. [Google Scholar] [CrossRef]
- Priyanka, S.; Varsha, R.; Ayenampudi, S.B. Spirulina: A Spotlight on Its Nutraceutical Properties and Food Processing Applications. J. Microb. Biotechnol. Food Sci. 2023, 12, e4785. [Google Scholar] [CrossRef]
- Benjamin, Y.N.; Nazo Edith, K.-K.; Koné Lacina, G.; Kablan, T. Caractérisation De Quelques Légumes-Feuilles Les Plus Consommés Dans La Ville De Daloa (Centre-Ouest, Côte d’Ivoire). Eur. Sci. J. ESJ 2020, 16, 257. [Google Scholar] [CrossRef]
- Santos, D.L.S.; Amorim, M.S.; Silva, M.R.; Carvalho, A.L.D.; Rocha, M.E.; Silva, D.G.D. Determination and Evaluation of Minerals in Legumes Consumed in Vitória Da Conquista-BA. Open Access Libr. J. 2022, 9, 1–10. [Google Scholar] [CrossRef]
- Margier, M.; Georgé, S.; Hafnaoui, N.; Remond, D.; Nowicki, M.; Du Chaffaut, L.; Amiot, M.-J.; Reboul, E. Nutritional Composition and Bioactive Content of Legumes: Characterization of Pulses Frequently Consumed in France and Effect of the Cooking Method. Nutrients 2018, 10, 1668. [Google Scholar] [CrossRef]
- Islam, Z.; Islam, S.M.R.; Hossen, F.; Mahtab-ul-Islam, K.; Hasan, M.R.; Karim, R. Moringa Oleifera Is a Prominent Source of Nutrients with Potential Health Benefits. Int. J. Food Sci. 2021, 2021, 6627265. [Google Scholar] [CrossRef]
- Andrade, L.M. Chlorella and Spirulina Microalgae as Sources of Functional Foods, Nutraceuticals, and Food Supplements; an Overview. MOJ Food Process. Technol. 2018, 6, 00144. [Google Scholar] [CrossRef]
- Anvar, A.A.; Nowruzi, B. Bioactive Properties of Spirulina: A Review. Microb. Bioact. 2021, 4, 134–142. [Google Scholar] [CrossRef]
- Gaur, K.; Wal, A.; Sharma, P.; Parveen, A.; Singh, P.; Mishra, P.; Wal, P.; Mishra, N.T.P. Exploring the Nutritional and Medicinal Potential of Spirulina. Nat. Resour. Hum. Health 2024, 4, 277–286. [Google Scholar] [CrossRef]
- Castel, T.; Léon, K.; Gandubert, C.; Gueguen, B.; Amérand, A.; Guernec, A.; Théron, M.; Pichavant-Rafini, K. Comparison of Sodium Selenite and Selenium-Enriched Spirulina Supplementation Effects After Selenium Deficiency on Growth, Tissue Selenium Concentrations, Antioxidant Activities, and Selenoprotein Expression in Rats. Biol. Trace Elem. Res. 2024, 202, 685–700. [Google Scholar] [CrossRef]
- Jalouli, M.; Rahman, M.A.; Biswas, P.; Rahman, H.; Harrath, A.H.; Lee, I.-S.; Kang, S.; Choi, J.; Park, M.N.; Kim, B. Targeting Natural Antioxidant Polyphenols to Protect Neuroinflammation and Neurodegenerative Diseases: A Comprehensive Review. Front. Pharmacol. 2025, 16, 1492517. [Google Scholar] [CrossRef]
- Wang, M.; Wang, Y.; Zhang, H. Dietary Polyphenols for Tumor Therapy: Bioactivities, Nano-Therapeutic Systems and Delivery Strategies. Food Funct. 2025, 16, 853–866. [Google Scholar] [CrossRef]
- Rungjiraphirat, R.; Cheepchirasuk, N.; Suriyaprom, S.; Tragoolpua, Y. Inhibitory Efficacy of Arthrospira Platensis Extracts on Skin Pathogenic Bacteria and Skin Cancer Cells. Biology 2025, 14, 502. [Google Scholar] [CrossRef]
- Vicente-Zurdo, D.; Gómez-Mejía, E.; Morante-Zarcero, S.; Rosales-Conrado, N.; Sierra, I. Analytical Strategies for Green Extraction, Characterization, and Bioactive Evaluation of Polyphenols, Tocopherols, Carotenoids, and Fatty Acids in Agri-Food Bio-Residues. Molecules 2025, 30, 1326. [Google Scholar] [CrossRef]
- El-Chaghaby, G.A.; Rashad, S.; Abdel-Kader, S.F.; Rawash, E.-S.A.; Moneem, M.A. Assessment of Phytochemical Components, Proximate Composition and Antioxidant Properties of Scenedesmus Obliquus, Chlorella Vulgaris and Spirulina Platensis Algae Extracts. Egypt. J. Aquat. Biol. Fish. 2019, 23, 521–526. [Google Scholar] [CrossRef]
- Hidayati, J.R.; Yudiati, E.; Pringgenies, D.; Oktaviyanti, D.T.; Kusuma, A.P. Comparative Study on Antioxidant Activities, Total Phenolic Compound and Pigment Contents of Tropical Spirulina Platensis, Gracilaria Arcuata and Ulva Lactuca Extracted in Different Solvents Polarity. E3S Web Conf. 2020, 147, 03012. [Google Scholar] [CrossRef]
- Miazek, K.; Beton, K.; Śliwińska, A.; Brożek-Płuska, B. The Effect of β-Carotene, Tocopherols and Ascorbic Acid as Anti-Oxidant Molecules on Human and Animal In Vitro/In Vivo Studies: A Review of Research Design and Analytical Techniques Used. Biomolecules 2022, 12, 1087. [Google Scholar] [CrossRef]
- Park, W.; Kim, H.-J.; Li, M.; Lim, D.; Kim, J.; Kwak, S.-S.; Kang, C.-M.; Ferruzzi, M.; Ahn, M.-J. Two Classes of Pigments, Carotenoids and C-Phycocyanin, in Spirulina Powder and Their Antioxidant Activities. Molecules 2018, 23, 2065. [Google Scholar] [CrossRef]
- Salvia-Trujillo, L.; Verkempinck, S.H.E.; Sun, L.; Van Loey, A.M.; Grauwet, T.; Hendrickx, M.E. Lipid Digestion, Micelle Formation and Carotenoid Bioaccessibility Kinetics: Influence of Emulsion Droplet Size. Food Chem. 2017, 229, 653–662. [Google Scholar] [CrossRef]
- Demarco, M.; Oliveira De Moraes, J.; Matos, Â.P.; Derner, R.B.; De Farias Neves, F.; Tribuzi, G. Digestibility, Bioaccessibility and Bioactivity of Compounds from Algae. Trends Food Sci. Technol. 2022, 121, 114–128. [Google Scholar] [CrossRef]
- European Commission. European Commission Regulation (EU): 2023/915 of the Commission of 25 April 2023 on Maximum Levels for Certain Contaminants in Food and Repealing Regulation (EC) No 1881/2006; European Commission: Belgium, Brussels, 2023; pp. 103–157. [Google Scholar]
- Chilaka, C.A.; Obidiegwu, J.E.; Chilaka, A.C.; Atanda, O.O.; Mally, A. Mycotoxin Regulatory Status in Africa: A Decade of Weak Institutional Efforts. Toxins 2022, 14, 442. [Google Scholar] [CrossRef]
- Alshannaq, A.; Yu, J.-H. Occurrence, Toxicity, and Analysis of Major Mycotoxins in Food. Int. J. Environ. Res. Public Health 2017, 14, 632. [Google Scholar] [CrossRef]
- Mannaa, M.; Kim, K.D. Influence of Temperature and Water Activity on Deleterious Fungi and Mycotoxin Production during Grain Storage. Mycobiology 2017, 45, 240–254. [Google Scholar] [CrossRef]
- Altyar, A.E.; Kensara, O.A.; Noreldin, A.E.; Albadrani, G.M.; El-Demerdash, F.M.; Sayed, A.A.; Piscopo, M.; Mohammedsaleh, Z.M.; Al-Ghadi, M.Q.; Ghaboura, N.; et al. Spirulina Platensis Ameliorates Hepatic Oxidative Stress and DNA Damage Induced by Aflatoxin B1 in Rats. Toxicon 2024, 237, 107553. [Google Scholar] [CrossRef]
- Johnson, M.; Gilbert, K.; Peter, M.; Gordon, N.; Charles, M. Spirulina Platensis Inhibits Aflatoxin B1 Induced Biochemical Changes in Male Swiss Albino Mice. J. Phytopharmacol. 2022, 11, 341–346. [Google Scholar] [CrossRef]
- Petrus, M.; Culerrier, R.; Campistron, M.; Barre, A.; Rougé, P. First Case Report of Anaphylaxis to Spirulin: Identification of Phycocyanin as Responsible Allergen. Allergy 2010, 65, 924–925. [Google Scholar] [CrossRef]
- Gromek, W.; Kołdej, N.; Kurowski, M.; Majsiak, E. Spirulina (Arthrospira platensis): Antiallergic Agent or Hidden Allergen? A Literature Review. Foods 2024, 13, 1052. [Google Scholar] [CrossRef]
- Pinchart, P.-E.; Leruste, A.; Pasqualini, V.; Mastroleo, F. Microcystins and Cyanobacterial Contaminants in the French Small-Scale Productions of Spirulina (Limnospira sp.). Toxins 2023, 15, 354. [Google Scholar] [CrossRef]
- Sativa, H.A.; Faridah, D.N.; Andarwulan, N. Methods Validation and Analysis of Spirulina (Arthrospira platensis) Microcystin Contaminants by Enzyme-Linked Immunosorbent Assay (ELISA). Braz. J. Food Technol. 2024, 27, e2024005. [Google Scholar] [CrossRef]
- Al-Dhabi, N.A. Heavy Metal Analysis in Commercial Spirulina Products for Human Consumption. Saudi J. Biol. Sci. 2013, 20, 383–388. [Google Scholar] [CrossRef]
- Masten Rutar, J.; Jagodic Hudobivnik, M.; Nečemer, M.; Vogel Mikuš, K.; Arčon, I.; Ogrinc, N. Nutritional Quality and Safety of the Spirulina Dietary Supplements Sold on the Slovenian Market. Foods 2022, 11, 849. [Google Scholar] [CrossRef]
- Gogna, S.; Kaur, J.; Sharma, K.; Prasad, R.; Singh, J.; Bhadariya, V.; Kumar, P.; Jarial, S. Spirulina—An Edible Cyanobacterium with Potential Therapeutic Health Benefits and Toxicological Consequences. J. Am. Nutr. Assoc. 2023, 42, 559–572. [Google Scholar] [CrossRef]
- Bernaerts, T.M.M.; Gheysen, L.; Foubert, I.; Hendrickx, M.E.; Van Loey, A.M. The Potential of Microalgae and Their Biopolymers as Structuring Ingredients in Food: A Review. Biotechnol. Adv. 2019, 37, 107419. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Protein Source | Daily Productivity (g/m2/day) | Protein Yield (g/m2/day) | Production Duration (days) | References |
|---|---|---|---|---|
| Spirulina (this study) | 7.8 ± 0.58 | 4.68 ± 0.35 | 10 (up to 300/year) | Present study |
| Soybean (grain) | 33–421 (seasonal) | 12–170 (annual) | 90–150 | [26,27,28,29] |
| Beef (ruminant meat) | 0.37–11.9 (extensive systems) | ~0.00074 | 600–900 | [30,31,32] |
| Macronutrient | Cool Season (May) | Hot Season (October) |
|---|---|---|
| Moisture (%) | 7.2 ± 0.11 ᵃ | 6.9 ± 0.20 ᵃ |
| Ash (%) | 9.15 ± 0.15 ᵃ | 9.32 ± 0.15 ᵃ |
| Crude Protein (%) | 44.61 ± 0.26 ᵇ | 65.12 ± 0.62 ᵃ |
| Lipids (%) | 1.05 ± 0.39 ᵃ | 1.75 ± 0.37 ᵃ |
| Total Sugars (%) | 37.98 ± 0.73 ᵃ | 16.90 ± 1.19 ᵇ |
| Component | Cool Season (May) | Hot Season (October) |
|---|---|---|
| Amino Acids (g/100 g of (DM)) | ||
| Alanine | 3.41 ± 0.05 ᵃ | 3.28 ± 0.06 ᵇ |
| Arginine | 2.72 ± 0.04 ᵇ | 3.91 ± 0.08 ᵃ |
| Aspartic acid | 4.32 ± 0.06 ᵇ | 5.87 ± 0.13 ᵃ |
| Glutamic acid | 6.19 ± 0.09 ᵇ | 9.23 ± 0.16 ᵃ |
| Glycine | 2.13 ± 0.03 ᵃ | 1.92 ± 0.04 ᵇ |
| Histidine | 0.646 ± 0.012 ᵇ | 0.985 ± 0.022 ᵃ |
| Hydroxyproline | <0.2 (LOQ) ᵇ | <0.2 (LOQ) ᵇ |
| Isoleucine | 2.40 ± 0.03 ᵇ | 3.66 ± 0.09 ᵃ |
| Leucine | 3.81 ± 0.05 ᵇ | 6.10 ± 0.14 ᵃ |
| Lysine | 1.82 ± 0.03 ᵃ | 1.75 ± 0.04 ᵃ |
| Ornithine | <0.05 (LOQ) ᵇ | <0.05 (LOQ) ᵇ |
| Phenylalanine | 1.89 ± 0.03 ᵇ | 2.14 ± 0.05 ᵃ |
| Proline | 1.68 ± 0.03 ᵇ | 1.71 ± 0.08 ᵇ |
| Serine | 2.08 ± 0.03 ᵃ | 2.01 ± 0.06 ᵃ |
| Threonine | 2.12 ± 0.03 ᵃ | 2.34 ± 0.12 ᵃ |
| Tyrosine | 1.73 ± 0.02 ᵇ | 2.90 ± 0.10 ᵃ |
| Valine | 2.84 ± 0.04 ᵇ | 4.05 ± 0.13 ᵃ |
| Cystine + Cysteine | 0.399 ± 0.008 ᵇ | 0.376 ± 0.015 ᵇ |
| Methionine | 0.855 ± 0.014 ᵇ | 0.898 ± 0.020 ᵃ |
| Minerals (mg/100 g of DM) | ||
| Sodium (Na) | 2140.0 ± 35.4 ᵇ | 687.2 ± 12.3 ᵃ |
| Potassium (K) | 1530.0 ± 21.8 ᵇ | 1228.35 ± 10.4 ᵃ |
| Calcium (Ca) | 580.0 ± 12.1 ᵃ | 968.0 ± 15.1 ᵇ |
| Magnesium (Mg) | 187.0 ± 8.2 ᵃ | 389.28 ± 6.4 ᵇ |
| Phosphorus (P) | 690.0 ± 14.7 ᵃ | 815.0 ± 13.2 ᵇ |
| Iron (Fe) | 235.0 ± 9.1 ᵇ | 65.74 ± 1.1 ᵃ |
| Zinc (Zn) | 12.4 ± 1.3 ᵇ | 2.5 ± 0.4 ᵃ |
| Manganese (Mn) | 1.7 ± 0.2 ᵃ | 3.26 ± 0.6 ᵇ |
| Copper (Cu) | 1.4 ± 0.1 ᵇ | 0.5 ± 0.1 ᵃ |
| Selenium (Se) | 0.06 ± 0.01 ᵃ | 0.08 ± 0.01 ᵇ |
| Standard Equivalent | Total Polyphenols (mg/kg of DM) |
|---|---|
| Gallic acid (GAE) | 2830 ± 0.03 |
| Tannic acid | 3410 ± 0.11 |
| EGCG (epigallocatechin gallate) | 4110 ± 0.03 |
| Epicatechin | 1720 ± 0.05 |
| Catechin | 2260 ± 0.04 |
| Extraction Solvent | TPC (g GAE/100 g of DE) | TFC (g QE/100 g of DE) | DPPH IC50 (µg/mL) |
|---|---|---|---|
| Hydroethanolic extract | 4.67 ± 0.062 | 2.05 ± 0.036 | 213.77 ± 2.15 |
| Hexanic extract | 1.61 ± 0.074 | 1.405 ± 0.036 | 101.03 ± 1.37 |
| Aqueous extract | 3.77 ± 0.027 | 1.65 ± 0.08 | – |
| Ascorbic acid | 1.87 ± 0.04 |
| Aflatoxin | Cool Season (May) | Hot Season (October) |
|---|---|---|
| Aflatoxin G1 (µg/kg) | 0.1 | 0.1 |
| Aflatoxin B1 (µg/kg) | 0.1 | 0.2 |
| Aflatoxin G2 (µg/kg) | <0.1 | <0.1 |
| Aflatoxin B2 (µg/kg) | <0.1 | <0.1 |
| Total aflatoxins | 0.2 | 0.3 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Telesphore, A.F.; Botezatu, A.V.; Istrati, D.I.; Furdui, B.; Dinica, R.M.; Razafindratovo, V.L.A. Arthrospira platensis var. toliarensis: A Local Sustainable Microalga for Food System Resilience. Foods 2025, 14, 2634. https://doi.org/10.3390/foods14152634
Telesphore AF, Botezatu AV, Istrati DI, Furdui B, Dinica RM, Razafindratovo VLA. Arthrospira platensis var. toliarensis: A Local Sustainable Microalga for Food System Resilience. Foods. 2025; 14(15):2634. https://doi.org/10.3390/foods14152634
Chicago/Turabian StyleTelesphore, Antonio Fidinirina, Andreea Veronica Botezatu, Daniela Ionela Istrati, Bianca Furdui, Rodica Mihaela Dinica, and Valérie Lalao Andriamanamisata Razafindratovo. 2025. "Arthrospira platensis var. toliarensis: A Local Sustainable Microalga for Food System Resilience" Foods 14, no. 15: 2634. https://doi.org/10.3390/foods14152634
APA StyleTelesphore, A. F., Botezatu, A. V., Istrati, D. I., Furdui, B., Dinica, R. M., & Razafindratovo, V. L. A. (2025). Arthrospira platensis var. toliarensis: A Local Sustainable Microalga for Food System Resilience. Foods, 14(15), 2634. https://doi.org/10.3390/foods14152634