
Effect of Boiling Treatment on Linoleic Acid-Induced Oxidation of Myofibrillar Protein in Grass Carp
 ,
,
Abstract

1. Introduction
2. Materials and Methods
2.1. Samples and Materials
2.2. Experimental Methods
2.2.1. MP Extraction
2.2.2. MP Oxidation
2.2.3. Determination of Protein Oxidation
Carbonyl Content Measurement
Total Sulfhydryl Content Measurement
Fluorescence Spectroscopy
2.2.4. Determination of Protein Thermal Aggregation Indices
Surface Hydrophobicity Determination
Particle Size and Zeta Potential
Turbidity and Solubility
2.2.5. Determination of Protein Structure
Ultraviolet–Visible Absorption Spectroscopy
Fourier Transform Infrared (FTIR) Spectroscopy
Scanning Electron Microscope (SEM)
SDS-PAGE
2.2.6. Amino Acid Content Determination
2.2.7. Determination of MP Gel Water Retention
Preparation of MP Gel
Water-Holding Capacity (WHC)
Low-Field Nuclear Magnetic Resonance (LF-NMR) Proton Relaxation
2.3. Statistical Analysis
3. Result and Discussion
3.1. Protein Oxidation Analysis
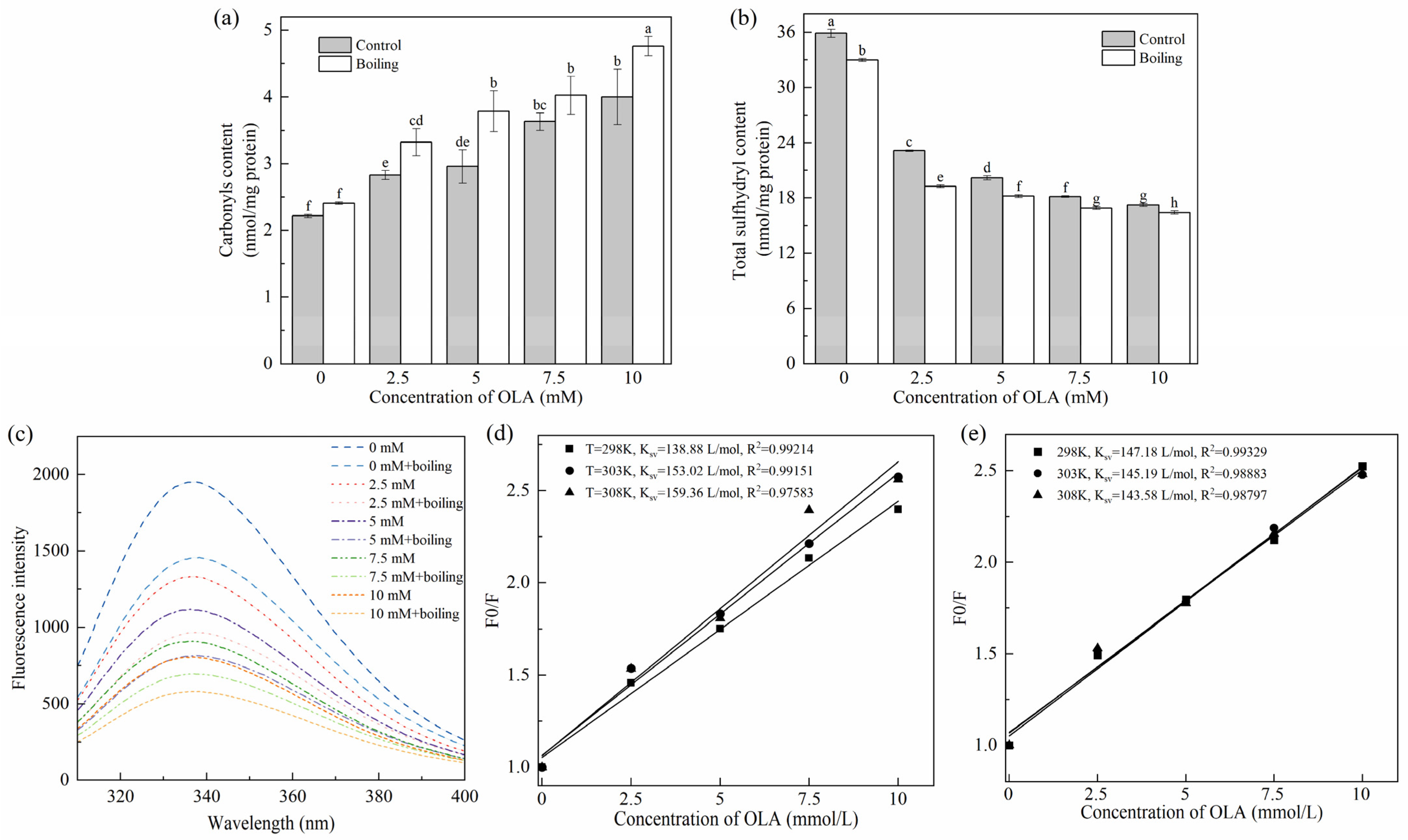
3.1.1. Carbonyl Content
3.1.2. Total Sulfhydryl Content
3.1.3. Amino Acid Analysis
3.1.4. Tryptophan Endogenous Fluorescence Intensity
3.2. Protein Thermal Aggregation Analysis
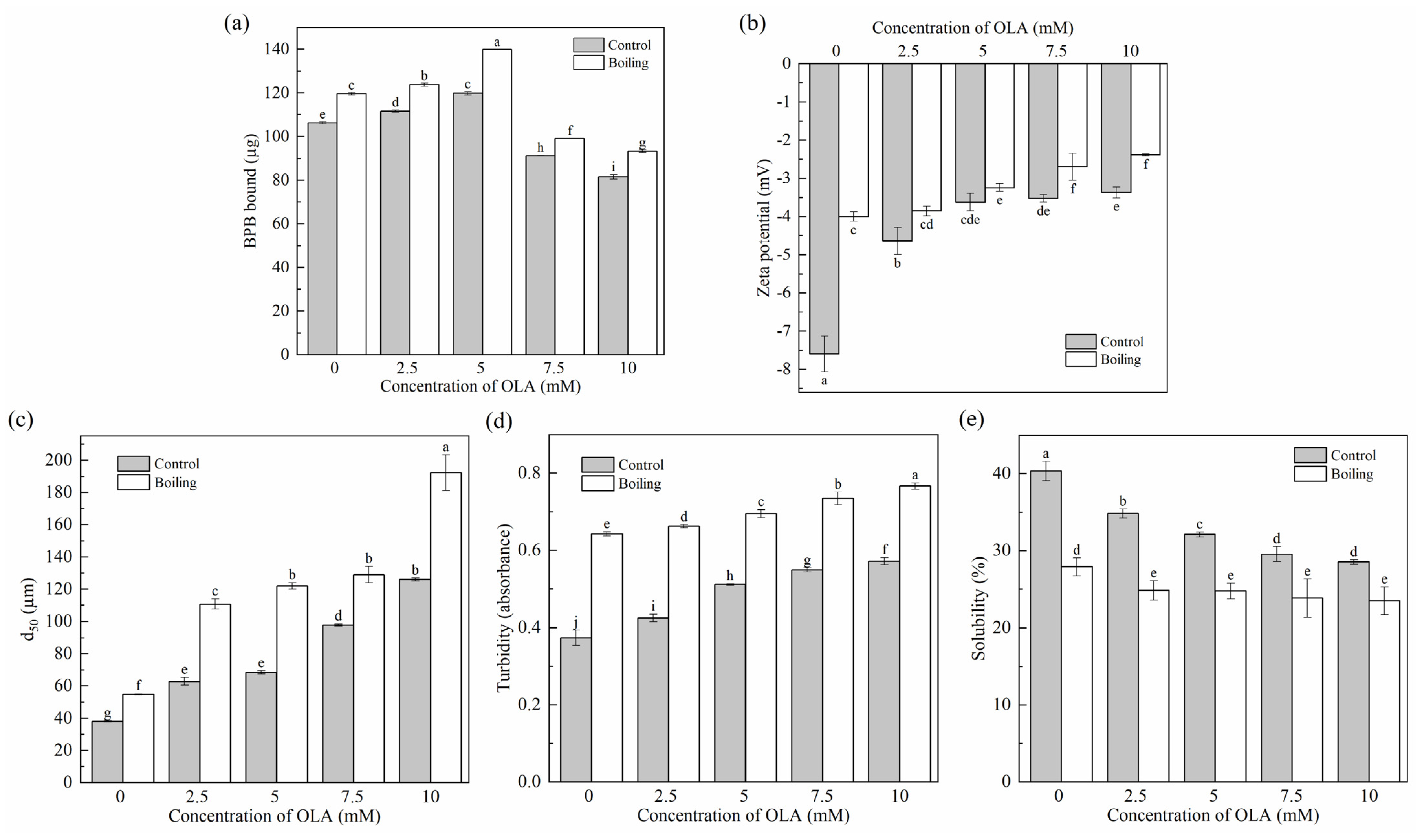
3.2.1. Surface Hydrophobicity
3.2.2. Zeta Potential
3.2.3. Particle Size, Turbidity, and Solubility
3.3. Protein Structure Analysis
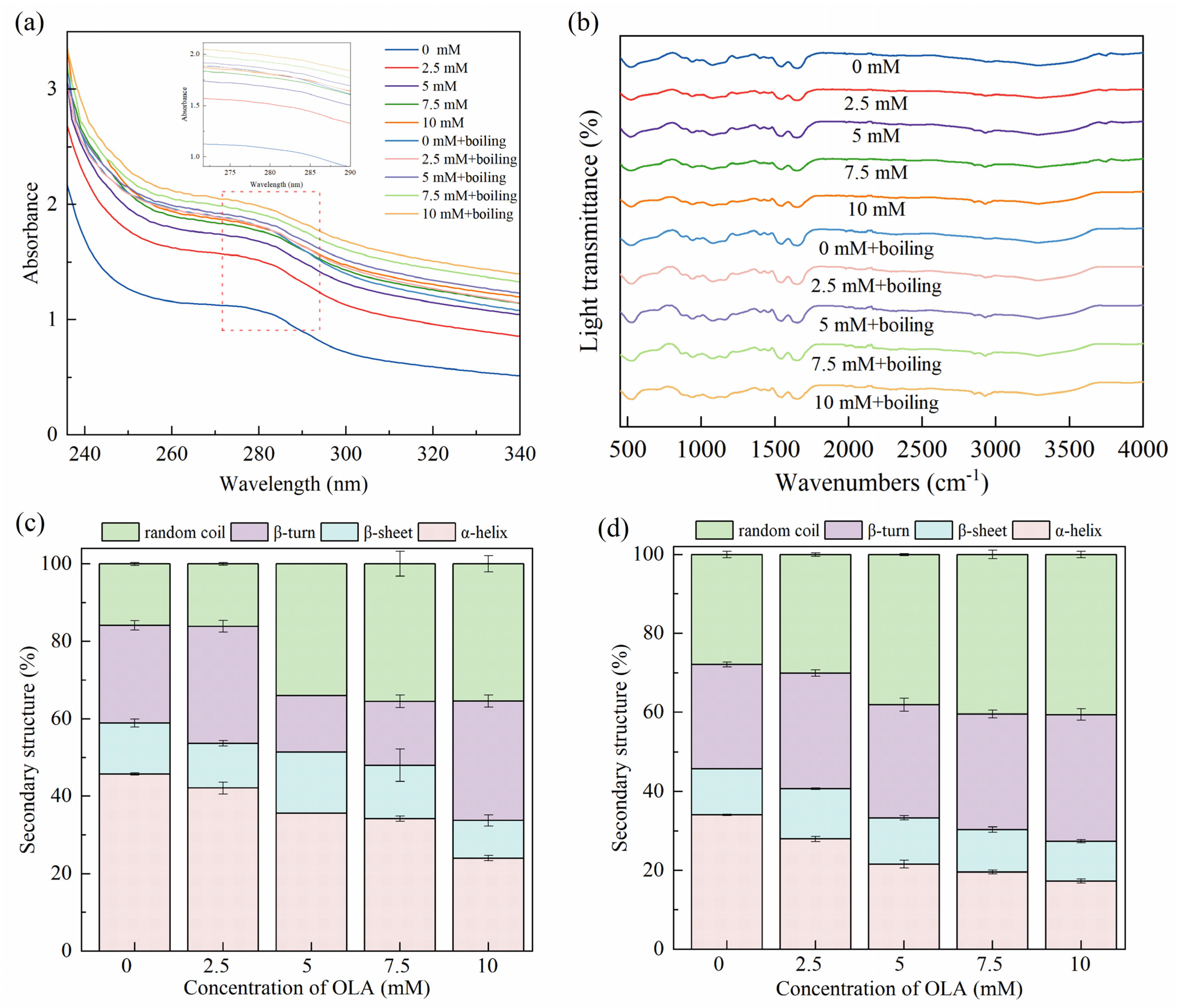
3.3.1. UV Absorption of MP
3.3.2. Secondary Structure
3.3.3. Microstructure
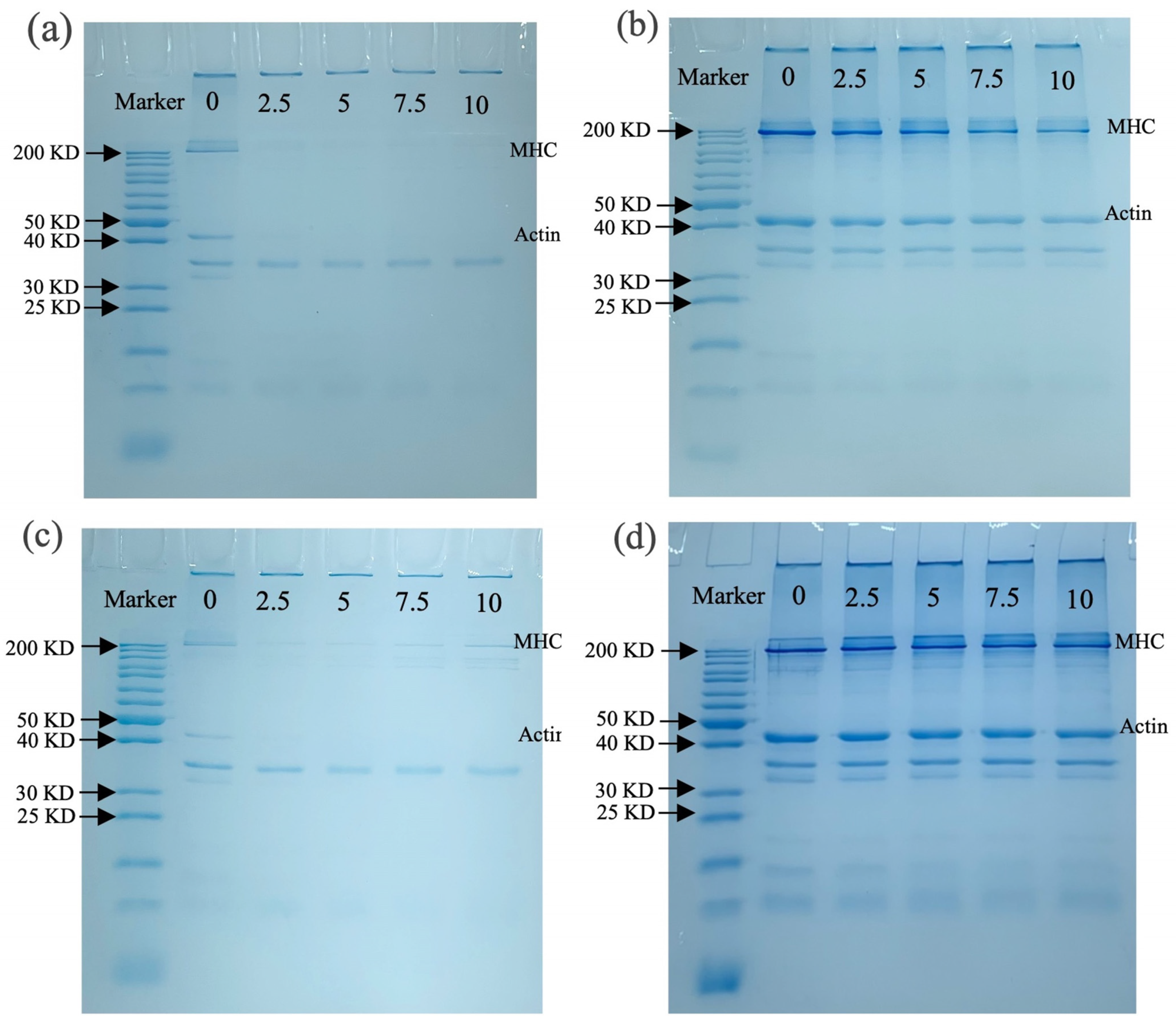
3.3.4. SDS-PAGE Analysis
3.4. Analysis of Water Retention of MP Gel
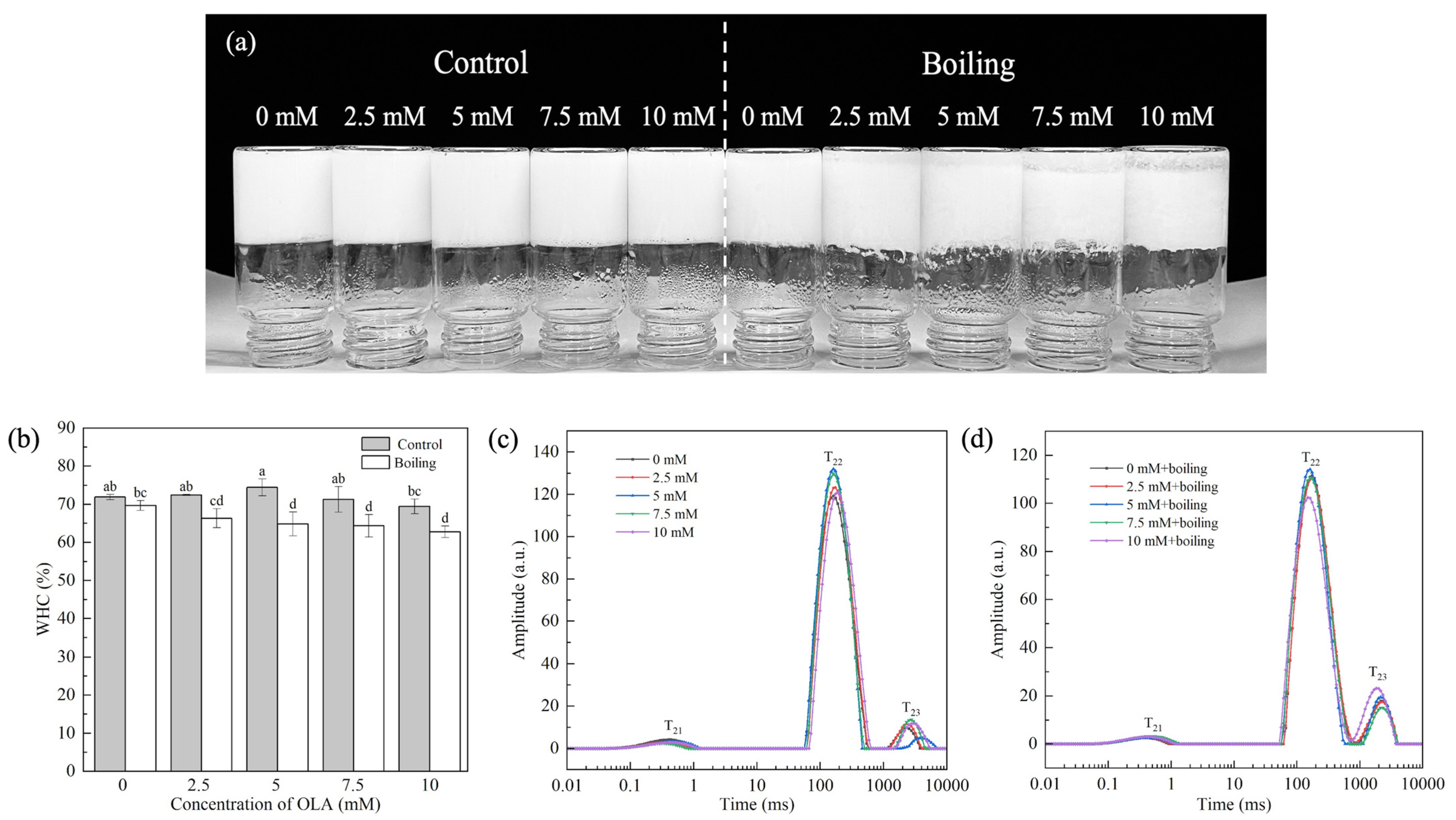
3.4.1. Gel WHC
3.4.2. Water Distribution
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Tan, C.; Xu, Q.; Chen, N.; He, Q.; Zeng, W. Cross-linking modifications of different phenolic compounds on myofibrillar protein of common carp. Food Bioprocess Technol. 2023, 16, 627–638. [Google Scholar] [CrossRef]
- Cao, J.; Zhou, C.; Wang, Y.; Sun, Y.; Pan, D. The effect of oxidation on the structure of G-actin and its binding ability with aroma compounds in carp grass skeletal muscle. Food Chem. 2018, 240, 346–353. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Bai, G.; Wang, J.; Wang, Y.; Jin, G.; Teng, W.; Geng, F.; Cao, J. Myofibrillar protein denaturation/oxidation in freezing-thawing impair the heat-induced gelation: Mechanisms and control technologies. Trends Food Sci. Technol. 2023, 138, 655–670. [Google Scholar] [CrossRef]
- Lan, M.; Li, T.; Li, L.; Wang, S.; Chen, J.; Yang, T.; Li, Z.; Yang, Y.; Zhang, X.; Li, B. Ultrasonic treatment treated sea bass myofibrillar proteins in low-salt solution: Emphasizing the changes on conformation structure, oxidation sites, and emulsifying properties. Food Chem. 2024, 435, 137564. [Google Scholar] [CrossRef]
- Zhang, L.; Li, Q.; Hong, H.; Luo, Y. Tracking structural modifications and oxidative status of myofibrillar proteins from silver carp (Hypophthalmichthys molitrix) fillets treated by different stunning methods and in vitro oxidizing conditions. Food Chem. 2021, 365, 130510. [Google Scholar] [CrossRef] [PubMed]
- Fang, M.; Xiong, S.; Yin, T.; Hu, Y.; Liu, R.; Du, H.; Liu, Y.; You, J. Proteomic profiling and oxidation site analysis of gaseous ozone oxidized myosin from silver carp (Hypophthalmichthys molitrix) with different oxidation degrees. Food Chem. 2021, 363, 130307. [Google Scholar] [CrossRef] [PubMed]
- Jiao, X.; Yan, B.; Huang, J.; Zhao, J.; Zhang, H.; Chen, W.; Fan, D. Redox proteomic analysis reveals microwave-induced oxidation modifications of myofibrillar proteins from silver carp (Hypophthalmichthys molitrix). J. Agric. Food Chem. 2021, 69, 9706–9715. [Google Scholar] [CrossRef] [PubMed]
- Nawaz, A.; Irshad, S.; Khan, I.A.; Khalifa, I.; Walayat, N.; Aadil, R.M.; Kumar, M.; Wang, M.; Chen, F.; Cheng, K.; et al. Protein oxidation in muscle-based products: Effects on physicochemical properties, quality concerns, and challenges to food industry. Food Res. Int. 2022, 157, 111322. [Google Scholar] [CrossRef] [PubMed]
- Bao, Y.; Ertbjerg, P. Effects of protein oxidation on the texture and water-holding of meat: A review. Crit. Rev. Food Sci. Nutr. 2019, 59, 3564–3578. [Google Scholar] [CrossRef] [PubMed]
- Ferreira, V.C.S.; Morcuende, D.; Madruga, M.S.; Silva, F.A.P.; Estévez, M. Role of protein oxidation in the nutritional loss and texture changes in ready-to-eat chicken patties. Int. J. Food Sci. Technol. 2018, 53, 1518–1526. [Google Scholar] [CrossRef]
- Mohanty, B.P.; Mahanty, A.; Ganguly, S.; Mitra, T.; Karunakaran, D.; Anandan, R. Nutritional composition of food fishes and their importance in providing food and nutritional security. Food Chem. 2019, 293, 561–570. [Google Scholar] [CrossRef] [PubMed]
- Bekhit, A.E.A.; Hopkins, D.L.; Fahri, F.T.; Ponnampalam, E.N. Oxidative processes in muscle systems and fresh meat: Sources, markers, and remedies. Compr. Rev. Food Sci. Food Saf. 2013, 12, 565–597. [Google Scholar] [CrossRef]
- Domínguez, R.; Pateiro, M.; Gagaoua, M.; Barba, F.; Zhang, W.; Lorenzo, J.M. A Comprehensive review on lipid oxidation in meat and meat products. Antioxidants 2019, 8, 429. [Google Scholar] [CrossRef] [PubMed]
- Mariutti, L.R.B.; Bragagnolo, N. Influence of salt on lipid oxidation in meat and seafood products: A review. Food Res. Int. 2017, 94, 90–100. [Google Scholar] [CrossRef]
- Cui, Y.; Hao, P.; Liu, B.; Meng, X. Effect of traditional Chinese cooking methods on fatty acid profiles of vegetable oils. Food Chem. 2017, 233, 77–84. [Google Scholar] [CrossRef] [PubMed]
- Cao, Y.; Ai, N.; True, A.D.; Xiong, Y.L. Effects of (−)-epigallocatechin-3-gallate incorporation on the physicochemical and oxidative stability of myofibrillar protein–soybean oil emulsions. Food Chem. 2018, 245, 439–445. [Google Scholar] [CrossRef] [PubMed]
- Cheng, L.; Li, X.; Tian, Y.; Wang, Q.; Li, X.; An, F.; Luo, Z.; Shang, P.; Liu, Z.; Huang, Q. Mechanisms of cooking methods on flavor formation of Tibetan pork. Food Chem. X 2023, 19, 100873. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Li, C.; Ullah, N.; Guo, Y.; Sun, X.; Wang, X.; Xu, X.; Hackman, R.M.; Zhou, G.; Feng, X. Different physicochemical, structural and digestibility characteristics of myofibrillar protein from PSE and normal pork before and after oxidation. Meat Sci. 2016, 121, 228–237. [Google Scholar] [CrossRef]
- Li, Q.; Sun, X.; Mubango, E.; Zheng, Y.; Liu, Y.; Zhang, Y.; Tan, Y.; Luo, Y.; Hong, H. Effects of protein and lipid oxidation on the water holding capacity of different parts of bighead carp: Eye, dorsal, belly and tail muscles. Food Chem. 2023, 423, 136238. [Google Scholar] [CrossRef] [PubMed]
- Hong, H.; Zhou, Y.; Wu, H.; Luo, Y.; Shen, H. Lipid content and fatty acid profile of muscle, brain and eyes of seven freshwater fish: A comparative study. J. Am. Oil Chem. Soc. 2014, 91, 795–804. [Google Scholar] [CrossRef]
- Jiang, Y.; Li, D.; Tu, J.; Zhong, Y.; Zhang, D.; Wang, Z.; Tao, X. Mechanisms of change in gel water-holding capacity of myofibrillar proteins affected by lipid oxidation: The role of protein unfolding and cross-linking. Food Chem. 2021, 344, 128587. [Google Scholar] [CrossRef] [PubMed]
- Liao, G.; Zhang, H.; Jiang, Y.; Javed, M.; Xiong, S.; Liu, Y. Effect of lipoxygenase-catalyzed linoleic acid oxidation on structural and rheological properties of silver carp (Hypophthalmichthys molitrix) myofibrillar protein. LWT-Food Sci. Technol. 2022, 161, 113388. [Google Scholar] [CrossRef]
- Zhou, F.; Zhao, M.; Cui, C.; Sun, W. Influence of linoleic acid-induced oxidative modifications on physicochemical changes and in vitro digestibility of porcine myofibrillar proteins. LWT-Food Sci. Technol. 2015, 61, 414–421. [Google Scholar] [CrossRef]
- Yu, T.Y.; Morton, J.D.; Clerens, S.; Dyer, J.M. Cooking-induced protein modifications in meat. Compr. Rev. Food Sci. Food Saf. 2017, 16, 141–159. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Zhu, J.; Zhang, H.; Chen, Q.; Kong, B. Understanding interactions among aldehyde compounds and porcine myofibrillar proteins by spectroscopy and molecular dynamics simulations. J. Mol. Liq. 2022, 349, 118190. [Google Scholar] [CrossRef]
- Yu, Y.; Wang, G.; Yin, X.; Ge, C.; Liao, G. Effects of different cooking methods on free fatty acid profile, water-soluble compounds and flavor compounds in Chinese Piao chicken meat. Food Res. Int. 2021, 149, 110696. [Google Scholar] [CrossRef]
- Zhang, L.; Li, Q.; Bao, Y.; Tan, Y.; Lametsch, R.; Hong, H.; Luo, Y. Recent advances on characterization of protein oxidation in aquatic products: A comprehensive review. Crit. Rev. Food Sci. Nutr. 2024, 64, 1572–1591. [Google Scholar] [CrossRef]
- Yu, M.; Fan, Y.; Liu, Y.; Yin, F.; Li, D.; Liu, X.; Zhou, D.; Zhu, B. Effects of antioxidants of bamboo leaves on protein digestion and transport of cooked abalone muscles. Food Funct. 2022, 13, 1785–1796. [Google Scholar] [CrossRef] [PubMed]
- Zhang, F.; Jiang, S.; Feng, X.; Wang, R.; Zeng, M.; Zhao, Y. Effect of heat treatment on physicochemical state and in vitro digestion of salt-soluble protein from Pacific oyster (Crassostrea gigas). LWT-Food Sci. Technol. 2020, 134, 110126. [Google Scholar] [CrossRef]
- Xia, C.; Wen, P.; Yuan, Y.; Yu, X.; Chen, Y.; Xu, H.; Cui, G.; Wang, J. Effect of roasting temperature on lipid and protein oxidation and amino acid residue side chain modification of beef patties. RSC Adv. 2021, 11, 21629–21641. [Google Scholar] [CrossRef] [PubMed]
- Huang, P.; Wang, Z.; Feng, X.; Kan, J. Promotion of fishy odor release by phenolic compounds through interactions with myofibrillar protein. Food Chem. 2022, 387, 132852. [Google Scholar] [CrossRef] [PubMed]
- Cao, Y.; Xiong, Y.L. Chlorogenic acid-mediated gel formation of oxidatively stressed myofibrillar protein. Food Chem. 2015, 180, 235–243. [Google Scholar] [CrossRef]
- Cheng, J.; Xu, L.; Xiang, R.; Liu, X.; Zhu, M. Effects of mulberry polyphenols on oxidation stability of sarcoplasmic and myofibrillar proteins in dried minced pork slices during processing and storage. Meat Sci. 2020, 160, 107973. [Google Scholar] [CrossRef] [PubMed]
- Zhang, D.; Li, H.; Wang, Z.; Emara, A.M.; Hu, Y.; He, Z. Effects of in vitro oxidation on myofibrillar protein charge, aggregation, and structural characteristics. Food Chem. 2020, 332, 127396. [Google Scholar] [CrossRef] [PubMed]
- Zhang, D.; Li, H.; Emara, A.M.; Hu, Y.; Wang, Z.; Wang, M.; He, Z. Effect of in vitro oxidation on the water retention mechanism of myofibrillar proteins gel from pork muscles. Food Chem. 2020, 315, 126226. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Hou, Y.; Liu, X.; Sun, J.; Wang, X.; Sang, Y. Improvement in the gelling properties of myofibrillar protein from the razor clam (Sinonovacula constricta) through phosphorylation and structural characterization of the modified protein. Food Chem. X 2023, 20, 101006. [Google Scholar] [CrossRef]
- Cheng, S.; Wang, X.; Li, R.; Yang, H.; Wang, H.; Wang, H.; Tan, M. Influence of multiple freeze-thaw cycles on quality characteristics of beef semimembranous muscle: With emphasis on water status and distribution by LF-NMR and MRI. Meat Sci. 2019, 147, 44–52. [Google Scholar] [CrossRef]
- Domínguez, R.; Pateiro, M.; Munekata, P.E.S.; Zhang, W.; Garcia-Oliveira, P.; Carpena, M.; Prieto, M.A.; Bohrer, B.; Lorenzo, J.M. Protein oxidation in muscle foods: A comprehensive review. Antioxidants 2022, 11, 60. [Google Scholar] [CrossRef] [PubMed]
- Gatellier, P.; Kondjoyan, A.; Portanguen, S.; Santé-Lhoutellier, V. Effect of cooking on protein oxidation in n-3 polyunsaturated fatty acids enriched beef. Implication on nutritional quality. Meat Sci. 2010, 85, 645–650. [Google Scholar] [CrossRef] [PubMed]
- Davies, K.J.A. Degradation of oxidized proteins by the 20S proteasome. Biochimie 2001, 83, 301–310. [Google Scholar] [CrossRef] [PubMed]
- Requena, J.R.; Fu, M.X.; Ahmed, M.U.; Jenkins, A.J.; Lyons, T.J.; Baynes, J.W.; Thorpe, S.R. Quantification of malondialdehyde and 4-hydroxynonenal adducts to lysine residues in native and oxidized human low-density lipoprotein. Biochem. J. 1997, 322, 317–325. [Google Scholar] [CrossRef] [PubMed]
- Ehrenshaft, M.; Deterding, L.J.; Mason, R.P. Tripping up Trp: Modification of protein tryptophan residues by reactive oxygen species, modes of detection, and biological consequences. Free Radic. Biol. Med. 2015, 89, 220–228. [Google Scholar] [CrossRef]
- Wang, K.; Luo, S.; Zhong, X.; Cai, J.; Jiang, S.; Zheng, Z. Changes in chemical interactions and protein conformation during heat-induced wheat gluten gel formation. Food Chem. 2017, 214, 393–399. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.; Wu, H.; Liu, M.; Liu, Z.; Xu, H.; Lai, F. Analysis of binding interaction between (−)-epigallocatechin (EGC) and β-lactoglobulin by multi-spectroscopic method. Spectrochim. Acta Part A Mol. Biomol. Spectrosc. 2011, 82, 164–168. [Google Scholar] [CrossRef]
- Shen, H.; Huang, M.; Zhao, M.; Sun, W. Interactions of selected ketone flavours with porcine myofibrillar proteins: The role of molecular structure of flavour compounds. Food Chem. 2019, 298, 125060. [Google Scholar] [CrossRef]
- Nyaisaba, B.M.; Hatab, S.; Liu, X.; Chen, Y.; Chen, X.; Miao, W.; Chen, M.; Deng, S. Physicochemical changes of myofibrillar proteins of squid (Argentinus ilex) induced by hydroxyl radical generating system. Food Chem. 2019, 297, 124941. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Zhang, M.; Fang, Z.; Bhandari, B.; Gao, Z. Influence of linoleic acid-induced oxidative modification on gel properties of myofibrillar protein from silver carp (Hypophthalmichthys molitrix) muscle. Food Biophys. 2016, 11, 266–274. [Google Scholar] [CrossRef]
- Liang, J.H. Fluorescence due to interactions of oxidizing soybean oil and soy proteins. Food Chem. 1999, 66, 103–108. [Google Scholar] [CrossRef]
- Zhang, W.; Xiao, S.; Ahn, D.U. Protein oxidation: Basic principles and implications for meat quality. Crit. Rev. Food Sci. Nutr. 2013, 53, 1191–1201. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; He, Z.; Wang, Z.; Li, H. Insight into the interaction of malondialdehyde with rabbit meat myofibrillar protein: Fluorescence quenching and protein oxidation. Foods 2023, 12, 2044. [Google Scholar] [CrossRef]
- Sante-Lhoutellier, V.; Aubry, L.; Gatellier, P. Effect of oxidation on in vitro digestibility of skeletal muscle myofibrillar proteins. J. Agric. Food Chem. 2007, 55, 5343–5348. [Google Scholar] [CrossRef] [PubMed]
- Runkana, V.; Somasundaran, P.; Kapur, P.C. Mathematical modeling of polymer-induced flocculation by charge neutralization. J. Colloid Interface Sci. 2004, 270, 347–358. [Google Scholar] [CrossRef]
- Cao, Y.; Ma, W.; Huang, J.; Xiong, Y.L. Effects of sodium pyrophosphate coupled with catechin on the oxidative stability and gelling properties of myofibrillar protein. Food Hydrocoll. 2020, 104, 105722. [Google Scholar] [CrossRef]
- Cai, L.; Nian, L.; Zhao, G.; Zhang, Y.; Sha, L.; Li, J. Effect of herring antifreeze protein combined with chitosan magnetic nanoparticles on quality attributes in red sea bream (Pagrosomus major). Food and Bioproc. Technol. 2019, 12, 409–421. [Google Scholar] [CrossRef]
- Dai, H.; Chen, X.; Peng, L.; Ma, L.; Sun, Y.; Li, L.; Wang, Q.; Zhang, Y. The mechanism of improved myosin gel properties by low dose rosmarinic acid addition during gel formation. Food Hydrocoll. 2020, 106, 105869. [Google Scholar] [CrossRef]
- Cao, H.; Jiao, X.; Fan, D.; Huang, J.; Zhao, J.; Yan, B.; Zhou, W.; Zhang, H.; Wang, M. Microwave irradiation promotes aggregation behavior of myosin through conformation changes. Food Hydrocoll. 2019, 96, 11–19. [Google Scholar] [CrossRef]
- Lange, R.; Balny, C. UV-visible derivative spectroscopy under high pressure. Biochim. Biophys. Acta 2002, 1595, 80–93. [Google Scholar] [CrossRef] [PubMed]
- Liu, F.; Sun, C.; Yang, W.; Yuan, F.; Gao, Y. Structural characterization and functional evaluation of lactoferrin–polyphenol conjugates formed by free-radical graft copolymerization. RSC Adv. 2015, 5, 15641–15651. [Google Scholar] [CrossRef]
- Sun, X.; Yu, Y.; Saleh, A.S.M.; Yang, X.; Ma, J.; Li, W.; Zhang, D.; Wang, Z. Understanding interactions among flavor compounds from spices and myofibrillar proteins by multi-spectroscopy and molecular docking simulation. Int. J. Biol. Macromol. 2023, 229, 188–198. [Google Scholar] [CrossRef]
- Zhou, F.; Zhao, M.; Su, G.; Cui, C.; Sun, W. Gelation of salted myofibrillar protein under malondialdehyde-induced oxidative stress. Food Hydrocoll. 2014, 40, 153–162. [Google Scholar] [CrossRef]
- Pei, Z.; Feng, Z.; Wang, H.; Xue, C. Preparation and characterization of Trachinotus ovatus myofibrillar protein emulsion gel. Sci. Technol. Food Ind. 2023, 44, 201–208. [Google Scholar] [CrossRef]
- Wu, W.; Jiang, Q.; Gao, P.; Yu, D.; Yu, P.; Xia, W. L-histidine-assisted ultrasound improved physicochemical properties of myofibrillar proteins under reduced-salt condition—Investigation of underlying mechanisms. Int. J. Biol. Macromol. 2023, 253, 126820. [Google Scholar] [CrossRef] [PubMed]
- Nie, X.; Liu, H.; Yu, N.; Tang, Q.; Wu, C.; Meng, X. Effect of high pressure homogenization on aggregation, conformation, and interfacial properties of bighead carp myofibrillar protein. J. Food Sci. 2021, 86, 5318–5328. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Zhao, G.; Yu, X.; Zhang, Q.; Zhu, C.; Tong, L.; Hao, J. Exploring in vitro gastrointestinal digestion of myofibrillar proteins at different heating temperatures. Food Chem. 2023, 414, 135694. [Google Scholar] [CrossRef]
- Chaijan, M.; Benjakul, S.; Visessanguan, W.; Faustman, C. Physicochemical properties, gel-forming ability and myoglobin content of sardine (Sardinella gibbosa) and mackerel (Rastrelliger kanagurta) surimi produced by conventional method and alkaline solubilisation process. Eur. Food Res. Technol. 2006, 222, 58–63. [Google Scholar] [CrossRef]
- Liu, X.; Feng, D.; Ji, L.; Zhang, T.; Xue, Y.; Xue, C. Effects of microwave heating on the gelation properties of heat-induced Alaska Pollock (Theragra chalcogramma) surimi. Food Sci. Technol. Int. 2018, 24, 497–506. [Google Scholar] [CrossRef]
- Yu, W.; Wang, Z.; Pan, Y.; Jiang, P.; Pan, J.; Yu, C.; Dong, X. Effect of κ-carrageenan on quality improvement of 3D printed Hypophthalmichthys molitrix-sea cucumber compound surimi product. LWT-Food Sci. Technol. 2022, 154, 112279. [Google Scholar] [CrossRef]
- Li, Y.; Li, X.; Wang, J.; Zhang, C.; Sun, H.; Wang, C.; Xie, X. Effects of oxidation on water distribution and physicochemical properties of porcine myofibrillar protein gel. Food Biophys. 2014, 9, 169–178. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Amino Acids/ | Concentration of OLA/(mM) | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| (mg/g Protein) | 0 | 2.5 | 5 | 7.5 | 10 | 0 + Boiling | 2.5 + Boiling | 5 + Boiling | 7.5 + Boiling | 10 + Boiling |
| Asp | 16.989 ± 0.402 a | 14.802 ± 0.571 b | 14.617 ± 0.700 bc | 13.852 ± 0.174 bcd | 12.558 ± 0.530 e | 15.924 ± 0.030 a | 13.820 ± 0.128 bcd | 13.589 ± 0.429 cde | 13.065 ± 0.076 de | 9.749 ± 0.962 f |
| Glu | 31.733 ± 0.942 a | 28.436 ± 1.210 bc | 27.294 ± 1.785 cd | 26.752 ± 0.765 cde | 24.683 ± 1.124 de | 30.514 ± 0.926 ab | 27.056 ± 2.162 cde | 25.623 ± 1.078 cde | 24.102 ± 0.377 e | 21.031 ± 1.500 f |
| Ser | 7.383 ± 0.260 a | 6.643 ± 0.472 b | 6.207 ± 0.327 bc | 6.038 ± 0.091 bcd | 5.656 ± 0.224 cd | 6.756 ± 0.276 ab | 6.054 ± 0.402 bcd | 5.637 ± 0.056 cd | 5.406 ± 0.057 d | 4.558 ± 0.424 e |
| His | 3.009 ± 0.101 a | 2.685 ± 0.141 bc | 2.579 ± 0.144 bcd | 2.526 ± 0.030 bcde | 2.408 ± 0.122 cde | 2.792 ± 0.190 ab | 2.543 ± 0.107 bcd | 2.320 ± 0.046 de | 2.229 ± 0.031 e | 1.808 ± 0.202 f |
| Gly | 7.532 ± 0.286 a | 6.424 ± 0.218 b | 5.939 ± 0.412 bcd | 5.655 ± 0.138 cde | 5.176 ± 0.284 ef | 6.268 ± 0.281 bc | 5.400 ± 0.237 def | 5.092 ± 0.093 ef | 4.980 ± 0.074 f | 4.161 ± 0.404 g |
| Thr | 8.814 ± 1.217 a | 8.321 ± 0.089 ab | 7.871 ± 0.710 ab | 7.851 ± 0.090 ab | 6.983 ± 0.098 bc | 8.085 ± 0.402 ab | 8.149 ± 0.930 ab | 7.441 ± 0.305 abc | 7.262 ± 0.482 bc | 6.117 ± 0.507 c |
| Arg | 10.716 ± 0.248 a | 9.137 ± 0.043 bc | 8.927 ± 0.547 bcd | 8.696 ± 0.098 cde | 8.125 ± 0.378 de | 9.738 ± 0.224 b | 8.611 ± 0.477 cde | 8.123 ± 0.026 de | 7.870 ± 0.102 e | 6.497 ± 0.661 f |
| Ala | 9.830 ± 0.344 a | 8.496 ± 0.154 b | 8.292 ± 0.514 bc | 8.071 ± 0.152 bc | 7.451 ± 0.330 c | 8.782 ± 0.083 b | 7.938 ± 0.560 bc | 7.452 ± 0.005 c | 7.387 ± 0.485 c | 5.922 ± 0.589 d |
| Tyr | 6.386 ± 0.088 a | 5.554 ± 0.337 bc | 5.403 ± 0.257 cd | 5.210 ± 0.052 cd | 5.038 ± 0.284 cd | 6.040 ± 0.302 ab | 5.297 ± 0.455 cd | 4.929 ± 0.032 d | 4.836 ± 0.102 d | 4.064 ± 0.204 e |
| Cys | 1.959 ± 0.235 a | 1.766 ± 0.023 ab | 1.642 ± 0.123 abc | 1.619 ± 0.086 abc | 1.468 ± 0.033 bcd | 1.734 ± 0.170 ab | 1.682 ± 0.170 ab | 1.534 ± 0.119 bcd | 1.263 ± 0.216 cd | 1.218 ± 0.229 d |
| Val | 7.910 ± 0.187 a | 6.863 ± 0.214 b | 6.703 ± 0.354 bc | 6.488 ± 0.124 bc | 6.082 ± 0.291 c | 7.193 ± 0.234 b | 6.485 ± 0.375 bc | 6.097 ± 0.050 c | 6.036 ± 0.371 c | 4.857 ± 0.477 d |
| Met | 4.509 ± 0.416 a | 3.001 ± 0.006 bc | 2.560 ± 0.064 cd | 2.252 ± 0.098 d | 2.235 ± 0.087 d | 3.287 ± 0.419 b | 2.547 ± 0.423 cd | 2.207 ± 0.421 d | 1.821 ± 0.320 de | 1.450 ± 0.330 e |
| Phe | 5.963 ± 0.120 a | 5.035 ± 0.025 b | 4.823 ± 0.127 b | 4.885 ± 0.094 b | 4.705 ± 0.352 b | 5.701 ± 0.272 a | 4.952 ± 0.344 b | 4.655 ± 0.089 b | 4.499 ± 0.148 b | 3.762 ± 0.313 c |
| Ile | 7.761 ± 0.419 a | 6.997 ± 0.253 bc | 6.557 ± 0.118 cd | 6.632 ± 0.097 cd | 6.122 ± 0.213 de | 7.310 ± 0.013 ab | 6.599 ± 0.343 cd | 6.261 ± 0.115 de | 5.870 ± 0.116 e | 4.947 ± 0.555 f |
| Leu | 13.835 ± 0.796 a | 12.165 ± 0.697 bc | 11.832 ± 0.986 bcd | 11.407 ± 0.018 bcd | 10.610 ± 0.504 d | 12.704 ± 0.496 ab | 11.410 ± 0.754 bcd | 10.766 ± 0.192 cd | 10.391 ± 0.406 d | 8.629 ± 0.767 e |
| Iys | 14.406 ± 0.023 a | 12.511 ± 0.588 bc | 11.862 ± 0.605 cd | 11.496 ± 0.441 cd | 11.081 ± 0.490 d | 13.220 ± 0.681 b | 11.726 ± 0.653 cd | 11.201 ± 0.040 d | 10.859 ± 0.123 d | 8.597 ± 0.815 e |
| Pro | 5.338 ± 0.664 a | 4.463 ± 0.033 ab | 4.171 ± 0.257 b | 4.044 ± 0.131 b | 3.737 ± 0.109 bc | 4.600 ± 0.680 ab | 3.988 ± 0.542 b | 3.715 ± 0.122 bc | 3.671 ± 0.141 bc | 2.868 ± 0.445 c |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, M.; Li, F.; Tang, Y.; Zhao, J.; Lei, X.; Ming, J. Effect of Boiling Treatment on Linoleic Acid-Induced Oxidation of Myofibrillar Protein in Grass Carp. Foods 2024, 13, 4153. https://doi.org/10.3390/foods13244153
Liu M, Li F, Tang Y, Zhao J, Lei X, Ming J. Effect of Boiling Treatment on Linoleic Acid-Induced Oxidation of Myofibrillar Protein in Grass Carp. Foods. 2024; 13(24):4153. https://doi.org/10.3390/foods13244153
Chicago/Turabian StyleLiu, Mengcong, Fuhua Li, Yuan Tang, Jichun Zhao, Xiaojuan Lei, and Jian Ming. 2024. "Effect of Boiling Treatment on Linoleic Acid-Induced Oxidation of Myofibrillar Protein in Grass Carp" Foods 13, no. 24: 4153. https://doi.org/10.3390/foods13244153
APA StyleLiu, M., Li, F., Tang, Y., Zhao, J., Lei, X., & Ming, J. (2024). Effect of Boiling Treatment on Linoleic Acid-Induced Oxidation of Myofibrillar Protein in Grass Carp. Foods, 13(24), 4153. https://doi.org/10.3390/foods13244153