
Deep Impact: Shifts of Native Cultivable Microbial Communities on Fresh Lettuce after Treatment with Plasma-Treated Water

Abstract

1. Introduction
2. Materials and Methods
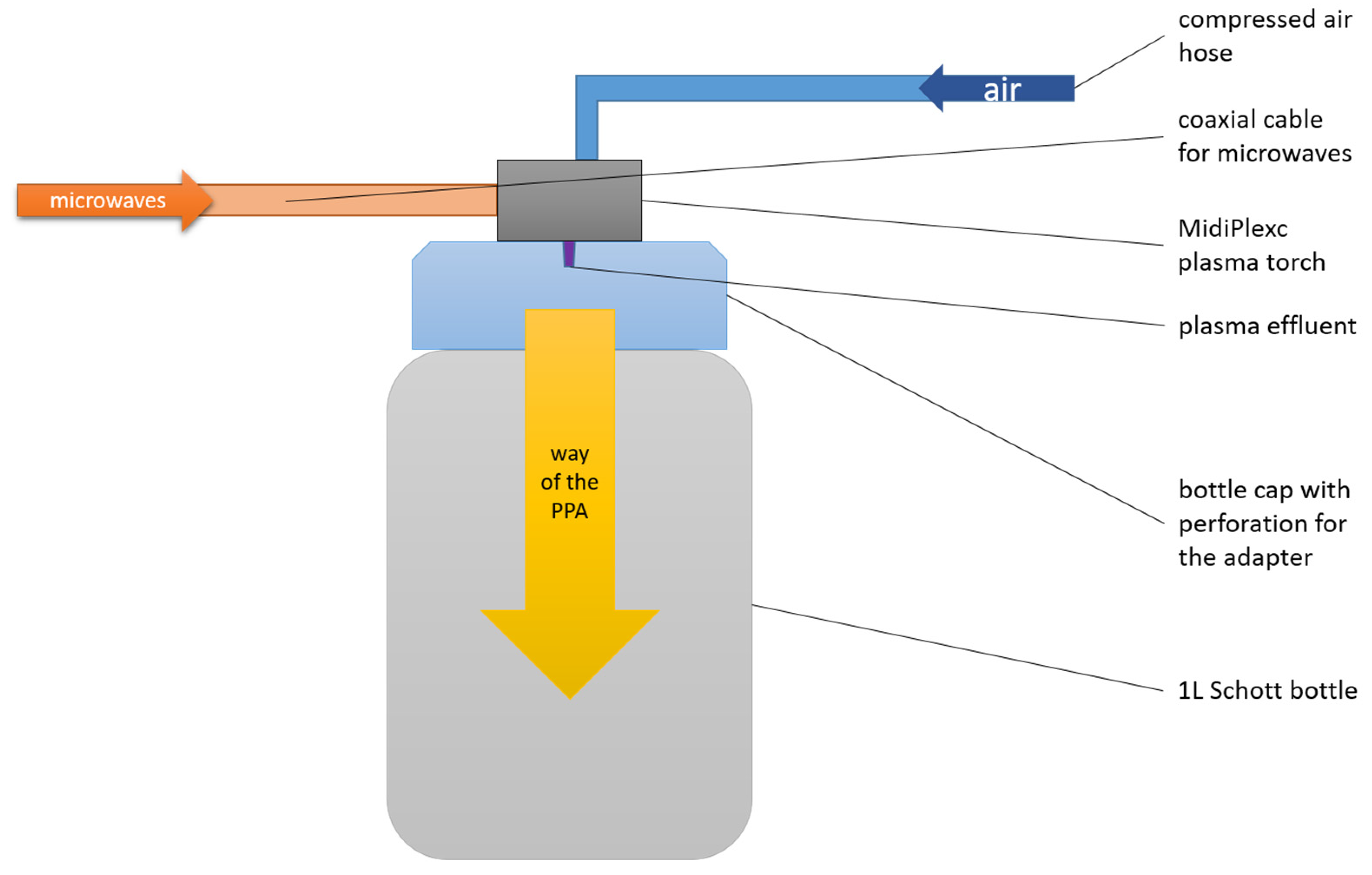
2.1. PTW Generation with Microwave Discharge
2.2. PTW Treatment of Bite-Sized Endive Pieces
2.3. Lettuce Conditions
2.4. Proliferation Assay Based on Total Viable Count (CFU/mL)
2.5. Live/Dead Assay
2.6. MALDI-ToF Analysis of Culturable Microbial Colonies
2.7. Ion Chromatography (IC) of the Lettuce Samples
2.8. Statistical Tests for Differences in the TVC
3. Results
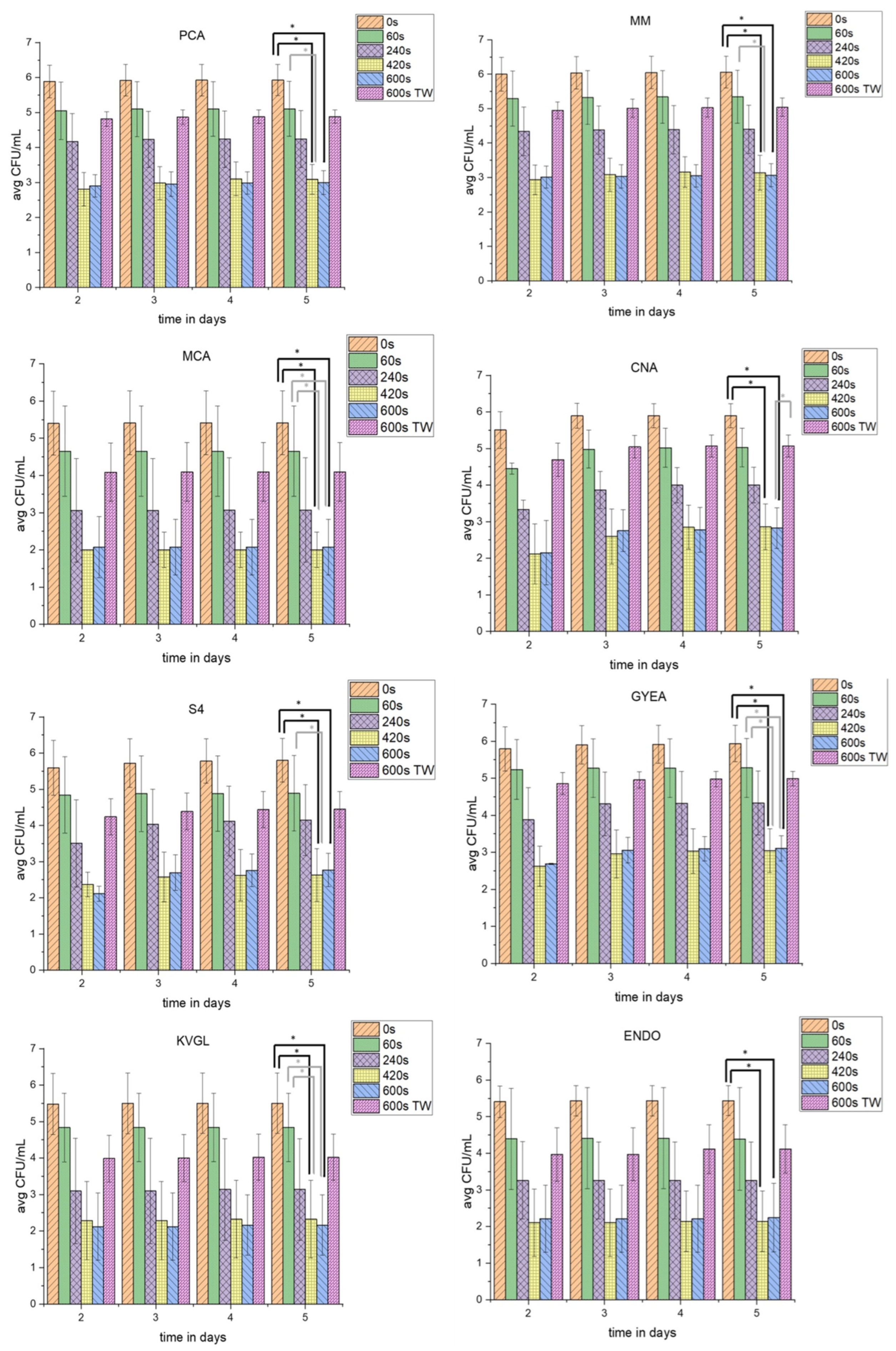
3.1. Proliferation Assay of PTW-Treated and TW-Treated Lettuce
3.1.1. Proliferation Assay of Full Medium (PCA) and Shortage Medium (MM)
3.1.2. TVC-Assay of Special Agars for Gram-Negative (MCA), Gram-Positive (CNA), and Yeast and Fungi (S4)
3.1.3. Proliferation Assay of Special Agar for Sporulating Bacteria (GYEA)
3.1.4. Proliferation Assay of the Special Agars for Coliform Bacteria (KVGL and ENDO)
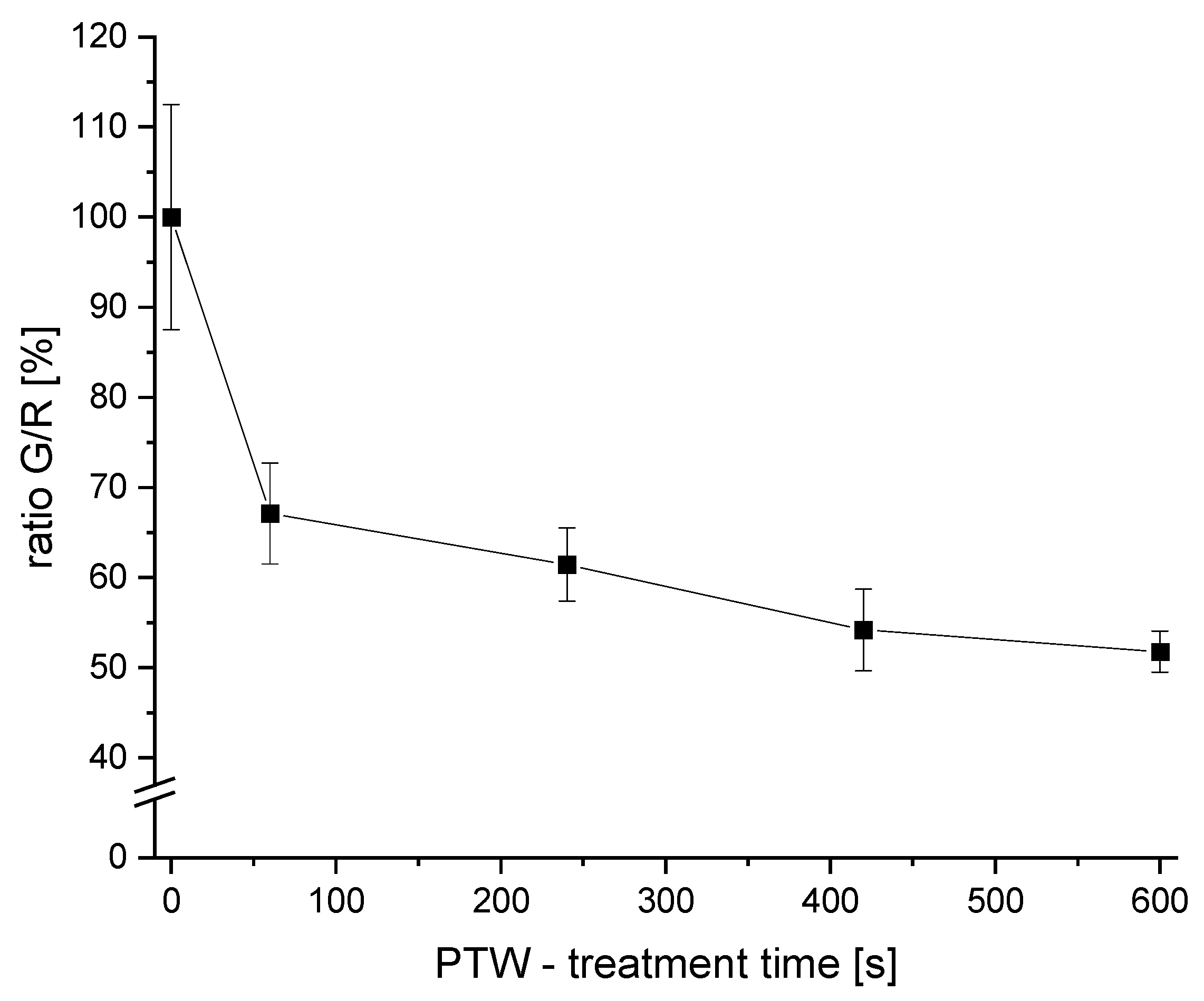
3.2. The Detected Live/Dead-Ratio after Different PTW-Treatment Times
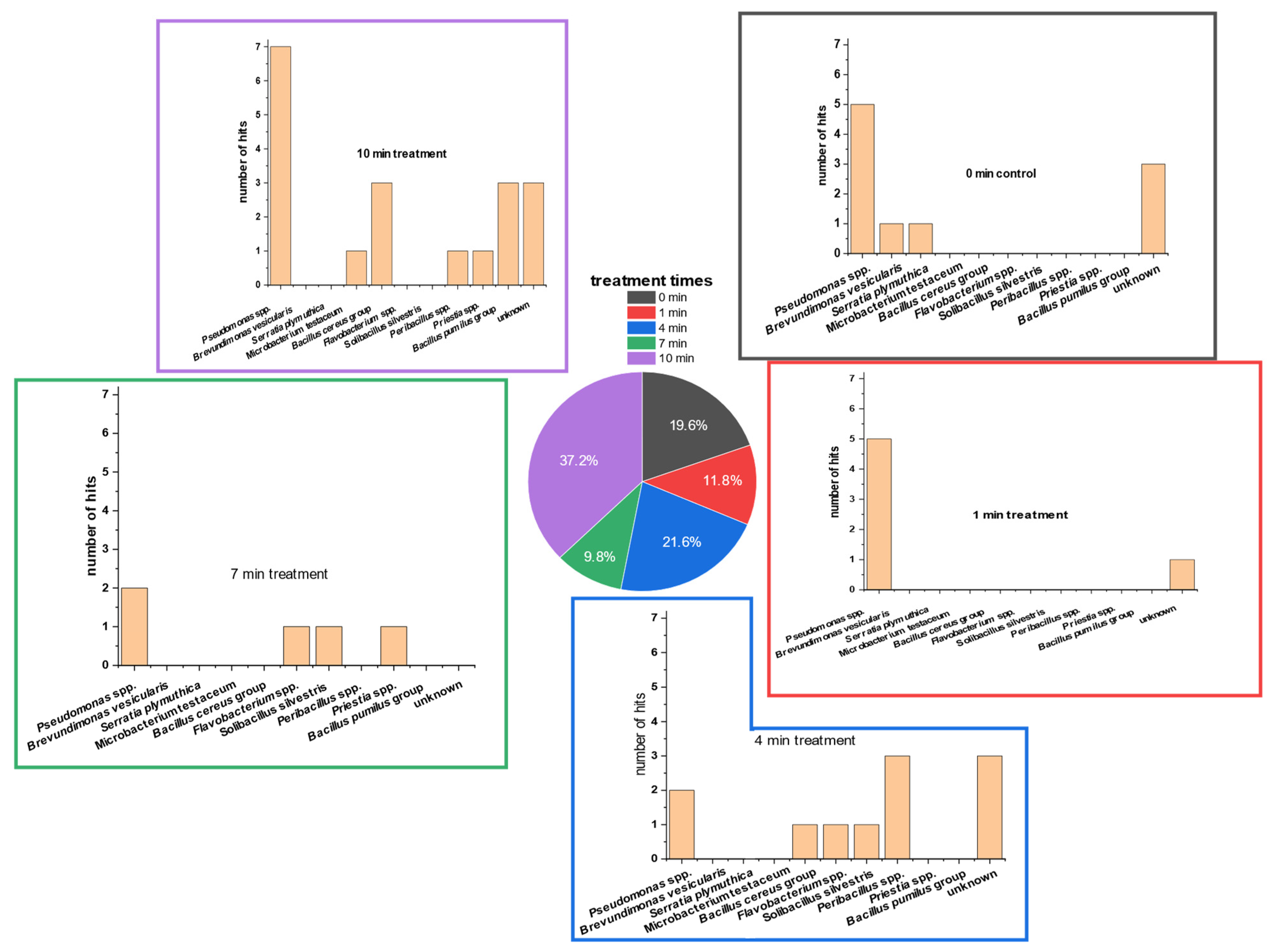
3.3. Detection of Bacterial Groups Using MALDI-ToF Analysis
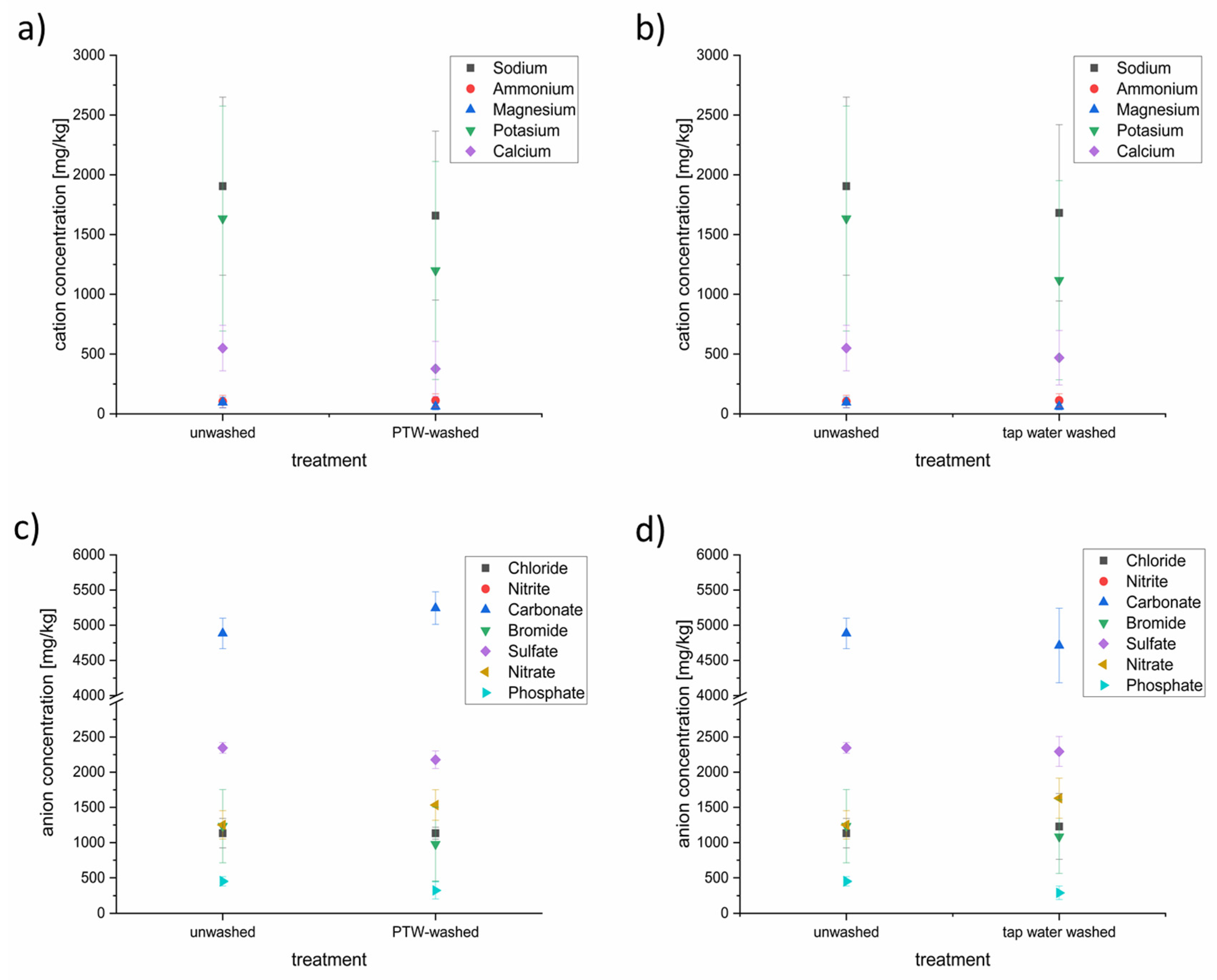
3.4. Characterization of the Anion and Cation Content of Lettuce after Washing with PTW or Tap Water with Ion Chromatography
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Edlund, A.; Yang, Y.; Yooseph, S.; He, X.; Shi, W.; McLean, J.S. Uncovering complex microbiome activities via metatranscriptomics during 24 hours of oral biofilm assembly and maturation. Microbiome 2018, 6, 217. [Google Scholar] [CrossRef]
- Turnbaugh, P.J.; Ley, R.E.; Hamady, M.; Fraser-Liggett, C.M.; Knight, R.; Gordon, J.I. The human microbiome project. Nature 2007, 449, 804–810. [Google Scholar] [CrossRef]
- Ursell, L.K.; Metcalf, J.L.; Parfrey, L.W.; Knight, R. Defining the human microbiome. Nutr. Rev. 2012, 70 (Suppl. S1), S38–S44. [Google Scholar] [CrossRef]
- Amon, P.; Sanderson, I. What is the microbiome? Arch. Dis. Child.-Educ. Pract. 2017, 102, 257–260. [Google Scholar] [CrossRef]
- Van Der Heijden, M.G.; Hartmann, M. Networking in the plant microbiome. PLoS Biol. 2016, 14, e1002378. [Google Scholar] [CrossRef]
- Liu, H.; Brettell, L.E.; Singh, B. Linking the phyllosphere microbiome to plant health. Trends Plant Sci. 2020, 25, 841–844. [Google Scholar] [CrossRef]
- Köberl, M.; Erschen, S.; Etemadi, M.; White, R.A.; El-Arabi, T.F.; Berg, G. Deciphering the microbiome shift during fermentation of medicinal plants. Sci. Rep. 2019, 9, 13461. [Google Scholar] [CrossRef]
- Lyu, D.; Zajonc, J.; Pagé, A.; Tanney, C.A.; Shah, A.; Monjezi, N.; Msimbira, L.A.; Antar, M.; Nazari, M.; Backer, R. Plant holobiont theory: The phytomicrobiome plays a central role in evolution and success. Microorganisms 2021, 9, 675. [Google Scholar] [CrossRef]
- Leveau, J.H. A brief from the leaf: Latest research to inform our understanding of the phyllosphere microbiome. Curr. Opin. Microbiol. 2019, 49, 41–49. [Google Scholar] [CrossRef]
- Ritpitakphong, U.; Falquet, L.; Vimoltust, A.; Berger, A.; Métraux, J.P.; L’Haridon, F. The microbiome of the leaf surface of Arabidopsis protects against a fungal pathogen. New Phytol. 2016, 210, 1033–1043. [Google Scholar] [CrossRef]
- Schlaeppi, K.; Bulgarelli, D. The plant microbiome at work. Mol. Plant Microbe Interact. 2015, 28, 212–217. [Google Scholar] [CrossRef]
- Thapa, S.; Prasanna, R. Prospecting the characteristics and significance of the phyllosphere microbiome. Ann. Microbiol. 2018, 68, 229–245. [Google Scholar] [CrossRef]
- Cernava, T.; Chen, X.; Krug, L.; Li, H.; Yang, M.; Berg, G. The tea leaf microbiome shows specific responses to chemical pesticides and biocontrol applications. Sci. Total Environ. 2019, 667, 33–40. [Google Scholar] [CrossRef]
- Gordon, J.; Knowlton, N.; Relman, D.A.; Rohwer, F.; Youle, M. Superorganisms and holobionts. Microbe 2013, 8, 152–153. [Google Scholar] [CrossRef]
- Erlacher, A.; Cardinale, M.; Grube, M.; Berg, G. Biotic stress shifted structure and abundance of Enterobacteriaceae in the lettuce microbiome. PLoS ONE 2015, 10, e0118068. [Google Scholar] [CrossRef]
- Cardinale, M.; Grube, M.; Erlacher, A.; Quehenberger, J.; Berg, G. Bacterial networks and co-occurrence relationships in the lettuce root microbiota. Environ. Microbiol. 2015, 17, 239–252. [Google Scholar] [CrossRef]
- Schnabel, U.; Handorf, O.; Winter, H.; Weihe, T.; Weit, C.; Schäfer, J.; Stachowiak, J.; Boehm, D.; Below, H.; Bourke, P. The effect of plasma treated water unit processes on the food quality characteristics of fresh-cut endive. Front. Nutr. 2021, 7, 627483. [Google Scholar] [CrossRef]
- Williams, T.R.; Moyne, A.L.; Harris, L.J.; Marco, M.L. Season, irrigation, leaf age, and Escherichia coli inoculation influence the bacterial diversity in the lettuce phyllosphere. PLoS ONE 2013, 8, e68642. [Google Scholar] [CrossRef]
- Schnabel, U.; Niquet, R.; Krohmann, U.; Winter, J.; Schlüter, O.; Weltmann, K.D.; Ehlbeck, J. Decontamination of microbiologically contaminated specimen by direct and indirect plasma treatment. Plasma Process. Polym. 2012, 9, 569–575. [Google Scholar] [CrossRef]
- McManamon, O.; Scollard, J.; Schmalenberger, A. Inoculation density is affecting growth conditions of Listeria monocytogenes on fresh cut lettuce. World J. Microbiol. Biotechnol. 2017, 33, 217. [Google Scholar] [CrossRef]
- Abadias, M.; Usall, J.; Oliveira, M.; Alegre, I.; Viñas, I. Efficacy of neutral electrolyzed water (NEW) for reducing microbial contamination on minimally-processed vegetables. Int. J. Food Microbiol. 2008, 123, 151–158. [Google Scholar] [CrossRef]
- Lopez-Velasco, G.; Welbaum, G.E.; Boyer, R.R.; Mane, S.P.; Ponder, M.A. Changes in spinach phylloepiphytic bacteria communities following minimal processing and refrigerated storage described using pyrosequencing of 16S rRNA amplicons. J. Appl. Microbiol. 2011, 110, 1203–1214. [Google Scholar] [CrossRef]
- Gram, L.; Ravn, L.; Rasch, M.; Bruhn, J.B.; Christensen, A.B.; Givskov, M. Food spoilage—Interactions between food spoilage bacteria. Int. J. Food Microbiol. 2002, 78, 79–97. [Google Scholar] [CrossRef]
- Rose, D.; Bianchini, A.; Martinez, B.; Flores, R. Methods for reducing microbial contamination of wheat flour and effects on functionality. Cereal Foods World 2012, 57, 104. [Google Scholar] [CrossRef]
- Wang, H.; Feng, H.; Luo, Y. Microbial reduction and storage quality of fresh-cut cilantro washed with acidic electrolyzed water and aqueous ozone. Food Res. Int. 2004, 37, 949–956. [Google Scholar] [CrossRef]
- Pirovani, M.; Piagentini, A.; Güemes, D.; Arkwright, S. Reduction of chlorine concentration and microbial load during washing-disinfection of shredded lettuce. Int. J. Food Sci. Technol. 2004, 39, 341–347. [Google Scholar] [CrossRef]
- Klaiber, R.G.; Baur, S.; Wolf, G.; Hammes, W.P.; Carle, R. Quality of minimally processed carrots as affected by warm water washing and chlorination. Innov. Food Sci. Emerg. Technol. 2005, 6, 351–362. [Google Scholar] [CrossRef]
- Delaquis, P.J.; Fukumoto, L.R.; Toivonen, P.M.; Cliff, M.A. Implications of wash water chlorination and temperature for the microbiological and sensory properties of fresh-cut iceberg lettuce. Postharvest Biol. Technol. 2004, 31, 81–91. [Google Scholar] [CrossRef]
- Najafi, M.B.H.; Khodaparast, M.H. Efficacy of ozone to reduce microbial populations in date fruits. Food Control 2009, 20, 27–30. [Google Scholar] [CrossRef]
- Ehlbeck, J.; Schnabel, U.; Polak, M.; Winter, J.; Von Woedtke, T.; Brandenburg, R.; Von dem Hagen, T.; Weltmann, K. Low temperature atmospheric pressure plasma sources for microbial decontamination. J. Phys. D Appl. Phys. 2010, 44, 013002. [Google Scholar] [CrossRef]
- Handorf, O.; Pauker, V.I.; Schnabel, U.; Weihe, T.; Freund, E.; Bekeschus, S.; Riedel, K.; Ehlbeck, J. Characterization of Antimicrobial Effects of Plasma-Treated Water (PTW) Produced by Microwave-Induced Plasma (MidiPLexc) on Pseudomonas fluorescens Biofilms. Appl. Sci. 2020, 10, 3118. [Google Scholar] [CrossRef]
- Kogelschatz, U. Atmospheric-pressure plasma technology. Plasma Phys. Control. Fusion 2004, 46, B63–B75. [Google Scholar] [CrossRef]
- Handorf, O.; Below, H.; Schnabel, U.; Riedel, K.; Ehlbeck, J. Investigation of the chemical composition of plasma-treated water by MidiPLexc and its antimicrobial effect onL. monocytogenesandPseudomonas fluorescensmonospecies suspension cultures. J. Phys. D Appl. Phys. 2020, 53, 305204. [Google Scholar] [CrossRef]
- Schmidt-Bleker, A.; Winter, J.; Bösel, A.; Reuter, S.; Weltmann, K.-D. On the plasma chemistry of a cold atmospheric argon plasma jet with shielding gas device. Plasma Sources Sci. Technol. 2015, 25, 015005. [Google Scholar] [CrossRef]
- Ammann, P.; Timmins, R. Chemical reactions during rapid quenching of oxygen-nitrogen mixtures from very high temperatures. AIChE J. 1966, 12, 956–963. [Google Scholar] [CrossRef]
- Shiva, S. Nitrite: A physiological store of nitric oxide and modulator of mitochondrial function. Redox Biol. 2013, 1, 40–44. [Google Scholar] [CrossRef]
- Stock, H. Prüfbericht 23-1116-001. In Untersuchung Nach Trinkwasserverordnung; Stadtwerke Greifswald: Greifswald, Germany, 2023; pp. 1–4. [Google Scholar]
- Weihe, T.; Schnabel, U.; Winter, H.; Möller, T.; Stachowiak, J.; Neumann, S.; Schlüter, O.; Ehlbeck, J. Reduce and refine: Plasma treated water vs conventional disinfectants for conveyor-belt cleaning in sustainable food-production lines. J. Appl. Phys. 2021, 129, 223304. [Google Scholar] [CrossRef]
- Jovićević, S.; Ivković, M.; Pavlović, Z.; Konjević, N. Parametric study of an atmospheric pressure microwave-induced plasma of the mini MIP torch—I. Two-dimensional spatially resolved electron-number density measurements. Spectrochim. Acta Part B At. Spectrosc. 2000, 55, 1879–1893. [Google Scholar] [CrossRef]
- Miles, A.A.; Misra, S.; Irwin, J. The estimation of the bactericidal power of the blood. Epidemiol. Infect. 1938, 38, 732–749. [Google Scholar] [CrossRef]
- Beveridge, T.J. Structures of gram-negative cell walls and their derived membrane vesicles. J. Bacteriol. 1999, 181, 4725–4733. [Google Scholar] [CrossRef]
- Chislett, M.; Guo, J.; Bond, P.L.; Wang, Y.; Donose, B.C.; Yuan, Z. Reactive nitrogen species from free nitrous acid (FNA) cause cell lysis. Water Res. 2022, 217, 118401. [Google Scholar] [CrossRef]
- van der Heijden, J.; Vogt, S.L.; Reynolds, L.A.; Peña-Díaz, J.; Tupin, A.; Aussel, L.; Finlay, B.B. Exploring the redox balance inside gram-negative bacteria with redox-sensitive GFP. Free Radic. Biol. Med. 2016, 91, 34–44. [Google Scholar] [CrossRef]
- Shockman, G.D.; Barren, J.F. Structure, function, and assembly of cell walls of gram-positive bacteria. Annu. Rev. Microbiol. 1983, 37, 501–527. [Google Scholar] [CrossRef]
- Schäffer, C.; Messner, P. The structure of secondary cell wall polymers: How Gram-positive bacteria stick their cell walls together. Microbiology 2005, 151, 643–651. [Google Scholar] [CrossRef]
- Silhavy, T.J.; Kahne, D.; Walker, S. The bacterial cell envelope. Cold Spring Harb. Perspect. Biol. 2010, 2, a000414. [Google Scholar] [CrossRef]
- Nicholson, W.L.; Fajardo-Cavazos, P.; Rebeil, R.; Slieman, T.A.; Riesenman, P.J.; Law, J.F.; Xue, Y. Bacterial endospores and their significance in stress resistance. Antonie Leeuwenhoek 2002, 81, 27–32. [Google Scholar] [CrossRef]
- Schnabel, U.; Andrasch, M.; Weltmann, K.D.; Ehlbeck, J. Inactivation of vegetative microorganisms and Bacillus atrophaeus endospores by reactive nitrogen species (RNS). Plasma Process. Polym. 2014, 11, 110–116. [Google Scholar] [CrossRef]
- Cotter, P.D.; Hill, C. Surviving the acid test: Responses of gram-positive bacteria to low pH. Microbiol. Mol. Biol. Rev. 2003, 67, 429–453. [Google Scholar] [CrossRef]
- Aeschlimann, J.R. The role of multidrug efflux pumps in the antibiotic resistance of Pseudomonas aeruginosa and other gram-negative bacteria: Insights from the Society of Infectious Diseases Pharmacists. Pharmacother. J. Hum. Pharmacol. Drug Ther. 2003, 23, 916–924. [Google Scholar] [CrossRef]
- Stratford, M. Another brick in the wall? Recent developments concerning the yeast cell envelope. Yeast 1994, 10, 1741–1752. [Google Scholar] [CrossRef]
- Winter, H.; Wagner, R.; Yao, Y.; Ehlbeck, J.; Schnabel, U. Influence of plasma-treated air on surface microbial communities on freshly harvested lettuce. Curr. Res. Food Sci. 2023, 7, 100649. [Google Scholar] [CrossRef]
- Özdemir, E.; Başaran, P.; Kartal, S.; Akan, T. Cold plasma application to fresh green leafy vegetables: Impact on microbiology and product quality. Compr. Rev. Food Sci. Food Saf. 2023, 22, 4484–4515. [Google Scholar] [CrossRef]
- Mann, M.S.; Schnabel, U.; Weihe, T.; Weltmann, K.-D.; von Woedtke, T. A reference technique to compare the antimicrobial properties of atmospheric pressure plasma sources. Plasma Med. 2015, 5, 27–47. [Google Scholar] [CrossRef]
- Oliver, J.D. The viable but nonculturable state in bacteria. J. Microbiol. 2005, 43, 93–100. [Google Scholar]
- Liu, J.; Yang, L.; Kjellerup, B.V.; Xu, Z. Viable but nonculturable (VBNC) state, an underestimated and controversial microbial survival strategy. Trends Microbiol. 2023, 31, 1013–1023. [Google Scholar] [CrossRef]
- Schottroff, F.; Fröhling, A.; Zunabovic-Pichler, M.; Krottenthaler, A.; Schlüter, O.; Jäger, H. Sublethal injury and viable but non-culturable (VBNC) state in microorganisms during preservation of food and biological materials by non-thermal processes. Front. Microbiol. 2018, 9, 2773. [Google Scholar] [CrossRef]
- Salma, M.; Rousseaux, S.; Sequeira-Le Grand, A.; Divol, B.; Alexandre, H. Characterization of the viable but nonculturable (VBNC) state in Saccharomyces cerevisiae. PLoS ONE 2013, 8, e77600. [Google Scholar] [CrossRef]
- Trevors, J. Viable but non-culturable (VBNC) bacteria: Gene expression in planktonic and biofilm cells. J. Microbiol. Methods 2011, 86, 266–273. [Google Scholar] [CrossRef]
- Schnabel, U.; Handorf, O.; Yarova, K.; Zessin, B.; Zechlin, S.; Sydow, D.; Zellmer, E.; Stachowiak, J.; Andrasch, M.; Below, H. Plasma-treated air and water—Assessment of synergistic antimicrobial effects for sanitation of food processing surfaces and environment. Foods 2019, 8, 55. [Google Scholar] [CrossRef]
- Takeuchi, K.; Matute, C.M.; Hassan, A.N.; Frank, J.F. Comparison of the attachment of Escherichia coli O157: H7, Listeria monocytogenes, Salmonella Typhimurium, and Pseudomonas fluorescens to lettuce leaves. J. Food Prot. 2000, 63, 1433–1437. [Google Scholar] [CrossRef]
- Cipriano, M.A.; Lupatini, M.; Lopes-Santos, L.; da Silva, M.J.; Roesch, L.F.; Destefano, S.A.; Freitas, S.S.; Kuramae, E.E. Lettuce and rhizosphere microbiome responses to growth promoting Pseudomonas species under field conditions. FEMS Microbiol. Ecol. 2016, 92, fiw197. [Google Scholar] [CrossRef]
- Grogan, R.; Misaghi, I.; Kimble, K.; Greathead, A.; Ririe, D.; Bardin, R. Varnish spot, destructive disease of lettuce in California caused by Pseudomonas cichorii. Phytopathology 1977, 67, 957–960. [Google Scholar] [CrossRef]
- Starkey, M.; Rahme, L.G. Modeling Pseudomonas aeruginosa pathogenesis in plant hosts. Nat. Protoc. 2009, 4, 117–124. [Google Scholar] [CrossRef]
- Hansen, B.M.; Hendriksen, N.B. Detection of enterotoxic Bacillus cereus and Bacillus thuringiensis strains by PCR analysis. Appl. Environ. Microbiol. 2001, 67, 185–189. [Google Scholar] [CrossRef]
- Leong, S.S.; Korel, F.; King, J.H. Bacillus cereus: A review of “fried rice syndrome” causative agents. Microb. Pathog. 2023, 185, 106418. [Google Scholar] [CrossRef]
- Cerna-Cortes, J.F.; Leon-Montes, N.; Cortes-Cueto, A.L.; Salas-Rangel, L.P.; Helguera-Repetto, A.C.; Lopez-Hernandez, D.; Rivera-Gutierrez, S.; Fernandez-Rendon, E.; Gonzalez-y-Merchand, J.A. Microbiological quality of ready-to-eat vegetables collected in Mexico City: Occurrence of aerobic-mesophilic bacteria, fecal coliforms, and potentially pathogenic nontuberculous mycobacteria. BioMed Res. Int. 2015, 2015, 789508. [Google Scholar] [CrossRef]
- Falomir, M.P.; Gozalbo, D.; Rico, H. Coliform bacteria in fresh vegetables: From cultivated lands to consumers. Curr. Res. Technol. Educ. Top. Appl. Microbiol. Microb. Biotechnol. 2010, 2, 1175–1181. [Google Scholar]
- Martin, N.H.; Trmčić, A.; Hsieh, T.-H.; Boor, K.J.; Wiedmann, M. The evolving role of coliforms as indicators of unhygienic processing conditions in dairy foods. Front. Microbiol. 2016, 7, 1549. [Google Scholar] [CrossRef]
- Bahadoran, Z.; Mirmiran, P.; Ghasemi, A.; Kabir, A.; Azizi, F.; Hadaegh, F. Is dietary nitrate/nitrite exposure a risk factor for development of thyroid abnormality? A systematic review and meta-analysis. Nitric Oxide 2015, 47, 65–76. [Google Scholar] [CrossRef]
- Quijano, L.; Yusà, V.; Font, G.; McAllister, C.; Torres, C.; Pardo, O. Risk assessment and monitoring programme of nitrates through vegetables in the Region of Valencia (Spain). Food Chem. Toxicol. 2017, 100, 42–49. [Google Scholar] [CrossRef]
- Hosseini, M.-J.; Dezhangah, S.; Esmi, F.; Gharavi-Nakhjavani, M.S.; Hashempour-Baltork, F.; Alizadeh, A.M. A worldwide systematic review, meta-analysis and meta-regression of nitrate and nitrite in vegetables and fruits. Ecotoxicol. Environ. Saf. 2023, 257, 114934. [Google Scholar] [CrossRef] [PubMed]
- Handorf, O.; Pauker, V.I.; Weihe, T.; Schafer, J.; Freund, E.; Schnabel, U.; Bekeschus, S.; Riedel, K.; Ehlbeck, J. Plasma-Treated Water Affects Listeria monocytogenes Vitality and Biofilm Structure. Front. Microbiol. 2021, 12, 652481. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Su, P.; Chen, H.; Qiao, M.; Yang, B.; Zhao, X. Impact of reactive oxygen species on cell activity and structural integrity of Gram-positive and Gram-negative bacteria in electrochemical disinfection system. Chem. Eng. J. 2023, 451, 138879. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Agar Type | Cultivable Microbial Groups | Agar Composition | Company |
|---|---|---|---|
| Plate count agar (PCA) | Full-medium agar | 23.5 g/L | Carl Roth (Karlsruhe, Germany) |
| Standard count agar (MM) | Minimal-medium agar | 25 g/L | Carl Roth (Germany) |
| MacConkey agar (MCA) | Special agar for Gram-negative bacteria | 50 g/L | Carl Roth (Germany) |
| PCA with collistinesulfate and nalidixinacid (CNA) | Special agar for Gram-positive bacteria | PCA After autoclaving: 10 mg/L collistinsulfat 15 mg/L nalidixinacid | PCA; Carl Roth (Germany) Collistinsulfat; Carl Roth (Germany) nalidixinacid; Carl Roth (Germany) |
| Sabouraud agar with 4% dextrose (S4) | Special agar for yeasts and molds | 65 g/L | Merck KGaA (Darmstadt, Germany) |
| Glycerol yeast extract agar (GYEA) | Special agar composition for sporulating bacteria | 15 g/L agar-agar 5 g/L glycerol 2 g/L yeast extract 1 g/L di-potassium phosphate | Agar-argar, Kobe I; Carl Roth (Germany) Glycerol; TH Geyer (Renningen, Germany) Yeast extract; Carl Roth (Germany) Di-potassium-phosphate; Carl Roth (Germany) |
| Crystal violet bile lactose agar (CVBLA) | Special agar for coliforms (room temperature) | 41.5 g/L | Carl Roth (Germany) |
| Endo agar (ENDO) | Special agar for coliforms (37 °C) | 36 g/L Endo agar base agar 5 mL/L ethanolic fuchsin solution | Endo agar base, Carl Roth (Germany) Fuchsin, Carl Roth (Germany) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Winter, H.; Wagner, R.; Ehlbeck, J.; Urich, T.; Schnabel, U. Deep Impact: Shifts of Native Cultivable Microbial Communities on Fresh Lettuce after Treatment with Plasma-Treated Water. Foods 2024, 13, 282. https://doi.org/10.3390/foods13020282
Winter H, Wagner R, Ehlbeck J, Urich T, Schnabel U. Deep Impact: Shifts of Native Cultivable Microbial Communities on Fresh Lettuce after Treatment with Plasma-Treated Water. Foods. 2024; 13(2):282. https://doi.org/10.3390/foods13020282
Chicago/Turabian StyleWinter, Hauke, Robert Wagner, Jörg Ehlbeck, Tim Urich, and Uta Schnabel. 2024. "Deep Impact: Shifts of Native Cultivable Microbial Communities on Fresh Lettuce after Treatment with Plasma-Treated Water" Foods 13, no. 2: 282. https://doi.org/10.3390/foods13020282
APA StyleWinter, H., Wagner, R., Ehlbeck, J., Urich, T., & Schnabel, U. (2024). Deep Impact: Shifts of Native Cultivable Microbial Communities on Fresh Lettuce after Treatment with Plasma-Treated Water. Foods, 13(2), 282. https://doi.org/10.3390/foods13020282