
Application of Microbial Fermentation in Caffeine Degradation and Flavor Modulation of Coffee Beans

 and
and 
Abstract
1. Introduction
2. Caffeine and Decaffeinated Coffee
3. Decaffeinated Coffee Production Methods: Advantages and Limitations
3.1. Industrial Decaffeinated Coffee: Physical and Chemical Methods
3.2. Naturally Low-Caffeine and Genetically Modified Coffee
4. Microbial Fermentation for Caffeine Degradation
4.1. Diversity and Mechanisms of Caffeine-Degrading Microorganisms

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Microorganism | Carbon Source | Degradation Pathway | Source/Application | Degradation Efficacy | Flavor Change | References | |
|---|---|---|---|---|---|---|---|
| 1 | R. oryzae FNCC6010 | — | — | Coffee Beans | 0.05 mg·g−1·h−1 | — | [15] |
| 2 | S. cerevisiae | — | — | Coffee Beans | 0.018 mg·g−1·h−1 | — | [15] |
| 3 | Leuconostoc mesenteroides | — | — | Coffee Beans | 0.020 mg·g−1·h−1 | [15] | |
| 4 | Lactobacillus casei | — | — | Coffee Beans | 0.020 mg·g−1·h−1 | [15] | |
| 5 | Torulaspora delbrueckii CCMA 0648 | — | — | Coffee Beans | 0.048 mg·g−1·h−1 | Chocolate, Caramel, Honey | [46] |
| 6 | Aspergillus ochraceus | — | — | Coffee Beans | — | — | [76] |
| 7 | Aspergillus sp. | — | N-demethylation | Coffee husk | — | — | [71] |
| 8 | Aspergillus sp. V12A25 | Caffeine + sucrose | — | Coffee grounds | 6.54 mg·L−1·h−1 | — | [77] |
| 9 | Aspergillus fumigatus C11B25 | Caffeine + sucrose | — | Coffee grounds | 5.00 mg·L−1·h−1 | — | [77] |
| 10 | Aspergillus niger C16A25 | Caffeine + sucrose | — | Coffee grounds | 5.07 mg·L−1·h−1 | — | [77] |
| 11 | Rhizopus sp. LPB-79 | — | — | Coffee husk | — | — | [71] |
| 12 | R. delemar | Caffeine + sugars | N-demethylation | Coffee husk | — | — | [78] |
| 13 | Rhizopus oryzae MUCL 28168 | _ | N-demethylation | Coffee pulp | — | — | [75] |
| 14 | Phanerochaete chrysosporium BK | — | — | Coffee husk | — | — | [71] |
| 15 | Pleurotus ostreatus | — | N-demethylation | Coffee grounds | — | — | [72] |
| 16 | Brevibacterium sp. MTCC 10313 | Glucose + Caffeine | — | Coffee Pulp | — | — | [79] |
| 17 | Penicillium simplicissimum 4–17 | — | — | Tea | 0.043 mg·g−1·h−1 | like the smell of fermented soyabean | [13] |
| 18 | Aspergillus niger NCBT110A | glucose | — | Pu’er Tea | — | — | [14] |
| 19 | Aspergillus sydowii NRRL250 | Caffaine | N-demethylation | Pu’er Tea | 2.57 mg·L−1·h−1 | — | [14] |
| 20 | Candida famata ACCC 2052 | — | — | Black/Green Tea | — | — | [80] |
| 21 | Candida albicans ACCC 2100 | — | — | Black/Green Tea | — | — | [80] |
4.2. Enzymes Involved in Caffeine Degradation
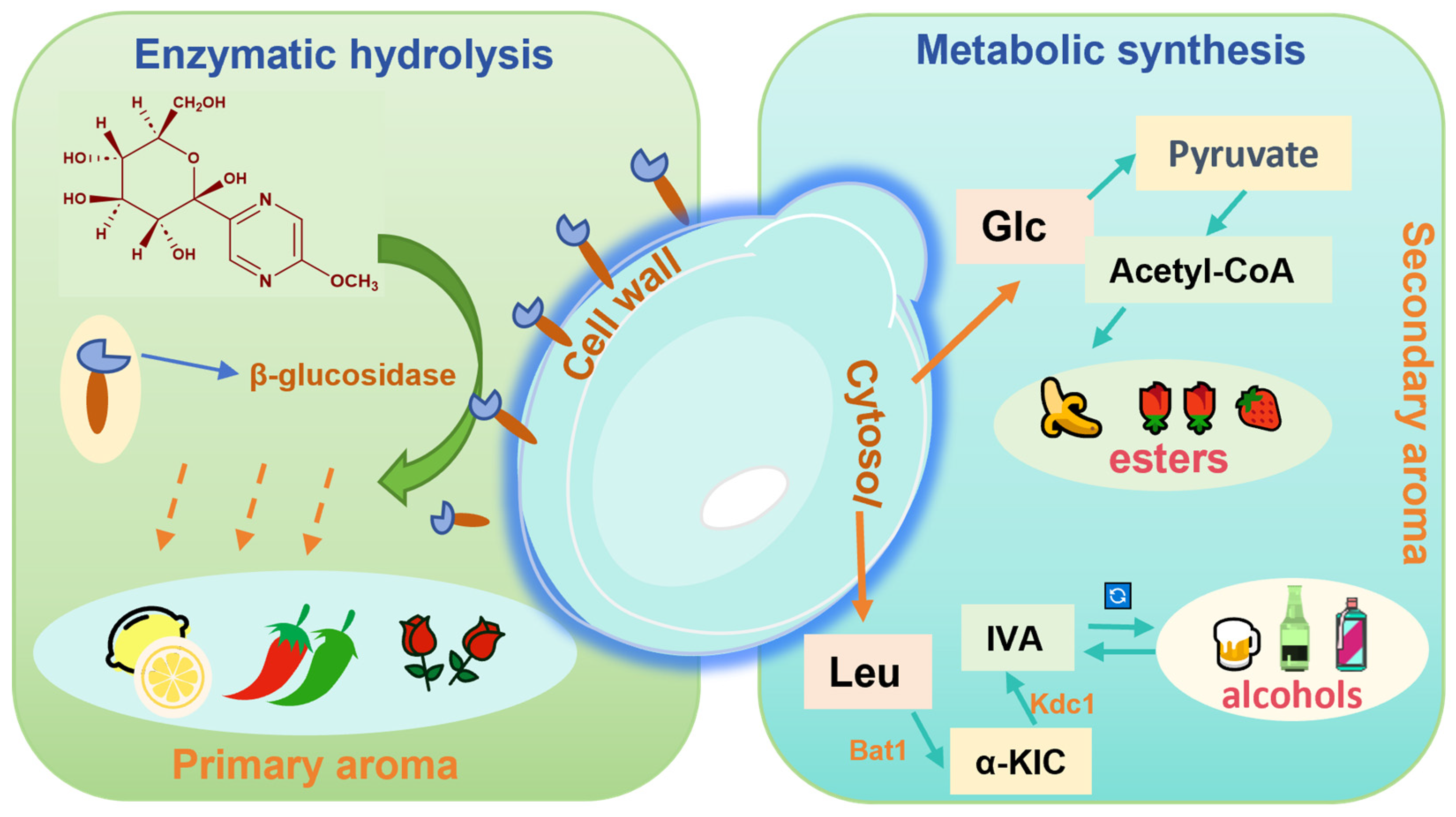
4.3. Metabolic Shifts in Caffeine-Degrading Fermentation
4.4. Screening of Caffeine-Degrading Strains
4.4.1. Caffeine Degradation Capacity of the Strains
4.4.2. Alkaloid Tolerance of the Strain
4.4.3. Environmental Adaptability of Alkaloid-Degrading Microorganisms
4.4.4. Safety and Quality of Low-Caffeine Coffee Products
5. Coffee Fermentation
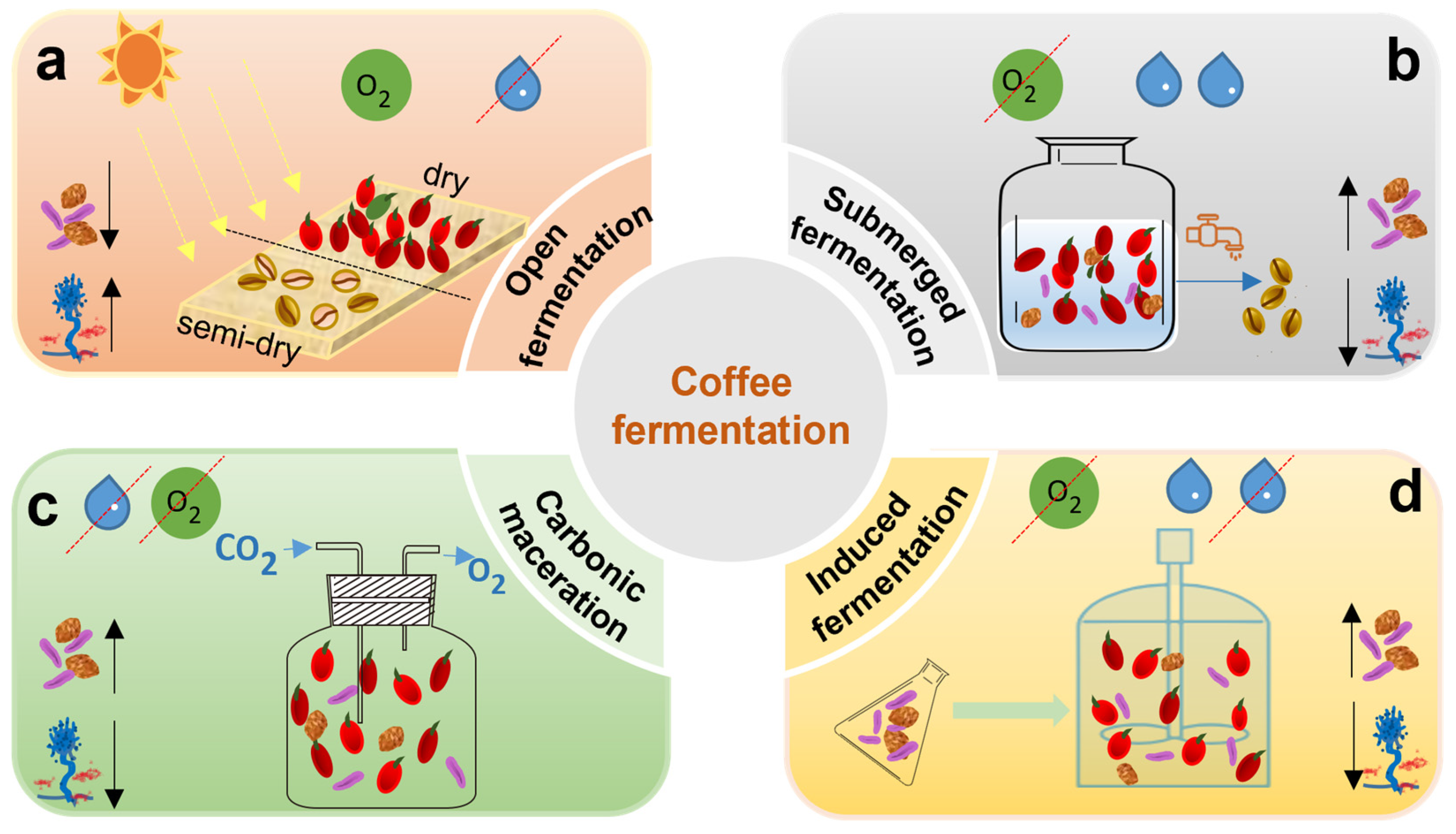
5.1. Types of Coffee Fermentation
5.2. Microbial Interaction Mechanisms in Coffee Fermentation
5.3. Mixed-Culture Coffee Fermentation
5.4. Optimization of Mixed-Culture Coffee Fermentation
6. Conclusions and Future Perspectives
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Cassimiro, D.M.J.; Batista, N.N.; Fonseca, H.C.; Oliveira Naves, J.A.; Coelho, J.M.; Bernardes, P.C.; Dias, D.R.; Schwan, R.F. Wet Fermentation of Coffea Canephora by Lactic Acid Bacteria and Yeasts Using the Self-Induced Anaerobic Fermentation (SIAF) Method Enhances the Coffee Quality. Food Microbiol. 2023, 110, 104161. [Google Scholar] [CrossRef]
- Lang, T.; Lang, R.; Di Pizio, A.; Mittermeier, V.K.; Schlagbauer, V.; Hofmann, T.; Behrens, M. Numerous Compounds Orchestrate Coffee’s Bitterness. J. Agric. Food Chem. 2020, 68, 6692–6700. [Google Scholar] [CrossRef] [PubMed]
- Ferreira, G.A.; Felippe, L.C.; Bertuzzi, R.; Bishop, D.J.; Ramos, I.S.; De-Oliveira, F.R.; Lima-Silva, A.E. Does Caffeine Ingestion before a Short-Term Sprint Interval Training Promote Body Fat Loss? Braz. J. Med. Biol. Res. 2019, 52, e9169. [Google Scholar] [CrossRef] [PubMed]
- Giles, G.E.; Spring, A.M.; Urry, H.L.; Moran, J.M.; Mahoney, C.R.; Kanarek, R.B. Caffeine Alters Emotion and Emotional Responses in Low Habitual Caffeine Consumers. Can. J. Physiol. Pharmacol. 2018, 96, 191–199. [Google Scholar] [CrossRef] [PubMed]
- Jahrami, H.; Al-Mutarid, M.; Penson, P.E.; Al-Islam Faris, M.; Saif, Z.; Hammad, L. Intake of Caffeine and Its Association with Physical and Mental Health Status among University Students in Bahrain. Foods 2020, 9, 473. [Google Scholar] [CrossRef]
- Mejia, E.G.D.; Ramirez-Mares, M.V. Impact of Caffeine and Coffee on Our Health. Trends Endocrinol. Metab. 2014, 25, 489–492. [Google Scholar] [CrossRef]
- Turnbull, D.; Rodricks, J.V.; Mariano, G.F.; Chowdhury, F. Caffeine and Cardiovascular Health. Regul. Toxicol. Pharm. 2017, 89, 165–185. [Google Scholar] [CrossRef]
- Guimares, P.S.; Schenk, J.C.M.; Cintra, L.C.; Giachetto, P.F.; Silvarolla, M.B.; Padilha, L.; Maluf, M.P. Large-Scale Prospection of Genes on Caffeine-Free Coffea arabica Plants-Discovery of Novel Markers Associated with Development and Secondary Metabolism. Plant Gene 2021, 27, 100314. [Google Scholar] [CrossRef]
- Leibrock, N.V.; Santegoets, J.; Mooijman, P.J.W.; Yusuf, F.; Zuijdgeest, X.C.L.; Zutt, E.A.; Jacobs, J.G.M.; Schaart, J.G. The Biological Feasibility and Social Context of Gene-Edited, Caffeine-Free Coffee. Food Sci. Biotechnol. 2022, 31, 635–655. [Google Scholar] [CrossRef]
- Zhou, B.; Ma, C.; Ren, X.; Xia, T.; Li, X. LC-MS/MS-Based Metabolomic Analysis of Caffeine-Degrading Fungus Aspergillus Sydowii during Tea Fermentation. J. Food Sci. 2020, 85, 477–485. [Google Scholar] [CrossRef]
- Fan, F.-Y.; Xu, Y.; Liang, Y.-R.; Zheng, X.-Q.; Borthakur, D.; Lu, J.-L. Isolation and Characterization of High Caffeine-Tolerant Bacterium Strains from the Soil of Tea Garden. Afr. J. Microbiol. Res. 2011, 5, 2278–2286. [Google Scholar]
- Babu, V.R.S.; Patra, S.; Thakur, M.S.; Karanth, N.G.; Varadaraj, M.C. Degradation of Caffeine by Pseudomonas alcaligenes CFR 1708. Enzym. Microb. Technol. 2005, 37, 617–624. [Google Scholar] [CrossRef]
- Yu, Z.; Deng, H.; Qu, H.; Zhang, B.; Lei, G.; Chen, J.; Feng, X.; Wu, D.; Huang, Y.; Ji, Z. Penicillium simplicissimum Possessing High Potential to Develop Decaffeinated Qingzhuan Tea. LWT 2022, 165, 113606. [Google Scholar] [CrossRef]
- Zhou, B.; Ma, C.; Wang, H.; Xia, T. Biodegradation of Caffeine by Whole Cells of Tea-Derived Fungi Aspergillus sydowii, Aspergillus niger and Optimization for Caffeine Degradation. BMC Microbiol. 2018, 18, 53. [Google Scholar] [CrossRef] [PubMed]
- Purwoko, T.; Suranto; Ratna, S.; Soerya, D.M. Chlorogenic Acid and Caffeine Content of Fermented Robusta Bean. Biodiversitas J. Biol. Divers. 2022, 23, 902–906. [Google Scholar] [CrossRef]
- Peng, C.; Zhang, Y.; Li, C.; Shang, Y.; Liu, J.; Min, Y.; Tang, J.; Xiang, W.; Zhang, Q. Elucidating the Biogenic Transformation Mechanisms of Polyphenol and Volatile Compound Metabolites in Wild Cherry Wine Co-Fermentation by Hanseniaspora uvarum and Saccharomyces cerevisiae. LWT 2025, 217, 117420. [Google Scholar] [CrossRef]
- Cassimiro, D.M.D.J.; Batista, N.N.; Fonseca, H.C.; Naves, J.A.O.; Dias, D.R.; Schwan, R.F. Coinoculation of Lactic Acid Bacteria and Yeasts Increases the Quality of Wet Fermented Arabica Coffee. Int. J. Food Microbiol. 2022, 369, 109627. [Google Scholar] [CrossRef]
- Bastian, F.; Hutabarat, O.S.; Dirpan, A.; Nainu, F.; Harapan, H.; Emran, T.B.; Simal-Gandara, J. From Plantation to Cup: Changes in Bioactive Compounds during Coffee Processing. Foods 2021, 10, 2827. [Google Scholar] [CrossRef]
- Areniello, M.; Matassa, S.; Esposito, G.; Lens, P.N.L. Biowaste Upcycling into Second-Generation Microbial Protein through Mixed-Culture Fermentation. Trends Biotechnol. 2023, 41, 197–213. [Google Scholar] [CrossRef]
- Helena Sances Rabelo, M.; Meira Borém, F.; Paula de Carvalho Alves, A.; Soares Pieroni, R.; Mendes Santos, C.; Nakajima, M.; Sugino, R. Fermentation of Coffee Fruit with Sequential Inoculation of Lactiplantibacillus plantarum and Saccharomyces cerevisiae: Effects on Volatile Composition and Sensory Characteristics. Food Chem. 2024, 444, 138608. [Google Scholar] [CrossRef]
- Su, Y.; Dong, Q.; Chen, Y.; Wang, R.; Jiang, J.; Qin, Y.; Song, Y.; Liu, Y. Impact of Sequential Inoculation Timing on the Quality of Wine Fermented by Indigenous Lachancea thermotolerans and Saccharomyces cerevisiae. LWT 2024, 204, 116438. [Google Scholar] [CrossRef]
- Lu, Y.; Putra, S.D.; Liu, S.Q. A Novel Non-Dairy Beverage from Durian Pulp Fermented with Selected Probiotics and Yeast. Int. J. Food Microbiol. 2018, 265, 1–8. [Google Scholar] [CrossRef]
- dePaula, J.; Farah, A. Caffeine Consumption through Coffee: Content in the Beverage, Metabolism, Health Benefits and Risks. Beverages 2019, 5, 37. [Google Scholar] [CrossRef]
- Bhardwaj, I.; Ansari, A.H.; Rai, S.P.; Singh, S.; Singh, D. Chapter Three-Molecular Targets of Caffeine in the Central Nervous System. In Progress in Brain Research; Moradikor, N., Chatterjee, I., Eds.; Neuroscience of Coffee Part A; Elsevier: Amsterdam, The Netherlands, 2024; Volume 288, pp. 35–58. [Google Scholar]
- López-Canales, O.A.; Ortiz-Hernández, M.; Lozano-Cuenca, J.; Herrera-Tolentino, O.C.; López-Canales, J.S.; Nicolás-Velázquez, P.; Paredes-Carbajal, M.C. Evaluation of Differences in Cardiovascular and Metabolic Effects of Chronic Caffeinated and Decaffeinated Coffee Intake. J. Funct. Foods 2024, 121, 106419. [Google Scholar] [CrossRef]
- Heckman, M.A.; Weil, J.; De Mejia, E.G. Caffeine (1, 3, 7-trimethylxanthine) in Foods: A Comprehensive Review on Consumption, Functionality, Safety, and Regulatory Matters. J. Food Sci. 2010, 75, R77–R87. [Google Scholar] [CrossRef] [PubMed]
- Mazzafera, P.; Baumann, T.W.; Shimizu, M.M.; Silvarolla, M.B. Decaf and the Steeplechase towards Decaffito—The Coffee from Caffeine-Free Arabica Plants. Trop. Plant. Biol. 2009, 2, 63–76. [Google Scholar] [CrossRef]
- Shlonsky, A. Traits of Persons Who Drink Decaffeinated Coffee. Ann. Epidemiol. 2003, 13, 273–279. [Google Scholar] [CrossRef]
- Chindapan, N.; Soydok, S.; Devahastin, S. Roasting Kinetics and Chemical Composition Changes of Robusta Coffee Beans during Hot Air and Superheated Steam Roasting. J. Food Sci. 2019, 84, 292–302. [Google Scholar] [CrossRef]
- Zhang, Y.; Ma, C.; Zhao, L.; Mucci, L.A.; Giovannucci, E.L. Decaffeinated Coffee Consumption and Risk of Total and Site-Specific Cancer. Ann. Oncol. 2025, 36, 819–831. [Google Scholar] [CrossRef]
- Lee, J.-Y.; Kim, Y.-S. Changes in Aroma Compounds of Decaffeinated Coffee Beans. Korean J. Food Preserv. 2023, 30, 492–501. [Google Scholar] [CrossRef]
- Liang, H.; Liang, Y.; Dong, J.; Lu, J.; Xu, H.; Wang, H. Decaffeination of Fresh Green Tea Leaf (Camellia sinensis) by Hot Water Treatment. Food Chem. 2007, 101, 1451–1456. [Google Scholar] [CrossRef]
- Chen, X.; Du, Y.; Wu, L.; Xie, J.; Chen, X.; Hu, B.; Wu, Z.; Yao, Q.; Li, Q. Effects of Tea-Polysaccharide Conjugates and Metal Ions on Precipitate Formation by Epigallocatechin Gallate and Caffeine, the Key Components of Green Tea Infusion. J. Agric. Food Chem. 2019, 67, 3744–3751. [Google Scholar] [CrossRef] [PubMed]
- Sotelo, J.L.; Ovejero, G.; Rodríguez, A.; Álvarez, S.; Galán, J.; García, J. Competitive Adsorption Studies of Caffeine and Diclofenac Aqueous Solutions by Activated Carbon. Chem. Eng. J. 2014, 240, 443–453. [Google Scholar] [CrossRef]
- Maluf, M.P.; Silva, C.C.D.; Oliveira, M.D.P.A.D.; Tavares, A.G.; Silvarolla, M.B.; Guerreiro Filho, O. Altered Expression of the Caffeine Synthase Gene in a Naturally Caffeine-Free Mutant of Coffea arabica. Genet. Mol. Biol. 2009, 32, 802–810. [Google Scholar] [CrossRef]
- Silvarolla, M.B.; Mazzafera, P.; Fazuoli, L.C. A Naturally Decaffeinated Arabica Coffee. Nature 2004, 429, 826. [Google Scholar] [CrossRef]
- Narko, T.; Wibowo, M.S.; Damayanti, S.; Wibowo, I. Effect of Kombucha Culture on Caffeine and Chlorogenic Acid Content in Fermentation of Robusta Green Coffee Beans (Coffea canephora L.). Rasayan J. Chem. 2020, 13, 1181–1186. [Google Scholar] [CrossRef]
- Rocha, T.R.; Carvalho, A.; Fazuoli, L.C. Melhoramento Do Cafeeiro: XXXVIII. Observações Sobre Progênies Do Cultivar Mundo-Novo de Coffea arabica Na Estação Experimental de Mococa. Bragantia 1980, 39, 147–160. [Google Scholar] [CrossRef]
- Jiang, Y.; Lu, Y.; Huang, Y.; Chen, S.; Ji, Z. Bacillus amyloliquefaciens HZ-12 Heterologously Expressing NdmABCDE with Higher Ability of Caffeine Degradation. LWT 2019, 109, 387–394. [Google Scholar] [CrossRef]
- Kim, J.H.; Kim, B.H.; Brooks, S.; Kang, S.Y.; Summers, R.M.; Song, H.K. Structural and Mechanistic Insights into Caffeine Degradation by the Bacterial N-Demethylase Complex. J. Mol. Biol. 2019, 431, 3647–3661. [Google Scholar] [CrossRef]
- Sledz, W.; Los, E.; Paczek, A.; Rischka, J.; Motyka, A.; Zoledowska, S.; Piosik, J.; Lojkowska, E. Antibacterial Activity of Caffeine against Plant Pathogenic Bacteria. Acta Biochim. Pol. 2015, 62, 605–612. [Google Scholar] [CrossRef]
- Mock, M.B.; Summers, R.M. Microbial Metabolism of Caffeine and Potential Applications in Bioremediation. J. Appl. Microbiol. 2024, 135, lxae080. [Google Scholar] [CrossRef]
- Lukman, K.; Ibrahim, S.; Muhammad, A.; Babandi, A.; Yakasai, H.M.; Muhammad, J.B.; Jagaba, A.H. Bacillus sp. KS38 Strain for Sustainable Caffeine Degradation: Isolation, Identification and Optimization Using Response Surface Methodology. Desalin. Water Treat. 2024, 320, 100628. [Google Scholar] [CrossRef]
- Cao, L.; Garcia, S.L.; Wurzbacher, C. Profiling Trace Organic Chemical Biotransformation Genes, Enzymes and Associated Bacteria in Microbial Model Communities. J. Hazard. Mater. 2025, 485, 136811. [Google Scholar] [CrossRef]
- Dave, B.; Moysa, E.L.; Kunik, A. Enhancing Fungal Adaptation for Efficient Caffeine Degradation in Wastewater: Biomimetic Approach and Environmental Optimization. Desalin. Water Treat. 2025, 321, 100938. [Google Scholar] [CrossRef]
- Bressani, A.P.P.; Batista, N.N.; Ferreira, G.; Martinez, S.J.; Simão, J.B.P.; Dias, D.R.; Schwan, R.F. Characterization of Bioactive, Chemical, and Sensory Compounds from Fermented Coffees with Different Yeasts Species. Food Res. Int. 2021, 150, 110755. [Google Scholar] [CrossRef]
- Summers, R.M.; Louie, T.M.; Yu, C.L.; Subramanian, M. Characterization of a Broad-Specificity Non-Haem Iron N-Demethyl ase from Pseudomonas putida CBB5 Capable of Utilizing Several Purine Alkaloids as Sole Carbon and Nitrogen Source. Microbi Ology 2011, 157, 583–592. [Google Scholar] [CrossRef]
- Mohanty, S.K.; Yu, C.-L.; Das, S.; Louie, T.M.; Gakhar, L.; Subramanian, M. Delineation of the Caffeine C-8 Oxidation Pathway in Pseudomonas sp. Strain CBB1 via Characterization of a New Trimethyluric Acid Monooxygenase and Genes Involved in Trimethyluric Acid Metabolism. J. Bacteriol. 2012, 194, 3872–3882. [Google Scholar] [CrossRef] [PubMed]
- Lin, Z.; Wei, J.; Hu, Y.; Pi, D.; Jiang, M.; Lang, T. Caffeine Synthesis and Its Mechanism and Application by Microbial Degradation, a Review. Foods 2023, 12, 2721. [Google Scholar] [CrossRef] [PubMed]
- Shanmugam, M.K.; Rathinavelu, S.; Gummadi, S.N. Self-Directing Optimization for Enhanced Caffeine Degradation in Synthetic Coffee Wastewater Using Induced Cells of Pseudomonas sp.: Bioreactor Studies. J. Water Process Eng. 2021, 44, 102341. [Google Scholar] [CrossRef]
- Summers, R.M.; Shao, J.; Mock, M.B.; Yu, C.L.; Vega, F.E. Draft Genome Sequence of Pseudomonas sp. Strain CES, Containing the Entire Alkylxanthine Gene Cluster for Caffeine Breakdown. Microbiol. Resour. Announc. 2020, 9, e00484-20. [Google Scholar] [CrossRef]
- Thathola, P.; Agnihotri, V.; Pandey, A. Microbial Degradation of Caffeine Using Himalayan Psychrotolerant Pseudomonas sp. GBPI_Hb5 (MCC 3295). Curr. Microbiol. 2021, 78, 3924–3935. [Google Scholar] [CrossRef]
- Shotaro, U.; Yasuhiro, U.; Hiroshi, I.T.T.M.E. Caffeine 7-N-Demethylation and C-8-Oxidation Mediated by Liver Microsomal Cytochrome P450 Enzymes in Common Marmosets. Xenobiotica Fate Foreign Compd. Biol. Syst. 2016, 46, 573–578. [Google Scholar]
- Summers, R.M.; Mohanty, S.K.; Gopishetty, S.; Subramanian, M. Genetic Characterization of Caffeine Degradation by Bacteria and Its Potential Applications. Microb. Biotechnol. 2015, 8, 369–378. [Google Scholar] [CrossRef]
- Ma, Y.-X.; Wu, X.-H.; Wu, H.-S.; Dong, Z.-B.; Ye, J.-H.; Zheng, X.-Q.; Liang, Y.-R.; Lu, J. Different Catabolism Pathways Triggered by Various 1 Methylxanthines in Caffeine-Tolerant Bacterium Pseudomonas putida CT25 Isolated from Tea Garden Soil. J. Microbiol. Biotechnol. 2018, 28, 1147–1155. [Google Scholar] [CrossRef]
- Ramarethinam, S.; Rajalakshmi, N. Caffeine in Tea Plants [Camellia sinensis (L) O. Kuntze]: In Situ Lowering by Bacillus licheniformis (Weigmann) Chester. Indian J. Exp. Biol. 2004, 42, 575–580. [Google Scholar] [CrossRef]
- Siddharth, S.; Renuka, J.; Anja, A.A.; Rounaq, N.; Suneetha, V. A Preliminary Study and First Report on Caffeine Degrading Bacteria Isolated from the Soils of Chittoor and Vellore. Int. Res. J. Pharm. 2012, 3, 305. [Google Scholar]
- Sun, D.; Yang, X.; Zeng, C.; Li, B.; Wang, Y.; Zhang, C.; Hu, W.; Li, X.; Gao, Z. Novel Caffeine Degradation Gene Cluster Is Mega-Plasmid Encoded in Paraburkholderia caffeinilytica CF1. Appl. Microbiol. Biotechnol. 2020, 104, 3025–3036. [Google Scholar] [CrossRef] [PubMed]
- Ashengroph, M. Salinivibrio Costicola GL6, a Novel Isolated Strain for Biotransformation of Caffeine to Theobromine under Hypersaline Conditions. Curr. Microbiol. 2017, 74, 34–41. [Google Scholar] [CrossRef] [PubMed]
- Mazzafera, P.; Olsson, O.; Sandberg, G. Degradation of Caffeine and Related Methylxanthines by Serratia marcescens Isolated from Soil under Coffee Cultivation. Microb. Ecol. 1996, 31, 199–207. [Google Scholar] [CrossRef] [PubMed]
- Mohapatra, B.R.; Harris, N.; Nordin, R.; Mazumder, A. Purification and Characterization of a Novel Caffeine Oxidase from Alcaligenes Species. J. Biotechnol. 2006, 125, 319–327. [Google Scholar] [CrossRef]
- Madyastha, K.M.; Sridhar, G.R. A Novel Pathway for the Metabolism of Caffeine by a Mixed Culture Consortium. Biochem. Biophys. Res. Commun. 1998, 249, 178–181. [Google Scholar] [CrossRef] [PubMed]
- Varadarasu, L.; Nilanjana, D. Bioremediation of Caffeine-Contaminated Soil by Immobilized Yeast—A Laboratory Based Study. Nat. Environ. Pollut. Technol. 2012, 11, 385–392. [Google Scholar]
- Ashengroph, M.; Borchaluei, M. Saccharomyces cerevisiae TFS9, a Novel Isolated Yeast Capable of High Caffeine-Tolerant and Its Application in Biodecaffeination Approach. Prog. Biol. Sci. 2013, 3, 145–156. [Google Scholar] [CrossRef]
- Liu, F.; Song, Z.; Zhang, T.; Tong, X.; Chen, M.Y.; Gao, D.; Chen, J.; Ho, C.L. Characterization of the Therapeutic Properties and Flavor Profile of Coffee via Monoculture Fermentation with Endophytic Microbial Isolates. ACS Food Sci. Technol. 2022, 2, 1039–1049. [Google Scholar] [CrossRef]
- Lakshmi; Devlina, D.; Nilanjana, D. Biodegradation of Caffeine by the Yeast Trichosporon asahii Immobilized in Single and Hybrid Matrices. Indian J. Chem. Technol. 2013, 20, 195–201. [Google Scholar]
- Ashengroph, M. A Novel Strain of Aureobasidium Sp. TeO12 for Theophylline Production from Caffeine. Biotech 2017, 7, 176. [Google Scholar] [CrossRef]
- Nayak, V.; Pai, P.V.; Pai, A.; Pai, S.; Sushma, Y.D.; Rao, C.V. A Comparative Study of Caffeine Degradation by Four Different Fungi. Bioremediat. J. 2013, 17, 79–85. [Google Scholar] [CrossRef]
- Kobetičová, K.; Nábělková, J.; Petříková, M.; Kočí, V.; Jerman, M.; Černý, R. Biodegradation of Methylxanthines by Coniophora puteana. In AIP Conference Proceeding, Proceedings of the Central European Symposium on Thermophysics 2021 (CEST 2021), Kazimierz Dolny, Poland, 1–3 September 2021; AIP Publishing LLC: New York, NY, USA, 2021; p. 20014. [Google Scholar] [CrossRef]
- Nanjundaiah, S.; Mutturi, S.; Bhatt, P. Modeling of Caffeine Degradation Kinetics during Cultivation of Fusarium solani Using Sucrose as Co-Substrate. Biochem. Eng. J. 2017, 125, 73–80. [Google Scholar] [CrossRef]
- Brand, D.; Pandey, A.; Roussos, S.; Soccol, C.R. Biological Detoxification of Coffee Husk by Filamentous Fungi Using a Solid State Fermentation System. Enzym. Microb. Technol. 2000, 27, 127–133. [Google Scholar] [CrossRef]
- Carrasco-Cabrera, C.P.; Bell, T.L.; Kertesz, M.A. Caffeine Metabolism during Cultivation of Oyster Mushroom (Pleurotus ostreatus) with Spent Coffee Grounds. Appl. Microbiol. Biotechnol. 2019, 103, 5831–5841. [Google Scholar] [CrossRef]
- Kurtzman, R.H.; Schwimmer, S. Caffeine Removal from Growth Media by Microorganisms. Experientia 1971, 27, 481–482. [Google Scholar] [CrossRef] [PubMed]
- Hussein, A.A.; Al-Janabi, S. The Ability of Dermatophytes to Utilize Methylxanthine as Sole Source of Carbon and Nitrogen. Curr. Res. J. Biol. Ences 2009, 1, 163–167. [Google Scholar]
- Peña-Lucio, E.M.; Londoño-Hernández, L.; Ascacio-Valdes, J.A.; Chavéz-González, M.L.; Bankole, O.E.; Aguilar, C.N. Use of Coffee Pulp and Sorghum Mixtures in the Production of N-Demethylases by Solid-State Fermentation. Bioresour. Technol. 2020, 305, 123112. [Google Scholar] [CrossRef] [PubMed]
- Tsubouchi, H.; Terada, H.; Yamamoto, K.; Hisada, K.; Sakabe, Y. Caffeine Degradation and Increased Ochratoxin a Production by Toxigenic Strains of Aspergillus ochraceus Isolated from Green Coffee Beans. Mycopathologia 1985, 90, 181–186. [Google Scholar] [CrossRef]
- Roussos, S.; De Los Angeles Aquiáhuatl, M.; Del Refugio Trejo-Hernández, M.; Gaime Perraud, I.; Favela, E.; Ramakrishna, M.; Raimbault, M.; Viniegra-González, G. Biotechnological Management of Coffee Pulp—Isolation, Screening, Characterization, Selection of Caffeine-Degrading Fungi and Natural Microflora Present in Coffee Pulp and Husk. Appl. Microbiol. Biotechnol. 1995, 42, 756–762. [Google Scholar] [CrossRef]
- Tagliari, C.V.; Sanson, R.K.; Zanette, A.; Franco, T.T.; Soccol, C.R. Caffeine Degradation by Rhizopus Delemar in Packed Bed Column Bioreactor Using Coffee Husk as Substrate. Braz. J. Microbiol. 2003, 34, 102–104. [Google Scholar] [CrossRef]
- Nayak, S.; Harshitha, M.J.; Maithili, C.; Sampath, H.S.; Anilkumar, C.V.; Rao, C.V. Isolation and Characterization of Caffeine Degrading Bacteria from Coffee Pulp. Indian J. Biotechnol. 2012, 57, 81–83. [Google Scholar] [CrossRef]
- Wang, X.; Wan, X.; Hu, S.; Pan, C. Study on the Increase Mechanism of the Caffeine Content during the Fermentation of Tea with Microorganisms. Food Chem. 2008, 107, 1086–1091. [Google Scholar] [CrossRef]
- Summers, R.M.; Louie, T.M.; Yu, C.-L.; Gakhar, L.; Louie, K.C.; Subramanian, M. Novel, Highly Specific N-Demethylases Enable Bacteria to Live on Caffeine and Related Purine Alkaloids. J. Bacteriol. 2012, 194, 2041–2049. [Google Scholar] [CrossRef]
- Summers, R.M.; Seffernick, J.L.; Quandt, E.M.; Yu, C.L.; Barrick, J.E.; Subramanian, M.V. Caffeine Junkie: An Unprecedented Glutathione S-Transferase-Dependent Oxygenase Required for Caffeine Degradation by Pseudomonas putida CBB5. J. Bacteriol. 2013, 195, 3933–3939. [Google Scholar] [CrossRef]
- Glück, M.; Lingens, F. Heteroxanthinedemethylase, a New Enzyme in the Degradation of Caffeine by Pseudomonas putida. Appl. Microbiol. Biotechnol. 1988, 28, 59–62. [Google Scholar] [CrossRef]
- Yamaoka-Yano, D.M.; Mazzafera, P. Catabolism of Caffeine and Purification of a Xanthine Oxidade Responsible for Methyluric Acids Production in Pseudomonas putida L. Rev. Microbiol. 1999, 30, 62–70. [Google Scholar] [CrossRef]
- Madyastha, K.M.; Sridhar, G.R.; Vadiraja, B.B.; Madhavi, Y.S. Purification and Partial Characterization of Caffeine Oxidase—A Novel Enzyme from a Mixed Culture Consortium. Biochem. Biophys. Res. Commun. 1999, 263, 460–464. [Google Scholar] [CrossRef] [PubMed]
- Gambacorta, F.V.; Dietrich, J.J.; Yan, Q.; Pfleger, B.F. Rewiring Yeast Metabolism to Synthesize Products beyond Ethanol. Curr. Opin. Chem. Biol. 2020, 59, 182. [Google Scholar] [CrossRef]
- Han, X.; Qin, Q.; Li, C.; Zhao, X.; Song, F.; An, M.; Chen, Y.; Wang, X.; Huang, W.; Zhan, J.; et al. Application of Non-Saccharomyces Yeasts with High β-Glucosidase Activity to Enhance Terpene-Related Floral Flavor in Craft Beer. Food Chem. 2023, 404, 134726. [Google Scholar] [CrossRef]
- Dzialo, M.C.; Park, R.; Steensels, J.; Lievens, B.; Verstrepen, K.J. Physiology, Ecology and Industrial Applications of Aroma Formation in Yeast. FEMS Microbiol. Rev. 2017, 41, S95–S128. [Google Scholar] [CrossRef]
- Bhatia, S.K.; Yang, Y.-H. Microbial Production of Volatile Fatty Acids: Current Status and Future Perspectives. Rev. Environ. Sci. Bio/Technol. 2017, 16, 327–345. [Google Scholar] [CrossRef]
- Cai, M.; Huang, L.; Dong, S.; Diao, N.; Ye, W.; Peng, Z.; Fang, X. Enhancing the Flavor Profile of Summer Green Tea via Fermentation with Aspergillus niger RAF106. Foods 2023, 12, 3420. [Google Scholar] [CrossRef]
- Gummadi, S.; Santhosh, D. How Induced Cells of Pseudomonas sp. Increase the Degradation of Caffeine. Open Life Sci. 2006, 1, 561–571. [Google Scholar] [CrossRef]
- Das, N. Caffeine Degradation by Yeasts Isolated from Caffeine Contaminated Samples. Int. J. Sci. Nat. 2010, 1, 47–52. [Google Scholar]
- Gummadi, S.N.; Bhavya, B. Enhanced Degradation of Caffeine and Caffeine Demethylase Production by Pseudomonas Sp. in Bioreactors under Fed-Batch Mode. Appl. Microbiol. Biotechnol. 2011, 91, 1007–1017. [Google Scholar] [CrossRef]
- Arimurti, S.; Afifuddin, R.B.; Siswanto, S.; Muzakhar, K. Characterization and Identification of Caffeine-Degrading Bacteria KAJ36. Pelita Perkeb. 2020, 36, 173–175. [Google Scholar] [CrossRef]
- Win, Y.Y.; Singh, M.; Sadiq, M.B.; Anal, A.K. Isolation and Identification of Caffeine-Degrading Bacteria from Coffee Plantation Area. Food Biotechnol. 2019, 33, 109–124. [Google Scholar] [CrossRef]
- Ibrahim, S.; Shukor, M.Y.; Syed, M.A.; Johari, W.L.W.; Shamaan, N.A.; Sabullah, M.K.; Ahmad, S.A. Enhanced Caffeine Degradation by Immobilised Cells of Leifsonia Sp. Strain SIU. J. Gen. Appl. Microbiol. 2016, 62, 18–24. [Google Scholar] [CrossRef] [PubMed]
- Pereira, G.V.D.M.; Thomaz, S.V.; Pedroni, M.A.B.; Rodrigues, A.L.J.M.; Luiz, G.A.; Ricardo, S.C. Isolation, Selection and Evaluation of Yeasts for Use in Fermentation of Coffee Beans by the Wet Process. Int. J. Food Microbiol. 2014, 188, 60–66. [Google Scholar] [CrossRef]
- Liu, T.-Y.; Hsu, W.-H.; Lee, B.-H. Evaluation of the Application of Wild Yeasts in Inhibiting Germination of Ochratoxin-Producing Fungi during Coffee Fermentation Process. Food Chem. X 2025, 25, 102077. [Google Scholar] [CrossRef]
- Zhang, S.J.; De Bruyn, F.; Pothakos, V.; Contreras, G.F.; Cai, Z.; Moccand, C.; Weckx, S.; De Vuyst, L. Influence of Various Processing Parameters on the Microbial Community Dynamics, Metabolomic Profiles, and Cup Quality during Wet Coffee Processing. Front. Microbiol. 2019, 10, 2621. [Google Scholar] [CrossRef]
- Rocha, H.A.; Borém, F.M.; Alves, A.P.D.C.; Santos, C.M.D.; Schwan, R.F.; Haeberlin, L.; Nakajima, M.; Sugino, R. Natural Fermentation with Delayed Inoculation of the Yeast Torulaspora delbrueckii: Impact on the Chemical Composition and Sensory Profile of Natural Coffee. Food Res. Int. 2023, 174, 113632. [Google Scholar] [CrossRef]
- Janne Carvalho Ferreira, L.; de Souza Gomes, M.; Maciel de Oliveira, L.; Diniz Santos, L. Coffee Fermentation Process: A Review. Food Res. Int. 2023, 169, 112793. [Google Scholar] [CrossRef]
- Bressani, A.P.P.; Martinez, S.J.; Sarmento, A.B.I.; Borém, F.M.; Schwan, R.F. Influence of Yeast Inoculation on the Quality of Fermented Coffee (Coffea arabica Var. Mundo Novo) Processed by Natural and Pulped Natural Processes. Int. J. Food Microbiol. 2021, 343, 109107. [Google Scholar] [CrossRef]
- Tangjaidee, P.; Braspaiboon, S.; Singhadechachai, N.; Phongthai, S.; Therdtatha, P.; Rachtanapun, P.; Sommano, S.R.; Seesuriyachan, P. Enhanced Bioactive Coffee Cherry: Infusion of Submerged-Fermented Green Coffee Beans via Vacuum Impregnation. Foods 2025, 14, 1165. [Google Scholar] [CrossRef]
- Pothakos, V.; De Vuyst, L.; Zhang, S.J.; De Bruyn, F.; Verce, M.; Torres, J.; Callanan, M.; Moccand, C.; Weckx, S. Temporal Shotgun Metagenomics of an Ecuadorian Coffee Fermentation Process Highlights the Predominance of Lactic Acid Bacteria. Curr. Res. Biotechnol. 2020, 2, 1–15. [Google Scholar] [CrossRef]
- Pereira, T.S.; Batista, N.N.; Santos Pimenta, L.P.; Martinez, S.J.; Ribeiro, L.S.; Oliveira Naves, J.A.; Schwan, R.F. Self-Induced Anaerobiosis Coffee Fermentation: Impact on Microbial Communities, Chemical Composition and Sensory Quality of Coffee. Food Microbiol. 2022, 103, 103962. [Google Scholar] [CrossRef] [PubMed]
- Junior, D.B.; Guarconi, R.C.; da Silva, M.D.C.S.; Veloso, T.G.R.; Kasuya, M.C.M.; da Silva Oliveira, E.C.; da Luz, J.M.R.; Moreira, T.R.; Debona, D.G.; Pereira, L.L. Microbial Fermentation Affects Sensorial, Chemical, and Microbial Profile of Coffee under Carbonic Maceration. Food Chem. 2021, 342, 128296. [Google Scholar] [CrossRef] [PubMed]
- da Silva, M.C.S.; Veloso, T.G.R.; Brioschi Junior, D.; Bullergahn, V.B.; da Luz, J.M.R.; Menezes, K.M.S.; Guarçoni, R.C.; Kasuya, M.C.M.; Pereira, L.L. Bacterial Community and Sensory Quality from Coffee Are Affected along Fermentation under Carbonic Maceration. Food Chem. Adv. 2023, 3, 100554. [Google Scholar] [CrossRef]
- Entringer, T.L.; da Luz, J.M.R.; Veloso, T.G.R.; Pereira, L.L.; Menezes, K.M.S.; Brioschi Júnior, D.; Kasuya, M.C.M.; da Silva, M.C.S. Genetic Diversity of the Fungal Community That Contributes to the Sensory Quality of Coffee Beverage after Carbonic Maceration and Fermentation. 3 Biotech 2024, 14, 272. [Google Scholar] [CrossRef]
- Pereira, L.L.; Júnior, D.B.; de Sousa, L.H.B.P.; dos Santos Gomes, W.; Cardoso, W.S.; Guarçoni, R.C. Relationship between Coffee Processing and Fermentation. In Quality Determinants in Coffee Production; Springer Nature: Berlin/Heidelberg, Germany, 2021. [Google Scholar] [CrossRef]
- Martinez, S.J.; Batista, N.N.; Bressani, A.P.P.; Leite, S.; Oliveira Naves, J.A.; Dias, D.R.; Schwan, R.F. Volatiles and Sensory Mapping of SIAF Fermentations with Selected Coffee Yeasts from Different Brazilian Regions. Food Biosci. 2024, 62, 105551. [Google Scholar] [CrossRef]
- Martinez, S.J.; Sances Rabelo, M.H.; Pereira Bressani, A.P.; Caroline, B.D.M.; Borem, F.M.; Schwan, R.F. Novel Stainless Steel Tanks Enhances Coffee Fermentation Quality. Food Res. Int. 2021, 139, 109921. [Google Scholar] [CrossRef]
- Braga, A.V.U.; Miranda, M.A.; Aoyama, H.; Schmidt, F.L. Study on Coffee Quality Improvement by Self-Induced Anaerobic Fermentation: Microbial Diversity and Enzymatic Activity. Food Res. Int. 2023, 165, 112528. [Google Scholar] [CrossRef]
- Batista Da Mota, M.C.; Batista, N.N.; Dias, D.R.; Schwan, R.F. Impact of Microbial Self-Induced Anaerobiosis Fermentation (SIAF) on Coffee Quality. Food Biosci. 2022, 47, 101640. [Google Scholar] [CrossRef]
- Cardoso, W.S.; Agnoletti, B.Z.; De Freitas, R.; De Abreu Pinheiro, F.; Pereira, L.L. Biochemical Aspects of Coffee Fermentation. In Quality Determinants in Coffee Production; Food Engineering Series; Springer: Berlin/Heidelberg, Germany, 2021; pp. 149–208. ISBN 978-3-030-54436-2. [Google Scholar]
- Janne Carvalho Ferreira, L.; Luiz Lima Bertarini, P.; Rodrigues Do Amaral, L.; Souza Gomes, M.D.; Maciel De Oliveira, L.; Diniz Santos, L. Coinoculation of Saccharomyces cerevisiae and Bacillus amyloliquefaciens in Solid-State and Submerged Coffee Fermentation: Influences on Chemical and Sensory Compositions. LWT 2024, 202, 116299. [Google Scholar] [CrossRef]
- Vinícius De Melo Pereira, G.; Soccol, V.T.; Brar, S.K.; Neto, E.; Soccol, C.R. Microbial Ecology and Starter Culture Technology in Coffee Processing. Crit. Rev. Food Sci. Nutr. 2017, 57, 2775–2788. [Google Scholar] [CrossRef]
- De Melo Pereira, G.V.; Neto, E.; Soccol, V.T.; Medeiros, A.B.P.; Woiciechowski, A.L.; Soccol, C.R. Conducting Starter Culture-Controlled Fermentations of Coffee Beans during on-Farm Wet Processing: Growth, Metabolic Analyses and Sensorial Effects. Food Res. Int. 2015, 75, 348–356. [Google Scholar] [CrossRef]
- Chen, Y.; Jiang, J.; Li, Y.; Xie, Y.; Cui, M.; Hu, Y.; Yin, R.; Ma, X.; Niu, J.; Cheng, W.; et al. Enhancing Physicochemical Properties, Organic Acids, Antioxidant Capacity, Amino Acids and Volatile Compounds for ‘Summer Black’ Grape Juice by Lactic Acid Bacteria Fermentation. LWT 2024, 209, 116791. [Google Scholar] [CrossRef]
- Zhao, Q.-S.; Yang, J.-G.; Zhang, K.-Z.; Wang, M.-Y.; Zhao, X.-X.; Su, C.; Cao, X.-Z. Lactic Acid Bacteria in the Brewing of Traditional Daqu Liquor. J. Inst. Brew. 2020, 126, 14–23. [Google Scholar] [CrossRef]
- Ponomarova, O.; Gabrielli, N.; Sévin, D.C.; Mülleder, M.; Zirngibl, K.; Bulyha, K.; Andrejev, S.; Kafkia, E.; Typas, A.; Sauer, U.; et al. Yeast Creates a Niche for Symbiotic Lactic Acid Bacteria through Nitrogen Overflow. Cell Syst. 2017, 5, 345–357.e6. [Google Scholar] [CrossRef] [PubMed]
- Da Silva Vale, A.; De Melo Pereira, G.V.; De Carvalho Neto, D.P.; Rodrigues, C.; Pagnoncelli, M.G.B.; Soccol, C.R. Effect of Co-Inoculation with Pichia Fermentans and Pediococcus acidilactici on Metabolite Produced during Fermentation and Volatile Composition of Coffee Beans. Fermentation 2019, 5, 67. [Google Scholar] [CrossRef]
- Gonçalves Bravim, D.; Mota De Oliveira, T.; Kaic Alves Do Rosário, D.; Nara Batista, N.; Freitas Schwan, R.; Moreira Coelho, J.; Campos Bernardes, P. Inoculation of Yeast and Bacterium in Wet-Processed Coffea canephora. Food Chem. 2023, 400, 134107. [Google Scholar] [CrossRef]
- Borém, F.M.; Rabelo, M.H.S.; De Carvalho Alves, A.P.; Santos, C.M.; Pieroni, R.S.; Nakajima, M.; Sugino, R. Fermentation of Coffee Fruit with Sequential Inoculation of Lactiplantibacillus plantarum and Saccharomyces cerevisiae: Effect on Sensory Attributes and Chemical Composition of the Beans. Food Chem. 2024, 446, 138820. [Google Scholar] [CrossRef] [PubMed]
- Chan, M.Z.A.; Lau, H.; Lim, S.Y.; Li, S.F.Y.; Liu, S.-Q. Untargeted LC-QTOF-MS/MS Based Metabolomics Approach for Revealing Bioactive Components in Probiotic Fermented Coffee Brews. Food Res. Int. 2021, 149, 110656. [Google Scholar] [CrossRef]
- Bressani, A.P.P.; Martinez, S.J.; Batista, N.N.; Simão, J.B.P.; Dias, D.R.; Schwan, R.F. Co-Inoculation of Yeasts Starters: A Strategy to Improve Quality of Low Altitude Arabica Coffee. Food Chem. 2021, 361, 130133. [Google Scholar] [CrossRef]
- Wang, C.; Sun, J.; Lassabliere, B.; Yu, B.; Liu, S.Q. Coffee flavour modification through controlled fermentation of green coffee beans by Saccharomyces cerevisiae and Pichia kluyveri: Part II. Mixed cultures with or without lactic acid bacteria. Food Res. Int. 2020, 136, 109452. [Google Scholar] [CrossRef]
- Elhalis, H.; Cox, J.; Frank, D.; Zhao, J. Microbiological and Chemical Characteristics of Wet Coffee Fermentation Inoculated with Hansinaspora uvarum and Pichia kudriavzevii and Their Impact on Coffee Sensory Quality. Front. Microbiol. 2021, 12, 713969. [Google Scholar] [CrossRef] [PubMed]
- Pereira Bressani, A.P.; Monteiro De Andrade, H.; Ribeiro Dias, D.; Freitas Schwan, R. Protein Profile and Volatile Compound Associated with Fermented Coffees with Yeast Co-Inoculation. Food Res. Int. 2023, 174, 113494. [Google Scholar] [CrossRef] [PubMed]
- Tenglics, R.; Szappanos, B.; Mülleder, M.; Kalapis, D.; Grézal, G.; Sajben, C.; Agostini, F.; Mokochinski, J.B.; Bálint, B.; Nagy, L.G.; et al. The Metabolic Domestication Syndrome of Budding Yeast. Proc. Natl. Acad. Sci. USA 2024, 121, e2313354121. [Google Scholar] [CrossRef]
- Qin, S.; Hu, F.; Yang, N.; Li, L.; Yang, H.; Suo, Y.; He, F. Influence of Native Coffee Yeast Fermentation on Phenolic Content, Organic Acids, and Volatile Compounds in Cascara. LWT 2024, 210, 116860. [Google Scholar] [CrossRef]
- Prakash, I.; Shankar, S.; Sneha, H.P.; Kumar, P.; Hari, O.; Basavaraj, K.; Murthy, P.S. Metabolomics and Volatile Fingerprint of Yeast Fermented Robusta Coffee: A Value Added Coffee. LWT 2022, 154, 112717. [Google Scholar] [CrossRef]
- Pereira, G.V.D.M.; De Mello Sampaio, V.; Wiele, N.; Da Silva Vale, A.; De Carvalho Neto, D.P.; Souza, A.D.F.D.D.; Nogueira Dos Santos, D.V.; Ruiz, I.R.; Rogez, H.; Soccol, C.R. How Yeast Has Transformed the Coffee Market by Creating New Flavors and Aromas through Modern Post-Harvest Fermentation Systems. Trends Food Sci. Technol. 2024, 151, 104641. [Google Scholar] [CrossRef]
- García, C.; Rendueles, M.; Díaz, M. Liquid-Phase Food Fermentations with Microbial Consortia Involving Lactic Acid Bacteria: A Review. Food Res. Int. 2019, 119, 207–220. [Google Scholar] [CrossRef]
- Freire, A.L.; Ramos, C.L.; Da Costa Souza, P.N.; Cardoso, M.G.B.; Schwan, R.F. Nondairy Beverage Produced by Controlled Fermentation with Potential Probiotic Starter Cultures of Lactic Acid Bacteria and Yeast. Int. J. Food Microbiol. 2017, 248, 39–46. [Google Scholar] [CrossRef]
- Liu, Y.; Chua, X.Y.; Dong, W.; Lu, Y.; Liu, S.-Q. Effects of Sequential Inoculation of Lachancea thermotolerans and Oenococcus oeni on Chemical Composition of Spent Coffee Grounds Hydrolysates. Curr. Res. Food Sci. 2022, 5, 1276–1286. [Google Scholar] [CrossRef]


| Types | Mixed Fermentation | Fermentation/Inoculation Mode | Flavor Characteristics | References |
|---|---|---|---|---|
| Yeast + Bacteria | L. plantarum + S. cerevisiae | Sequential inoculation/wet | Greater acidity, sweetness, body | [123] |
| Lactobacillus rhamnosus GG+ Saccharomyces boulardii CNCM-I745 | Simultaneous inoculation/wet | _ | [124] | |
| S. cerevisiae + Bacillus amyloliquefaciens | Simultaneous inoculation/SIAF | Nutty, cocoa, and sweet | [115] | |
| Pichia fermentans YC5.2 + Pediococcus acidilactici LPBC161 | Combined fermentation/wet | Honey, floral, sweet and grass fragrance | [121] | |
| Leuconostoc mesenteroides + Torulaspora delbrueckii | SIAF/wet | Chocolate, wheat, green, woody, floral, caramel, | [1] | |
| Leuconostoc mesenteroides + S. cerevisiae | SIAF/wet | Chocolate, caramel, fruity, spices | ||
| L. plantarum + Torulaspora delbrueckii | SIAF/wet | Chocolate, dark chocolate, nutty, fruity, spices | ||
| L. plantarum + S. cerevisiae | SIAF/wet | Dark chocolate, caramel, nutty, spices | ||
| L. plantarum CCMA1065 + S. cerevisiae CCMA0543 | Co-inoculation/wet | Citrus, floral | [17] | |
| Bacillus licheniformis + Meyerozyma guilliermondii | Co-inoculation/wet | Almond, chocolate | [122] | |
| Yeast + Yeast | S. cerevisiae CCMA 0543 + Candida parapsilosis CCMA 0544 | Co-inoculation/dry | Fruity, nutty, roasted, sweet, almond, biter, cocoa, coffee, roasted nutty, caramel and peppermint, rose–honey-like floral–woody | [125] |
| S. cerevisiae CCMA 0543 + Torulaspora delbrueckii CCMA 0684 | Co-inoculation/dry | Fruity, caramel and peppermint, rose–honey-like floral–woody | ||
| Candida parapsilosis CCMA 0544 + Torulaspora delbrueckii CCMA 0684 | Co-inoculation/dry | Caramel and peppermint, rose–honey-like, floral–woody | ||
| Torulaspora delbrueckii+ Candida parapsilosis | SIAF/wet | Citrus, caramel, honey, chocolate, chestnut | [126] | |
| S. cerevisiae CCMA 0543 + Candida parapsilosis CCMA 0544 + Torulaspora delbrueckii CCMA 0684 | Co-inoculation/dry | Caramel and peppermint, rose–honey-like, floral–woody, citric acidity, fruity, banana and pear | [46] | |
| Hansinaspora uvarum +Pichia kudriavzevii | Combined fermentation/wet | Earthy, apple cider, walnut notes, smooth mouthfeel | [127] | |
| S. cerevisiae CCMA 0543 + Candida parapsilosis CCMA 0544 + Torulaspora delbrueckii CCMA 0684 | SIAF/wet | Caramel, peppermint, citric acidity | [128] | |
| Bacteria + Bacteria | Leuconostoc mesenteroides + L. plantarum | SIAF/wet | Caramel, fruity, spices | [1] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ran, L.-X.; Wei, X.-Y.; Ren, E.-F.; Qin, J.-F.; Rasheed, U.; Chen, G.-L. Application of Microbial Fermentation in Caffeine Degradation and Flavor Modulation of Coffee Beans. Foods 2025, 14, 2606. https://doi.org/10.3390/foods14152606
Ran L-X, Wei X-Y, Ren E-F, Qin J-F, Rasheed U, Chen G-L. Application of Microbial Fermentation in Caffeine Degradation and Flavor Modulation of Coffee Beans. Foods. 2025; 14(15):2606. https://doi.org/10.3390/foods14152606
Chicago/Turabian StyleRan, Lu-Xia, Xiang-Ying Wei, Er-Fang Ren, Jian-Feng Qin, Usman Rasheed, and Gan-Lin Chen. 2025. "Application of Microbial Fermentation in Caffeine Degradation and Flavor Modulation of Coffee Beans" Foods 14, no. 15: 2606. https://doi.org/10.3390/foods14152606
APA StyleRan, L.-X., Wei, X.-Y., Ren, E.-F., Qin, J.-F., Rasheed, U., & Chen, G.-L. (2025). Application of Microbial Fermentation in Caffeine Degradation and Flavor Modulation of Coffee Beans. Foods, 14(15), 2606. https://doi.org/10.3390/foods14152606