Abstract
Yersiniosis, one of the leading foodborne infections in the European Union, is caused by Yersinia enterocolitica. In this study, the antibacterial and antibiofilm effects of cinnamon (Cinnamomum zeylanicum Nees), clove (Syzygium aromaticum L.), oregano (Origanum vulgare L.), rosemary (Rosmarinus officinalis L.), thyme (Thymus vulgaris L.), and winter savory (Satureja montana L.) essential oils were investigated against Y. enterocolitica strains belonging to the bioserotype 4/O:3. Cinnamon essential oil showed the highest antibacterial activity, with an MIC value 0.09 µL/mL, followed by oregano and thyme essential oils, with MIC values from 0.09 to 0.18 µL/mL, and from 0.18 to 0.23 µL/mL, respectively. Thyme essential oil at 0.23 µL/g (MIC) and at 0.46 µL/g (2MIC) significantly (p < 0.05) reduced the number of Y. enterocolitica by 0.38 log CFU/g and 0.64 log CFU/g, respectively, in minced pork meat during storage at 4 °C for 4 days. The Y. enterocolitica strains formed biofilms at 15 °C and 37 °C in tryptic soy broth and Luria–Bertani broth, while no biofilms were obtained at 5 °C, and in meat broth nutrient media. Applying the minimum bactericidal concentrations of cinnamon, clove, oregano, rosemary, thyme, and winter savory essential oils on preformed biofilms led to significant reductions being observed in the range from 45.34% to 78.89%. A scanning electron microscopy assay showed the devastating impact of oregano and thyme essential oils on the morphology of Y. enterocolitica bacterial cells. In conclusion, the results of this study show that essential oils possess high anti-Yersinia and antibiofilm effects.
1. Introduction
Currently, the genus Yersinia consists of 28 species (Y. aldovae, Y. aleksiciae, Y. alsatica, Y. artesiana, Y. bercovieri, Y. canariae, Y. enterocolitica, Y. entomophaga, Y. frederiksenii, Y. hibernica, Y. intermedia, Y. kristensenii, Y. massiliensis, Y. mollaretii, Y. nurmii, Y. occitanica, Y. pekkanenii, Y. pestis, Y. philomiragia, Y. proxima, Y. rochesterensis, Y. pseudotuberculosis, Y. rohdei, Y. ruckeri, Y. similis, Y. thracica, Y. vastinensis, and Y. wautersii) [1]. Three species of Yersinia are human pathogens, including Y. pestis, the causative agent of plague, and Y. enterocolitica and Y. pseudotuberculosis, the enteropathogen species [2,3]. Y. enterocolitica is divided into six biotypes, and into over 70 serotypes [4,5]. According to pathogenicity, biotype 1A is non-pathogenic, biotypes 2, 3, 4, and 5 are low-pathogenic, while the biotype 1B is highly pathogenic [3,6]. This facultative anaerobic, Gram-negative bacterium may cause human infections with various symptoms, including fever, abdominal pain, diarrhea, ileitis, pseudoappendicitis, mesenteric lymphadenitis, arthritis, septicemia, and mortality [7,8].
Y. enterocolitica, the subject of our study, is transmitted to humans as a foodborne or waterborne pathogen [4,5]. Although yersiniosis occurs after the consumption of various contaminated food of both animal and plant origin, contaminated pork is the main source of Y. enterocolitica in European countries [5]. Pigs are considered the main asymptomatic reservoir of Y. enterocolitica, especially bioserotype 4/O:3 [9]. The prevalence in Europe is estimated to be up to 93% [10]. The prevalence of Y. enterocolitica in pigs varies worldwide [11,12], and depends on several factors including the farm management system, conditions in slaughterhouses, and the detection methodology [13]. During slaughter, Y. enterocolitica from tonsils and the intestine can contaminate carcasses and work surfaces at slaughterhouses. This can lead to the cross-contamination of the meat. A low number of Y. enterocolitica in minced meat is characterized as a high microbiological risk factor for consumers [14], because Y. enterocolitica can survive and replicate at refrigerated temperatures [9].
One of the survival strategies of bacterial pathogens is the ability to form biofilms [15]. In the food industry, biofilms are mainly responsible for microbial contamination. Biofilms are complex three-dimensional communities of surface-attached bacteria protected by an extracellular matrix. Here, bacterial cells communicate through a signaling mechanism known as quorum sensing (QS) [15]. Y. enterocolitica are able to form biofilms through five stages of activating the QS system [16]. In the first stage, bacterial cells adhere to a surface. Next, extracellular polymeric substances (EPSs) are made, and bacterial cells become irreversibly attached. The EPSs form a complex organic polymer matrix which consists of polysaccharides, proteins, lipids, nucleic acid, and other substances [17]. In the third stage, the biofilm architecture is formed, and the biofilm becomes mature. The maximum bacterial cell density is reached at the next stage. After that, the biofilm releases bacterial cells that can attach and contaminate new surfaces and form new biofilms [15]. Insufficient and ineffective sanitation allows the biofilm to persist on food-contacting surfaces, presenting a constant source of microbial contamination of food. Contaminated food may cause foodborne diseases, which are, along with antimicrobial resistance, a global public health problem [18]. Further, biofilms can damage equipment and cause food spoilage all leading to increased costs in the food sector [16,19].
Although Y. enterocolitica infection is mainly asymptomatic, antibiotics are often used as a conventional method of treatment. The World Health Organization suggests the use of chloramphenicol, gentamicin, cotrimoxazole, tetracyclines, fluoroquinolones, and third-generation cephalosporins. Yet, antibiotic resistance has been observed [20]. Because of this, it is important to find an adequate treatment strategy against Y. enterocolitica biofilms, and against the Y. enterocolitica present on meat.
One of the measures against Y. enterocolitica may be natural volatile liquids, named as essential oils (EOs). EOs are complex mixtures of secondary metabolites originating from aromatic and/or medicinal plants with wide biological activities [21]. They are obtained from different parts of aromatic plants (oregano, rosemary, thyme, sage, lavender, basil, and many others) [22,23], including roots, leaves, flowers, fruits, seeds, by extraction methods, of which hydrodistillation is the most common [24]. A wide range of EO biological activities have been thoroughly examined, including antimicrobial, antiviral, antifungal, anticancer, antioxidant, and anti-inflammatory activity [24,25]. The corresponding mechanisms of action are many, and they have been previously reported [24]. Different EOs show strong antibacterial activity against many foodborne pathogens, including Salmonella Enteritidis, Salmonella Typhimurium, Listeria monocytogenes, Campylobacter jejuni, Camplylobacter coli, Escherichia coli, Staphylococcus aureus, Bacillus subtilis, Bacillus cereus, and others [21]. The chemical composition of EOs is responsible for their antimicrobial properties [25]. According to Durofil et al. [20], there are some promising data about EOs’ activity against Y. enterocolitica. In the last two decades, extensive investigations have involved EOs of Ocimum basilicum, Origanum vulgare, Rosmarinus officinalis, and Thymus vulgaris. Articles have shown several minimum inhibitory concentration (MIC) values of the Cinnamomum zeylanicum (17.5 µL/L, 75 µg/mL), Syzygium aromaticum (8.7 µL/L), Origanum vulgare (75 µg/mL, 20 µL/mL, 2.5 µL/mL, 0.6 mg/mL, 4.4 µL/L), Rosmarinus officinalis (0.075 mg/mL, 20 µL/mL, 8 µg/mL), Thymus vulgaris (1.2 mg/mL, 32 µg/mL, 34.9 µL/L, <0.2 µL/mL), and Satureja montana (0.32%) EOs against Y. enterocolitica, all depending on their origin, plant part, chemical composition, and performed assay [20]. However, in addition to in vitro studies, EOs deserve further research in terms of their effect on Y. enterocolitica.
To the best of our knowledge, no research has examined the reduction in Y. enterocolitica biofilms by cinnamon (Cinnamomum zeylanicum Nees), clove (Syzygium aromaticum L.), oregano (Origanum vulgare L.), rosemary (Rosmarinus officinalis L.), thyme (Thymus vulgaris L.), and winter savory (Satureja montana L.). The novelty of this research lies in the different approach of measuring the antimicrobial and antibiofilm activity of six commercially available EOs against Y. enterocolitica 4/O:3. Withal, all Y. enterocolitica strains included in this research were isolated from a slaughterhouse located in Serbia. Additionally, the winter savory EO was obtained from organic plants cultivated in Southeast Serbia, making it a locally available option for the Y. enterocolitica control. The aims of this research were (I) to determine the antibacterial activity of selected EOs against Y. enterocolitica in minced pork meat, (II) to determine the ability of Y. enterocolitica strains to form biofilms in different nutrient media and temperature conditions, and (III) to investigate the influence of the selected EOs on the formed Y. enterocolitica biofilms.
2. Materials and Methods
2.1. Essential Oils
Essential oils of cinnamon (Cinnamomum zeylanicum Nees, Sri Lanka) (CIEO), clove (Syzygium aromaticum L., India) (CLEO), oregano (Origanum vulgare L., India) (OREO), rosemary (Rosmarinus officinalis L., Spain) (ROEO), thyme (Thymus vulgaris L., India) (THEO), and winter savory (Satureja montana L., Serbia) (WSEO) were selected for this experiment. All EOs were commercially available from Terra Co, Novi Sad, Serbia, and Siempreviva oils, Niš, Serbia. For identification of EO compounds, a gas chromatograph GC 7890B coupled with an MS 5977A mass spectrometer (Agilent Technologies, Santa Clara, CA, USA) was used, as in the previous study by Vidaković Knežević et al. [26].
2.2. Bacterial Strains
Three Y. enterocolitica strains (Y4/1, Y9, and Y14) were previously isolated from pig tonsils and belonged to the bioserotype 4/O:3 [27]. Until examination, strains were stored frozen in tryptic soy broth (TSB) (Oxoid, UK) with the addition of 20% glycerol at −80 °C.
2.3. Determination of Minimum Inhibitory Concentration (MIC) and Minimum Bactericidal Concentration (MBC)
The EOs were prepared in DMSO (Lach-Ner SRO, Prague, Czech Republic). The broth microdilution method reported by Kocić-Tanackov et al. [28] was used to determine the MICs and MBCs of the EOs. Briefly, 100 µL of EO was mixed with 100 µL of Muller–Hinton broth (Oxoid, Basingstoke, UK) in the first well of a microtiter plate. After that, a 1:1 serial dilution was made, reaching the concentration 0.23 µL/mL. All wells were filled with 10 µL Y. enterocolitica suspension (108 CFU/mL). Incubation was maintained at 37 °C for 24 h. Then, the content of each well was inoculated onto Muller–Hinton agar (Biokar Diagnostic, Beauvais, France) and incubated at 37 °C for 24 h. The lowest concentrations of EOs that inhibited the visible growth of Y. enterocolitica were defined as MICs, while the lowest concentrations of EOs with no growth after subculturing onto Muller–Hinton agar were defined as MBCs.
2.4. Preparation of Minced Meat
The fresh pork Quadriceps femoris was minced in a sterile grinder with a No. 4 disc, and portions of 10 g were placed into sterile Petri dishes. The minced pork meat samples were inoculated with ca. 104–105 CFU Y. enterocolitica strain Y4/1. Then, the samples were treated with MIC and 2MIC values of OREO (0.09 µL/g and 0.18 µL/g), and THEO (0.23 µL/g and 0.46 µL/g). All samples were homogenized using a sterile glass rod, and stored in Petri dishes for 4 days at 4 ± 1 °C.
2.5. Bacterial Enumeration
Y. enterocolitica count was performed by adding all 10 g of minced pork meat sample into a stomacher bag containing 90 mL sterilized peptone water (Biokar Diagnostics, Beauvais, France). After homogenization, decimal dilution was performed followed by spread-plating on CIN agar (Yersinia selective agar base CM0653, with Yersinia Selective Supplement SR0109, Oxoid, UK), and incubation at 30 ± 1 °C for 24 h. The suspect colonies, small (≤1 mm), smooth, with a red center and translucent rim, were further examined according to the standard method [29].
2.6. Biofilm Formation
The capability of Y. enterocolitica strains to produce biofilms in TSB, meat broth (MB) (Oxoid, UK), and Luria–Bertani broth (LB) (Oxoid, UK) at three different temperatures, including 5 °C, 15 °C, and 37 °C for 48 h was tested according to a previously reported protocol [30]. Briefly, Y. enterocolitica strains were subcultured in TSB overnight at 37 °C, and then diluted in TSB, MB, and LB at the ratio of 1:40. Aliquots of 200 µL were inoculated into wells of a sterile 96-well microtiter plate with flat bottoms (Sarstedt, Nimbrecht, Germany) and incubated at different temperatures for 48 h. After incubation, the Y. enterocolitica cells that did not adhere were discarded, rinsed with physiological saline (3 × 250 μL/well), and air-dried. Following this, fixation (250 μL/well of 96% ethanol for 20 min), staining (250 μL of 0.3% crystal violet (Fluka, Sigma-Aldrich, Germany) for 20 min), rinsing (tapped water), air drying, and recording the optical density at 550 nm (OD550) on the ASYS Expert Plus Microtitration Reader (Biochrom, Cambridge, UK) were performed. All Y. enterocolitica strains were classified as non-biofilm formers (OD ≤ ODC), weak biofilm formers (ODC ≤ OD ≤ (2 × ODC)), moderate biofilm formers ((2 × ODC) < OD ≤ (4 × ODC)), or strong biofilm formers (OD > (4 × ODC)) [31].
2.7. Reduction in Biofilm
The reduction in biofilms by EOs was performed according to a previously reported methodology [30]. Following the procedure from Section 2.6, the biofilms of Y. enterocolitica strains were formed. After discarding and washing non-adherent Y. enterocolitica cells, the adhered biofilm was treated with an aliquot of 200 µL solution of MBC concentrations of EOs in TSB for 48 h at 15 °C and 37 °C. As in the biofilm formation procedure, after washing, the remained biomass was stained with 0.3% crystal violet. OD550 of remaining biofilms were measured, and the reductions were calculated following Equation (1):
Biofilm reduction (%) = [(ODGROWTH CONTROL − ODSAMPLE)/ODGROWTH CONTROL] × 100,
2.8. Scanning Electron Microscopy (SEM) Assay
Suspensions of Y. enterocolitica strain Y9 (100 μL) were inoculated on the surfaces of sterile SS 304 stainless steel coupons (10 × 10 × 1 mm) for 3 h at 37 °C, and placed in a 12-well plate. Then, the SS 304 stainless steel coupons were rinsed with 3 mL of physiological saline, treated with 2 mL LB broth (control), MIC of OREO (0.18 µL/mL), and MIC of THEO (0.45 µL/mL), and incubated at 37 °C for 24 h. After incubation, the SS 304 stainless steel coupons were rinsed with physiological saline. The fixation was performed using 4% glutaraldehyde at 5 °C overnight. Once again, the SS 304 stainless steel coupons were rinsed with physiological saline and dehydrated with a series of graded ethanol (30%, 50%, 60%, 70%, 90%, and 96%). The SS 304 stainless steel coupons were air-dried and coated with gold (Bal-Tec SCD005 Sputter Coater, BAL-TEC AG, Balzers, Liechtenstein) prior to SEM analysis (JMS SEM 6460 LV, JEOL Ltd., Tokyo, Japan).
2.9. Statistical Analysis
The research findings are presented as mean ± standard deviation (SD). Using statistical software R version 3.2.2 (R Foundation for Statistical Computing, Vienna, Austria), the outcomes were evaluated with an analysis of variance (ANOVA) test, followed by a Duncan’s test. The statistical comparison was evaluated at p < 0.05. All assays were performed in triplicate.
3. Results and Discussion
3.1. Chemical Composition of Essential Oils
The detailed chemical compositions of CIEO, CLEO, OREO, ROEO, THEO, and WSEO have been reported in a previous study [30]. The main component of CIEO was cinnamaldehyde (74.93%), followed by ascabin (9.01%). CLEO and OREO were rich in eugenol (85.14%) and carvacrol (81.00%), respectively. The main components of ROEO were α-pinene (28.23%) and borneol (24.87%), followed by α-terpineol (11.86%) and 1,8-cineole (11.54%). THEO was rich in p-cymene (40.91%) and thymol (40.36%). The main component of WSEO was carvacrol (50.45%), followed by p-cymene (15.73%) and γ-terpinene (11.43%). Usually, EOs contain one to three main components at a high concentration, while the others may be at a significantly lower concentration, even as traces [24]. The chemical composition of EOs varies quantitatively and qualitatively. These differences are the outcome of the affect of endogenous and exogenous factors. Endogenous factors include the anatomical and physiological characteristics of plants, i.e., plant genetics (species, ecotype, chemotype), plant origin, season, vegetative phase and parts of plants, physiological and biochemical pathways, degree of development, and metabolic processes of plants. Exogenous factors include the external environment, i.e., climate and habitat conditions (temperature, humidity, windiness, soil composition, geographical origin), cultivation conditions, i.e., agrotechnical measures (method and time of harvesting) and techniques applied after harvesting (methods of drying, extraction, time distillations, and storage conditions) [25,32].
3.2. Antibacterial Effect of Essential Oils
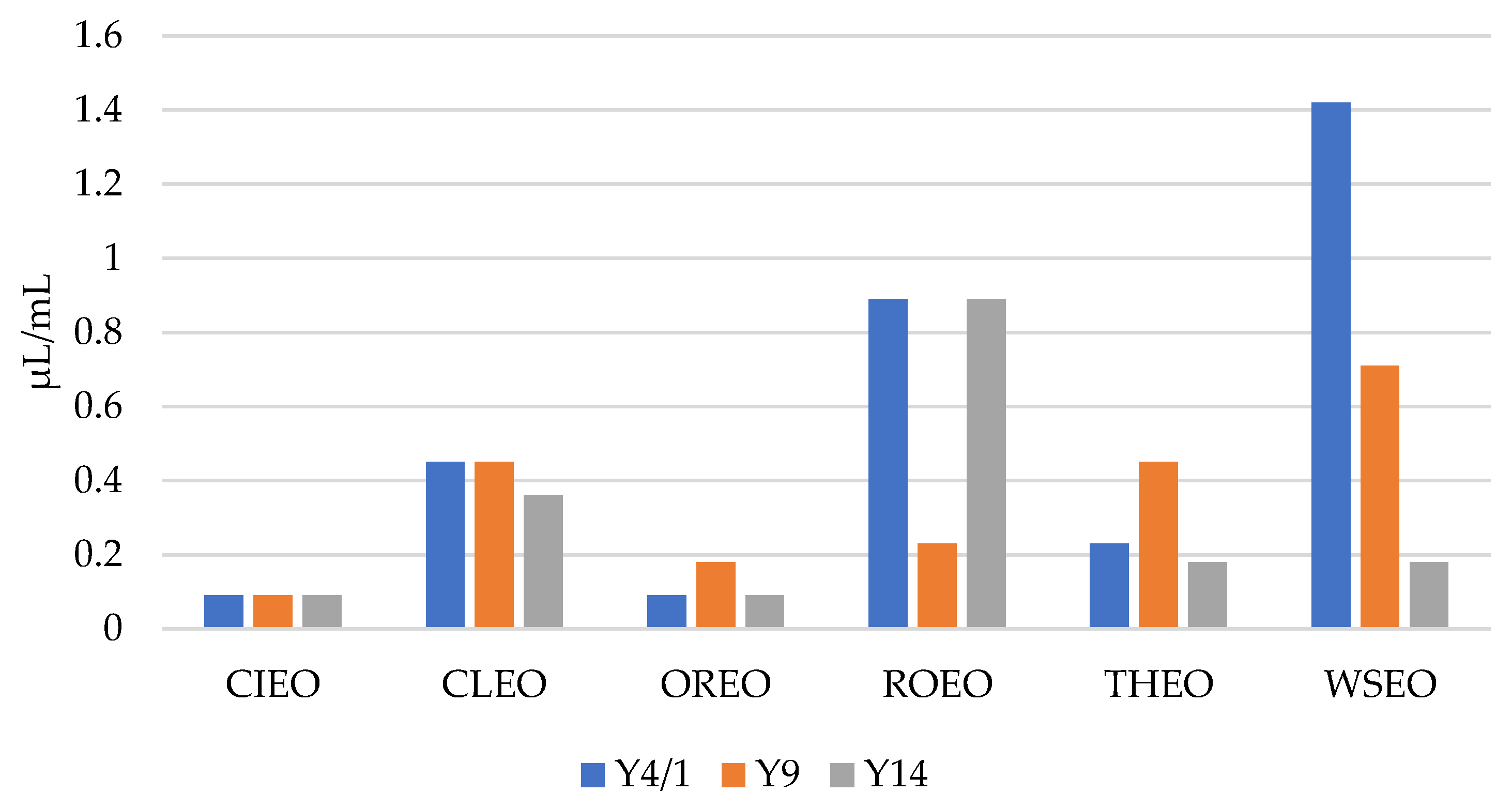
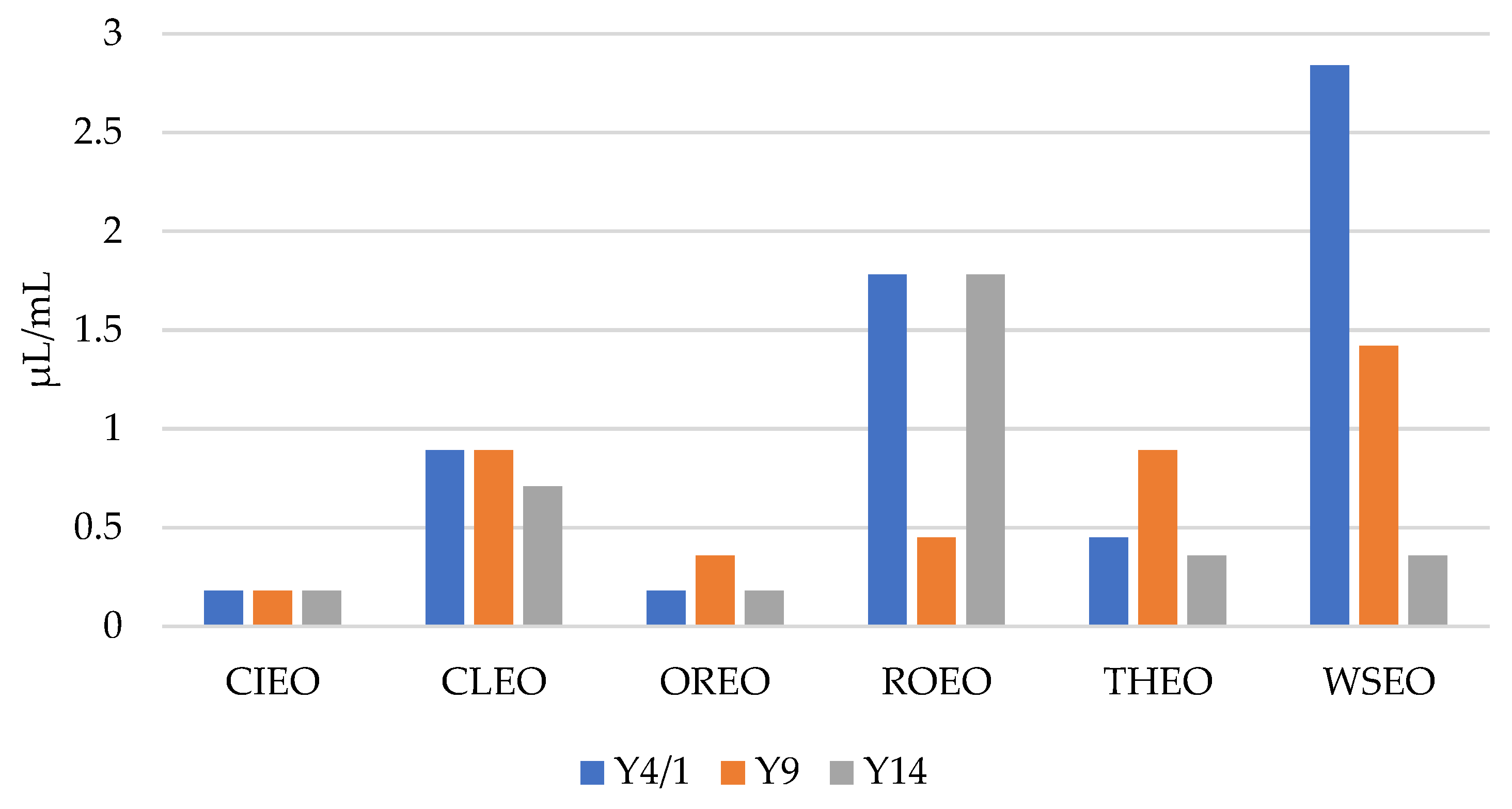
As presented in Figure 1 and Figure 2, the results show that the MICs and MBCs of the EOs for the Y. enterocolitica strains ranged from 0.09 µL/mL to 1.42 µL/mL, and 0.18 µL/mL to 2.84 µL/mL, respectively. CIEO manifested the strongest antibacterial activity, with an MIC value of 0.09 µL/mL for all three Y. enterocolitica strains, followed by OREO, with an MIC value of 0.09 µL/mL for two Y. enterocolitica strains, Y4/1 and Y14. The MIC value for the Y9 strain was 0.18 µL/mL. The widest range of MICs, from 0.18 µL/mL to 1.42 µL/mL, was obtained with WSEO. The antibacterial activity of different EOs, including CIEO, OREO, ROEO, and THEO, against Y. enterocolitica have been previously reported [9,20]. The MIC and MBC values of CIEO, CLEO, OREO, ROEO, THEO, and WSEO for Y. enterocolitica are similar to the values for S. Enteritidis, S. Typhimurium, and L. monocytogenes obtained using the EOs of same origin [26,30]. According to the review of Durofil et al. [20], which included results from more than 50 researchers, EOs originating from plants that belong to the Lamiaceae and Lauracea families possess high activity against Yersinia. This statement is in accordance with our study, with the fact that oregano, thyme, and winter savory belong to the Lamiaceae family, while cinnamon belongs to the Lauracea family.
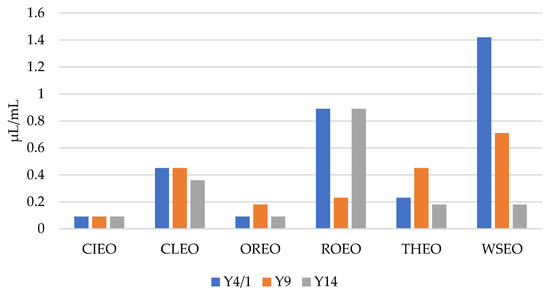
Figure 1.
MICs of cinnamon (Cinnamomum zeylanicum Nees) (CIEO), clove (Syzygium aromaticum L.) (CLEO), oregano (Origanum vulgare L.) (OREO), rosemary (Rosmarinus officinalis L.) (ROEO), thyme (Thymus vulgaris L.) (THEO), and winter savory (Satureja montana L.) (WSEO) against the three strains of Y. enterocolitica.
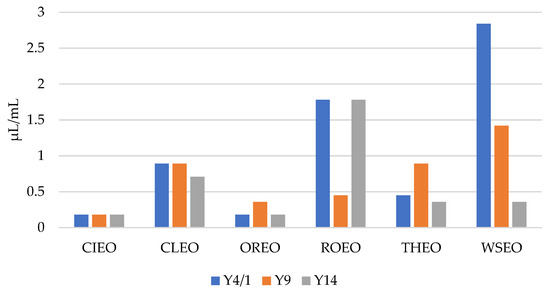
Figure 2.
MBCs of cinnamon (Cinnamomum zeylanicum Nees) (CIEO), clove (Syzygium aromaticum L.) (CLEO), oregano (Origanum vulgare L.) (OREO), rosemary (Rosmarinus officinalis L.) (ROEO), thyme (Thymus vulgaris L.) (THEO), and winter savory (Satureja montana L.) (WSEO) against the three strains of Y. enterocolitica.
3.3. Antibacterial Effect of OREO and THEO on Y. enterocolitica Inoculated in Minced Pork Meat
As reported in Table 1, the initial population of Y. enterocolitica was between 4.31 log CFU/g and 4.46 log CFU/g in all the treatments, without a significant difference (p > 0.05). The OREO at 0.09 µL/g and 0.18 µL/g did not lower the number of Y. enterocolitica in the minced pork meat during the 4 days of storage at 4 ± 1 °C. On the contrary, the THEO significantly (p < 0.05) lowered the number of Y. enterocolitica in the minced pork meat at 0.23 µL/g and 0.46 µL/g, for 0.38 log CFU/g and 0.64 log CFU/g, respectively. Similar to the outcomes of our research, the addition of OREO in barbecued chicken meat did not reduce the number of Y. enterocolitica during 72 h of storage [33]. According to our knowledge, this is the first reported research to examine the efficiency of THEO in minced pork meat against Y. enterocolitica. The reduction in Y. enterocolitica may be due to the high content of thymol, a phenolic monoterpene able to alter the composition of the fatty acids of the cell membrane, and violate its integrity, causing the leakage of intracellular materials [34]. Oregano and thyme are regularly used as spices in meat products, and can elevate the antibacterial, antioxidant, and sensory properties, and consequently extend the shelf-life [35]. However, the required amounts of both EOs in food models are much higher than those obtained in in vitro studies, and can cause negative organoleptic effects [36], with an emphasis on strong aroma. Food components, such as protein and fat, are able to reduce the antibacterial effect of EOs [32]. In addition, Y. enterocolitica is able to activate its adaption mechanisms when exposed to oregano EO to protect itself, including a reduction in energy consumption for mobility, flagellum formation, and QS to ensure normal physiological function [37].

Table 1.
Antibacterial effect of oregano (Origanum vulgare L.) (OREO) and thyme (Thymus vulgaris L.) (THEO) on Y. enterocolitica inoculated in minced pork meat (log CFU/g).
3.4. Formation of Biofilms by Y. enterocolitica
The production of biofilms by Y. enterocolitica strains on the surface of polystyrene wells with the three different nutrient media (TSB, MB, and LB broth) at three different temperatures (5 °C, 15 °C, and 37 °C) over 48 h are shown in Table 2. It is well known that the formation of biofilms is under the influence of the characteristics of bacteria, the surface, and the environment conditions [22]. In this study, at 5 °C, none of Y. enterocolitica strains formed a biofilm. The same was noticed with the MB, regardless of the temperature. The Y. enterocolitica strains were capable of forming weak and moderate biofilms, while no strong biofilms were observed in this study. The Y4/1 strain formed one moderate and three weak biofilms. The Y9 strain formed only a moderate biofilm under two different conditions, while the Y14 strain formed one moderate and two weak biofilms. Similar findings have been reported previously [38,39,40]. According to Wang et al. [40] and our results, Y. enterocolitica can form biofilms under conditions simulating a pork slaughterhouse. Unlike the MB used in our study, the MJ (meat juice) was more suitable for biofilm formation compared to TSB. The biofilm in MJ was more resistant to sanitization treatment, physical washing, and starvation when compared to the biofilm formed in TSB [40].
Table 2.
Production of biofilm by Y. enterocolitica on polystyrene surface under different conditions of nutrient media and temperatures.
In our study, only 3 h of adhesion was enough for the Y. enterocolitica cells to attach to the surface and start forming a biofilm. Other studies have shown that adhesion can be observed after 6–8 h, while the formation of a mature biofilm takes 48–72 h [41]. The plasmid for Yersinia virulence (pYV) is essential for the surface properties of Y. enterocolitica [42]. The capability of Y. enterocolitica to produce biofilms contributes to its pathogenicity and adaptability [43], which increases the risk of contaminating food-contacting surfaces in processing plants and final products. Y. enterocolitica biofilm formations are more common at higher temperatures, so keeping the temperature cool may reduce the bacterial adhesion on food-contacting surfaces and reduce the biofilm formation in production plants.
3.5. Biofilm Reduction
Biofilm reduction was conducted on selected moderate biofilm formers. The MBC concentrations of EOs reduced the 48 h old Y. enterocolitica biofilms in the range from 45.34% to 78.89%, as shown in Table 3. To reduce established biofilms, EO must penetrate the exopolysaccharide matrix, reach the protected surface-attached bacterial cells, and alter the QS system. The QS system controls biofilm formation, so the strategy in biofilm reduction is targeting QS [16].
Table 3.
Reduction (%) in Y. enterocolitica biofilms formed at polystyrene surface exposed to cinnamon (Cinnamomum zeylanicum Nees) (CIEO), clove (Syzygium aromaticum L.) (CLEO), oregano (Origanum vulgare L.) (OREO), rosemary (Rosmarinus officinalis L.) (ROEO), thyme (Thymus vulgaris L.) (THEO), and winter savory (Satureja montana L.) (WSEO) essential oils.
The major components of EOs, such as cinnamaldehyde, eugenol, carvacrol, p-cymene, and thymol, are responsible for their antibiofilm effects. However, the components of EOs present in smaller amounts play a significant synergistic role [22]. Former studies have suggested that low concentrations of cinnamaldehyde (0.078 mg/mL) can repress the production of Y. enterocolitica biofilm [16]. In a previous study, it was shown that extracts from 12 edible plants inhibited the biofilm production of Y. enterocolitica [44]. Natural antibacterial agents cause abnormal expression of a few important genes, including luxS, glgC, envZ, ompF, kdpD, and cydB, resulting in the damage of the biofilm [45]. Generally, planktonic cells are more sensitive to EOs compared with cells protected in biofilms. Therefore, the strategy to control biofilms in production plants is an important part of food safety. The antibiofilm mechanisms of EOs are different and are not entirely clear [22], which can also be concluded from the results of this study. Namely, the achieved reduction in Y. enterocolitica biofilms varied within the same applied EO. The lowest difference was observed applying ROEO (14.75%), while the highest difference was observed applying THEO (32.8%). Differences in biofilm reduction could be explained by the influence of temperature. Lower reduction rates were observed at 15 °C, compared with 37 °C. The temperature of 30 °C is the optimal growth temperature for Y. enterocolitica, so the bacterial cells in the biofilm formed and treated at 37 °C could be stronger and more resistant to the applied EOs. Additionally, EOs could exhibit different affinities for different surfaces. Polystyrene has a hydrophobic surface, and attracts EOs more than hydrophilic stainless steel [22].
3.6. SEM
The SEM assay was performed on the Y. enterocolitica strain Y9. This strain was chosen because of its ability to form the strongest biofilms compared with the other two Y. enterocolitica strains. Figure 3 shows SEM micrographs of the untreated Y. enterocolitica cells, and the Y. enterocolitica cells treated with MIC amounts of OREO (0.18 µL/mL), and THEO (0.45 µL/mL). The untreated Y. enterocolitica cells have their typical morphological appearance; a smooth surface and rod-shaped structure [45], while the Y. enterocolitica cells treated with OREO and THEO showed damage, with deformed shapes, rough surfaces, and membrane rupture. The EO components, such as carvacrol, p-cymene, and thymol, act on bacterial cells by various antimicrobial mechanisms, including the attack of the phospholipid bilayer, disrupting enzyme systems, metabolic pathways, and the genetic material of bacterial cells, causing structural and functional damages to the bacterial cell membrane, that eventually lead to cell death [36,46]. Some of the bacteria cells treated with thymol showed swelling [47]. The bacteria cells treated with carvacrol and thymol modified the lipid profile, resulting in an increase in saturated C16 and C18 fatty acids, and a decrease in unsaturated C18 fatty acids, causing membrane structural alterations and permeability [48]. Carvacrol has an effect on the proteins of the outer membrane of Gram-negative bacteria, which allows Gram-negative bacteria to be more resistant to EOs and their compounds compared to Gram-positive bacteria [47].

Figure 3.
SEM micrographs of Y. enterocolitica cells on SS 304 stainless steel coupons: untreated (A) and treated with oregano (Origanum vulgare L.) essential oil (B) and thyme (Thymus vulgaris L.) essential oil (C). Magnifications: (A) ×20,000, (B) ×20,000, (C) ×30,000.
4. Conclusions
The prevalence of Y. enterocolitica is high, and as the leading foodborne infection agent, this pathogen deserves more attention. In vitro studies have shown that the use of EOs may be an effective treatment against Y. enterocolitica. CIEO, CLEO, OREO, ROEO, THEO, and WSEO showed high anti-Yersinia effects, with MIC values > 0.09 µL/mL. However, the fact that only the THEO (MIC and 2MIC) was able to reduce the number of Y. enterocolitica in minced pork meat during storage at 4 ± 1 °C for 4 days shows the limited practical application of EOs in food at the concentrations obtained in in vitro studies. This study also shows the ability of Y. enterocolitica to form biofilms in different conditions, regarding temperatures and available nutrients. Y. enterocolitica was able to attach to polystyrene and stainless steel surfaces, and started to form biofilms in just 3 h. The CIEO, CLEO, OREO, ROEO, THEO, and WSEO were able to reduce the biofilms of Y. enterocolitica strains formed at 15 °C and 37 °C in TSB and LB broth. The applied EOs reduced the biomass of the preformed biofilms by up to 78.89%. SEM showed that OREO and THEO influenced the typical morphological appearance of the Y. enterocolitica cells, causing a deformed shape and membrane rupture. This is evidence that environmentally friendly EOs may be used to control foodborne pathogenic biofilms present on food-contacting surfaces made of polystyrene and stainless steel. However, further research is necessary to find adequate EO concentrations applicable to food and surfaces in order to control Y. enterocolitica, before the industrial application of EOs.
Author Contributions
Conceptualization, S.V.K., S.K.-T. and N.K.; methodology, S.V.K., D.M., S.K.-T. and N.K.; formal analysis, S.V.K., S.K. and Z.R.; investigation, S.V.K., S.K. and J.V.; resources, S.V.K., D.M. and N.K.; writing—original draft preparation, S.V.K.; writing—review and editing, S.K.-T.; supervision, S.K.-T. and N.K.; project administration, S.V.K., S.K.-T. and N.K. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the Ministry of Science, Technological Development and Innovation of the Republic of Serbia by the contract of implementation and funding of research work of NIV-NS in 2024, Contract No: 451-03-66/2024-03/200031 and by the Provincial Secretariat for Higher Education and Scientific Research, Republic of Serbia, Autonomous Province of Vojvodina (No. 142-451-3498/2023-01/01).
Institutional Review Board Statement
An institutional review board statement is not required for this paper in Serbia.
Informed Consent Statement
Not applicable.
Data Availability Statement
The original contributions presented in the study are included in the article, further inquiries can be directed to the corresponding author.
Acknowledgments
The authors are grateful to the TerraCo d.o.o., Novi Sad, Republic of Serbia, for providing the essential oils, and to the “Đurđević” meat industry, Industrijska zona BB, Subotište, Pećinci, Republic of Serbia, for providing the pork meat.
Conflicts of Interest
The authors declare that they have no conflicts of interest.
References
- LPSN—List of Prokaryotic Names with Standing in Nomenclature. Available online: https://lpsn.dsmz.de/ (accessed on 25 December 2023).
- Adeolu, M.; Alnajar, S.; Naushad, S.; Gupta, R.S. Genome-based phylogeny and taxonomy of the ‘Enterobacteriales’: Proposal for Enterobacterales ord. nov. divided into the families Enterobacteriaceae, Erwiniaceae fam. nov., Pectobacteriaceae fam. nov., Yersiniaceae fam. nov., Hafniaceae fam. nov., Morganellaceae fam. nov., and Budviciaceae fam. nov. Int. J. Syst. Evol. Microbiol. 2016, 66, 5575–5599. [Google Scholar]
- Savin, C.; Criscuolo, A.; Guglielmini, J.; Le Guern, A.S.; Carniel, E.; Pizarro-Cerdá, J.; Brisse, S. Genus-wide Yersinia core-genome multilocus sequence typing for species identification and strain characterization. Microb. Genom. 2019, 5, e000301. [Google Scholar] [CrossRef]
- Barton, M.D.; Robins-Browne, R.M. Foodborne microorganisms of public health significance. In Yersinia Enterocolitica, 6th ed.; Hocking, A.D., Ed.; Australian Institute of Food Science and Technology Incorporated (AIFST Inc.): Waterloo, Australia, 2003; pp. 577–595. [Google Scholar]
- Gupta, V.; Gulati, P.; Bhagat, N.; Dhar, M.S.; Virdi, J.S. Detection of Yersinia enterocolitica in food: An overview. Eur. J. Clin. Microbiol. Infect. Dis. 2015, 34, 641–650. [Google Scholar] [CrossRef]
- Karlsson, P.A.; Tano, E.; Jernberg, C.; Hickman, R.A.; Guy, L.; Järhult, J.D.; Wang, H. Molecular characterization of multidrug-resistant Yersinia enterocolitica from foodborne outbreaks in Sweden. Front. Microbiol. 2021, 12, 664665. [Google Scholar] [CrossRef]
- Fukushima, H.; Shimizu, S.; Inatsu, Y. Yersinia enterocolitica and Yersinia pseudotuberculosis detection in foods. J. Pathog. 2011, 2011, 735308. [Google Scholar] [CrossRef]
- Gruber, J.F.; Morris, S.; Warren, K.A.; Kline, K.E.; Schroeder, B.; Dettinger, L.; Husband, B.; Pollard, K.; Davis, C.; Miller, J.; et al. Yersinia enterocolitica outbreak associated with pasteurized milk. Foodborne Pathog. Dis. 2021, 18, 448–454. [Google Scholar] [CrossRef]
- Hulankova, R. Higher Resistance of Yersinia enterocolitica in Comparison to Yersinia pseudotuberculosis to Antibiotics and Cinnamon, Oregano and Thyme Essential Oils. Pathogens 2022, 11, 1456. [Google Scholar] [CrossRef]
- Angelovska, M.; Zaharieva, M.M.; Dimitrova, L.L.; Dimova, T.; Gotova, I.; Urshev, Z.; Ilieva, Y.; Kaleva, M.D.; Kim, T.C.; Naydenska, S.; et al. Prevalence, Genetic Homogeneity, and Antibiotic Resistance of Pathogenic Yersinia enterocolitica Strains Isolated from Slaughtered Pigs in Bulgaria. Antibiotics 2023, 12, 716. [Google Scholar] [CrossRef]
- Pegoraro, K.; Sereno, M.J.; Viana, C.; Martins, B.T.F.; Yamatogi, R.S.; Nero, L.A.; Bersot, L.D.S. Pathogenic potential and antibiotic resistance of Yersinia enterocolitica, a foodborne pathogen limited to swine tonsils in a pork production chain from Southern Brazil. Braz. J. Microbiol. 2021, 52, 2335–2342. [Google Scholar] [CrossRef]
- Rivas, L.; Strydom, H.; Paine, S.; Wang, J.; Wright, J. Yersiniosis in New Zealand. Pathogens 2021, 10, 191. [Google Scholar] [CrossRef]
- Angelovska, M.; Zaharieva, M.M.; Najdenski, H. Yersinia enterocolitica—Isolation, Pathogenicity, and Prevalence in Farms for Slaughtered Pigs. Acta Microbiol. Bulg. 2023, 39, 118–129. [Google Scholar] [CrossRef]
- Zdolec, N.; Kiš, M. Meat Safety from Farm to Slaughter—Risk-Based Control of Yersinia enterocolitica and Toxoplasma gondii. Processes 2021, 9, 815. [Google Scholar] [CrossRef]
- Verderosa, A.D.; Totsika, M.; Fairfull-Smith, K.E. Bacterial biofilm eradication agents: A current review. Front. Chem. 2019, 7, 824. [Google Scholar] [CrossRef]
- Gao, C.; Tian, L.; Lu, J.; Zhang, S.; Wu, D.; Xiao, P.; Gong, G. Cinnamaldehyde reduces Yersinia enterocolitica activity and biofilm formation by altering membrane permeability and signalling. Food Biosci. 2023, 53, 102791. [Google Scholar] [CrossRef]
- Siddharth, T.; Sridhar, P.; Vinila, V.; Tyagi, R.D. Environmental applications of microbial extracellular polymeric substance (EPS): A review. J. Environ. Manag. 2021, 287, 112307. [Google Scholar] [CrossRef]
- Abebe, G.M. The role of bacterial biofilm in antibiotic resistance and food contamination. Int. J. Microbiol. 2020, 2020, 1705814. [Google Scholar] [CrossRef]
- Shineh, G.; Mobaraki, M.; Perves Bappy, M.J.; Mills, D.K. Biofilm formation, and related impacts on healthcare, food processing and packaging, industrial manufacturing, marine industries, and sanitation—A review. Appl. Microbiol. 2023, 3, 629–665. [Google Scholar] [CrossRef]
- Durofil, A.; Maddela, N.R.; Naranjo, R.A.; Radice, M. Evidence on antimicrobial activity of essential oils and herbal extracts against Yersinia enterocolitica—A review. Food Biosci. 2022, 47, 101712. [Google Scholar] [CrossRef]
- Mutlu-Ingok, A.; Devecioglu, D.; Dikmetas, D.N.; Karbancioglu-Guler, F.; Capanoglu, E. Antibacterial, antifungal, antimycotoxigenic, and antioxidant activities of essential oils: An updated review. Molecules 2020, 25, 4711. [Google Scholar] [CrossRef] [PubMed]
- Rossi, C.; Chaves-López, C.; Serio, A.; Casaccia, M.; Maggio, F.; Paparella, A. Effectiveness and mechanisms of essential oils for biofilm control on food-contact surfaces: An updated review. Crit. Rev. Food Sci. Nutr. 2022, 62, 2172–2191. [Google Scholar] [CrossRef] [PubMed]
- Ramos da Silva, L.R.; Ferreira, O.O.; Cruz, J.N.; de Jesus Pereira Franco, C.; Oliveira Dos Anjos, T.; Cascaes, M.M.; Almeida da Costa, W.; Helena de Aguiar Andrade, E.; Santana de Oliveira, M. Lamiaceae essential oils, phytochemical profile, antioxidant, and biological activities. Evid. Based Complement. Altern. Med. 2021, 2021, 6748052. [Google Scholar] [CrossRef] [PubMed]
- Jugreet, B.S.; Suroowan, S.; Rengasamy, R.K.; Mahomoodally, M.F. Chemistry, bioactivities, mode of action and industrial applications of essential oils. Trends Food Sci. Technol. 2020, 101, 89–105. [Google Scholar] [CrossRef]
- Falleh, H.; Jemaa, M.B.; Saada, M.; Ksouri, R. Essential oils: A promising eco-friendly food preservative. Food Chem. 2020, 330, 127268. [Google Scholar] [CrossRef]
- Vidaković Knežević, S.; Kocić-Tanackov, S.; Kravić, S.; Knežević, S.; Vranešević, J.; Savić Radovanović, R.; Karabasil, N. In vitro antibacterial activity of some essential oils against Salmonella Enteritidis and Salmonella Typhimurium isolated from meat. J. Food Saf. Food Qual. 2021, 72, 4–11. [Google Scholar]
- Arsić, M.; Vićić, I.; Galić, N.; Dmitrić, M.; Kureljušić, J.; Dimitrijević, M.; Petrović, M.; Šarić, L.; Karabasil, N. Risk factors and the overall characterization of Yersinia enterocolitica as an initial model of pathogen surveillance in the pig production system in Serbia. Res. Vet. Sci. 2022, 152, 167–174. [Google Scholar] [CrossRef]
- Kocić-Tanackov, S.; Blagojev, N.; Suturović, I.; Dimić, G.; Pejin, J.; Tomović, V.; Šojić, B.; Savanović, J.; Kravić, S.; Karabasil, N. Antibacterial activity essential oils against Escherichia coli, Salmonella enterica and Listeria monocytogenes. J. Food Saf. Food Qual. 2017, 68, 88–95. [Google Scholar]
- ISO 10273:2017; Microbiology of the Food Chain—Horizontal Method for the Detection of Pathogenic Yersinia enterocolitica. International Organization for Standardization: Geneva, Switzerland, 2017.
- Vidaković Knežević, S.; Knežević, S.; Vranešević, J.; Kravić, S.Ž.; Lakićević, B.; Kocić-Tanackov, S.; Karabasil, N. Effects of Selected Essential Oils on Listeria monocytogenes in Biofilms and in a Model Food System. Foods 2023, 12, 1930. [Google Scholar] [CrossRef]
- Stepanović, S.; Vuković, D.; Hola, V.; Bonaventura, G.D.; Djukić, S.; Ćirković, I.; Ruzicka, F. Quantification of biofilm in microtiter plates: Overview of testing conditions and practical recommendations for assessment of biofilm production by staphylococci. APMIS J. Pathol. Microbiol. Immunol. 2007, 115, 891–899. [Google Scholar] [CrossRef]
- Pateiro, M.; Barba, F.J.; Domínguez, R.; Sant’Ana, A.S.; Khaneghah, A.M.; Gavahian, M.; Gómez, B.; Lorenzo, J.M. Essential oils as natural additives to prevent oxidation reactions in meat and meat products: A review. Food Res. Int. 2018, 113, 156–166. [Google Scholar] [CrossRef]
- Firouzi, R.; Shekarforoush, S.S.; Nazer, A.H.K.; Borumand, Z.; Jooyandeh, A.R. Effects of essential oils of oregano and nutmeg on growth and survival of Yersinia enterocolitica and Listeria monocytogenes in barbecued chicken. J. Food Prot. 2007, 70, 2626–2630. [Google Scholar] [CrossRef]
- Posgay, M.; Greff, B.; Kapcsándi, V.; Lakatos, E. Effect of Thymus vulgaris L. essential oil and thymol on the microbiological properties of meat and meat products: A review. Heliyon 2022, 8, e10812. [Google Scholar] [CrossRef] [PubMed]
- Yu, H.H.; Chin, Y.-W.; Paik, H.-D. Application of Natural Preservatives for Meat and Meat Products against Food-Borne Pathogens and Spoilage Bacteria: A Review. Foods 2021, 10, 2418. [Google Scholar] [CrossRef] [PubMed]
- Rao, J.; Chen, B.; McClements, D.J. Improving the efficacy of essential oils as antimicrobials in foods: Mechanisms of action. Annu. Rev. Food Sci. Technol. 2019, 10, 365–387. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Wang, R.; Zhao, L.; Wang, M.; Wang, R.; Guo, D.; Yang, Y.; Li, Y.; Guan, N.; Shi, Y.; et al. Stress tolerance and transcriptomic response analysis of Yersinia enterocolitica adapted to Origanum vulgare L. essential oil. LWT Food Sci. Technol. 2022, 168, 113930. [Google Scholar] [CrossRef]
- Ioannidis, A.; Kyratsa, A.; Ioannidou, V.; Bersimis, S.; Chatzipanagiotou, S. Detection of biofilm production of Yersinia enterocolitica strains isolated from infected children and comparative antimicrobial susceptibility of biofilm versus planktonic forms. Mol. Diagn. Ther. 2014, 18, 309–314. [Google Scholar] [CrossRef]
- Zadernowska, A.; Chajęcka-Wierzchowska, W. Prevalence, biofilm formation and virulence markers of Salmonella sp. and Yersinia enterocolitica in food of animal origin in Poland. LWT 2017, 75, 552–556. [Google Scholar] [CrossRef]
- Wang, H.; Tay, M.; Palmer, J.; Flint, S. Biofilm formation of Yersinia enterocolitica and its persistence following treatment with different sanitation agents. Food Control 2017, 73, 433–437. [Google Scholar] [CrossRef]
- Lenchenko, E.; Lozovoy, D.; Strizhakov, A.; Vatnikov, Y.; Byakhova, V.; Kulikov, E.; Sturov, N.; Kuznetsov, V.; Avdotin, V.; Grishin, V. Features of formation of biofilms. Vet. World 2019, 12, 136. [Google Scholar] [CrossRef]
- Wang, H.; Palmer, J.; Flint, S. Function of pYV plasmid on biofilm formation of Yersinia enterocolitica ERL032123 in the presence of Ca2+. J. Food Prot. 2019, 82, 1683–1687. [Google Scholar] [CrossRef] [PubMed]
- Wang, H. Factors Contributing to Biofilm Formation of Yersinia enterocolitica. Ph.D. Thesis, Massey University, Palmerston North, New Zealand, December 2016. [Google Scholar]
- Ham, Y.; Kim, T.J. Inhibition of biofilm formation in Yersinia enterocolitica by edible plant extracts including Polygoni Multiflori Radix. J. Korean Wood Sci. Technol. 2022, 50, 448–457. [Google Scholar] [CrossRef]
- Chen, K.; Peng, C.; Chi, F.; Yu, C.; Yang, Q.; Li, Z. Antibacterial and antibiofilm activities of chlorogenic acid against Yersinia enterocolitica. Front. Microbiol. 2022, 13, 885092. [Google Scholar] [CrossRef] [PubMed]
- Tajkarimi, M.M.; Ibrahim, S.A.; Cliver, D.O. Antimicrobial herb and spice compounds in food. Food Control 2010, 21, 1199–1218. [Google Scholar] [CrossRef]
- Di Pasqua, R.; Betts, G.; Hoskins, N.; Edwards, M.; Ercolini, D.; Mauriello, G. Membrane toxicity of antimicrobial compounds from essential oils. J. Agric. Food Chem. 2007, 55, 4863–4870. [Google Scholar] [CrossRef] [PubMed]
- Nazzaro, F.; Fratianni, F.; De Martino, L.; Coppola, R.; De Feo, V. Effect of essential oils on pathogenic bacteria. Pharmaceuticals 2013, 6, 1451–1474. [Google Scholar] [CrossRef] [PubMed]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).