
Decoding Flavonoid Metabolism for Nutritional Enhancement: A Transcriptome–Metabolome Integration Study of Biosynthesis in Edible Chrysanthemum indicum L.
Abstract

1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Detection of Linarin
2.3. Electronic Tongue Analysis
2.4. Extraction of Flavonoids
2.5. Instrument Parameter Settings
2.6. Metabolomics Data Processing
2.7. RNA Extraction and Transcriptome Sequencing
2.8. Transcriptome Data Processing
2.9. Integrated Analysis of Metabolomics and Transcriptomics
2.10. RT-qPCR Analysis
3. Results
3.1. Metabolomic Differential Analysis
3.1.1. Analysis of Flavonoid Metabolomes
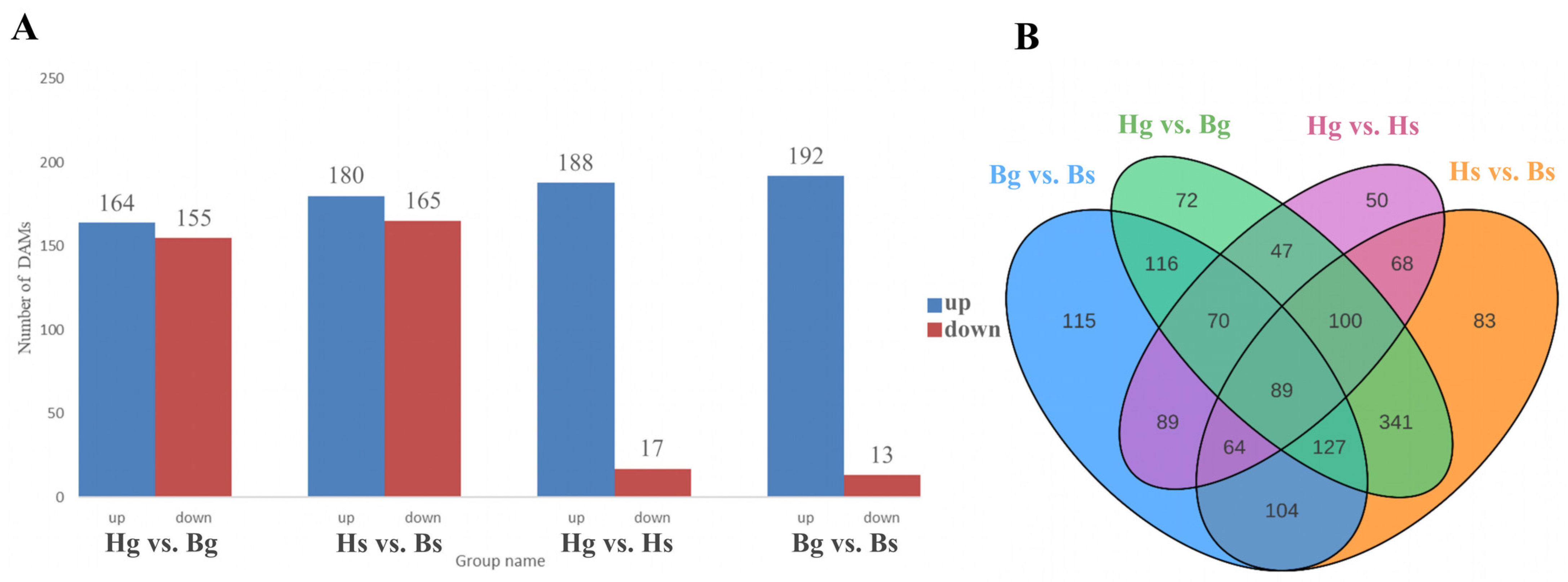
3.1.2. Analysis of Flavonoid Differentially Accumulated Metabolites (DAMs)
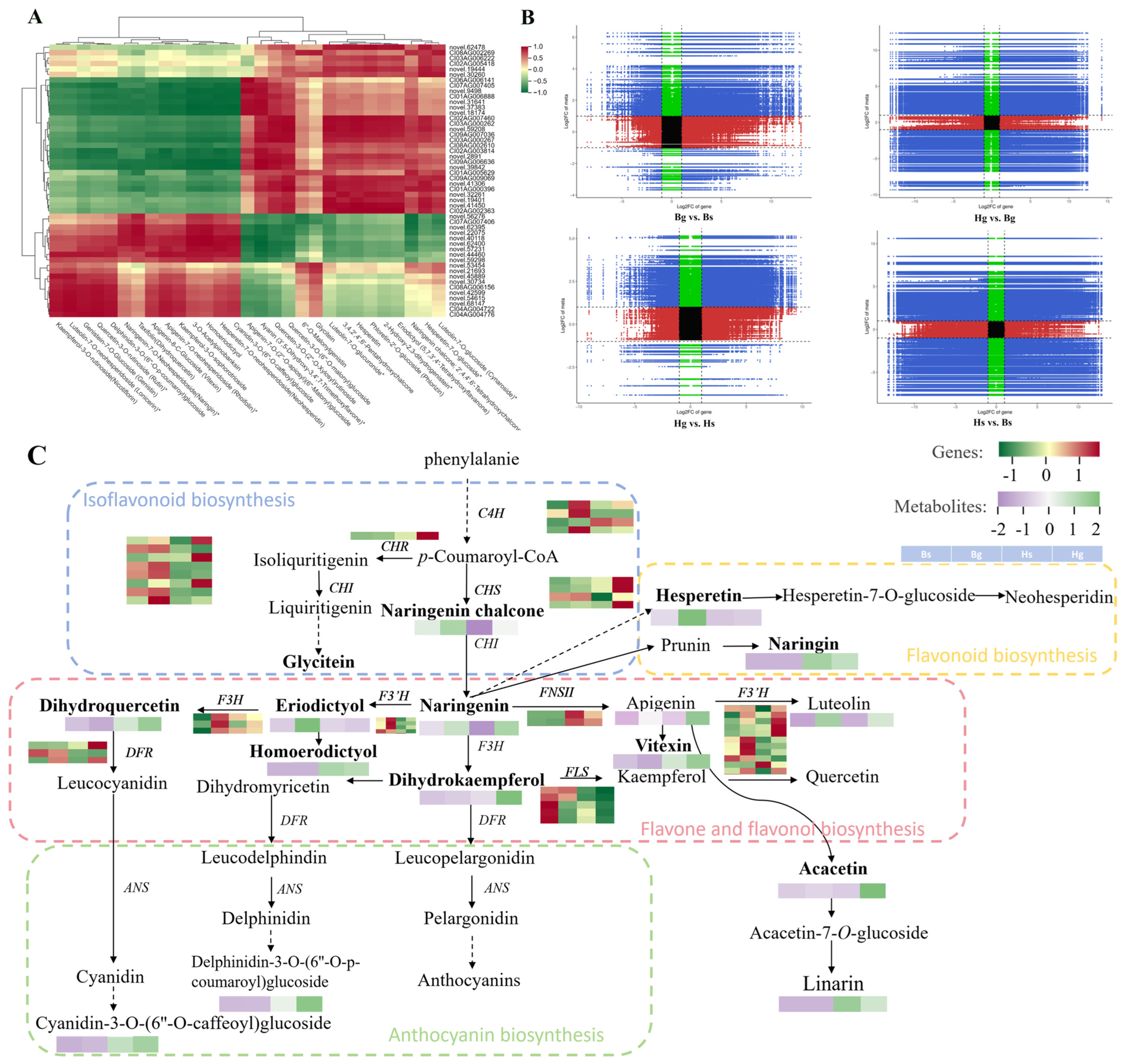
3.1.3. Cluster Analysis of Flavonoid Metabolites
3.1.4. Electronic Tongue Analysis of BHYJ and HJ06
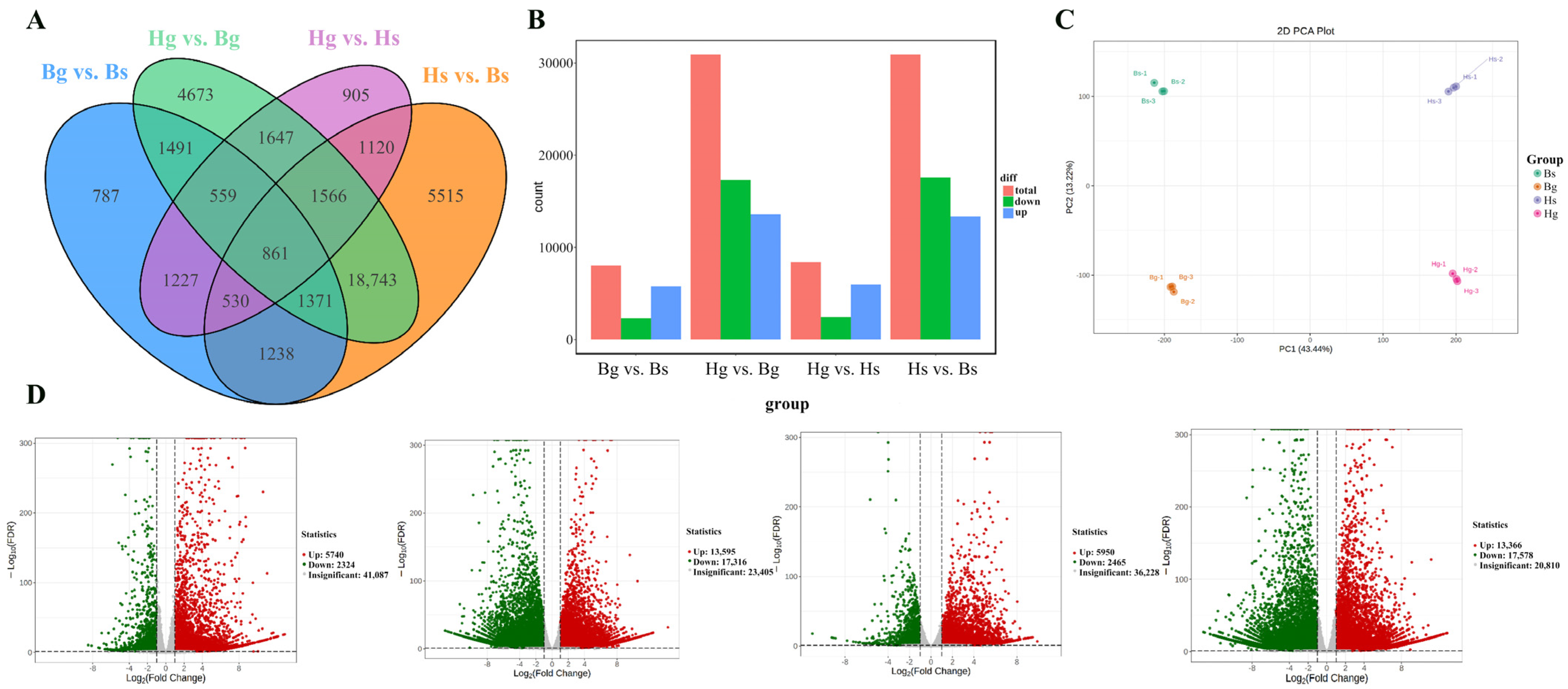
3.2. Transcriptome Differential Analysis
3.2.1. Library Construction and Functional Analysis
3.2.2. Functional Analysis of DEGs
3.2.3. Differential Gene and Key Enzyme Gene Selection
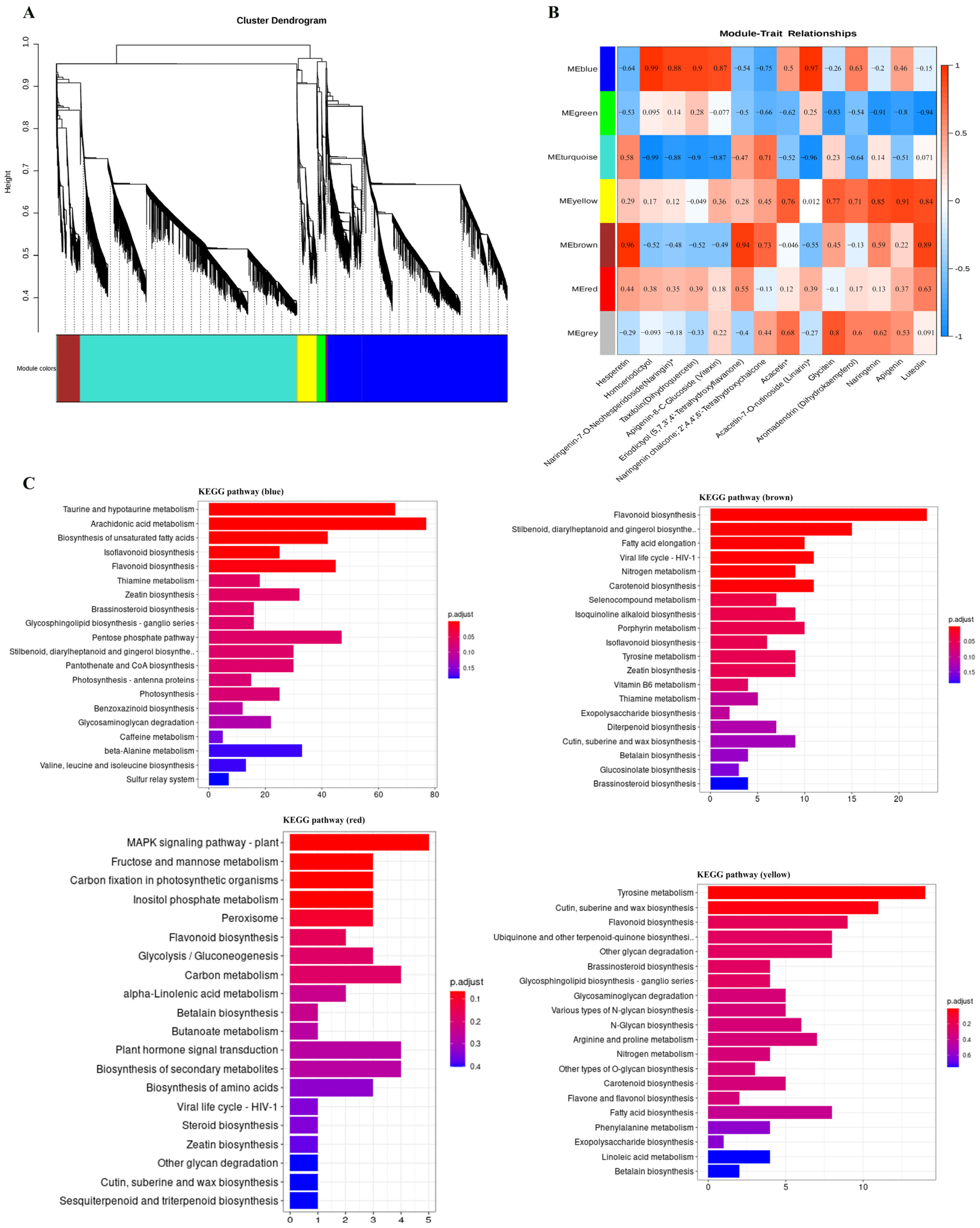
3.3. Gene Co-Expression Network Analysis
3.4. Integration Analysis of Metabolomics and Transcriptomics
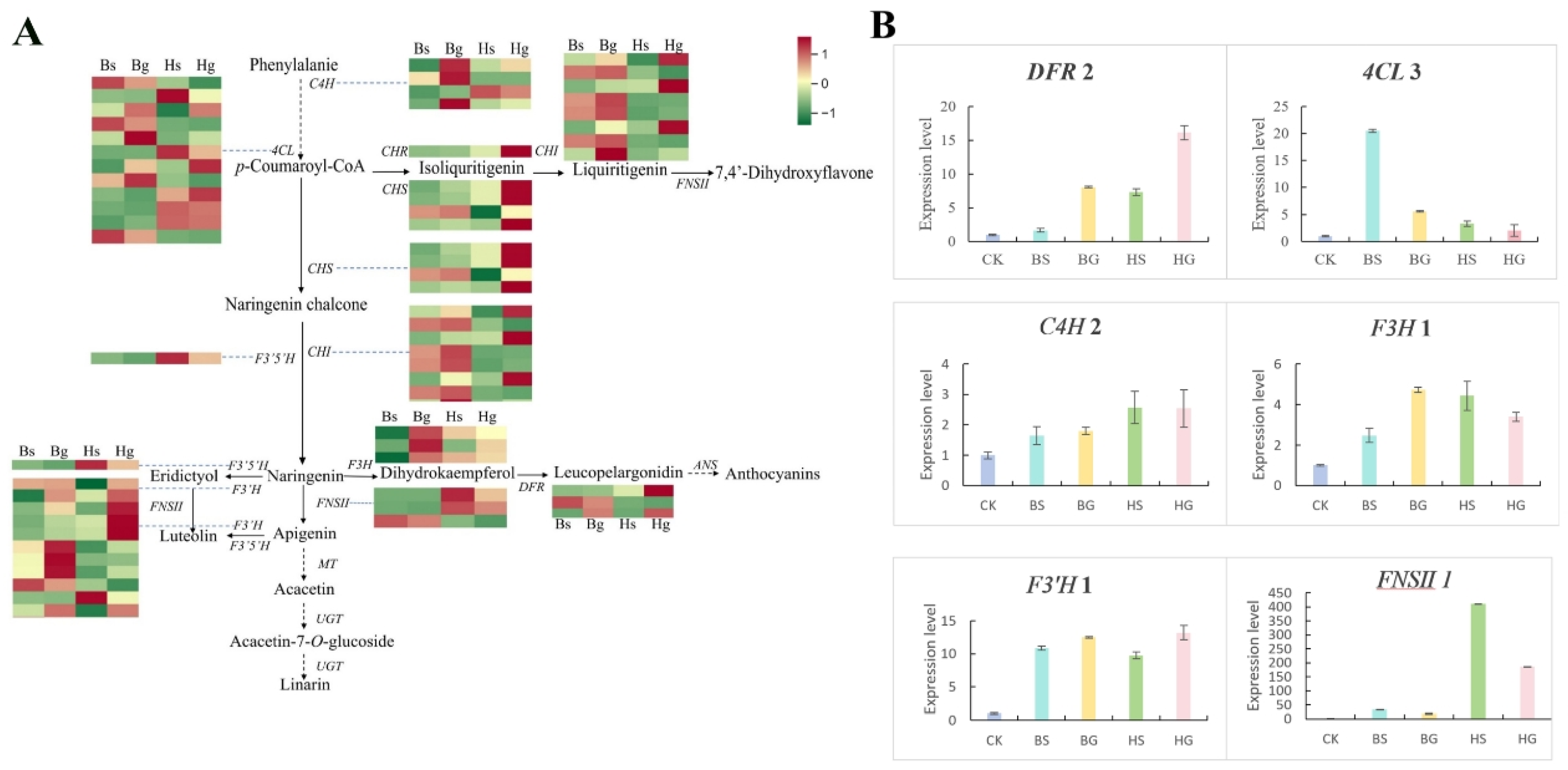
3.5. Verification of Key Enzyme Genes
4. Discussion
4.1. Elucidating Flavonoid Synthesis Pathways in BHYJ and HJ06
4.2. Unraveling the Key Enzyme Genes in Flavonoid Biosynthesis Pathways of C. indicum Through Integrated Transcriptomics and Metabolomics Analysis
4.3. Flower Color Variations from a Medicinal and Eatable Perspective
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Zou, Q.; Guo, Q.; Wang, T.; Chen, J.; Yang, F.; Yang, C. Comparison of metabolome characteristics and screening of chemical markers in Chrysanthemum indicum from different habitats. Physiol. Mol. Biol. Plants 2022, 28, 65–76. [Google Scholar] [CrossRef] [PubMed]
- Song, J.Y.; Xu, J.L.; Miao, J.H.; Xiao, P.G. Research progress in Chrysanthemum indicum L. Mordern Chin. Med. 2020, 22, 1762. [Google Scholar] [CrossRef]
- Editorial Committee of Flora of China. Chinese Academy of Sciences Flora of China; Science Press: Beijing, China, 1999. [Google Scholar]
- Jiang, Y.F.Y.; Duan, X.Y.; Ye, L.X.; Ye, P.; Yang, J.F.; Zhan, R.T.; Chen, W.W.; Ma, D.M. Gene mining and identification of a flavone synthase II involved in flavones biosynthesis by transcriptomic analysis and targeted flavonoid profiling in Chrysanthemum indicum L. Ind. Crops Prod. 2019, 134, 244–256. [Google Scholar] [CrossRef]
- Sheng, H.J.; Guo, Q.S.; Fang, H.L. Analyses on phenolic compositions, abilities of scavenging free radicals and mildewproof of 60 % ethanol extract from Flos Chrysanthemi Indici. JPRE 2010, 19, 20–25. [Google Scholar]
- Zhang, H.J. Study on the Preparation Process of Natural Wild Chrysanthemum Flavor. Ph.D. Dissertation, Shanghai Jiao Tong University, Shanghai, China, 2013. [Google Scholar]
- Deng, Y.; Yang, P.; Zhang, Q.; Wu, Q.W.; Feng, L.F.; Shi, W.J.; Peng, Q.; Ding, L.; Tan, X.K.; Zhan, R.T.; et al. Genomic insights into the evolution of flavonoid biosynthesis and O-methyltransferase and glucosyltransferase in Chrysanthemum indicum. Cell Res. 2020, 43, 13725. [Google Scholar] [CrossRef]
- National Pharmacopoeia Committee. Pharmacopoeia of the People’s Republic of China; China Medical Science Press: Beijing, China, 2020. [Google Scholar]
- Ohmiya, A. Molecular mechanisms underlying the diverse array of petal colors in chrysanthemum flowers. Breed. Sci. 2018, 68, 119–127. [Google Scholar] [CrossRef]
- Han, A.R.; Nam, B.; Kim, B.R.; Lee, K.C.; Song, B.S.; Kim, S.H.; Kim, J.B.; Jin, C.H.; Giampieri, F.; Naughton, D.P. Phytochemical composition and antioxidant activities of two different color Chrysanthemum flower teas. Molecules 2019, 24, 329. [Google Scholar] [CrossRef]
- Li, J.L.; Ye, Z.; Wei, M.; Deng, C.R.; Chi, L.F.; Xu, L.; Han, Z.Z.; Wei, W.F. Evaluation of Chrysanthemi Indici Flos germplasms based on nine bioactive constituents and color parameters. Public Libr. Sci. 2023, 18, e0283498. [Google Scholar] [CrossRef]
- Wang, X.; Zhang, J.; Liu, Z.; Wang, S.B.; Huang, B.S.; Hu, Z.G.; Liu, Y.F. Comparative transcriptome analysis of three chrysanthemums provides insights into flavonoid and terpenoid biosynthesis. J. Plant Biol. 2021, 64, 389–401. [Google Scholar] [CrossRef]
- Li, P.; Huang, Z.; She, Y.; Qin, S.; Gao, W.R.; Cao, Y.N.; Liu, X. An assessment of the interaction for three Chrysanthemum indicum flavonoids and α-amylase by surface plasmon resonance. Food Sci. Nutr. 2019, 8, 620–628. [Google Scholar] [CrossRef] [PubMed]
- Chen, Q.; Kuang, A.C.; Wu, H.H.; Liu, D.; Zhang, X.; Mao, H.Y. Physiological response of CmWRKY15-1 to chrysanthemum white rust based on TRV-VIGS. Genes 2023, 14, 1140596. [Google Scholar] [CrossRef] [PubMed]
- Tian, X.; Wang, H.D.; Chen, L.; Yuan, H.W.; Peng, C.Y.; Wang, W. Distinct Changes in Metabolic Profile and Sensory Quality with Different Varieties of Chrysanthemum (Juhua) Tea Measured by LC-MS-Based Untargeted Metabolomics and Electronic Tongue. Foods 2024, 13, 1080. [Google Scholar] [CrossRef]
- Coskun, O.; Kanter, M.; Korkmaz, A.; Oter, S. Quercetin, a flavonoid antioxidant, prevents and protects streptozotocin-induced oxidative stress and beta-cell damage in rat pancreas. Pharmacol. Res. 2005, 51, 117–123. [Google Scholar] [CrossRef]
- Ren, S.G. Flavonoid composition, antibacterial and antioxidant properties of tartary buckwheat bran extract. Ind. Crops Prod. 2013, 49, 312–317. [Google Scholar] [CrossRef]
- Sobral, F.; Calhelha, R.C.; Barros, L.; Duenas, M.; Tomas, A.; Santos-Buelga, C.; Vilas-Boas, M.; Ferreira, L.C.F.R.; McPhee, D.J. Flavonoid Composition and Antitumor Activity of Bee Bread Collected in Northeast Portugal. Molecules 2017, 22, 248. [Google Scholar] [CrossRef]
- Grassi, D.; Desideri, G.; Croce, G.; Tiberti, S.; Aggio, A.; Ferri, C. Flavonoids, vascular function and cardiovascular protection. Curr. Pharm. Des. 2009, 15, 1072–1084. [Google Scholar] [CrossRef] [PubMed]
- Cha, J.Y.; Cho, Y.S.; Kim, I.; Anno, T.; Rahman, S.M.; Yanagita, T. Effect of hesperetin, a citrus flavonoid, on the liver triacylglycerol content and phosphatidate phosphohydrolase activity in orotic acid-fed rats. Plant Food Hum. Nutr. 2001, 56, 349–358. [Google Scholar] [CrossRef]
- Zhu, L.; Ding, Y.; Wang, S.; Wang, Z.M.; Dai, L.P. Genome-Wide Identification, Characterization, and Expression Analysis of CHS Gene Family Members in Chrysanthemum nankingense. Genes 2022, 13, 2145. [Google Scholar] [CrossRef] [PubMed]
- Kang, K. Anti-Inflammatory Effects of Chrysanthemum indicum Water Extract in RAW 264.7 Cell as a Whole Plant. J. Korean Med. Sci. 2015, 17, 341–347. [Google Scholar] [CrossRef]
- Fan, J.; Huang, J.; Pu, Y.; Niu, Y.J.; Zhang, M.M.; Dai, S.L.; Huang, H. Transcriptomic analysis reveals the formation mechanism of anemone-type flower in chrysanthemum. BMC Genom. 2022, 23, 846. [Google Scholar] [CrossRef]
- Gao, W.; Meng, Q.; Wang, X.; Chen, F.; Zhou, Y.W.; He, M. Overexpression of CiMYC2 Transcription Factor from Chrysanthemum indicum var. aromaticum Resulted in Modified Trichome Formation and Terpenoid Biosynthesis in Transgenic tobacco. J. Plant Growth Regul. 2023, 42, 4161–4175. [Google Scholar] [CrossRef]
- Liao, J.H.; Chen, S.; Liu, D.; Zhang, J.J.; Liu, Y.F. Identification and expression of genome of uridine diphosphate glycosyltransferase (UGT) gene family from Chrysanthemum indicum L. China J. Chin. Mater. Medica 2024, 49, 702–716. [Google Scholar] [CrossRef]
- Su, Y.; Huang, J.; Guo, Q.; Shi, H.; Wei, M.; Wang, C.; Zhao, K.; Bao, T. Combined metabolomic and transcriptomic analysis reveals the characteristics of the lignan in Isatis indigotica Fortune. Gene 2023, 888, 147752. [Google Scholar] [CrossRef] [PubMed]
- Fraga, C.G.; Clowers, B.H.; Moore, R.J.; Zink, E.M. Signature-discovery approach for sample matching of a nerve-agent precursor using liquid chromatography-mass spectrometry, XCMS, and chemometrics. Anal. Chem. 2010, 82, 4165–4173. [Google Scholar] [CrossRef]
- Pisu, D.; Johnston, L.; Mattila, J.T.; Russell, D.G. The frequency of CD38+ alveolar macrophages correlates with early control of M. tuberculosis in the murine lung. Nat. Commun. 2024, 15, 8522. [Google Scholar] [CrossRef]
- Roepke, J.; Jean, T.; Perkel, K.J. Daminozide Alters Anthocyanin Metabolism in Ray Florets of Bronze Chrysanthemum (Chrysanthemum morifolium Ramat.). J. Plant Growth Regul. 2013, 32, 453–460. [Google Scholar] [CrossRef]
- Zhu, L.; Liao, J.; Liu, Y.; Zhou, C.M.; Wang, X.; Hu, Z.G.; Huang, B.S.; Zhang, J.Z. Integrative metabolome and transcriptome analyses reveal the molecular mechanism underlying variation in floral scent during flower development of Chrysanthemum indicum var. aromaticum. Front. Plant Sci. 2022, 13, 919151. [Google Scholar] [CrossRef]
- Shao, Y.; Sun, Y.; Li, D.; Chen, Y. Chrysanthemum indicum L.: A Comprehensive Review of its Botany, Phytochemistry and Pharmacology. Am. J. Chin. Med. 2020, 4, 871–897. [Google Scholar] [CrossRef]
- Uehara, A.; Nakata, M.; Kitajima, J.; Iwashina, T. Internal and external flavonoids from the leaves of Japanese Chrysanthemum species (Asteraceae). Biochem. Syst. Ecol. 2012, 41, 142–149. [Google Scholar] [CrossRef]
- Guo, N.; Zou, Y.P.; Li, H.K.; Kou, P.; Fu, Y. Effective extraction and recovery of linarin from Chrysanthemum indicum L., flower using deep eutectic solvents. Microchem. J. 2020, 159, 105586. [Google Scholar] [CrossRef]
- Huang, L.Q.; Yuan, Y. Molecular Pharmacognosy in Daodi herbs. Chin. J. 2020, 65, 1093–1102. [Google Scholar] [CrossRef]
- Tibaduiza, D.; Anaya, M.; Gómez, J. Electronic Tongues and Noses: A General Overview. Biosensors 2024, 14, 190. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Zhang, Z.J.; Fang, S.Z.; Liu, Y.; Shang, X.L. Integrative analysis of metabolome and transcriptome reveals molecular regulatory mechanism of flavonoid biosynthesis in cyclocarya paliurus under salt stress. Ind. Crops Prod. 2021, 170, 113823. [Google Scholar] [CrossRef]
- Zhang, J.Y.; Long, Y.Q.; Zeng, J.; Fu, X.S.; He, J.W.; Zhou, R.B.; Liu, X.D. Transcriptional regulation mechanism of differential accumulation of flavonoids in different varieties of Lonicera macranthoides based on metabonomics and transcriptomics. Chin. J. Chin. Mater. Med. 2024, 49, 2666–2679. [Google Scholar] [CrossRef]
- Liu, H.; Chen, H.X.; Chen, X.X.; Lu, J.; Chen, D.L.; Luo, C.; Cheng, X.; Huang, C.L. Transcriptomic and metabolomic analyses reveal that MYB transcription factors regulate anthocyanin synthesis and accumulation in the disc florets of the anemone form of Chrysanthemum morifolium. Sci. Hortic. 2023, 307, 110847. [Google Scholar] [CrossRef]
- Chen, S.; Wang, X.; Cheng, Y.; Gao, H.S.; Chen, X.H. A Review of Classification, Biosynthesis, Biological Activities and Potential Applications of Flavonoids. Molecules. 2023, 28, 4982. [Google Scholar] [CrossRef]
- Noda, N.; Aida, R.; Kishimoto, S.; Ishiguro, K.; Fukuchi-Mizutani, M.; Tanaka, Y.; Ohmiya, A. Genetic engineering of novel bluer-colored chrysanthemums produced by accumulation of delphinidin-based anthocyanins. Plant Cell Physiol. 2013, 54, 1684–1695. [Google Scholar] [CrossRef]
- Wang, T.; Zou, Q.J.; Guo, Q.S.; Yang, F.; Wu, L.W.; Zhang, W.Y. Widely Targeted Metabolomics Analysis Reveals the Effect of Flooding Stress on the Synthesis of Flavonoids in Chrysanthemum morifolium. Molecules 2019, 24, 3695. [Google Scholar] [CrossRef]
- Luo, C.; Liu, L.; Zhao, J.; Xu, Y.; Liu, H.; Chen, D.; Cheng, X.; Gao, J.; Hong, B.; Huang, C. CmHY5 functions in apigenin biosynthesis by regulating flavone synthase II expression in chrysanthemum flowers. Planta 2022, 257, 7. [Google Scholar] [CrossRef]
- Sun, W.; Shen, H.; Xu, H.; Tang, X.; Tang, M.; Ju, Z.; Yi, Y. Chalcone Isomerase a Key Enzyme for Anthocyanin Biosynthesis in Ophiorrhiza japonica. Front. Plant Sci. 2019, 10, 865. [Google Scholar] [CrossRef]
- Busche, M.; Acatay, C.; Martens, S.; Weisshaar, B.; Stracke, R. Functional Characterisation of Banana (Musa spp.) 2-Oxoglutarate-Dependent Dioxygenases Involved in Flavonoid Biosynthesis. Front. Plant Sci. 2021, 12, 701780. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.; Wang, X.C.; Liu, Y.; Du, H.; Shu, Q.Y.; Wang, J.W.; Li, S.S.; Wang, L.S. Flavone synthases from Lonicera japonica and L. macranthoides reveal differential flavone accumulation. Sci. Rep. 2016, 6, 19245. [Google Scholar] [CrossRef]
- Ragupathy, S.; Thirugnanasambandam, A.; Henry, T.; Vinayagam, V.; Sneha, R.; Newmaster, S.G. Flower Species Ingredient Verification Using Orthogonal Molecular Methods. Foods 2024, 13, 1862. [Google Scholar] [CrossRef]
- Ma, C.Y. Research on the Breeding Basis of Licorice Varieties and Cloning of HMGR Gene. Ph.D. Dissertation, BUCM, Beijing, China, 2009. [Google Scholar]
- Ni, L.H.; Zhao, L.Z.; Wu, J.R.; Ga, W.; Mi, M. Analysis of transcriptomes to explore genes contributing to iridoid Analysis of transcriptomes to explore genes contributing to iridoid. Acta Pharm. Sin. 2019, 54, 944–953. [Google Scholar] [CrossRef]
- Liu, Y.P.; Wang, Y.; Jia, Z.; Zhang, W.; Zhang, C. Q-Marker prediction analysis of Lonicerae Japonicae Flos based oncolor-characteristic components association analysis and network pharmacology. Chin. Herb. Med. 2024, 55, 4882–4896. [Google Scholar]
- Chen, Y.H.; Cao, X.Y.; Feng, Z.L.; Tan, B.; Hu, Y.; Zhang, H.P.; Jian, Z.H.; Meng, H.J.; Feng, J.C.; Wan, R. Genome-Wide Analysis of the Pomegranate F3’H Family and their Roles in Aril Anthocyanin Biosynthesis. Int. J. Fruit Sci. 2024, 41, 1064–1677. [Google Scholar] [CrossRef]
- Ohmiya, A.; Kishimoto, S.; Aida, R.; Satoshi, K.; Katsuhiko, S. Carotenoid cleavage dioxygenase (CmCCD4a) contributes to white color formation in chrysanthemum petals. Plant Physiol. 2006, 142, 1193–1201. [Google Scholar] [CrossRef]
- Kim, S.J.; Cho, H.I.; Park, J.H.; Kim, J.S.; Kim, Y.H.; Lee, S.K.; HwanKwak, J.; Lee, S.M. Protective effect of linarin against D-galactosamine and lipopolysaccharide-induced fulminant hepatic failure. Eur. J. Pharmacol. 2014, 738, 66–73. [Google Scholar] [CrossRef]
- Li, Y.; Bao, T.; Zhang, J.; Li, H.J.; Shan, X.T.; Yan, H.J.; Kimani, S.; Zhang, L.S.; Gao, X. The coordinated interaction or regulation between floral pigments and volatile organic compounds. Hortic. Plant J. 2024, 11, 463–485. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No. | Q1 (Da) | Q3 (Da) | Molecular Weight (Da) | Formula | Ionization Model | Compounds | CAS | Level |
|---|---|---|---|---|---|---|---|---|
| 1 | 287 | 125 | 288 | C15H12O6 | [M-H]− | 2-Hydroxy-2,3-dihydrogenistein * | - | 2 |
| 2 | 519 | 271 | 518 | C24H22O13 | [M+H]+ | 6″-O-Malonylgenistin | 51011-05-3 | 1 |
| 3 | 431 | 269 | 432 | C21H20O10 | [M-H]− | Genistein-7-O-Glucoside (Genistin) | 529-59-9 | 2 |
| 4 | 285 | 242 | 284 | C16H12O5 | [M+H]+ | Glycitein | 40957-83-3 | 2 |
| 5 | 289 | 163 | 288 | C15H12O6 | [M+H]+ | 3,4,2′,4′,6′-Pentahydroxychalcone | 73692-51-0 | 2 |
| 6 | 313 | 253 | 314 | C17H14O6 | [M-H]− | 3-O-Acetylpinobanksin | 52117-69-8 | 3 |
| 7 | 431 | 311 | 432 | C21H20O10 | [M-H]− | Apigenin-8-C-Glucoside (Vitexin) | 3681-93-4 | 1 |
| 8 | 287 | 135 | 288 | C15H12O6 | [M-H]− | Eriodictyol (5,7,3′,4′-Tetrahydroxyflavanone) | 552-58-9 | 1 |
| 9 | 301 | 164 | 302 | C16H14O6 | [M-H]− | Hesperetin | 520-33-2 | 1 |
| 10 | 465 | 303 | 464 | C22H24O11 | [M+H]+ | Hesperetin-7-O-glucoside * | 31712-49-9 | 3 |
| 11 | 611 | 303 | 610 | C28H34O15 | [M+H]+ | Hesperetin-7-O-neohesperidoside (Neohesperidin) | 13241-33-3 | 3 |
| 12 | 303 | 153 | 302 | C16H14O6 | [M+H]+ | Homoeriodictyol | 446-71-9 | 2 |
| 13 | 273 | 153 | 272 | C15H12O5 | [M+H]+ | Naringenin chalcone; 2′,4,4′,6′-Tetrahydroxychalcone | 73692-50-9 | 3 |
| 14 | 579 | 271 | 580 | C27H32O14 | [M-H]− | Naringenin-7-O-Neohesperidoside (Naringin) * | 10236-47-2 | 3 |
| 15 | 435 | 167 | 436 | C21H24O10 | [M-H]− | Phloretin-2′-O-glucoside (Phlorizin) | 60-81-1 | 3 |
| 16 | 303 | 125 | 304 | C15H12O7 | [M-H]− | Taxifolin (Dihydroquercetin) | 480-18-2 | 3 |
| 17 | 651 | 271 | 650 | C29H30O17 | [M+H]+ | Apigenin-7-O-(2″-O-apiosyl)(6′’-Malonyl)glucoside | - | 3 |
| 18 | 579 | 271 | 578 | C27H30O14 | [M+H]+ | Apigenin-7-O-neohesperidoside (Rhoifolin) * | 17306-46-6 | 1 |
| 19 | 431 | 311 | 432 | C21H20O10 | [M-H]− | Apigenin-8-C-Glucoside (Vitexin) | 3681-93-4 | 1 |
| 20 | 345 | 330 | 344 | C18H16O7 | [M+H]+ | Ayanin (3′,5-Dihydroxy-3,4′,7-Trimethoxyflavone) * | 572-32-7 | 1 |
| 21 | 593 | 285 | 594 | C27H30O15 | [M-H]− | Kaempferol-3-O-rutinoside (Nicotiflorin) | 17650-84-9 | 2 |
| 22 | 773 | 449 | 772 | C33H40O21 | [M+H]+ | Kaempferol-3-O-sophorotrioside | 80714-53-0 | 2 |
| 23 | 449 | 287 | 448 | C21H20O11 | [M+H]+ | Luteolin-7-O-glucoside (Cynaroside) * | 5373-11-5 | 1 |
| 24 | 463 | 287 | 462 | C21H18O12 | [M+H]+ | Luteolin-7-O-glucuronide * | 29741-10-4 | 1 |
| 25 | 595 | 287 | 594 | C27H30O15 | [M+H]+ | Luteolin-7-O-neohesperidoside (Lonicerin) * | 25694-72-8 | 1 |
| 26 | 743 | 449 | 742 | C32H38O20 | [M+H]+ | Quercetin-3-O-(2″-O-Xylosyl)rutinoside | 129235-39-8 | 3 |
| 27 | 549 | 300 | 550 | C24H22O15 | [M-H]− | Quercetin-3-O-(6″-O-malonyl)glucoside | 96862-01-0 | 3 |
| 28 | 611 | 303 | 610 | C27H30O16 | [M+H]+ | Quercetin-3-O-rutinoside (Rutin) * | 153-18-4 | 1 |
| 29 | 611 | 287 | 611 | C30H27O14+ | [M]+ | Cyanidin-3-O-(6″-O-caffeoyl)glucoside | - | 3 |
| 30 | 611 | 303 | 611 | C30H27O14+ | [M]+ | Delphinidin-3-O-(6″-O-p-coumaroyl)glucoside | - | 3 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, C.; Su, Y.; Wei, M.; Guo, Q.; Zou, Q.; Wang, T. Decoding Flavonoid Metabolism for Nutritional Enhancement: A Transcriptome–Metabolome Integration Study of Biosynthesis in Edible Chrysanthemum indicum L. Foods 2025, 14, 1896. https://doi.org/10.3390/foods14111896
Wang C, Su Y, Wei M, Guo Q, Zou Q, Wang T. Decoding Flavonoid Metabolism for Nutritional Enhancement: A Transcriptome–Metabolome Integration Study of Biosynthesis in Edible Chrysanthemum indicum L. Foods. 2025; 14(11):1896. https://doi.org/10.3390/foods14111896
Chicago/Turabian StyleWang, Chengxiang, Yong Su, Min Wei, Qiaosheng Guo, Qingjun Zou, and Tao Wang. 2025. "Decoding Flavonoid Metabolism for Nutritional Enhancement: A Transcriptome–Metabolome Integration Study of Biosynthesis in Edible Chrysanthemum indicum L." Foods 14, no. 11: 1896. https://doi.org/10.3390/foods14111896
APA StyleWang, C., Su, Y., Wei, M., Guo, Q., Zou, Q., & Wang, T. (2025). Decoding Flavonoid Metabolism for Nutritional Enhancement: A Transcriptome–Metabolome Integration Study of Biosynthesis in Edible Chrysanthemum indicum L. Foods, 14(11), 1896. https://doi.org/10.3390/foods14111896