
Antilisterial Properties of Selected Strains from the Autochthonous Microbiota of a Swiss Artisan Soft Smear Cheese

 , , , , , , and
, , , , , , and
Abstract
1. Introduction
2. Materials and Methods
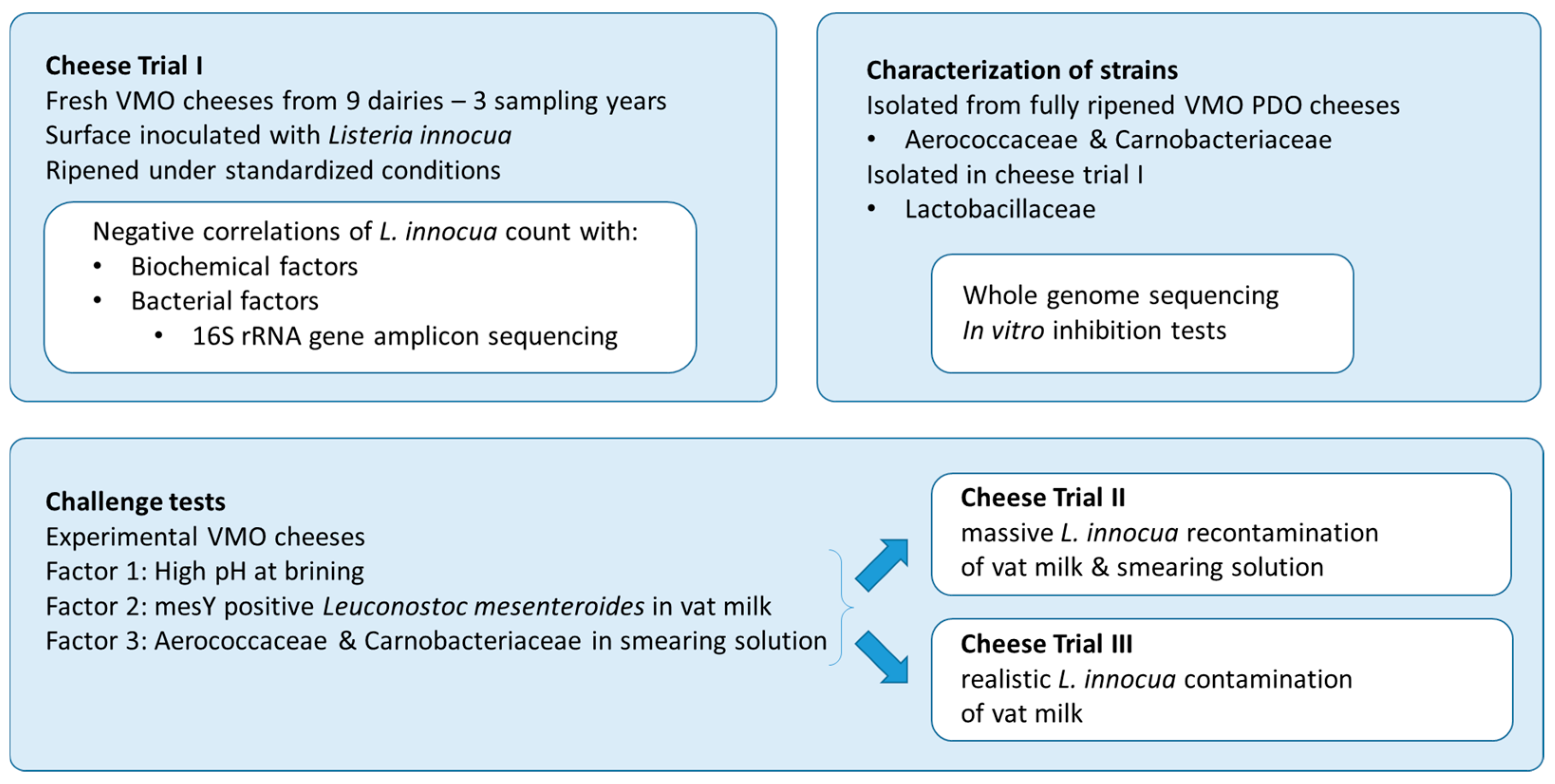
2.1. Experimental Design
2.2. Cheese Trial I
2.3. Listeria innocua Detection
2.4. Cheese Rind Microbiota Assessment
2.5. Biochemical Parameters
2.6. Isolation of Bacterial Strains
2.7. In Vitro Inhibition Tests
2.8. Whole-Genome Sequencing (WGS) and In Silico Analysis
2.9. Challenge Tests
2.9.1. Cheese Trial II
2.9.2. Cheese Trial III
2.10. Cultivation of Bacterial Strains for the Cheese Trials
2.11. Real-Time PCR Quantification (qPCR) in the Cheese Matrix
2.12. Statistical Analysis
3. Results
3.1. Cheese Trial I: Factors That Influence L. innocua Growth in the Rind of VMO Cheese
3.1.1. The Rind of VMO Cheese Supports the Growth of L. innocua During Ripening in a Facility-Dependent Manner
3.1.2. L. innocua Counts Correlate with Multiple Biochemical Parameters
3.1.3. Development of a Rich Autochthonous Microbiota During Ripening in an Experimental Cellar, Including Seven ASVs Negatively Correlated with L. innocua Counts
3.2. Isolation and Characterization of Putative Antagonistic Strains
3.2.1. Aerococcaceae and Carnobacteriaceae Show Strain-Specific Antilisterial Properties
3.2.2. Lactobacillaceae Antilisterial Properties
3.3. Challenge Tests at the Pilot Scale: The Effect of pH at Brining and the Addition of Protective Cultures in the Vat Milk/Smearing Solution
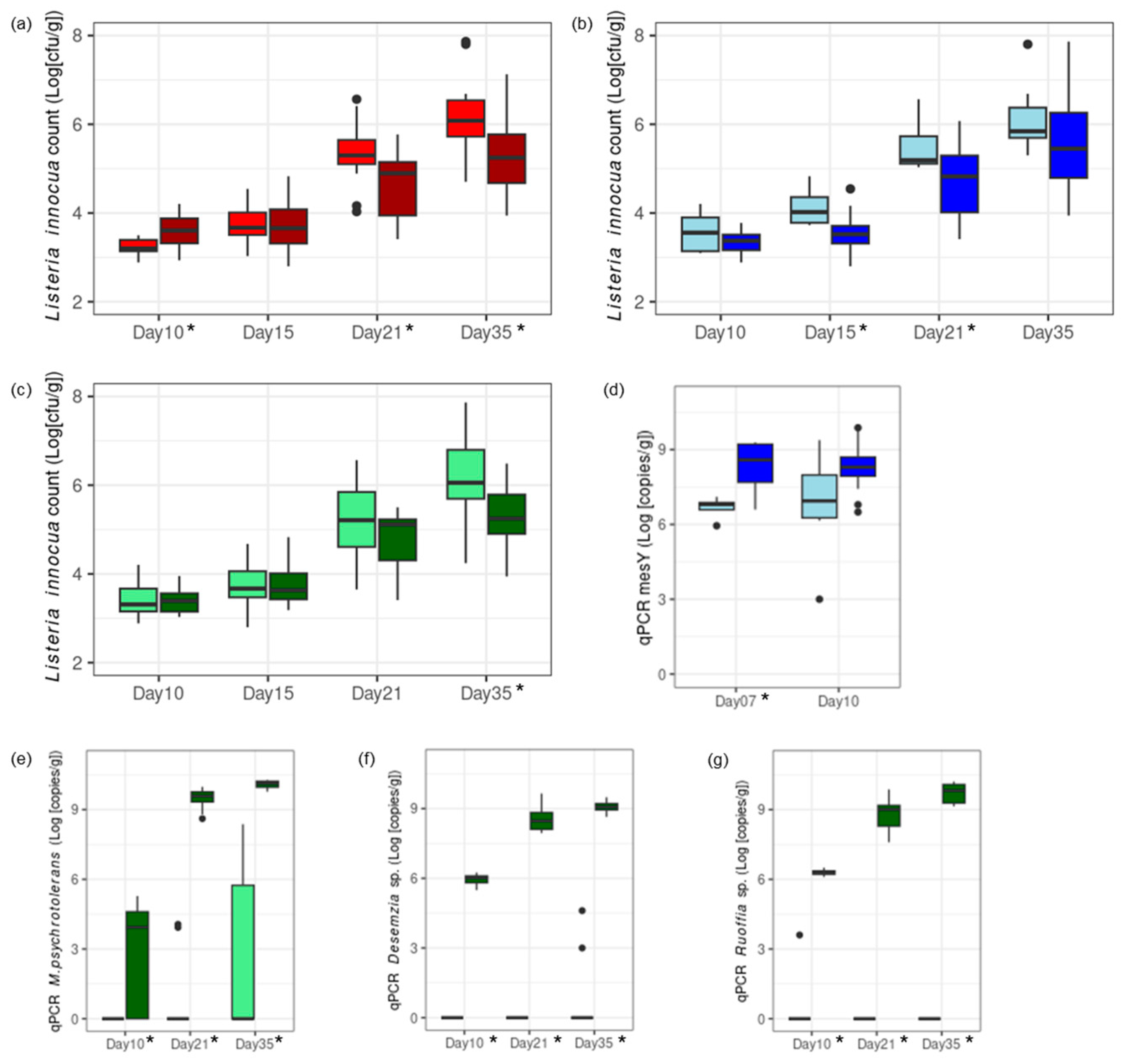
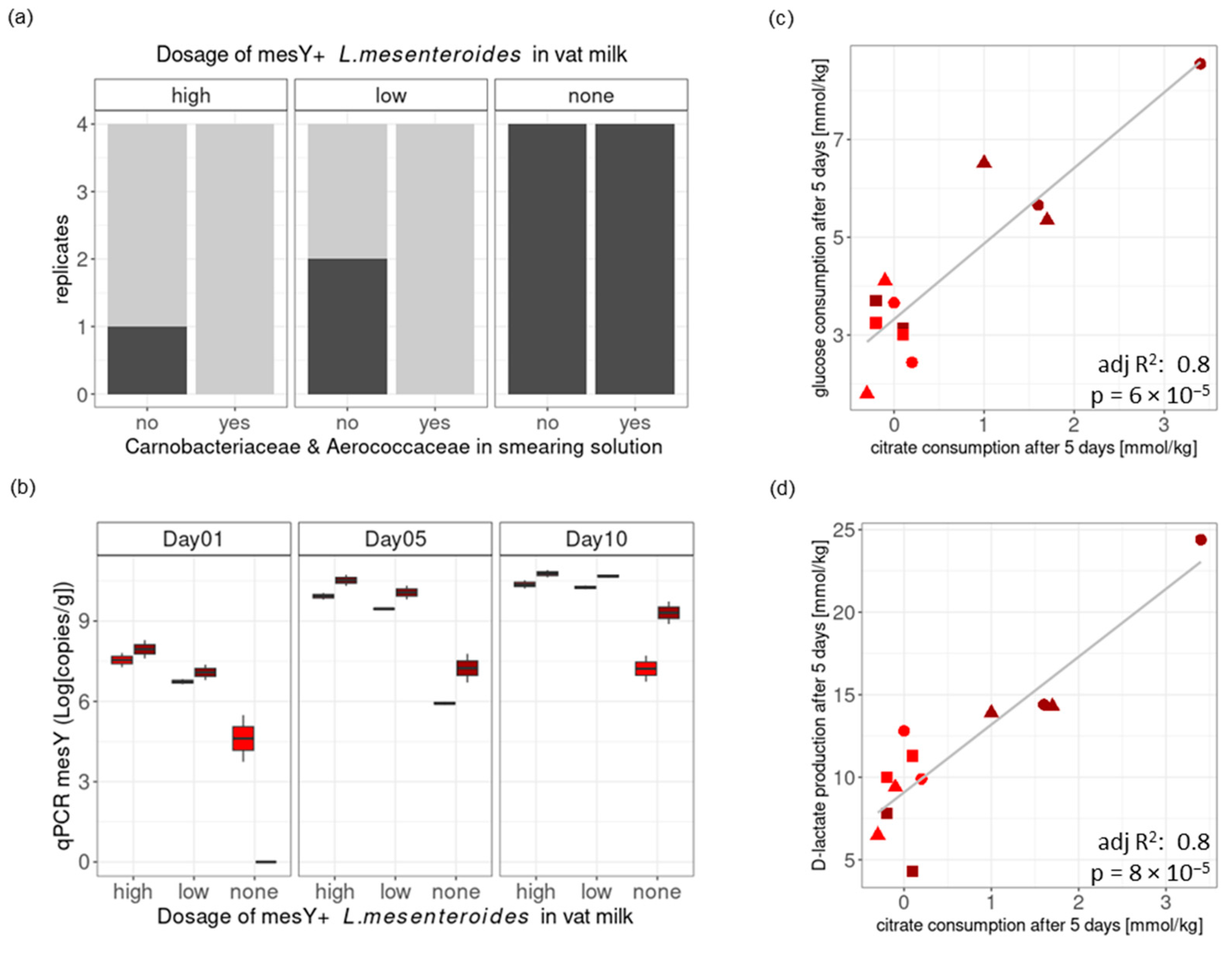
3.3.1. Worst-Case Scenario: Cheese Trials II: Conditions Mimicking a Massive Listeria Recontamination of the Vat Milk and the Ripening Facility
3.3.2. Realistic Scenario: Cheese Trial III—Conditions Mimicking a Low Listeria Level Remaining in Vat Milk After the Thermization of Contaminated Raw Milk
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kennedy, M.S.; Chang, E.B. The microbiome: Composition and locations. Prog. Mol. Biol. Transl. Sci. 2020, 176, 1–42. [Google Scholar] [CrossRef]
- Averill, C.; Anthony, M.A.; Baldrian, P.; Finkbeiner, F.; Hoogen, J.v.D.; Kiers, T.; Kohout, P.; Hirt, E.; Smith, G.R.; Crowther, T.W. Defending Earth’s terrestrial microbiome. Nat. Microbiol. 2022, 7, 1717–1725. [Google Scholar] [CrossRef]
- Galimberti, A.; Bruno, A.; Agostinetto, G.; Casiraghi, M.; Guzzetti, L.; Labra, M. Fermented food products in the era of globalization: Tradition meets biotechnology innovations. Curr. Opin. Biotechnol. 2021, 70, 36–41. [Google Scholar] [CrossRef]
- Mannaa, M.; Han, G.; Seo, Y.-S.; Park, I. Evolution of Food Fermentation Processes and the Use of Multi-Omics in Deciphering the Roles of the Microbiota. Foods 2021, 10, 2861. [Google Scholar] [CrossRef]
- Louw, N.L.; Lele, K.; Ye, R.; Edwards, C.B.; Wolfe, B.E. Microbiome Assembly in Fermented Foods. Annu. Rev. Microbiol. 2023, 77, 381–402. [Google Scholar] [CrossRef]
- Sooresh, M.M.; Willing, B.P.; Bourrie, B.C.T. Opportunities and Challenges of Understanding Community Assembly in Spontaneous Food Fermentation. Foods 2023, 12, 673. [Google Scholar] [CrossRef]
- Quigley, L.; O’Sullivan, O.; Beresford, T.P.; Ross, R.P.; Fitzgerald, G.F.; Cotter, P.D. High-Throughput sequencing for detection of subpopulations of bacteria Not previously associated with artisanal cheeses. Appl. Environ. Microbiol. 2012, 78, 5717–5723. [Google Scholar] [CrossRef]
- Irlinger, F.; Layec, S.; Hélinck, S.; Dugat-Bony, E. Cheese rind microbial communities: Diversity, composition and origin. FEMS Microbiol. Lett. 2015, 362, 1–11. [Google Scholar] [CrossRef]
- Dreier, M.; Meola, M.; Berthoud, H.; Shani, N.; Wechsler, D.; Junier, P. High-throughput qPCR and 16S rRNA gene amplicon sequencing as complementary methods for the investigation of the cheese microbiota. BMC Microbiol. 2022, 22, 48. [Google Scholar] [CrossRef]
- Breitenwieser, F.; Doll, E.V.; Clavel, T.; Scherer, S.; Wenning, M. Complementary use of cultivation and high-throughput amplicon sequencing reveals high biodiversity within raw milk microbiota. Front. Microbiol. 2020, 11, 1557. [Google Scholar] [CrossRef]
- Vermote, L.; Verce, M.; De Vuyst, L.; Weckx, S. Amplicon and shotgun metagenomic sequencing indicates that microbial ecosystems present in cheese brines reflect environmental inoculation during the cheese production process. Int. Dairy J. 2018, 87, 44–53. [Google Scholar] [CrossRef]
- Quijada, N.M.; Mann, E.; Wagner, M.; Rodríguez-Lázaro, D.; Hernández, M.; Schmitz-Esser, S. Autochthonous facility-specific microbiota dominates washed-rind Austrian hard cheese surfaces and its production environment. Int. J. Food Microbiol. 2018, 267, 54–61. [Google Scholar] [CrossRef] [PubMed]
- Wolfe, B.E.; Button, J.E.; Santarelli, M.; Dutton, R.J. Cheese rind communities provide tractable systems for in situ and in vitro studies of microbial diversity. Cell 2014, 158, 422–433. [Google Scholar] [CrossRef] [PubMed]
- Dugat-Bony, E.; Garnier, L.; Denonfoux, J.; Ferreira, S.; Sarthou, A.-S.; Bonnarme, P.; Irlinger, F. Highlighting the microbial diversity of 12 French cheese varieties. Int. J. Food Microbiol. 2016, 238, 265–273. [Google Scholar] [CrossRef]
- Montel, M.-C.; Buchin, S.; Mallet, A.; Delbes-Paus, C.; Vuitton, D.A.; Desmasures, N.; Berthier, F. Traditional cheeses: Rich and diverse microbiota with associated benefits. Int. J. Food Microbiol. 2014, 177, 136–154. [Google Scholar] [CrossRef]
- Cocolin, L.; Foschino, R.; Comi, G.; Grazia Fortina, M. Description of the bacteriocins produced by two strains of Enterococcus faecium isolated from Italian goat milk. Food Microbiol. 2007, 24, 752–758. [Google Scholar] [CrossRef]
- Roth, E.; Miescher Schwenninger, S.; Hasler, M.; Eugster-Meier, E.; Lacroix, C. Population dynamics of two antilisterial cheese surface consortia revealed by temporal temperature gradient gel electrophoresis. BMC Microbiol. 2010, 10, 74. [Google Scholar] [CrossRef]
- Lianou, A.; Samelis, J. Addition to thermized milk of Lactococcus lactis subsp. cremoris M104, a wild, novel nisin a-producing strain, replaces the natural antilisterial activity of the autochthonous raw milk microbiota reduced by thermization. J. Food Prot. 2014, 77, 1289–1297. [Google Scholar] [CrossRef]
- Callon, C.; Arliguie, C.; Montel, M.-C. Control of Shigatoxin-producing Escherichia coli in cheese by dairy bacterial strains. Food Microbiol. 2016, 53, 63–70. [Google Scholar] [CrossRef]
- Bourdichon, F.; Arias, E.; Babuchowski, A.; Bückle, A.; Bello, F.D.; Dubois, A.; Fontana, A.; Fritz, D.; Kemperman, R.; Laulund, S.; et al. The forgotten role of food cultures. FEMS Microbiol. Lett. 2021, 368, fnab085. [Google Scholar] [CrossRef]
- Mayo, B.; Rodríguez, J.; Vázquez, L.; Flórez, A.B. Microbial interactions within the cheese ecosystem and their application to improve quality and safety. Foods 2021, 10, 602. [Google Scholar] [CrossRef] [PubMed]
- Callon, C.; Saubusse, M.; Didienne, R.; Buchin, S.; Montel, M.-C. Simplification of a complex microbial antilisterial consortium to evaluate the contribution of its flora in uncooked pressed cheese. Int. J. Food Microbiol. 2011, 145, 379–389. [Google Scholar] [CrossRef] [PubMed]
- Roth, E.; Schwenninger, S.M.; Eugster-Meier, E.; Lacroix, C. Facultative anaerobic halophilic and alkaliphilic bacteria isolated from a natural smear ecosystem inhibit Listeria growth in early ripening stages. Int. J. Food Microbiol. 2011, 147, 26–32. [Google Scholar] [CrossRef]
- Gérard, A.; El-Hajjaji, S.; Burteau, S.; Fall, P.A.; Pirard, B.; Taminiau, B.; Daube, G.; Sindic, M. Study of the microbial diversity of a panel of Belgian artisanal cheeses associated with challenge studies for Listeria monocytogenes. Food Microbiol. 2021, 100, 103861. [Google Scholar] [CrossRef]
- EFSA Panel on Biological Hazards (BIOHAZ); Ricci, A.; Allende, A.; Bolton, D.; Chemaly, M.; Davies, R.; Fernández Escámez, P.S.; Girones, R.; Herman, L.; Koutsoumanis, K.; et al. Listeria monocytogenes contamination of ready-to-eat foods and the risk for human health in the EU. Eur. Food Saf. Auth. J. 2018, 16, e05134. [Google Scholar] [CrossRef]
- Maury, M.M.; Bracq-Dieye, H.; Huang, L.; Vales, G.; Lavina, M.; Thouvenot, P.; Disson, O.; Leclercq, A.; Brisse, S.; Lecuit, M. Hypervirulent Listeria monocytogenes clones’ adaption to mammalian gut accounts for their association with dairy products. Nat. Commun. 2019, 10, 2488. [Google Scholar] [CrossRef]
- Martinez-Rios, V.; Dalgaard, P. Prevalence of Listeria monocytogenes in European cheeses: A systematic review and meta-analysis. Food Control 2018, 84, 205–214. [Google Scholar] [CrossRef]
- Rudolf, M.; Scherer, S. High incidence of Listeria monocytogenes in European red smear cheese. Int. J. Food Microbiol. 2001, 63, 91–98. [Google Scholar] [CrossRef]
- Zangerl, P.; Schoder, D.; Eliskases-Lechner, F.; Zangana, A.; Frohner, E.; Stessl, B.; Wagner, M. Monitoring by a sensitive liquid-based sampling strategy reveals a considerable reduction of Listeria monocytogenes in smeared cheese production over 10 years of testing in Austria. Foods 2021, 10, 1977. [Google Scholar] [CrossRef]
- Eugster, E.; Jakob, E. Pre-treatments of milk and their effect on the food safety of cheese. Milk Sci. Int. 2019, 72, 45–52. [Google Scholar] [CrossRef]
- Bille, J. Epidemiology of human listeriosis in Europe with special reference to the Swiss outbreak. In Foodborne Listeriosis; Miller, A.J., Smith, J.L., Somkuti, G.A., Eds.; Elsevier: Amsterdam, The Netherlands, 1990; pp. 71–74. [Google Scholar]
- Carminati, D.; Perrone, A.; Neviani, E.; Mucchetti, G.; Carminati, A.P.D.; Gatti, M.; Bonvini, B. Influence of traditional brine washing of smear taleggio cheese on the surface spreading of Listeria innocua. J. Food Prot. 2000, 63, 1353–1358. [Google Scholar] [CrossRef] [PubMed]
- Hammer, P.; Bockelmann, W.; Hoffmann, W. Fate of listeria innocua during production and ripening of smeared hard cheese made from raw milk. J. Dairy Sci. 2017, 100, 7846–7856. [Google Scholar] [CrossRef] [PubMed]
- Berthoud, H.; Wüthrich, D.; Bruggmann, R.; Wechsler, D.; Fröhlich-Wyder, M.-T.; Irmler, S. Development of new methods for the quantitative detection and typing of lactobacillus parabuchneri in dairy products. Int. Dairy J. 2017, 70, 65–71. [Google Scholar] [CrossRef]
- Meola, M.; Rifa, E.; Shani, N.; Delbès, C.; Berthoud, H.; Chassard, C. DAIRYdb: A manually curated reference database for improved taxonomy annotation of 16S rRNA gene sequences from dairy products. BMC Genom. 2019, 20, 560. [Google Scholar] [CrossRef] [PubMed]
- IDF Standard 4; Cheese and Processed Cheese. Determination of the Total Solids Content. International Dairy Federation: Brussels, Belgium, 2004.
- Badertscher, R.; Blaser, C.; Noth, P. Validated method for the determination of free volatile carboxylic acids in cheese and bacterial cultures by GC-FID after esterification in aqueous extract and headspace injection. Food Chem. 2023, 398, 133932. [Google Scholar] [CrossRef]
- Kuhnert, P.; Capaul, S.E.; Nicolet, J.; Frey, J. Phylogenetic positions of clostridium chauvoei and clostridium septicum based on 16S rRNA gene sequences. Int. J. Syst. Evol. Microbiol. 1996, 46, 1174–1176. [Google Scholar] [CrossRef]
- Versalovic, J.; Schneider, M.; de Bruijn, F.J.; Lupsiki, J.R. Genomic fingerprinting of bacteria using repetitive sequence-based polymerase chain reaction. Methods Mol. Cell. Biol. 1994, 5, 24–40. [Google Scholar]
- Singhal, N.; Kumar, M.; Kanaujia, P.K.; Virdi, J.S. MALDI-TOF mass spectrometry: An emerging technology for microbial identification and diagnosis. Front. Microbiol. 2015, 6, 791. [Google Scholar] [CrossRef]
- Tagg, J.R.; McGiven, A.R. Assay system for bacteriocins. Appl. Microbiol. 1971, 21, 943. [Google Scholar] [CrossRef]
- Shani, N.; Oberhaensli, S.; Arias-Roth, E. Antibiotic susceptibility profiles of Pediococcus pentosaceus from various origins and their implications for the safety assessment of strains with food-technology applications. J. Food Prot. 2021, 84, 1160–1168. [Google Scholar] [CrossRef]
- Bankevich, A.; Nurk, S.; Antipov, D.; Gurevich, A.A.; Dvorkin, M.; Kulikov, A.S.; Lesin, V.M.; Nikolenko, S.I.; Pham, S.; Prjibelski, A.D.; et al. SPAdes: A new genome assembly algorithm and its applications to single-cell sequencing. J. Comput. Biol. 2012, 19, 455–477. [Google Scholar] [CrossRef] [PubMed]
- Richter, M.; Rosselló-Móra, R.; Oliver Glöckner, F.O.; Peplies, J. JSpeciesWS: A web server for prokaryotic species circumscription based on pairwise genome comparison. Bioinformatics 2016, 32, 929–931. [Google Scholar] [CrossRef] [PubMed]
- Meier-Kolthoff, J.P.; Göker, M. TYGS is an automated high-throughput platform for state-of-the-art genome-based taxonomy. Nat. Commun. 2019, 10, 2182. [Google Scholar] [CrossRef] [PubMed]
- Medema, M.H.; Blin, K.; Cimermancic, P.; De Jager, V.; Zakrzewski, P.; Fischbach, M.A.; Weber, T.; Takano, E.; Breitling, R. antiSMASH: Rapid identification, annotation, and analysis of secondary metabolite biosynthesis gene clusters in bacterial and fungal genome sequences. Nucleic Acids Res. 2011, 39, W339–W346. [Google Scholar] [CrossRef]
- Van Heel, A.J.; De Jong, A.; Song, C.; Viel, J.H.; Kok, J.; Kuipers, O.P. BAGEL4: A user-friendly web server to thoroughly mine RiPPs and bacteriocins. Nucleic Acids Res. 2018, 46, W278–W281. [Google Scholar] [CrossRef]
- Moser, A.; Berthoud, H.; Eugster, E.; Meile, L.; Irmler, S. Detection and enumeration of lactobacillus helveticus in dairy products. Int. Dairy J. 2017, 68, 52–59. [Google Scholar] [CrossRef]
- Pinheiro, J.; Bates, D.; R Core Team. nlme: Linear and Nonlinear Mixed Effects Models. R Package Version 3.1-162. 2023. Available online: https://CRAN.R-project.org/package=nlme (accessed on 22 November 2023).
- Anderson, M.J.; Gorley, R.N.; Clarke, K.R. PERMANOVA+ for PRIMER: Guide to Software and Statistical Methods; Primer-E: Plymouth, UK, 2008; pp. 1–214. [Google Scholar]
- Korena, K.; Krzyzankova, M.; Florianova, M.; Karasova, D.; Babak, V.; Strakova, N.; Juricova, H. Microbial succession in the cheese ripening process-Competition of the starter cultures and the microbiota of the cheese plant environment. Microorganisms 2023, 11, 1735. [Google Scholar] [CrossRef]
- Bokulich, N.A.; Mills, D.A. Facility-specific “house” microbiome drives microbial landscapes of artisan cheesemaking plants. Appl. Environ. Microbiol. 2013, 79, 5214–5223. [Google Scholar] [CrossRef]
- Callon, C.; Retureau, E.; Didienne, R.; Montel, M.C. Microbial biodiversity in cheese consortia and comparative listeria growth on surfaces of uncooked pressed cheeses. Int. J. Food Microbiol. 2014, 174, 98–109. [Google Scholar] [CrossRef]
- Imran, M.; Desmasures, N.; Vernoux, J.-P. From undefined red smear cheese consortia to minimal model communities both exhibiting similar anti-listerial activity on a cheese-like matrix. Food Microbiol. 2010, 27, 1095–1103. [Google Scholar] [CrossRef]
- Imran, M.; Bré, J.-M.; Guéguen, M.; Vernoux, J.-P.; Desmasures, N. Reduced growth of Listeria monocytogenes in two model cheese microcosms is not associated with individual microbial strains. Food Microbiol. 2013, 33, 30–39. [Google Scholar] [CrossRef] [PubMed]
- Aucher, W.; Simonet, V.; Fremaux, C.; Dalet, K.; Simon, L.; Cenatiempo, Y.; Frère, J.; Berjeaud, J.M. Differences in mesentericin secretion systems from two leuconostoc strains. FEMS Microbiol. Lett. 2004, 232, 15–22. [Google Scholar] [CrossRef] [PubMed]
- Muchaamba, F.; Eshwar, A.K.; Stevens MJ, A.; von Ah, U.; Tasara, T. Variable carbon source utilization, stress resistance, and virulence profiles among Listeria monocytogenes strains responsible for listeriosis outbreaks in Switzerland. Front. Microbiol. 2019, 10, 957. [Google Scholar] [CrossRef] [PubMed]
- Kastman, E.K.; Kamelamela, N.; Norville, J.W.; Cosetta, C.M.; Dutton, R.J.; Wolfe, B.E. Biotic interactions shape the ecological distributions of staphylococcus species. mBio 2016, 7, e01157-16. [Google Scholar] [CrossRef] [PubMed]
- Pierce, E.C.; Morin, M.; Little, J.C.; Liu, R.B.; Tannous, J.; Keller, N.P.; Pogliano, K.; Wolfe, B.E.; Sanchez, L.M.; Dutton, R.J. Bacterial–fungal interactions revealed by genome-wide analysis of bacterial mutant fitness. Nat. Microbiol. 2021, 6, 87–102. [Google Scholar] [CrossRef]
- Wemmenhove, E.; Wells-Bennik MH, J.; Zwietering, M.H. A model to predict the fate of listeria monocytogenes in different cheese types—A major role for undissociated lactic acid in addition to pH, water activity, and temperature. Int. J. Food Microbiol. 2021, 357, 109350. [Google Scholar] [CrossRef]
- Adams, T.J.; Vartivarian, S.; E Cowart, R. Iron acquisition systems of Listeria monocytogenes. Infect. Immun. 1990, 58, 2715–2718. [Google Scholar] [CrossRef]
- Lanciotti, R.; Patrignani, F.; Bagnolini, F.; Guerzoni, M.; Gardini, F. Evaluation of diacetyl antimicrobial activity against Escherichia coli, Listeria monocytogenes and Staphylococcus aureus. Food Microbiol. 2003, 20, 537–543. [Google Scholar] [CrossRef]
- Özcan, E.; Selvi, S.S.; Nikerel, E.; Teusink, B.; Toksoy Öner, E.; Çakır, T. A genome-scale metabolic network of the aroma bacterium leuconostoc mesenteroides subsp. cremoris. Appl. Microbiol. Biotechnol. 2019, 103, 3153–3165. [Google Scholar] [CrossRef]
- Budde, B.B.; Hornbæk, T.; Jacobsen, T.; Barkholt, V.; Koch, A.G. Leuconostoc carnosum 4010 has the potential for use as a protective culture for vacuum-packed meats: Culture isolation, bacteriocin identification, and meat application experiments. Int. J. Food Microbiol. 2003, 83, 171–184. [Google Scholar] [CrossRef]
- Colás-Medà, P.; Viñas, I.; Alegre, I. Evaluation of commercial anti-listerial products for improvement of food safety in ready-to-eat meat and dairy products. Antibiotics 2023, 12, 414. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Factor | r | p-Value | adj. p-Value | |
|---|---|---|---|---|
| Biochemical | ||||
| Acetic acid at 23 days | −0.66 | 0.000 | 0.013 | * |
| Lactose in fresh cheese | −0.63 | 0.001 | 0.013 | * |
| Glucose at 10 days | −0.62 | 0.001 | 0.013 | * |
| Citrate at 10 days | 0.55 | 0.005 | 0.029 | * |
| L-lactate in fresh cheese | 0.61 | 0.002 | 0.013 | * |
| Lactate in fresh cheese | 0.62 | 0.001 | 0.013 | * |
| Bacterial | ||||
| ASV_005 Leuconostoc carnosum/mesenteroides | −0.82 | 0.000 | 0.004 | ** |
| ASV_212 Weissella hellenica | −0.79 | 0.001 | 0.010 | * |
| ASV_147 Latilactobacillus curvatus | −0.77 | 0.001 | 0.017 | * |
| ASV_110 Latilactobacillus curvatus | −0.74 | 0.002 | 0.031 | * |
| ASV_068 Marinomonas sp./flavescens/ushuaiensis | −0.73 | 0.002 | 0.032 | * |
| ASV_227 Celerinatantimonas | −0.73 | 0.002 | 0.032 | * |
| ASV_007 Leuconostoc carnosum/mesenteroides | −0.71 | 0.003 | 0.047 | * |
| Df | R2 | Pseudo-F | p-Value | √CV 1 | |
|---|---|---|---|---|---|
| Dairy | 8 | 39.6% | 1.1 | 0.3703 | 0.09 |
| Replication | 6 | 27.5% | 9.9 | 0.0001 | 0.39 |
| Cheese ripening and at storage | 3 | 13.4% | 9.7 | 0.0001 | 0.20 |
| Residuals | 42 | 19.4% | 0.26 | ||
| Total | 59 | 100% |
| Strain 1 | Source 2 | Dairy | Inhibition of L. innocua by Live Bacteria 3 | Putative Bacteriocin Genes 4 |
|---|---|---|---|---|
| Aerococcaceae | ||||
| Ruoffia sp. | ||||
| FAM 24227 * | VMO | E | +++ | B1 |
| Carnobacteriaceae | ||||
| Desemzia sp. | ||||
| FAM 23990 | VMO | B | + | B2 |
| FAM 23991 | VMO | B | + | no |
| FAM 24101 * | VMO | D | +++ | no |
| FAM 23989 | VMO | F | ++ | no |
| FAM 23988 | VMO | H | + | B3 |
| Marinilactibacillus psychrotolerans | ||||
| FAM 24229 | VMO | A | ++ | no |
| FAM 23992 | VMO | D | (+) | no |
| FAM 23995 | VMO | F | (+) | B4 |
| FAM 23631 | VMO | G | + | B4, B5, B6 |
| FAM 23997 * | VMO | G | ++ | B7 |
| FAM 23998 | VMO | G | (+) | B8 |
| FAM 23993 | VMO | H | + | no |
| FAM 24102 | VMO | H | + | B7 |
| FAM 24106 | VMO | I | ++ | no |
| FAM 24231 | Cheese brine | G | B8 | |
| FAM 24230 | Cheese brine | I | no | |
| Strain 1 | Dairy | Inhibition of L. innocua by Supernatant 3 | Mesentericin Y105 Operon 4 | Other Putative Bacteriocin 5 | Citrate 6 Operon |
|---|---|---|---|---|---|
| Lacticaseibacillus paracasei | |||||
| FAM 25336 | D | (−) | |||
| Lactiplantibacillus plantarum | |||||
| FAM 25263 | D | (−) | 0 | B9 | incomplete |
| Latilactobacillus curvatus | |||||
| FAM 25311 | A | (−) | |||
| FAM 24637 | D | (−) | 0 | no | no |
| FAM 25309 | D | (−) | |||
| FAM 25316 | E | (−) | |||
| FAM 25313 | G | (−) | |||
| FAM 25314 | H | (−) | 0 | B10 | yes (>52%) |
| FAM 25315 | H | (−) | |||
| Latilactobacillus sakei | |||||
| FAM 24915 | E | (−) | 0 | B11 | yes (>53%) |
| FAM 24914 | G | (−) | 0 | B11 | yes (>53%) |
| FAM 24916 | I | (−) | 0 | no | incomplete |
| Leuconostoc carnosum | |||||
| FAM 24634 | D | (−) | 0 | B12 | yes (>67%) |
| FAM 24918 | D | (−) | |||
| FAM 25319 | D | (−) | |||
| FAM 25322 | G | (−) | |||
| FAM 25323 | G | (−) | |||
| FAM 24917 | H | (−) | |||
| FAM 24919 | I | (−) | |||
| Leuconostoc mesenteroides | |||||
| FAM 25299 | A | (−) | 0 | B13 | no |
| FAM 25300 * | A | + | 100 | B13 | no |
| FAM 24636 | D | + | 100 | B13 | yes (>99%) |
| FAM 25292 * | D | + | 100 | B13 | yes (>99%) |
| FAM 25293 * | D | ++ | 100 | B13 | yes (>99%) |
| FAM 25295 | D | + | 100 | B13 | no |
| FAM 25301 | G | (−) | 0 | B13 | no |
| FAM 25302 | G | (−) | 0 | B13 | no |
| FAM 25281 | H | (−) | 0 | B13 | no |
| FAM 25285 | I | (+) | 0 | B13 | no |
| FAM 24179 *,2 | - | +++ | 100 | B14 | yes (>98%) |
| Limosilactobacillus fermentum | |||||
| FAM 25337 | D | (−) | 0 | B15, B16 | incomplete |
| FAM 25338 | H | (−) | 0 | B15, B16 | no |
| Loigolactobacillus zhaoyuanensis | |||||
| FAM 25317 | E | (−) | 0 | B17 | incomplete |
| Weissella hellenica | |||||
| FAM 25328 | D | (−) | |||
| FAM 25329 | F | (−) | |||
| FAM 25330 | F | (−) | 0 | no | no |
| FAM 25332 | G | (−) | 0 | no | no |
| FAM 25333 | G | (−) | |||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Roetschi, A.; Baumeyer, A.; Berthoud, H.; Braillard, L.; Gschwend, F.; Guisolan, A.; Haldemann, J.; Hummerjohann, J.; Joller, C.; Loosli, F.; et al. Antilisterial Properties of Selected Strains from the Autochthonous Microbiota of a Swiss Artisan Soft Smear Cheese. Foods 2024, 13, 3473. https://doi.org/10.3390/foods13213473
Roetschi A, Baumeyer A, Berthoud H, Braillard L, Gschwend F, Guisolan A, Haldemann J, Hummerjohann J, Joller C, Loosli F, et al. Antilisterial Properties of Selected Strains from the Autochthonous Microbiota of a Swiss Artisan Soft Smear Cheese. Foods. 2024; 13(21):3473. https://doi.org/10.3390/foods13213473
Chicago/Turabian StyleRoetschi, Alexandra, Alexandra Baumeyer, Hélène Berthoud, Lauriane Braillard, Florian Gschwend, Anne Guisolan, John Haldemann, Jörg Hummerjohann, Charlotte Joller, Florian Loosli, and et al. 2024. "Antilisterial Properties of Selected Strains from the Autochthonous Microbiota of a Swiss Artisan Soft Smear Cheese" Foods 13, no. 21: 3473. https://doi.org/10.3390/foods13213473
APA StyleRoetschi, A., Baumeyer, A., Berthoud, H., Braillard, L., Gschwend, F., Guisolan, A., Haldemann, J., Hummerjohann, J., Joller, C., Loosli, F., Meola, M., Naskova, J., Oberhänsli, S., Shani, N., von Ah, U., & Arias-Roth, E. (2024). Antilisterial Properties of Selected Strains from the Autochthonous Microbiota of a Swiss Artisan Soft Smear Cheese. Foods, 13(21), 3473. https://doi.org/10.3390/foods13213473