
Fermentation to Increase the Value of Roasted Coffee Silverskin as a Functional Food Ingredient




Abstract
1. Introduction
2. Materials and Methods
2.1. Materials
2.1.1. Roasted Silver Skin of Coffee Beans
2.1.2. Microorganisms
2.2. Methods
2.2.1. Food Safety of RCSS
2.2.2. Fermentation
2.2.3. Number of Viable Microbial Cells in RCSS During Fermentation
2.2.4. Measurements of pH
2.2.5. Overall Antioxidant Capacity and Total Phenolic Compounds
2.2.6. Prebiotic Properties
2.2.7. Statistical Analysis
3. Results and Discussion
3.1. Food Safety of RCSS
3.2. Changes in pH Values Due to RCSS Fermentation
3.3. RCSS as a Medium for the Microorganisms’ Growth
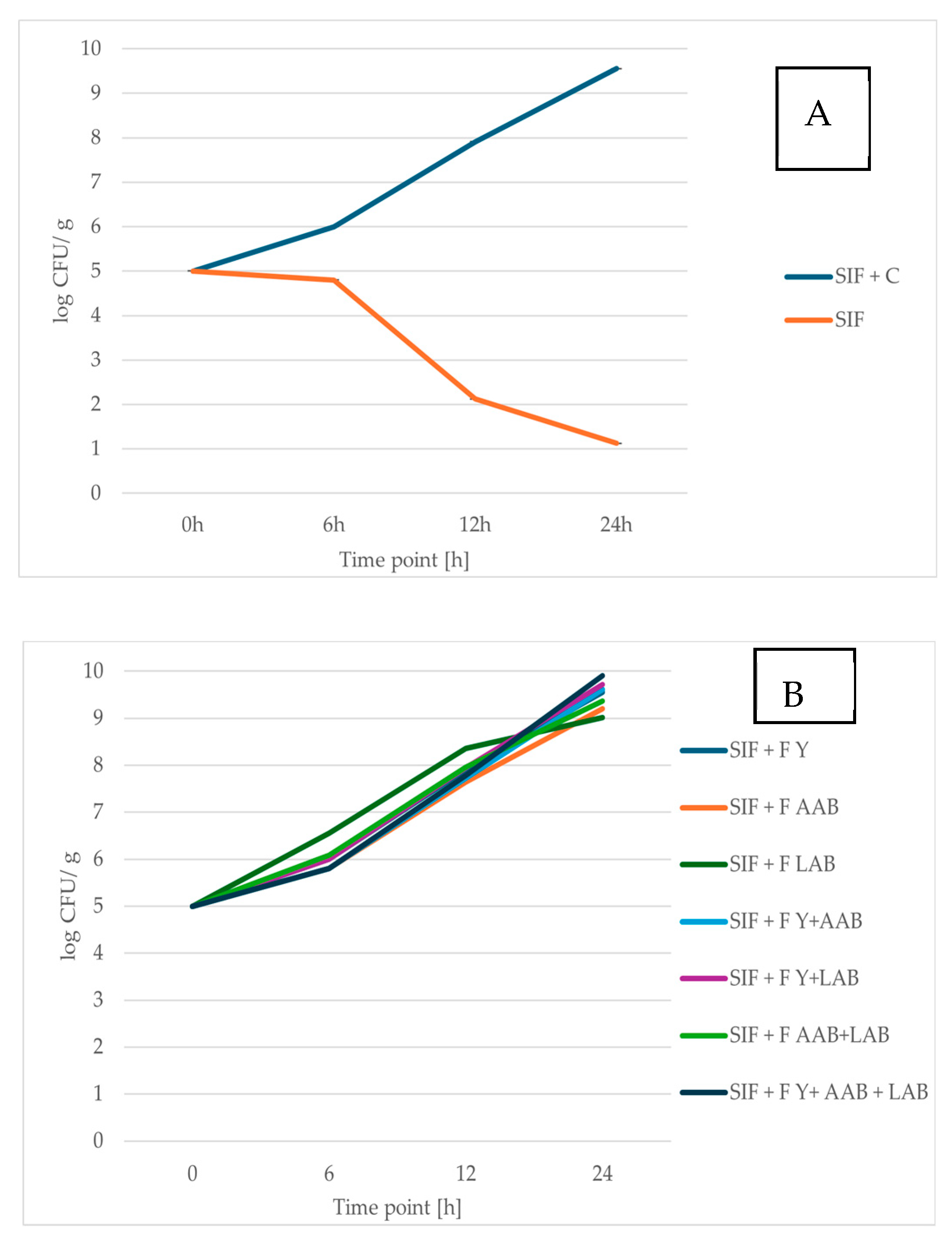
3.3.1. Growth of Microorganisms During RCSS Fermentation
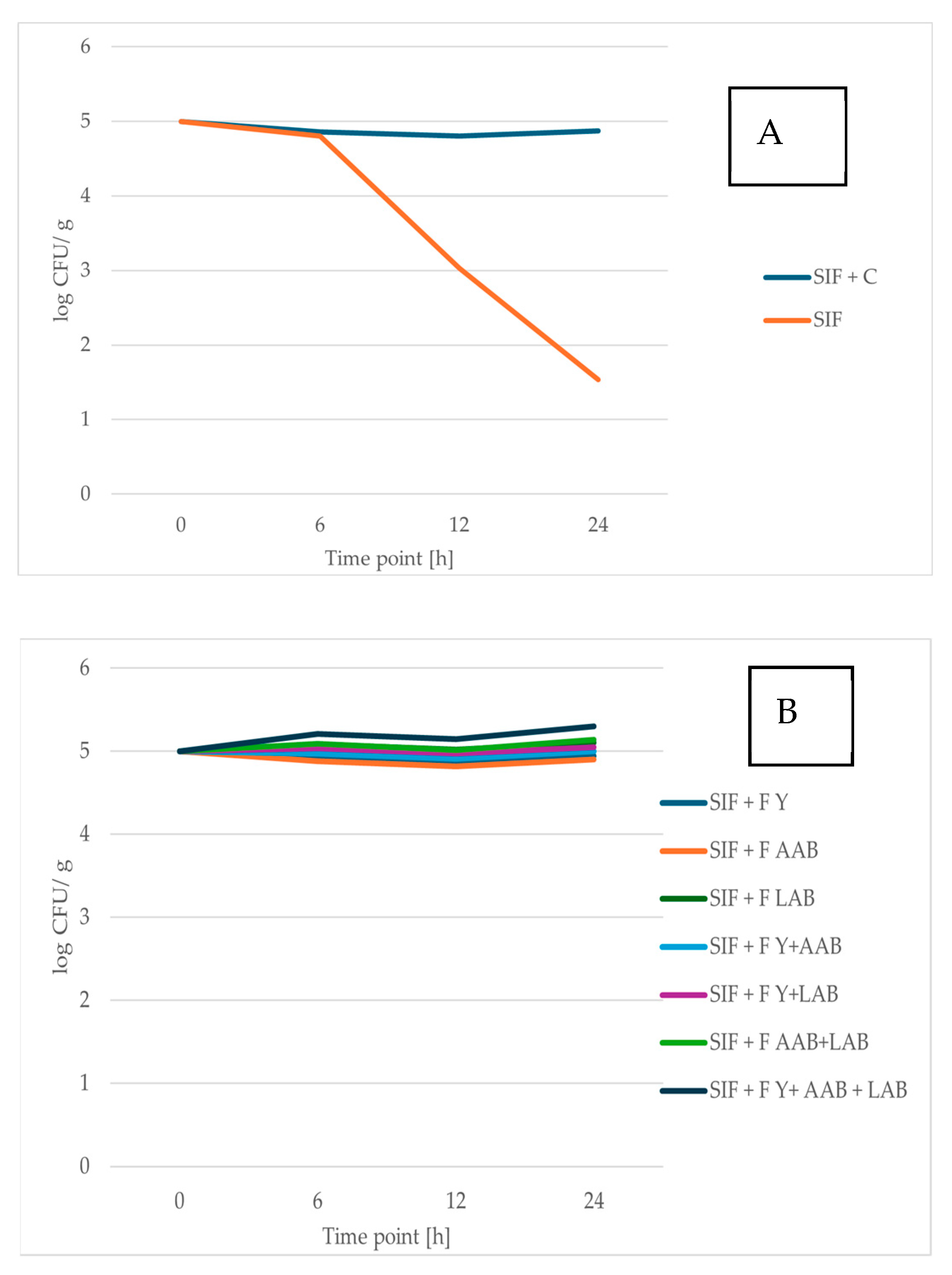
3.3.2. Survival of Probiotic Microorganisms Exposed to SIF Containing RCSS
3.4. Total Phenolic Compounds and Overall Antioxidant Capacity
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| RCSS | Roasted Coffee Silver Skin |
| CSS | Coffee Silver Skin |
| SCOBY | Symbiotic Cultures of Bacteria and Yeast |
| CL | Clean Label |
| ROS | Reactive Oxygen Species |
| Y | The yeast S. ludwigii WSL/A3 |
| AAB | The acetic acid bacteria G. oxydans ŁOCK 1153 |
| LAB | The lactic acid bacteria L. brevis ŁOCK 1152 |
| YPG | Yeast Peptone Glucose Medium |
| GFYP | Glucose-Free Yeast Peptone Medium |
| MRS | De Man, Rogosa, and Sharpe Medium |
| PCA | Plate Count Agar |
| VRBGA | Violet Red Bile Glucose Agar Medium |
| SDA | Sabouraud Dextrose Agar Medium |
| OSA | Orange Serum Agar |
| CFU | Colony-Forming Units |
| GAE | Gallic Acid |
| SIF | Simulated Intestinal Fluid |
| FC | Folin–Ciocalteu |
| FBBB | Fast Blue BB |
References
- Galanakis, C.M. Food Waste Recovery: Prospects and Opportunities. In Sustainable Food Systems from Agriculture to Industry; Elsevier: Amsterdam, The Netherlands, 2018; pp. 401–419. [Google Scholar]
- Iriondo-DeHond, M.; Miguel, E.; del Castillo, M.D. Food byproducts as sustainable ingredients for innovative and healthy dairy foods. Nutrients 2018, 10, 1358. [Google Scholar] [CrossRef] [PubMed]
- Sloan, A.E. Food expo highlights clean labels and functional foods. Food Technol. 2011, 65, 48–51. [Google Scholar]
- Chen, A.; Kayrala, N.; Trapeau, M.; Aoun, M.; Bordenave, N. The clean label trend: An ineffective heuristic that disserves both consumers and the food industry? Compr. Rev. Food Sci. Food Saf. 2022, 21, 4921–4938. [Google Scholar] [CrossRef] [PubMed]
- Šálková, D.; Maierová, O.; Kvasničková Stanislavská, L.; Pilař, L. The relationship between “zero waste” and food: Insights from social media trends. Foods 2023, 12, 3280. [Google Scholar] [CrossRef] [PubMed]
- Guiné, R.P.; Florença, S.G.; Barroca, M.J.; Anjos, O. The link between the consumer and the innovations in food product development. Foods 2020, 9, 1317. [Google Scholar] [CrossRef] [PubMed]
- Skorupa, A.; Worwąg, M.; Kowalczyk, M. Coffee industry and ways of using by-products as bioadsorbents for removal of pollutants. Water 2022, 15, 112. [Google Scholar] [CrossRef]
- Bevilacqua, E.; Cruzat, V.; Singh, I.; Rose’Meyer, R.B.; Panchal, S.K.; Brown, L. The potential of spent coffee grounds in functional food development. Nutrients 2023, 15, 994. [Google Scholar] [CrossRef] [PubMed]
- Wibowo, N.A.; Wanita, Y.P.; Novitasari, E.; Amri, A.F.; Purwanto, E.H.; Yulianti, Y.; Aurum, F.S. Innovative of cascara as potential in beverage, food and their functional impact: A review. Int. J. Food Sci. Technol. 2024, 59, 8082–8092. [Google Scholar] [CrossRef]
- Bessada, S.M.; Alves, R.C.; Oliveira, M.B.P.P. Coffee silverskin: A review on potential cosmetic applications. Cosmetics 2018, 5, 5. [Google Scholar] [CrossRef]
- Salim, H.; Ahmed, C.S.; Nassima, M.S.; Fadia, B.S.; Mimoun, S.; Merzouk, H. Coffee silverskin waste as alternative to produce alkaloids with antioxidant, antihemolytic, and alpha-amylase inhibitory activities. Biomass Convers. Biorefin. 2023, 15, 1069–1082. [Google Scholar] [CrossRef]
- Ballesteros, L.F.; Teixeira, J.A.; Mussatto, S.I. Chemical, functional, and structural properties of spent coffee grounds and coffee silverskin. Food Bioprocess Technol. 2014, 7, 3493–3503. [Google Scholar] [CrossRef]
- Narita, Y.; Inouye, K. Review on utilization and composition of coffee silverskin. Food Res. Int. 2014, 61, 16–22. [Google Scholar] [CrossRef]
- Machado, F.; Gómez-Domínguez, I.; Hurtado-Ribeira, R.; Martin, D.; Coimbra, M.A.; Del Castillo, M.D.; Coreta-Gomes, F. In vitro human colonic fermentation of coffee arabinogalactan and melanoidin-rich fractions. Int. J. Biol. Macromol. 2024, 275, 133740. [Google Scholar] [CrossRef] [PubMed]
- Vinelli, V.; Biscotti, P.; Martini, D.; Del Bo’, C.; Marino, M.; Meroño, T.; Riso, P. Effects of Dietary Fibers on Short-Chain Fatty Acids and Gut Microbiota Composition in Healthy Adults: A Systematic Review. Nutrients 2022, 14, 2559. [Google Scholar] [CrossRef] [PubMed]
- Belmiro, R.H.; de Carvalho Oliveira, L.; Tribst, A.A.L.; Cristianini, M. Techno-Functional Properties of Coffee By-Products Are Modified by Dynamic High Pressure: A Case Study of Clean Label Ingredient in Cookies. LWT 2022, 154, 112601. [Google Scholar] [CrossRef]
- Machado, M.; Ferreira, H.; Oliveira, M.B.P.; Alves, R.C. Coffee By-Products: An Underexplored Source of Prebiotic Ingredients. Crit. Rev. Food Sci. Nutr. 2024, 64, 7181–7200. [Google Scholar] [CrossRef] [PubMed]
- Martuscelli, M.; Esposito, L.; Di Mattia, C.D.; Ricci, A.; Mastrocola, D. Characterization of coffee silver skin as potential food-safe ingredient. Foods 2021, 10, 1367. [Google Scholar] [CrossRef] [PubMed]
- Gocmen, D.; Sahan, Y.; Yildiz, E.; Coskun, M.; Aroufai, I.A. Use of coffee silverskin to improve the functional properties of cookies. J. Food Sci. Technol. 2019, 56, 2979–2988. [Google Scholar] [CrossRef] [PubMed]
- Bertolino, M.; Barbosa-Pereira, L.; Ghirardello, D.; Botta, C.; Rolle, L.; Guglielmetti, A.; Zeppa, G. Coffee silverskin as nutraceutical ingredient in yogurt: Its effect on functional properties and its bioaccessibility. J. Sci. Food Agric. 2019, 99, 4267–4275. [Google Scholar] [CrossRef] [PubMed]
- Martuscelli, M.; Esposito, L.; Mastrocola, D. The role of coffee silver skin against oxidative phenomena in newly formulated chicken meat burgers after cooking. Foods 2021, 10, 1833. [Google Scholar] [CrossRef] [PubMed]
- European Commission. Novel Food Catalogue. 2024. Available online: https://ec.europa.eu/food/food-feed-portal/screen/novel-food-catalogue/search (accessed on 23 April 2025).
- Ortiz-Sanchez, M.; Inocencio-García, P.J.; Alzate-Ramírez, A.F.; Alzate, C.A.C. Potential and restrictions of food-waste valorization through fermentation processes. Fermentation 2023, 9, 274. [Google Scholar] [CrossRef]
- Sadh, P.K.; Kumar, S.; Chawla, P.; Duhan, J.S. Fermentation: A boon for production of bioactive compounds by processing of food industries wastes (by-products). Molecules 2018, 23, 2560. [Google Scholar] [CrossRef] [PubMed]
- Tran, T.; Roullier-Gall, C.; Verdier, F.; Martin, A.; Schmitt-Kopplin, P.; Alexandre, H.; Tourdot-Maréchal, R. Microbial interactions in kombucha through the lens of metabolomics. Metabolites 2022, 12, 235. [Google Scholar] [CrossRef] [PubMed]
- Ditrych, M.; Jędrasik, J.; Królak, K.; Guzińska, N.; Pielech-Przybylska, K.; Ścieszka, S.; Kordialik-Bogacka, E. Kombucha Fortified with Cascade Hops (Humulus lupulus L.): Enhanced Antioxidative and Sensory Properties. Appl. Microbiol. Biotechnol. 2025, 109, 27. [Google Scholar] [CrossRef] [PubMed]
- Sales, A.L.; Iriondo-DeHond, A.; DePaula, J.; Ribeiro, M.; Ferreira, I.M.; Miguel, M.A.L.; Farah, A. Intra-cellular antioxidant and anti-inflammatory effects and bioactive profiles of coffee cascara and black tea kombucha beverages. Foods 2023, 12, 1905. [Google Scholar] [CrossRef] [PubMed]
- Candra, A.; Ahmed, Y.W.; Kitaw, S.L.; Anley, B.E.; Chen, K.J.; Tsai, H.C. A green method for fabrication of a biocompatible gold-decorated-bacterial cellulose nanocomposite in spent coffee grounds kombucha: A sustainable approach for augmented wound healing. J. Drug Deliv. Sci. Technol. 2024, 94, 105477. [Google Scholar] [CrossRef]
- Callejo, M.J.; García Navas, J.J.; Alba, R.; Escott, C.; Loira, I.; González, M.C.; Morata, A. Wort fermentation and beer conditioning with selected non-Saccharomyces yeasts in craft beers. Eur. Food Res. Technol. 2019, 245, 1229–1238. [Google Scholar] [CrossRef]
- Silva de Souza, H.L.; de Almeida, D.P.; Dos Santos, A.S.; Ramos, C.L. Physiological and fermentative performance of non-Saccharomyces yeasts isolated from kombucha for beer production. Food Sci. Technol. Int. 2025, 10820132251322288. [Google Scholar] [CrossRef] [PubMed]
- Vejarano, R. Saccharomycodes ludwigii, control and potential uses in winemaking processes. Fermentation 2018, 4, 71. [Google Scholar] [CrossRef]
- Pilap, W.; Thanonkeo, S.; Klanrit, P.; Thanonkeo, P. The potential of multistress tolerant yeast, Saccharomycodes ludwigii, for second-generation bioethanol production. Sci. Rep. 2022, 12, 22062. [Google Scholar] [CrossRef] [PubMed]
- Bovo, B.; Carlot, M.; Lombardi, A.; Lomolino, G.; Lante, A.; Giacomini, A.; Corich, V. Exploring the use of Saccharomyces cerevisiae commercial strain and Saccharomycodes ludwigii natural isolate for grape marc fermentation to improve sensory properties of spirits. Food Microbiol. 2014, 41, 33–41. [Google Scholar] [CrossRef] [PubMed]
- Neffe-Skocińska, K.; Długosz, E.; Szulc-Dąbrowska, L.; Zielińska, D. Novel Gluconobacter oxydans strains selected from Kombucha with potential postbiotic activity. Appl. Microbiol. Biotechnol. 2024, 108, 27. [Google Scholar] [CrossRef] [PubMed]
- Tenea, G.N.; Cifuentes, V.; Reyes, P.; Cevallos-Vallejos, M. Unveiling the microbial signatures of Arabica coffee cherries: Insights into ripeness specific diversity, functional traits, and implications for quality and safety. Foods 2025, 14, 614. [Google Scholar] [CrossRef] [PubMed]
- Ali, S.S.; Ahsan, H.; Zia, M.K.; Siddiqui, T.; Khan, F.H. Understanding oxidants and antioxidants: Classical team with new players. J. Food Biochem. 2020, 44, e13145. [Google Scholar] [CrossRef] [PubMed]
- Lee, B.J.; Kim, J.S.; Kang, Y.M.; Lim, J.H.; Kim, Y.M.; Lee, M.S.; Je, J.Y. Antioxidant activity and γ-aminobutyric acid (GABA) content in sea tangle fermented by Lactobacillus brevis BJ20 isolated from traditional fermented foods. Food Chem. 2010, 122, 271–276. [Google Scholar] [CrossRef]
- Noureen, S.; Riaz, A.; Arshad, M.; Arshad, N. In vitro selection and in vivo confirmation of the antioxidant ability of Lactobacillus brevis MG000874. J. Appl. Microbiol. 2019, 126, 1221–1232. [Google Scholar] [CrossRef] [PubMed]
- Rajoka, M.S.R.; Mehwish, H.M.; Zhang, H.; Ashraf, M.; Fang, H.; Zeng, X.; He, Z. Antibacterial and anti-oxidant activity of exopolysaccharide mediated silver nanoparticle synthesized by Lactobacillus brevis isolated from Chinese koumiss. Colloid. Surface B 2020, 186, 110734. [Google Scholar] [CrossRef] [PubMed]
- Jin, Y.; Wu, J.; Hu, D.; Li, J.; Zhu, W.; Yuan, L.; Yao, J. Gamma-aminobutyric acid-producing Levilactobacillus brevis strains as probiotics in litchi juice fermentation. Foods 2023, 12, 302. [Google Scholar] [CrossRef] [PubMed]
- Pineiro, M.; Asp, N.G.; Reid, G.; Macfarlane, S.; Morelli, L.; Brunser, O.; Tuohy, K. FAO Technical meeting on prebiotics. J. Clin. Gastroenterol. 2008, 42, S156–S159. [Google Scholar] [CrossRef] [PubMed]
- Gibson, G.R.; Hutkins, R.; Sanders, M.E.; Prescott, S.L.; Reimer, R.A.; Salminen, S.J.; Reid, G. Expert consensus document: The International Scientific Association for Probiotics and Prebiotics (ISAPP) consensus statement on the definition and scope of prebiotics. Nat. Rev. Gastroenterol. Hepatol. 2017, 14, 491–502. [Google Scholar] [CrossRef] [PubMed]
- Osimani, A.; Ferrocino, I.; Corvaglia, M.R.; Roncolini, A.; Milanović, V.; Garofalo, C.; Clementi, F. Microbial dynamics in rearing trials of Hermetia illucens larvae fed coffee silverskin and microalgae. Food Res. Int. 2021, 140, 110028. [Google Scholar] [CrossRef] [PubMed]
- Herald, T.J.; Gadgil, P.; Tilley, M. High-throughput micro plate assays for screening flavonoid content and DPPH-scavenging activity in sorghum bran and flour. J. Sci. Food Agric. 2012, 92, 2326–2331. [Google Scholar] [CrossRef] [PubMed]
- Iriondo-DeHond, A.; Martorell, P.; Genovés, S.; Ramón, D.; Stamatakis, K.; Fresno, M.; Del Castillo, M.D. Coffee silverskin extract protects against accelerated aging caused by oxidative agents. Molecules 2016, 21, 721. [Google Scholar] [CrossRef] [PubMed]
- Ainsworth, E.A.; Gillespie, K.M. Estimation of total phenolic content and other oxidation substrates in plant tissues using Folin–Ciocalteu reagent. Nat. Protoc. 2007, 2, 875–877. [Google Scholar] [CrossRef] [PubMed]
- Medina, M.B. Determination of the Total Phenolics in Juices and Superfruits by a Novel Chemical Method. J. Funct. Foods 2011, 3, 79–87. [Google Scholar] [CrossRef]
- Siroli, L.; Giordani, B.; Rossi, S.; Gottardi, D.; McMahon, H.; Augustyniak, A.; Tabanelli, G.; Lanciotti, R. Antioxidant and Functional Features of Pre-Fermented Ingredients Obtained by the Fermentation of Milling By-Products. Fermentation 2022, 8, 722. [Google Scholar] [CrossRef]
- Mladenović, K.G.; Grujović, M.Ž.; Kiš, M.; Furmeg, S.; Tkalec, V.J.; Stefanović, O.D.; Kocić-Tanackov, S.D. Enterobacteriaceae in food safety with an emphasis on raw milk and meat. Appl. Microbiol. Biotechnol. 2021, 105, 8615–8627. [Google Scholar] [CrossRef] [PubMed]
- Nolasco, A.; Squillante, J.; Esposito, F.; Velotto, S.; Romano, R.; Aponte, M.; Cirillo, T. Coffee silverskin: Chemical and biological risk assessment and health profile for its potential use in functional foods. Foods 2022, 11, 2834. [Google Scholar] [CrossRef] [PubMed]
- Hashimoto, A.; Sugimoto, Y.; Suehara, K.I.; Kameoka, T. Influences of pH and temperature on infrared spectroscopic features of brewed coffee. Procedia Food Sci. 2011, 1, 1132–1138. [Google Scholar] [CrossRef]
- Monente, C.; Ludwig, I.A.; Irigoyen, A.; De Peña, M.-P.; Cid, C. Assessment of total (free and bound) phenolic compounds in spent coffee extracts. J. Agric. Food Chem. 2015, 63, 4327–4334. [Google Scholar] [CrossRef] [PubMed]
- Yadav, P.; Chauhan, A.K.; Singh, R.B.; Khan, S.; Halabi, G. Singh, R.B., Ed.; Organic acids: Microbial sources, production, and applications. In Functional Foods and Nutraceuticals in Metabolic and Non-Communicable Diseases; Academic Press: Cambridge, MA, USA, 2022; pp. 325–337. [Google Scholar]
- Borrelli, R.C.; Esposito, F.; Napolitano, A.; Ritieni, A.; Fogliano, V. Characterization of a new potential functional ingredient: Coffee silverskin. J. Agric. Food Chem. 2004, 52, 1338–1343. [Google Scholar] [CrossRef] [PubMed]
- Renaudie, M.; Dumas, C.; Vuilleumier, S.; Ernst, B. New way of valorization of raw coffee silverskin: Biohydrogen and acetate production by dark fermentation without exogenous inoculum. Bioresour. Technol. Rep. 2022, 17, 100918. [Google Scholar] [CrossRef]
- Hijosa-Valsero, M.; Garita-Cambronero, J.; Paniagua-García, A.I.; Díez-Antolínez, R. Biobutanol production from coffee silverskin. Microb. Cell Fact. 2018, 17, 154. [Google Scholar] [CrossRef] [PubMed]
- Rebollo-Hernanz, M.; Fernández-Gómez, B.; Herrero, M.; Aguilera, Y.; Martín-Cabrejas, M.A.; Uribarri, J.; Del Castillo, M.D. Inhibition of the Maillard reaction by phytochemicals composing an aqueous coffee silverskin extract via a mixed mechanism of action. Foods 2019, 8, 438. [Google Scholar] [CrossRef] [PubMed]
- Einarsson, H.; Snygg, B.G.; Eriksson, C. Inhibition of bacterial growth by Maillard reaction products. J. Agric. Food Chem. 1983, 31, 1043–1047. [Google Scholar] [CrossRef]
- Ferdes, M. Antimicrobial compounds from plants. In Fighting Antimicrobial Resistance; IAPC-OBP: Zagreb, Croatia, 2018; pp. 243–271. [Google Scholar]
- Geck, R.C.; Moresi, N.G.; Anderson, L.M.; Brewer, R.; Renz, T.R.; Taylor, M.B.; Dunham, M.J. Experimental evolution of Saccharomyces cerevisiae for caffeine tolerance alters multidrug resistance and target of rapamycin signaling pathways. G3 Genes Genomes Genet. 2024, 14, jkae148. [Google Scholar] [CrossRef] [PubMed]
- Nilsson, R.L.K.; Holmgren, M.; Madavi, B.; Nilsson, R.T.; Sellstedt, A. Adaptability of Trametes versicolor to the lignocellulosic inhibitors furfural, HMF, phenol and levulinic acid during ethanol fermentation. Biomass Bioenergy 2016, 90, 95–100. [Google Scholar]
- Milić, M.D.; Buntić, A.V.; Mihajlovski, K.R.; Ilić, N.V.; Davidović, S.Z.; Dimitrijević-Branković, S.I. The development of a combined enzymatic and microbial fermentation as a viable technology for the spent coffee ground full utilization. Biomass Convers. Biorefin. 2023, 13, 6747–6759. [Google Scholar] [CrossRef]
- Machado, M.; Galrinho, M.F.; Passos, C.P.; Santo, L.E.; Chiș, M.S.; Ranga, F.; Alves, R.C. Prebiotic potential of a coffee silverskin extract obtained by ultrasound-assisted extraction on Lacticaseibacillus paracasei subsp. paracasei. J. Funct. Foods 2024, 120, 106378. [Google Scholar] [CrossRef]
- Tomičić, Z.; Šarić, L.; Tomičić, R. Novel insights in the application of probiotic yeast Saccharomyces boulardii in dairy products and health promotion. Foods 2024, 13, 2866. [Google Scholar] [CrossRef] [PubMed]
- De Paula, B.P.; Chávez, D.W.H.; Lemos Junior, W.J.F.; Guerra, A.F.; Corrêa, M.F.D.; Pereira, K.S.; Coelho, M.A.Z. Growth parameters and survivability of Saccharomyces boulardii for probiotic alcoholic beverages development. Front. Microbiol. 2019, 10, 2092. [Google Scholar] [CrossRef] [PubMed]
- Azam, M.; Saeed, M.; Ahmad, T.; Yamin, I.; Khan, W.A.; Iqbal, M.W.; Riaz, T. Characterization of biopolymeric encapsulation system for improved survival of Lactobacillus brevis. J. Food Meas. Charact. 2022, 16, 2292–2299. [Google Scholar] [CrossRef]
- Alves, R.C.; Rodrigues, F.; Nunes, M.A.; Vinha, A.F.; Oliveira, M.B.P. Alves, R.C., Ed.; State of the art in coffee processing by-products. In Handbook of Coffee Processing By-Products; Academic Press Inc: Cambridge, MA, USA, 2017; pp. 1–26. [Google Scholar]
- de Miranda, J.F.; Belo, G.M.P.; de Lima, L.S.; Silva, K.A.; Uekane, T.M.; Gonzalez, A.G.M.; Lima, A.R. Arabic coffee infusion based kombucha: Characterization and biological activity during fermentation, and in vivo toxicity. Food Chem. 2023, 412, 135556. [Google Scholar] [CrossRef] [PubMed]
- Muzaifa, M.; Abubakar, Y.; Nilda, C. Phytochemicals and Sensory Quality of Cascara Kombucha Made From Coffee By-Products. Curr. Res. Nutr. Food Sci. 2023, 11, 2. [Google Scholar] [CrossRef]
- Lester, G.E.; Lewers, K.S.; Medina, M.B.; Saftner, R.A. Comparative Analysis of Strawberry Total Phenolics via Fast Blue BB vs. Folin–Ciocalteu: Assay Interference by Ascorbic Acid. J. Food Compos. Anal. 2012, 27, 102–107. [Google Scholar] [CrossRef]
- Pico, J.; Pismag, R.Y.; Laudouze, M.; Martinez, M.M. Systematic Evaluation of the Folin–Ciocalteu and Fast Blue BB Reactions during the Analysis of Total Phenolics in Legumes, Nuts and Plant Seeds. Food Funct. 2020, 11, 9868–9880. [Google Scholar] [CrossRef] [PubMed]
- Zou, L.; Zhang, H.; Liu, Z.; Sun, J.; Hu, Y.; Ding, Y.; Hu, B. Analyzing the Effect of Microbial Consortia Fermentation on the Quality of HnB by Untargeted Metabolomics. J. Microbiol. Biotechnol. 2024, 34, 1890. [Google Scholar] [CrossRef] [PubMed]
- Yaqoob, S.; Imtiaz, A.; Khalifa, I.; Maqsood, S.; Ullah, R.; Shahat, A.A.; Ma, Y. Multi-frequency sono-fermentation with mono and co-cultures of LAB synergistically enhance mulberry juice: Evidence from metabolic, micromorphological, sensorial, and computational approaches. Ultrason. Sonochem. 2024, 107, 117. [Google Scholar] [CrossRef] [PubMed]
- Ghosh, S.; Chowdhury, R.; Bhattacharya, P. Mixed consortia in bioprocesses: Role of microbial interactions. Appl. Microbiol. Biotechnol. 2016, 100, 4283–4295. [Google Scholar] [CrossRef] [PubMed]
- AOAC. Official Methods of Analysis of AOAC International, 18th ed.; AOAC International: Gaithersburg, MD, USA, 2010; Volume 6, pp. 90022–90025. [Google Scholar]
- Gottstein, V.; Bernhardt, M.; Dilger, E.; Keller, J.; Breitling-Utzmann, C.M.; Schwarz, S.; Bunzel, M. Coffee Silver Skin: Chemical Characterization with Special Consideration of Dietary Fiber and Heat-Induced Contaminants. Foods 2021, 10, 1705. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
{kind=link}
| Type of Microorganisms | Number of Cells [log CFU/g] |
|---|---|
| Mesophilic aerobic bacteria | 0.2 ± 0.10 |
| Lactic acid bacteria | n.d. |
| Enterobacteriaceae | n.d. |
| Bacterial spores | n.d. |
| Eumycetes | n.d. |
| Sample | 0 h | 24 h | 48 h |
|---|---|---|---|
| C | 4.9 ± 0.00 Aa | 4.89 ± 0.00 Ca | 4.89 ± 0.00 Ca |
| F Y | 4.91 ± 0.00 Ac | 4.45 ± 0.00 Bb | 4.36 ± 0.00 Ba |
| F AAB | 4.91 ± 0.00 Ac | 4.40 ± 0.00 ABb | 4.33 ± 0.00 ABa |
| F LAB | 4.90 ± 0.00 Ac | 4.45 ± 0.00 Bb | 4.36 ± 0.00 Ba |
| F Y + AAB | 4.91 ± 0.00 Ac | 4.41 ± 0.00 ABb | 4.32 ± 0.00 ABa |
| F Y + LAB | 4.91 ± 0.00 Ac | 4.39 ± 0.00 ABb | 4.30 ± 0.00 ABa |
| F AAB + LAB | 4.91 ± 0.00 Ac | 4.4 ± 0.00 ABb | 4.32 ± 0.00 ABa |
| F Y + AAB + LAB | 4.92 ± 0.00 Ac | 4.38 ± 0.00 Ab | 4.28 ± 0.00 Aa |
| Microbial Strain | Time Point [h] | C | F Y | F AAB | F LAB | F Y + AAB | F Y + LAB | F AAB + LAB | F Y + AAB + LAB |
|---|---|---|---|---|---|---|---|---|---|
| S. ludwigii | 0 | n.d. | 5.5 ± 0.10 Aa | n.d. | n.d. | 5.54 ± 0.03 Aa | 5.52 ± 0.21 Aa | n.d. | 5.49 ± 0.14 Aa |
| 24 | n.d. | 5.69 ± 0.08 Ac | n.d. | n.d. | 5.69 ± 0.23 Ac | 5.69 ± 0.07 Ac | n.d. | 5.7 ± 0.91 Ac | |
| 48 | n.d. | 5.64 ± 0.56 Ab | n.d. | n.d. | 5.68 ± 0.08 Bb | 5.66 ± 0.13 Bb | n.d. | 5.66 ± 0.02 Bb | |
| G. oxydans | 0 | n.d. | n.d. | 5.53 ± 0.45 Aa | n.d. | 5.52 ± 0.34 Aa | n.d. | 5.52 ± 0.54 Aa | 5.51 ± 0.21 Aa |
| 24 | n.d. | n.d. | 7.34 ± 0.13 Ac | n.d. | 7.31 ± 0.19 Bc | n.d. | 7.61 ± 0.21 Cc | 7.64 ± 0.63 Cc | |
| 48 | n.d. | n.d. | 7.23 ± 0.26 Ab | n.d. | 7.27 ± 0.42 Bb | n.d. | 7.58 ± 0.21 Db | 7.61 ± 0.06 Cb | |
| L. brevis | 0 | n.d. | n.d. | n.d. | 5.53 ± 0.18 Aa | n.d. | 5.52 ± 0.15 Aa | 5.5 ± 0.06 Aa | 5.51 ± 0.02 Aa |
| 24 | n.d. | n.d. | n.d. | 7.68 ± 0.18 Ac | n.d. | 7.89 ± 0.21 Ac | 7.90 ± 0.14 Bc | 7.90 ± 0.16 Bc | |
| 48 | n.d. | n.d. | n.d. | 7.63 ± 0.03 Ab | n.d. | 7.82 ± 0.21 Ab | 7.87 ± 0.02 Bb | 7.83 ± 0.04 Bb |
| Sample | FC | FBBB | DPPH | ABTS |
|---|---|---|---|---|
| (GAE mg/g) | Inhibition (%) | |||
| C | 10.72 ± 0.01 AB | 3.41 ± 0.02 A | 10.03 ± 0.19 A | 21.48 ± 1.28 A |
| F Y | 9.29 ± 0.04 A | 4.19 ± 0.04 AB | 17.68 ± 2.87 B | 25.36 ± 1.86 B |
| F AAB | 8.25 ± 0.04 A | 4.12 ± 0.01 AB | 18.04 ± 3.15 B | 25.47 ± 2.31 B |
| F LAB | 8.70 ± 0.06 A | 4.02 ± 0.13 AB | 18.34 ± 2.12 B | 25.23 ± 1.06 B |
| F Y + AAB | 12.28 ± 0.09 BC | 4.15 ± 0.18 AB | 19.25 ± 0.29 B | 23.60 ± 1.16 B |
| F Y + LAB | 14.09 ± 0.13 C | 3.81 ± 0.11 AB | 19.59 ± 0.55 B | 25.84 ± 1.79 B |
| F AAB + LAB | 13.94 ± 0.11 C | 4.30 ± 0.06 B | 20.35 ± 3.44 B | 23.64 ± 3.96 B |
| F Y + AAB + LAB | 14.15 ± 0.06 C | 7.12 ± 0.31 C | 20.43 ± 0.33 B | 27. 69 ± 0.86 B |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Guzińska, N.; Castillo, M.D.d.; Kordialik-Bogacka, E. Fermentation to Increase the Value of Roasted Coffee Silverskin as a Functional Food Ingredient. Foods 2025, 14, 2608. https://doi.org/10.3390/foods14152608
Guzińska N, Castillo MDd, Kordialik-Bogacka E. Fermentation to Increase the Value of Roasted Coffee Silverskin as a Functional Food Ingredient. Foods. 2025; 14(15):2608. https://doi.org/10.3390/foods14152608
Chicago/Turabian StyleGuzińska, Nadia, Maria Dolores del Castillo, and Edyta Kordialik-Bogacka. 2025. "Fermentation to Increase the Value of Roasted Coffee Silverskin as a Functional Food Ingredient" Foods 14, no. 15: 2608. https://doi.org/10.3390/foods14152608
APA StyleGuzińska, N., Castillo, M. D. d., & Kordialik-Bogacka, E. (2025). Fermentation to Increase the Value of Roasted Coffee Silverskin as a Functional Food Ingredient. Foods, 14(15), 2608. https://doi.org/10.3390/foods14152608