
Effects of Pomelo Peel-Derived Dietary Fibers on Simulated Intestinal Digestion and Fermentation of Fish Balls In Vitro

 , , and
, , and 
Abstract
1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Preparation of Different Pomelo Peel-Derived Dietary Fiber
2.2.1. Preparation of Total Dietary Fiber of Pomelo Peel
2.2.2. Preparation of Pomelo Peel-Derived Cellulose
2.2.3. Preparation of Pomelo Peel-Derived Microcrystalline Cellulose
2.3. Preparation of Fish Balls with Pomelo Peel-Derived Dietary Fiber
2.4. Simulated Gastrointestinal Digestion In Vitro
2.5. Analysis of Gastric Digestive Products
2.5.1. Determination of Protein Digestibility
2.5.2. Analysis of Free Amino Acids in Gastric Digestive Products
2.6. Analysis of Intestinal Digestive Products
2.6.1. Determination of Starch Digestibility
2.6.2. Determination of Relative Contents of Starch Components
2.7. Simulated Colonic Fermentation with Gut Microbiota In Vitro
2.7.1. Determination of pH of Fermentation Products
2.7.2. Determination of Short-Chain Fatty Acids in Fermentation Products
2.7.3. Extraction of DNA and Sequencing of 16S rRNA
2.8. Statistical Analysis
3. Results and Discussion
3.1. Effects of Different Pomelo Peel-Derived Dietary Fibers on Simulated Gastric Digestive Characteristics of Fish Balls In Vitro
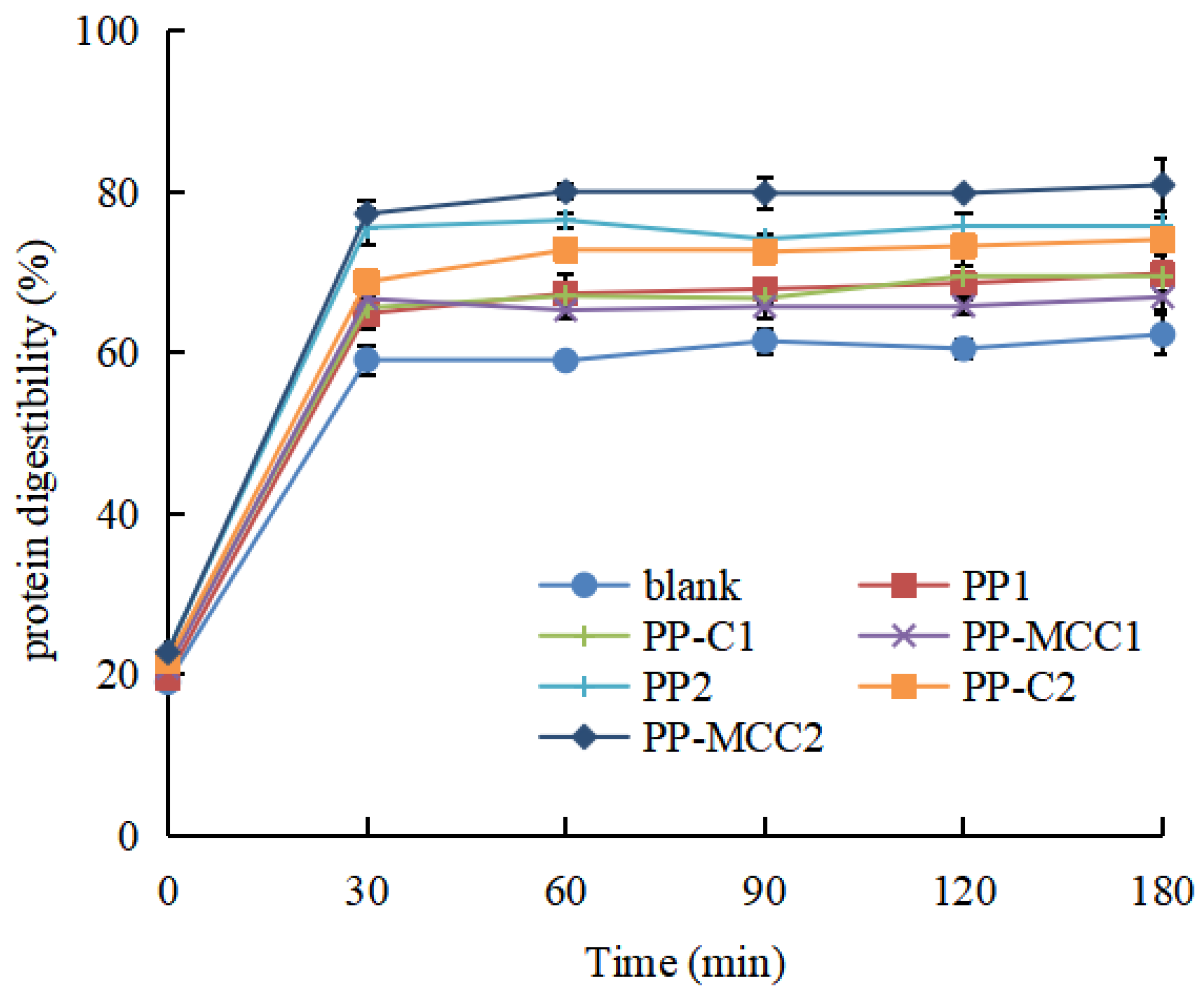
3.1.1. Changes in Protein Digestibility of Fish Balls
3.1.2. Changes in Free Amino Acids in Gastric Digestive Products
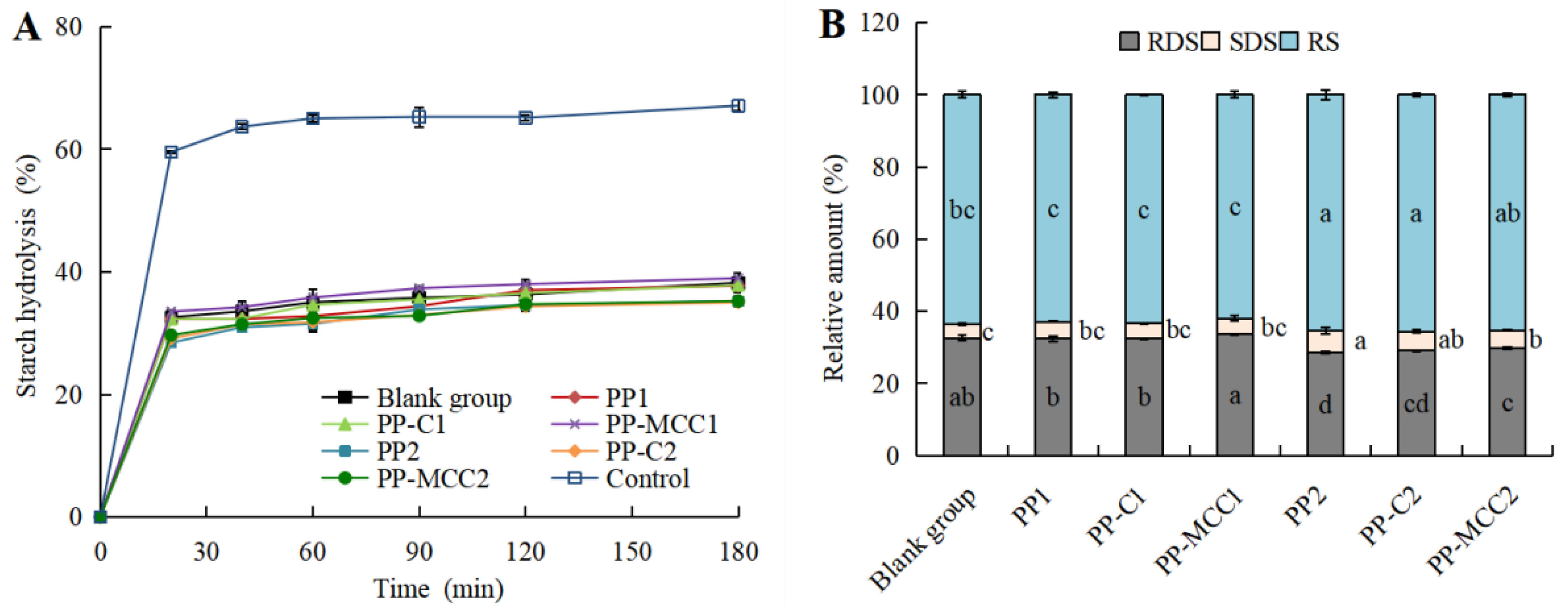
3.2. Effects of Different Pomelo Peel-Derived Dietary Fibers on Simulated Intestinal Digestive Characteristics of Fish Balls In Vitro
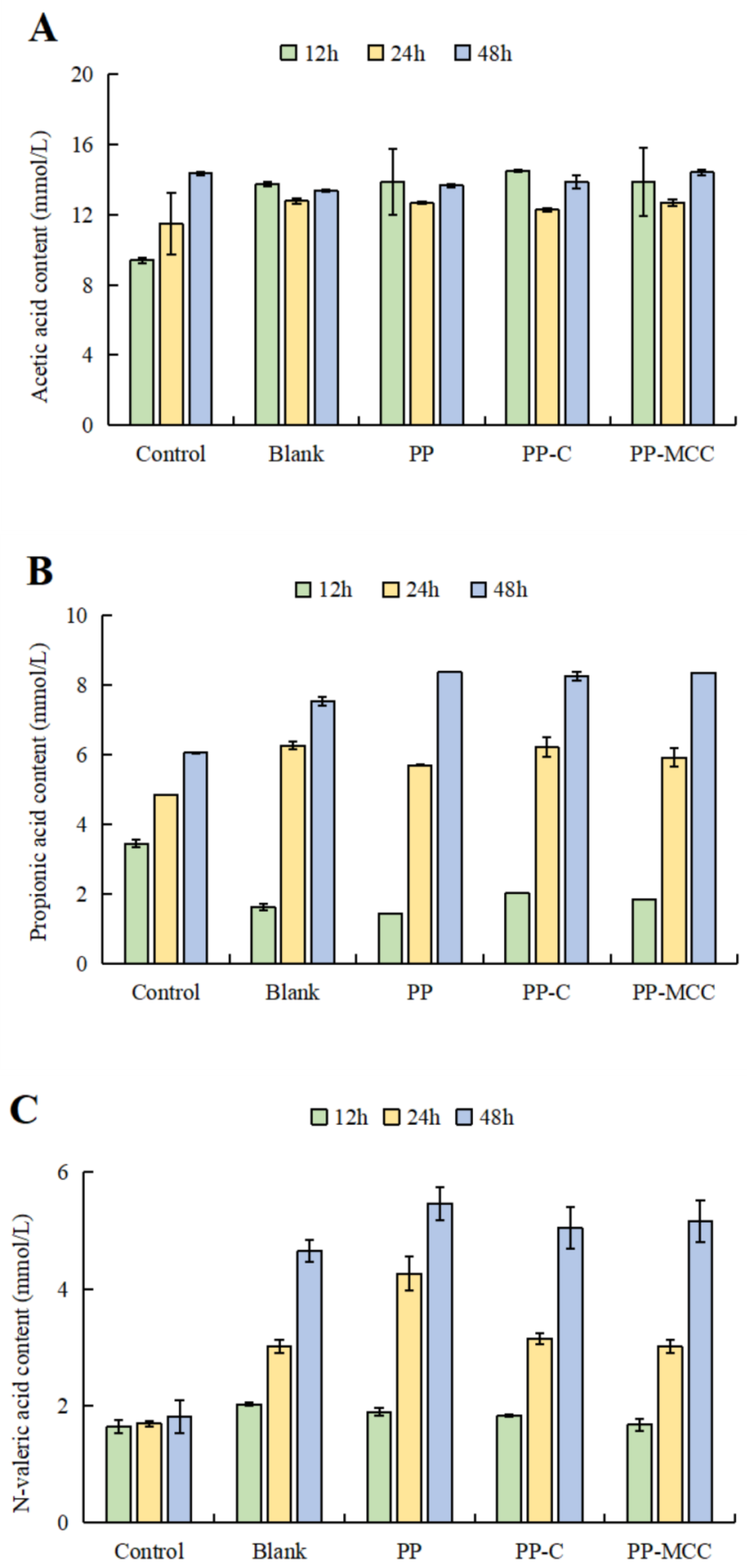
3.3. Effects of Different Pomelo Peel-Derived Dietary Fibers on SCFAs in Fish Balls in In Vitro Simulated Fermentation
3.4. Effects of Different Pomelo Peel-Derived Dietary Fibers on Gut Microbiota Diversity in Fish Balls in In Vitro Simulated Fermentation
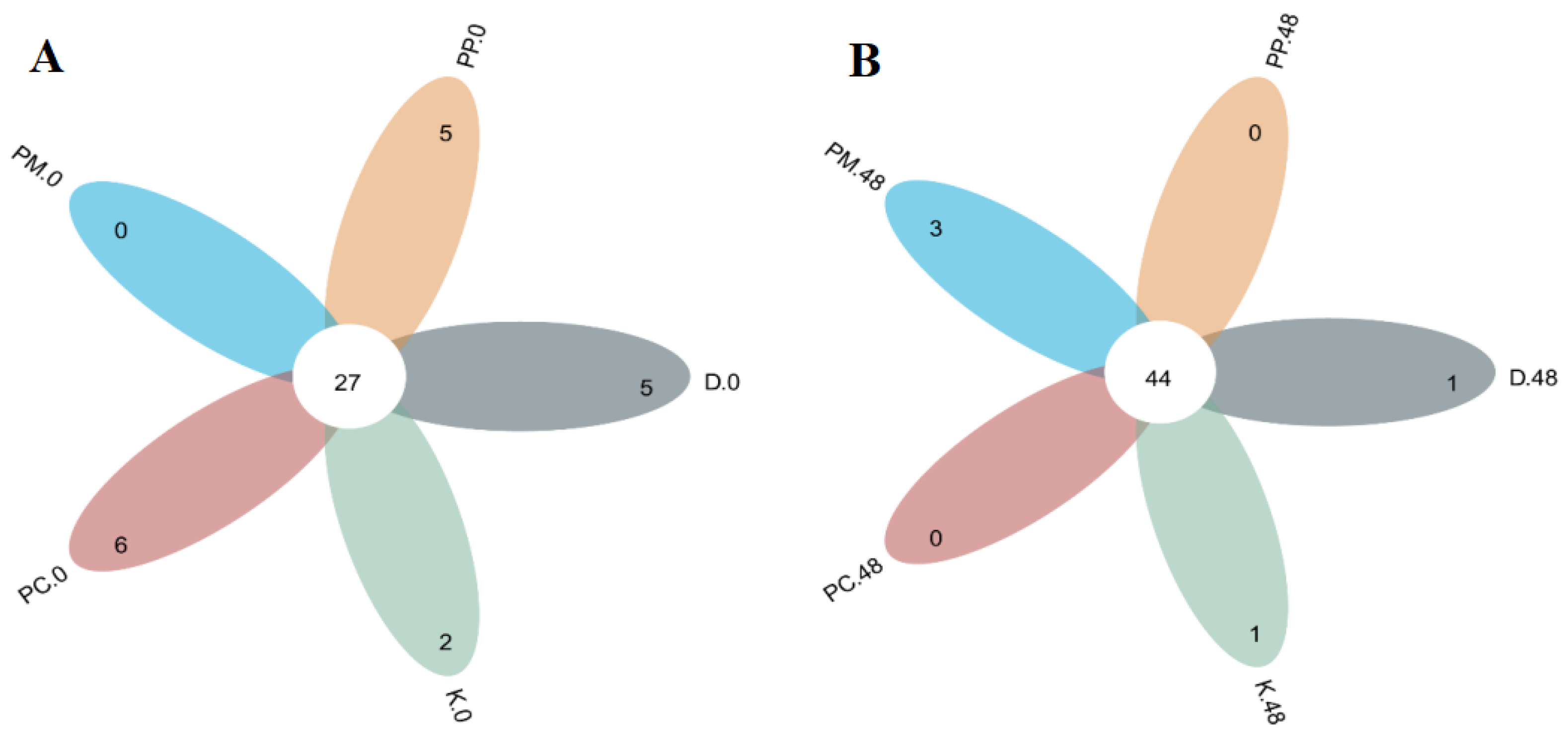
3.4.1. OTU Level of Gut Microbiota
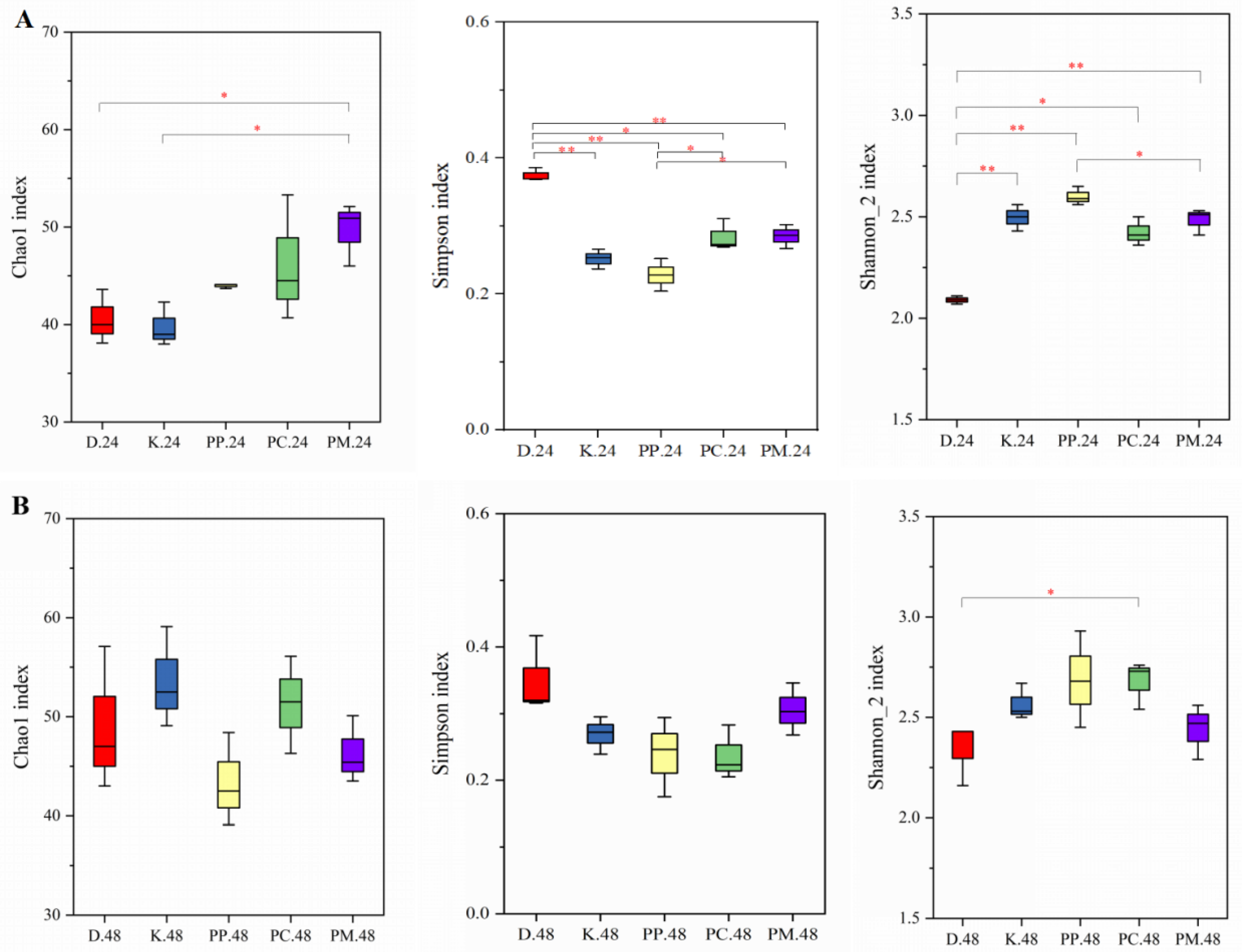
3.4.2. α-Diversity of Gut Microbiota
3.5. Effects of Different Pomelo Peel-Derived Dietary Fibers on Structure of Gut Microbiota During Fish Ball In Vitro Simulated Fermentation
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| PP | Fish balls supplemented with total dietary fiber of pomelo peel |
| PP-C | Fish balls with pomelo peel cellulose |
| PP-MCC | Fish balls with pomelo peel microcrystalline cellulose |
References
- Xiao, L.; Ye, F.; Zhou, Y.; Zhao, G. Utilization of pomelo peels to manufacture value-added products: A review. Food Chem. 2021, 351, 129247. [Google Scholar] [CrossRef]
- Liu, Y.; Liu, A.; Ibrahim, S.A.; Yang, H.; Huang, W. Isolation and characterization of microcrystalline cellulose from pomelo peel. Int. J. Biol. Macromol. 2018, 111, 717–721. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Xu, H.; Yuan, F.; Pan, Q.; Fan, R.; Gao, Y. Physicochemical characterization of five types of citrus dietary fibers. Biocatal. Agric. Biotechnol. 2015, 4, 250–258. [Google Scholar] [CrossRef]
- Zhu, Y. Studies on Dietary Fiber of Citrus Fruits. Master’s Thesis, Zhejiang University, Hangzhou, China, 2003. [Google Scholar]
- Liu, A.L. Preparation of Microcrystalline Cellulose from Pomelo Peel and Application in Emulsified Sausage. Master’s Thesis, Huazhong Agricultural University, Wuhan, China, 2017. [Google Scholar]
- Ni, J.; Shangguan, Y.; Jiang, L.; He, C.; Ma, Y.; Xiong, H. Pomelo peel dietary fiber ameliorates alterations in obesity-related features and gut microbiota dysbiosis in mice fed on a high-fat diet. Food Chem. X 2023, 20, 100993. [Google Scholar] [CrossRef] [PubMed]
- Gao, K.; Liu, Y.; Liu, T.; Song, X.; Ruan, R.; Feng, S.; Wang, X.; Cui, X. OSA improved the stability and applicability of emulsions prepared with enzymatically hydrolyzed pomelo peel insoluble fiber. Food Hydrocoll. 2022, 132, 107806. [Google Scholar] [CrossRef]
- Xu, J.; Zhao, Z.; Liang, P.; Chen, Z.; Cai, G.; Yang, H.; Lu, K.; Lin, J.; Sun, Y. Pomelo (citrus grandis) peel and soybean meal co-fermented protein improved immune response and intestinal health, but not growth performance in largemouth bass (micropterus Salmoides). Aquacult. Rep. 2025, 41, 102660. [Google Scholar] [CrossRef]
- Qi, J.; Song, L.; Zeng, W.; Liao, J. Citrus fiber for the stabilization of O/W emulsion through combination of pickering effect and fiber-based network. Food Chem. 2020, 343, 128523. [Google Scholar] [CrossRef]
- Liu, T.; Wang, K.; Xue, W.; Wang, L.; Zhang, C.; Zhang, X.; Chen, Z. In vitro starch digestibility, edible quality and microstructure of instant rice noodles enriched with rice bran insoluble dietary fiber. Lwt-Food Sci. Technol. 2021, 142, 111008. [Google Scholar] [CrossRef]
- Chen, Y.; Jin, L.; Li, Y.; Xia, G.; Chen, C.; Zhang, Y. Bamboo-shaving polysaccharide protects against high-diet induced obesity and modulates the gut microbiota of mice. J. Funct. Foods 2018, 49, 20–31. [Google Scholar] [CrossRef]
- Fang, T.; Liu, D.; Ning, H.; Liu, D.; Sun, J.; Huang, X.; Dong, Y.; Geng, M.; Yun, S.; Yan, J. Modified citrus pectin inhibited bladder tumor growth through downregulation of galectin-3. Acta Pharmacol. Sin. 2018, 39, 1885–1893. [Google Scholar] [CrossRef]
- Chen, J.; Deng, T.; Wang, C.; Mi, H.; Yi, S.; Li, X.; Li, J. Effect of hydrocolloids on gel properties and protein secondary structure of silver carp surimi. J. Sci. Food Agric. 2020, 100, 2252–2260. [Google Scholar] [CrossRef] [PubMed]
- Zeng, H.; Chen, J.; Zhai, J.; Wang, H.; Xia, W.; Xiong, Y.L. Reduction of the fat content of battered and breaded fish balls during deep-fat frying using fermented bamboo shoot dietary fiber. Lwt-Food Sci. Technol. 2016, 73, 425–431. [Google Scholar] [CrossRef]
- Chen, Y.; Hong, J.; Jiang, Z.; Zhu, Y.; Jiang, Z.; Ni, H.; Zheng, M. Effect of red algae powder on gel properties and in vitro hypolipidemic activity of fish balls. Algal Res. 2023, 74, 103223. [Google Scholar] [CrossRef]
- He, C.; Li, H.; Hong, J.; Xiong, H.; Ni, H.; Zheng, M. Characterization and functionality of cellulose from pomelo fruitlets by different extraction methods. Polymers 2022, 14, 518. [Google Scholar] [CrossRef] [PubMed]
- Brodkorb, A.; Egger, L.; Alminger, M.; Alvito, P.; Assuncao, R.; Ballance, S.; Bohn, T.; Bourlieu-Lacanal, C.; Boutrou, R.; Carriere, F. INFOGEST static in vitro simulation of gastrointestinal food digestion. Nat. Protoc. 2019, 14, 991–1014. [Google Scholar] [CrossRef]
- Zheng, M.; Ye, A.; Singh, H.; Zhang, Y. The in vitro digestion of differently structured starch gels with different amylose contents. Food Hydrocoll. 2021, 116, 106647. [Google Scholar] [CrossRef]
- Minekus, M.; Alminger, M.; Alvito, P.; Ballance, S.; Bohn, T.; Bourlieu, C.; Carriere, F.; Boutrou, R.; Corredig, M.; Dupont, D. A standardised static in vitro digestion method suitable for food—an international consensus. Food Funct. 2014, 5, 1113–1124. [Google Scholar] [CrossRef]
- Xie, J.; Sun, N.; Huang, H.; Xie, J.; Chen, Y.; Hu, X.; Hu, X.; Dong, R.; Yu, Q. Catabolism of polyphenols released from mung bean coat and its effects on gutmicrobiota during in vitro simulated digestion and colonic fermentation. Food Chem. 2022, 396, 133719. [Google Scholar] [CrossRef]
- Cao, C.; Yuan, D.; Kong, B.; Chen, Q.; He, J.; Liu, Q. Effect of different κ-carrageenan incorporation forms on the gel properties and in vitro digestibility of frankfurters. Food Hydrocoll. 2022, 129, 107637. [Google Scholar] [CrossRef]
- Virag, D.; Kiraly, M.; Drahos, L.; Edes, A.E.; Gecse, K.; Bagdy, G.; Juhasz, G.; Antal, I.; Klebovich, I.; Kiss, B.D. Development, validation and application of LC—MS/MS method for quantification of amino acids, kynurenine and serotonin in human plasma. J. Pharm. Biomed. Anal. 2020, 180, 8. [Google Scholar] [CrossRef]
- Fuertig, R.; Ceci, A.; Camus, S.M.; Bezard, E.; Luippold, A.H.; Hengerer, B. LC-MS/MS-based quantification of kynurenine metabolites, tryptophan, monoamines and neopterin in plasma, cerebrospinal fluid and brain. Bioanalysis 2016, 8, 1903–1917. [Google Scholar] [CrossRef] [PubMed]
- Liyanaarachchi, G.V.V.; Mahanama, K.R.R.; Somasiri, H.P.P.S.; Punyasiri, P.A.N. Development and validation of a method for direct, underivatized analysis of free amino acids in rice using liquid chromatography-tandem mass spectrometry. J. Chromatogr. A 2018, 1568, 131–139. [Google Scholar] [CrossRef] [PubMed]
- Thiele, B.; Hupert, M.; Santiago-Schuebel, B.; Oldiges, M.; Hofmann, D. Direct analysis of underivatized amino acids in plant extracts by LC-MS/MS (improved method). Methods Mol. Biol. 2019, 2030, 403–414. [Google Scholar] [CrossRef]
- Englyst, K.N.; Liu, S.; Englyst, H.N. Nutritional characterization and measurement of dietary carbohydrates. Eur. J. Clin. Nutr. 2007, 61, S19–S39. [Google Scholar] [CrossRef]
- Englyst, H.N.; Hudson, G.J. The classification and measurement of dietary carbohydrates. Food Chem. 1996, 57, 15–21. [Google Scholar] [CrossRef]
- Englyst, H.N.; Kingman, S.M.; Cummings, J.H. Classification and measurement of nutritionally important starch fractions. Eur. J. Clin. Nutr. 1992, 462, S33–S50. [Google Scholar]
- López-Barrera, D.M.; Vázquez-Sánchez, K.; Loarca-Piña, M.G.F.; Campos-Vega, R. Spent coffee grounds, an innovative source of colonic fermentable compounds, inhibit inflammatory mediators in vitro. Food Chem. 2016, 212, 282–290. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Hu, G.; Zhu, L.; Zhao, Z.; Yun, J.; Gao, M.; Zhan, X. In vitro digestion and fecal fermentation of highly resistant starch rice and its effect on the gut microbiota. Food Chem. 2021, 361, 130095. [Google Scholar] [CrossRef] [PubMed]
- Kaur, L.; Mao, B.; Beniwal, A.S.; Abhilasha; Kaur, R.; Chian, F.M.; Singh, J. Alternative proteins vs animal proteins: The influence of structure and processing on their gastro-small intestinal digestion. Trends Food Sci. Technol. 2022, 122, 275–286. [Google Scholar] [CrossRef]
- Agyekum, A.K.; Nyachoti, C.M. Nutritional and Metabolic Consequences of feeding high-fiber diets to swine: A Review. Engineering 2017, 3, 716–725. [Google Scholar] [CrossRef]
- Zhang, S.W. Effects of Celery on Gel Quality and in Vitro Digestion Characteristics of Surimi-Based Production. Master’s Thesis, Bohai University, Jinzhou, China, 2019. [Google Scholar]
- Erickson, R.H.; Kim, Y.S. Digestion and absorption of dietary protein. Annu. Rev. Med. 1990, 41, 133–139. [Google Scholar] [CrossRef] [PubMed]
- Barbé, F.; Ménard, O.; Le Gouar, Y.; Buffière, C.; Famelart, M.; Laroche, B.; Le Feunteun, S.; Remond, D.; Dupont, D. Acid and rennet gels exhibit strong differences in the kinetics of milk protein digestion and amino acid bioavailability. Food Chem. 2013, 143, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Igual, M.; Garcia-Segovia, P.; Martinez-Monzo, J. Amino acids release from enriched bread with edible insect or pea protein during in vitro gastrointestinal digestion. Int. J. Gastron. Food Sci. 2021, 24, 100351. [Google Scholar] [CrossRef]
- Graca, C.; Raymundo, A.; Sousa, I.D. Yoghurt and curd cheese addition to wheat bread dough: Impact on in vitro starch digestibility and estimated glycemic index. Food Chem. 2021, 339, 127887. [Google Scholar] [CrossRef]
- Goñi, I.; Garcia-Alonso, A.; Saura-Calixto, F. A starch hydrolysis procedure to estimate glycemic index. Nutr. Res. 1997, 17, 427–437. [Google Scholar] [CrossRef]
- Xu, Z.J. Study on the Quality Properties of Sweet Potato Fish Balls and Its Application. Master’s Thesis, Fujian Agriculture and Forestry University, Fuzhou, China, 2017. [Google Scholar]
- Krishnan, J.G.; Menon, R.; Padmaja, G.; Sajeev, M.S.; Moorthy, S.N. Evaluation of nutritional and physico-mechanical characteristics of dietary fiber-enriched sweet potato pasta. Eur. Food Res. Technol. 2012, 234, 467–476. [Google Scholar] [CrossRef]
- Cui, C.; Li, M.; Ji, N.; Qin, Y.; Shi, R.; Qiao, Y.; Xiong, L.; Dai, L.; Sun, Q. Calcium alginate/curdlan/corn starch @ calcium alginate macrocapsules for slowly digestible and resistant starch. Carbohydr. Polym. 2022, 285, 11. [Google Scholar] [CrossRef]
- Zhou, D.; Ma, Z.; Xu, J.; Li, X.; Hu, X. Resistant starch isolated from enzymatic, physical, and acid treated pea starch: Preparation, structural characteristics, and in vitro bile acid capacity. Lwt-Food Sci. Technol. 2019, 116, 7. [Google Scholar] [CrossRef]
- Kang, J.; Yin, S.; Liu, J.; Li, C.; Wang, N.; Sun, J.; Li, W.; He, J.; Guo, Q.; Cui, S.W. Fermentation models of dietary fibre in vitro and in vivo—A review. Food Hydrocoll. 2022, 131, 17. [Google Scholar] [CrossRef]
- Li, D.; Yao, X.; Yang, Y.; Cao, G.; Yi, G. In vitro digestibility and fermentability profiles of wheat starch modified by chlorogenic acid. Int. J. Biol. Macromol. 2022, 215, 92–101. [Google Scholar] [CrossRef]
- Yao, H.; Wang, L.; Tang, X.; Yang, Z.; Li, H.; Sun, C.; Wu, X.; Xu, D. Two novel polysaccharides from Solanum nigrum L. exert potential prebiotic effects in an in vitro fermentation model. Int. J. Biol. Macromol. 2020, 159, 648–658. [Google Scholar] [CrossRef]
- Perry, R.J.; Peng, L.; Barry, N.A.; Cline, G.W.; Zhang, D.; Cardone, R.L.; Petersen, K.F.; Kibbey, R.G.; Goodman, A.L.; Shulman, G.I. Acetate mediates a microbiome-brain-β-cell axis to promote metabolic syndrome. Nature 2016, 534, 213. [Google Scholar] [CrossRef] [PubMed]
- He, M.; Wu, Y.; Wang, Y.; Hong, M.; Li, T.; Deng, T.; Jiang, Y. Valeric acid suppresses cell wall polysaccharides disassembly to maintain fruit firmness of harvested ‘Waizuili‘ plum (Prunus salicina Lindl). Sci. Hortic. 2022, 291, 7. [Google Scholar] [CrossRef]
- Luu, M.; Pautz, S.; Kohl, V.; Singh, R.; Romero, R.; Lucas, S.; Hofmann, J.; Raifer, H.; Vachharajani, N.; Carrascosa, L.C. The short-chain fatty acid pentanoate suppresses autoimmunity by modulating the metabolic-epigenetic crosstalk in lymphocytes. Nat. Commun. 2019, 10, 760. [Google Scholar] [CrossRef]
- Sowah, S.A.; Riedl, L.; Damms-Machado, A.; Johnson, T.S.; Schübel, R.; Graf, M.; Kartal, E.; Zeller, G.; Schwingshackl, L.; Stangl, G.I. Effects of weight-loss interventions on short-chain fatty acid concentrations in blood and feces of adults: A systematic review. Adv. Nutr. 2019, 10, 673–684. [Google Scholar] [CrossRef] [PubMed]
- Du, J.J.; Zhang, P.W.; Luo, J.; Shen, L.; Zhang, S.; Gu, H.; He, J.; Wang, L.; Zhao, X.; Gan, M.; et al. Dietary betaine prevents obesity through gut microbiota-drived microRNA-378a family. Gut Microbes 2021, 13, 1862612. [Google Scholar] [CrossRef]
- Li, M.; van Esch, B.C.A.M.; Henricks, P.A.J.; Folkerts, G.; Garssen, J. The Anti-inflammatory effects of short chain fatty acids on lipopolysaccharide or tumor necrosis factor α-stimulated endothelial cells via activation of GPR41/43 and inhibition of HDACs. Front. Pharmacol. 2018, 9, 533. [Google Scholar] [CrossRef]
- Bai, X.; Zhang, M.; Zhang, Y.; Zhang, Y.; Huo, R.; Guo, X. In vitro fermentation of pretreated oat bran by human fecal inoculum and impact on microbiota. J. Funct. Foods 2022, 98, 12. [Google Scholar] [CrossRef]
- Kaoutari, A.E.; Armougom, F.; Gordon, J.I.; Raoult, D.; Henrissat, B. The abundance and variety of carbohydrate-active enzymes in the human gut microbiota. Nat. Rev. Microbiol. 2013, 11, 497–504. [Google Scholar] [CrossRef]
- Sassone-Corsi, M.; Nuccio, S.; Liu, H.; Hernandez, D.; Vu, C.T.; Takahashi, A.A.; Edwards, R.A.; Raffatellu, M. Microcins mediate competition among Enterobacteriaceae in the inflamed gut. Nature 2016, 540, 280. [Google Scholar] [CrossRef]
- Yang, M.; Ashraf, J.; Tong, L.; Wang, L.; Zhang, X.; Li, N.; Zhou, S.; Liu, L. Effects of Rhizopus oryzae and Aspergillus oryzae on prebiotic potentials of rice bran pretreated with superheated steam in an in vitro fermentation system. Lwt-Food Sci. Technol. 2021, 139, 6. [Google Scholar] [CrossRef]
- Karboune, S.; Davis, E.J.; Fliss, I.; Andreani, E.S. In-vitro digestion and fermentation of cranberry extracts rich in cell wall oligo/polysaccharides. J. Funct. Foods 2022, 92, 11. [Google Scholar] [CrossRef]
- Zhang, Q.; Li, J.; Wang, G.; Wang, L.; Zhang, Z.; Fang, Z.; Lin, Y.; Xu, S.; Feng, B.; Zhuo, Y.; et al. The replacement of bacitracin methylene disalicylate with Bacillus subtilis PB6 in the diet of male Cherry Valley Ducks reduces the feed conversion ratio by improving intestinal health and modulating gut microbiota. Poult. Sci. 2022, 101, 13. [Google Scholar] [CrossRef] [PubMed]
- Xie, Z.; Ding, L.; Huang, Q.; Fu, X.; Liu, F.; Dhital, S.; Zhang, B. In vitro colonic fermentation profiles and microbial responses of propionylated high-amylose maize starch by individual Bacteroides-dominated enterotype inocula. Food Res. Int. 2021, 144, 8. [Google Scholar] [CrossRef]
- Ortega-Hernández, A.; Martínez-Martínez, E.; Gómez-Gordo, R.; López-Andrés, N.; Fernández-Celis, A.; Gutiérrrez-Miranda, B.; Nieto, M.L.; Alarcón, T.; Alba, C.; Gómez-Garre, D. The interaction between mitochondrial oxidative stress and gut microbiota in the cardiometabolic consequences in diet-induced obese rats. Antioxidants 2020, 9, 640. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Content (μg/mL) | Blank Group | PP1 | PP-C1 | PP-MCC1 | PP2 | PP-C2 | PP-MCC2 | |
|---|---|---|---|---|---|---|---|---|
| Special amino acids | GABA | 0.76 ± 0.00 d | 0.86 ± 0.00 b | 0.80 ± 0.01 cd | 0.82 ± 0.01 bc | 0.94 ± 0.05 a | 0.81 ± 0.02 cd | 0.83 ± 0.01 bc |
| Orn | 15.22 ± 0.36 c | 15.82 ± 0.38 bc | 16.11 ± 0.31 ab | 15.77 ± 0.10 bc | 16.65 ± 0.57 a | 15.83 ± 0.25 bc | 15.96 ± 0.34 abc | |
| Hcy | 0.22 ± 0.03 ab | 0.22 ± 0.01 ab | 0.18 ± 0.01 c | 0.18 ± 0.01 bc | 0.18 ± 0.03 c | 0.19 ± 0.00 bc | 0.24 ± 0.02 a | |
| Non-essential amino acids | Gly | 43.93 ± 0.30 d | 48.24 ± 0.62 bc | 42.71 ± 0.62 d | 49.22 ± 0.19 ab | 51.01 ± 2.20 a | 48.37 ± 0.43 bc | 46.86 ± 1.09 c |
| Ala | 62.27 ± 0.07 c | 66.52 ± 1.87 b | 67.21 ± 1.67 b | 68.15 ± 0.53 ab | 72.02 ± 3.71 a | 67.84 ± 1.72 b | 68.56 ± 0.19 ab | |
| Ser | 28.33 ± 0.53 c | 29.63 ± 0.76 bc | 30.05 ± 0.41 b | 30.14 ± 0.78 b | 31.99 ± 0.99 a | 30.72 ± 0.12 ab | 30.13 ± 0.38 b | |
| Pro | 15.01 ± 0.17 c | 15.79 ± 0.40 bc | 16.29 ± 0.21 ab | 16.43 ± 0.00 ab | 17.02 ± 0.86 a | 15.99 ± 0.07 b | 16.10 ± 0.15 b | |
| Asn | 42.35 ± 3.23 de | 53.33 ± 3.74 ab | 44.01 ± 1.01 cd | 56.22 ± 1.42 ab | 50.30 ± 3.82 bc | 58.27 ± 4.61 a | 36.18 ± 2.52 e | |
| Asp | 37.20 ± 0.04 c | 40.42 ± 0.93 b | 40.53 ± 1.60 b | 39.45 ± 0.40 bc | 45.31 ± 2.75 a | 40.67 ± 0.91 b | 40.18 ± 0.20 b | |
| Gln | 8.68 ± 0.03 a | 8.92 ± 0.07 a | 9.21 ± 0.00 a | 8.82 ± 0.34 a | 8.72 ± 0.50 a | 8.96 ± 0.23 a | 9.00 ± 0.04 a | |
| Glu | 46.95 ± 1.24 c | 51.54 ± 0.31 ab | 50.10 ± 1.59 bc | 51.83 ± 0.14 ab | 54.27 ± 2.86 a | 51.69 ± 1.77 ab | 51.88 ± 1.05 ab | |
| Arg | 19.83 ± 0.26 b | 20.65 ± 0.15 b | 20.94 ± 0.14 b | 20.24 ± 0.06 b | 22.54 ± 1.73 a | 20.64 ± 0.60 b | 20.29 ± 0.39 b | |
| Tyr | 47.96 ± 0.81 b | 47.63 ± 0.87 b | 49.01 ± 1.05 ab | 48.28 ± 0.61 ab | 52.03 ± 4.40 a | 47.74 ± 0.66 b | 46.74 ± 0.21 b | |
| Essential amino acid | Val | 47.95 ± 0.97 c | 53.22 ± 1.13 b | 54.09 ± 0.60 b | 53.99 ± 0.13 b | 58.33 ± 1.87 a | 54.86 ± 0.77 b | 53.33 ± 2.49 b |
| Thr | 27.80 ± 0.18 c | 30.79 ± 0.06 ab | 30.22 ± 0.01 b | 30.46 ± 0.22 b | 31.52 ± 0.73 a | 30.89 ± 0.77 ab | 30.37 ± 0.08 b | |
| Ile | 42.23 ± 0.10 c | 44.52 ± 0.67 bc | 45.29 ± 1.33 abc | 46.77 ± 0.02 ab | 48.29 ± 2.75 a | 47.05 ± 2.11 ab | 45.92 ± 0.36 ab | |
| Leu | 56.91 ± 0.56 b | 60.21 ± 1.22 ab | 59.98 ± 2.77 ab | 59.72 ± 0.46 ab | 62.26 ± 2.51 a | 58.89 ± 1.90 ab | 58.75 ± 0.74 ab | |
| Lys | 38.10 ± 0.09 b | 40.51 ± 1.15 a | 41.35 ± 0.81 a | 40.84 ± 0.05 a | 42.14 ± 1.66 a | 40.59 ± 0.63 a | 40.21 ± 1.34 ab | |
| Met | 34.42 ± 0.05 b | 35.94 ± 1.83 ab | 35.91 ± 0.84 ab | 35.97 ± 1.09 ab | 37.71 ± 1.48 a | 38.38 ± 0.00 a | 36.00 ± 1.51 ab | |
| Phe | 150.37 ± 0.19 d | 159.75 ± 2.59 abc | 161.19 ± 4.01 abc | 156.25 ± 1.19 cd | 166.67 ± 8.04 a | 165.79 ± 2.87 ab | 157.48 ± 4.05 bcd | |
| Trp | 20.54 ± 0.05 bc | 20.92 ± 0.37 abc | 20.85 ± 0.75 abc | 21.59 ± 0.27 ab | 21.90 ± 1.25 a | 20.80 ± 0.05 abc | 20.28 ± 0.20 c | |
| His | 7.81 ± 0.07 b | 8.26 ± 0.07 ab | 8.35 ± 0.14 a | 8.10 ± 0.21 ab | 8.51 ± 0.54 a | 8.22 ± 0.11 ab | 8.09 ± 0.13 ab | |
| Total essential amino acid | 426.12 ± 0.06 c | 454.11 ± 8.83 b | 457.24 ± 9.26 ab | 453.70 ± 2.03 b | 477.33 ± 20.82 a | 465.47 ± 9.21 ab | 450.43 ± 10.02 b | |
| Total free amino acid | 794.83 ± 4.52 c | 853.69 ± 17.54 b | 844.40 ± 15.78 b | 859.26 ± 5.54 ab | 900.31 ± 37.66 a | 873.18 ± 18.30 ab | 833.39 ± 16.12 bc | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zheng, M.; Wei, Y.; Hong, J.; Li, Z.; Zhu, Y.; Hong, T.; Jiang, Z.; Ni, H. Effects of Pomelo Peel-Derived Dietary Fibers on Simulated Intestinal Digestion and Fermentation of Fish Balls In Vitro. Foods 2025, 14, 1818. https://doi.org/10.3390/foods14101818
Zheng M, Wei Y, Hong J, Li Z, Zhu Y, Hong T, Jiang Z, Ni H. Effects of Pomelo Peel-Derived Dietary Fibers on Simulated Intestinal Digestion and Fermentation of Fish Balls In Vitro. Foods. 2025; 14(10):1818. https://doi.org/10.3390/foods14101818
Chicago/Turabian StyleZheng, Mingjing, Yiman Wei, Jinling Hong, Zhipeng Li, Yanbing Zhu, Tao Hong, Zedong Jiang, and Hui Ni. 2025. "Effects of Pomelo Peel-Derived Dietary Fibers on Simulated Intestinal Digestion and Fermentation of Fish Balls In Vitro" Foods 14, no. 10: 1818. https://doi.org/10.3390/foods14101818
APA StyleZheng, M., Wei, Y., Hong, J., Li, Z., Zhu, Y., Hong, T., Jiang, Z., & Ni, H. (2025). Effects of Pomelo Peel-Derived Dietary Fibers on Simulated Intestinal Digestion and Fermentation of Fish Balls In Vitro. Foods, 14(10), 1818. https://doi.org/10.3390/foods14101818