
Short-Term Soy Bread Intervention Leads to a Dose-Response Increase in Urinary Isoflavone Metabolites and Satiety in Chronic Pancreatitis
 ,
,
Abstract
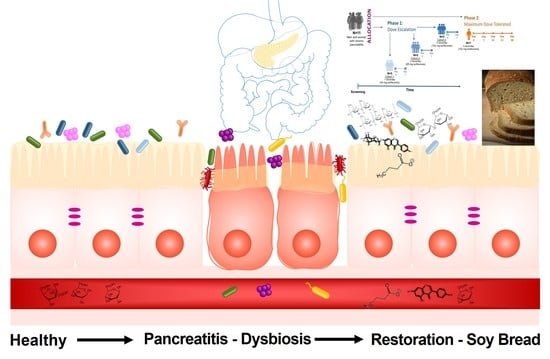
1. Introduction
2. Materials and Methods
2.1. Soy Bread Materials
2.2. Soy Bread Preparation and Isoflavone Analysis
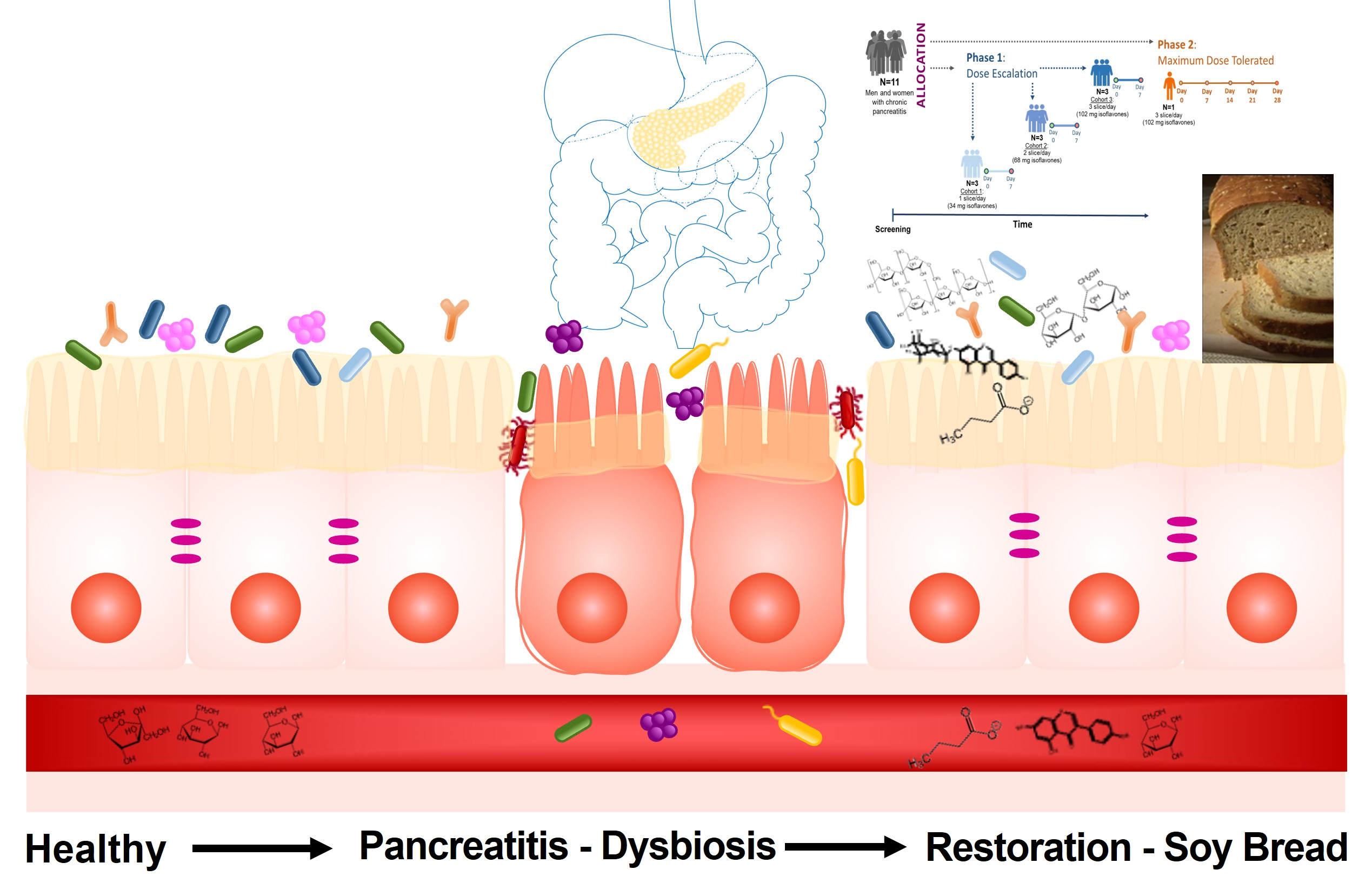
2.3. Study Design
2.4. Participant Description
2.5. Sensory and Satiety Assessments
2.6. Laboratory and Clinical Measures of Toxicity
2.7. Isoflavone Quantification in Urine
2.7.1. Chemicals
2.7.2. Sample Handling and Isoflavone Quantification
2.8. Statistical Analysis
3. Results and Discussion
3.1. Soy Bread Nutrient and Isoflavone Composition
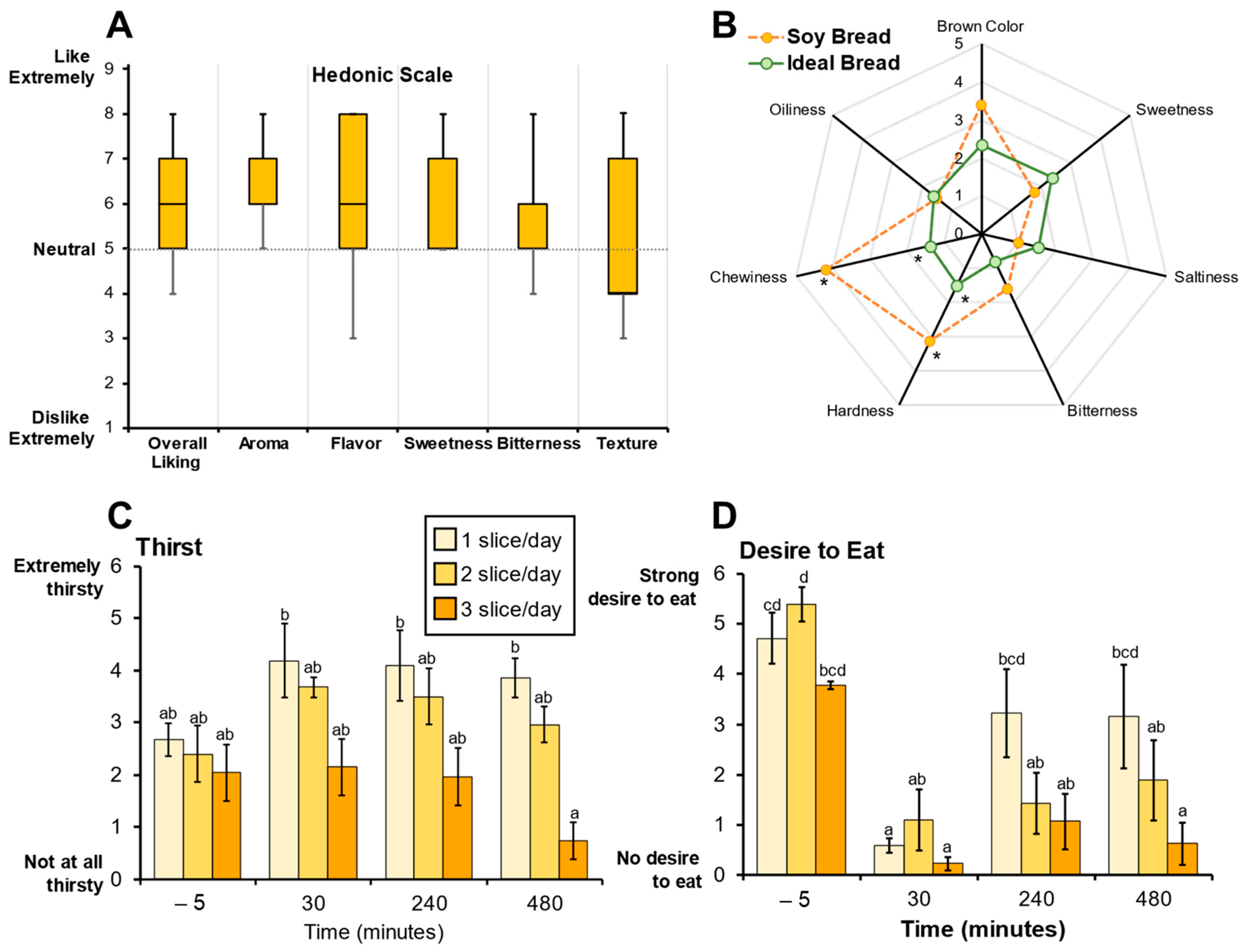
3.2. Soy Bread Palatability and Its Impact on Satiety
3.3. Participant Clinical Features and Dietary Adhrence
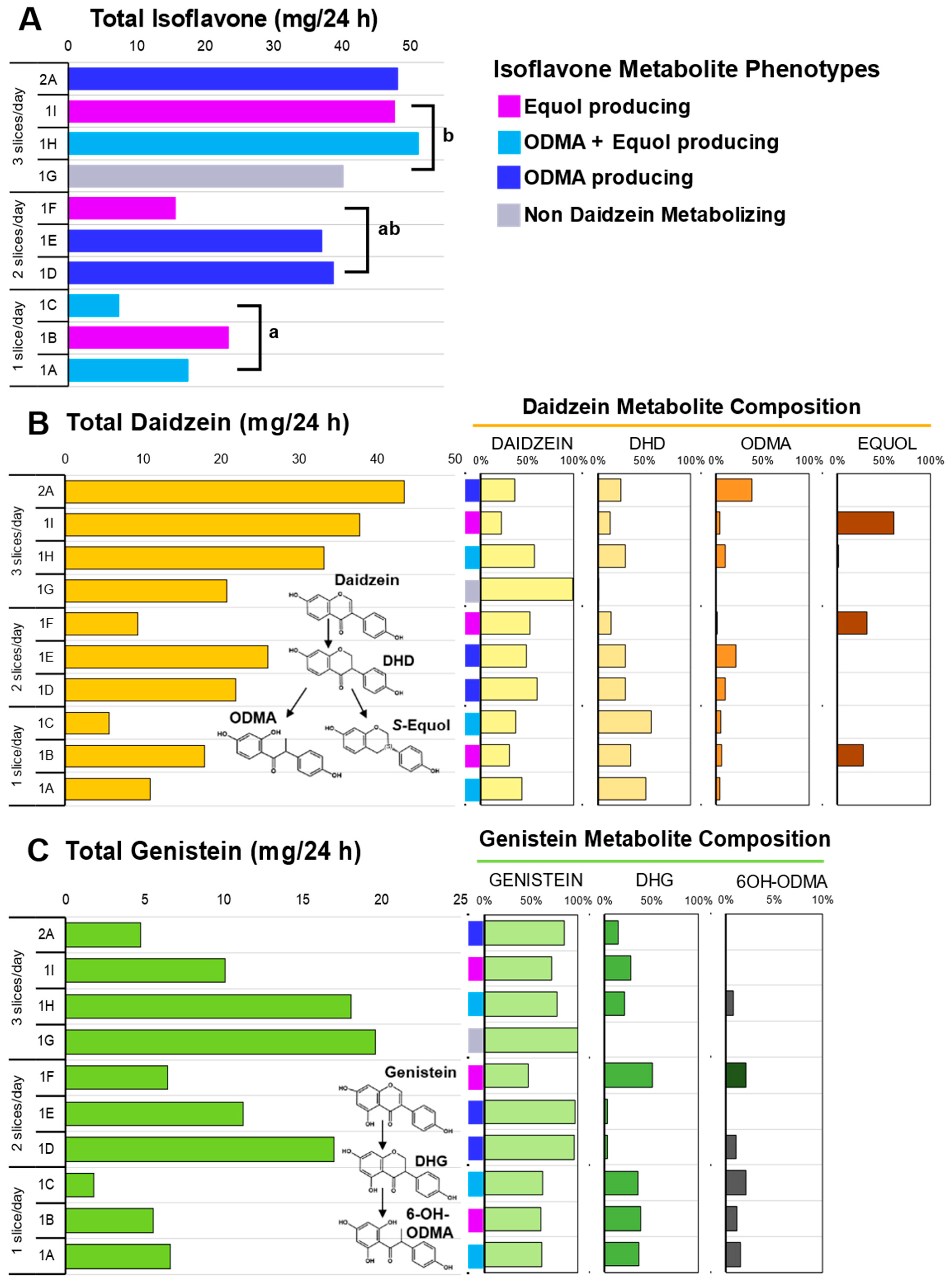
3.4. Isoflavone Metabolism
3.5. Isoflavonoid Phenotypes
3.6. Potential Implications for Clinical Care
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
References
- Kleeff, J.; Whitcomb, D.C.; Shimosegawa, T.; Esposito, I.; Lerch, M.M.; Gress, T.; Mayerle, J.; Drewes, A.M.; Rebours, V.; Akisik, F.; et al. Chronic pancreatitis. Nat. Rev. Dis. Primers 2017, 3, 17060. [Google Scholar] [CrossRef] [PubMed]
- Hart, P.A.; Conwell, D.L. Chronic Pancreatitis: Managing a Difficult Disease. Am. J. Gastroenterol. 2020, 115, 49–55. [Google Scholar] [CrossRef] [PubMed]
- Rasmussen, H.H.; Irtun, O.; Olesen, S.S.; Drewes, A.M.; Holst, M. Nutrition in chronic pancreatitis. World J. Gastroenterol. 2013, 19, 7267–7275. [Google Scholar] [CrossRef] [PubMed]
- Duggan, S.N. Negotiating the complexities of exocrine and endocrine dysfunction in chronic pancreatitis. Proc. Nutr. Soc. 2017, 76, 484–494. [Google Scholar] [CrossRef] [PubMed]
- Ramsey, M.L.; Conwell, D.L.; Hart, P.A. Complications of chronic pancreatitis. Dig. Dis. Sci. 2017, 62, 1745–1750. [Google Scholar] [CrossRef]
- Capurso, G.; Signoretti, M.; Archibugi, L.; Stigliano, S.; Fave, G.D. Systematic review and meta-analysis: Small intestinal bacterial overgrowth in chronic. Pancreatitis. United Eur. Gastroenterol. J. 2016, 4, 697–705. [Google Scholar] [CrossRef]
- Jandhyala, S.M.; Madhulika, A.; Deepika, G.; Rao, G.V.; Reddy, D.N.; Subramanyam, C.; Sasikala, M.; Talukdar, R. Altered intestinal microbiota in patients with chronic pancreatitis: Implications in diabetes and metabolic abnormalities. Sci. Rep. 2017, 7, srep43640. [Google Scholar] [CrossRef]
- Schepis, T.; De Lucia, S.S.; Nista, E.C.; Manilla, V.; Pignataro, G.; Ojetti, V.; Piccioni, A.; Gasbarrini, A.; Franceschi, F.; Candelli, M. Microbiota in pancreatic diseases: A review of the literature. J. Clin. Med. 2021, 10, 5920. [Google Scholar] [CrossRef]
- Komar, H.M.; Hart, P.A.; Cruz-Monserrate, Z.; Conwell, D.L.; Lesinski, G.B. Local and systemic expression of immunomodulatory factors in chronic pancreatitis. Pancreas 2017, 46, 986. [Google Scholar] [CrossRef]
- Frost, F.; Weiss, F.U.; Sendler, M.; Kacprowski, T.; Rühlemann, M.; Bang, C.; Franke, A.; Völker, U.; Völzke, H.; Lamprecht, G.; et al. The gut microbiome in patients with chronic pancreatitis is characterized by significant dysbiosis and overgrowth by opportunistic pathogens. Clin. Transl. Gastroenterol. 2020, 11, e00232. [Google Scholar] [CrossRef]
- Petrov, M.S. Metabolic trifecta after pancreatitis: Exocrine pancreatic dysfunction, altered gut microbiota, and new-onset diabetes. Clin. Transl. Gastroenterol. 2019, 10, e00086. [Google Scholar] [CrossRef]
- De Kort, S.; Keszthelyi, D.; Masclee, A.A.M. Leaky gut and diabetes mellitus: What is the link? Obes. Rev. 2011, 12, 449–458. [Google Scholar] [CrossRef]
- Canani, R.B.; Di Costanzo, M.; Leone, L.; Pedata, M.; Meli, R.; Calignano, A. Potential beneficial effects of butyrate in intestinal and extraintestinal diseases. World J. Gastroenterol. 2011, 17, 1519–1528. [Google Scholar] [CrossRef]
- Arvanitakis, M.; Ockenga, J.; Bezmarevic, M.; Gianotti, L.; Krznarić, Ž.; Lobo, D.N.; Löser, C.; Madl, C.; Meier, R.; Phillips, M.; et al. ESPEN guideline on clinical nutrition in acute and chronic pancreatitis. Clin. Nutr. Open Sci. 2020, 39, 612–631. [Google Scholar] [CrossRef]
- Eldridge, A.C.; Black, L.T.; Wolf, W.J. Carbohydrate composition of soybean flours, protein concentrates, and isolates. J. Agric. Food Chem. 1979, 27, 799–802. [Google Scholar] [CrossRef]
- Redondo-Cuenca, A.; Villanueva-Suárez, M.J.; Rodríguez-Sevilla, M.D.; Mateos-Aparicio, I. Chemical composition and dietary fibre of yellow and green commercial soybeans (Glycine max). Food Chem. 2007, 101, 1216–1222. [Google Scholar] [CrossRef]
- Kocot, A.M.; Jarocka-Cyrta, E.; Drabińska, N. Overview of the importance of biotics in gut barrier integrity. Int. J. Mol. Sci. 2022, 23, 2896. [Google Scholar] [CrossRef]
- Sheflin, A.M.; Melby, C.L.; Carbonero, F.; Weir, T.L. Linking dietary patterns with gut microbial composition and function. Gut Microbes 2017, 8, 113–129. [Google Scholar] [CrossRef]
- Wu, Q.; Chen, T.; El-Nezami, H.; Savidge, T.C. Food ingredients in human health: Ecological and metabolic perspectives implicating gut microbiota function. Trends Food Sci. Technol. 2020, 100, 103–117. [Google Scholar] [CrossRef]
- Clavel, T.; Fallani, M.; Lepage, P.; Levenez, F.; Mathey, J.; Rochet, V.; Serezat, M.; Sutren, M.; Henderson, G.; Bennetau-Pelissero, C.; et al. Isoflavones and functional foods alter the dominant intestinal microbiota in postmenopausal women. J. Nutr. 2005, 135, 2786–2792. [Google Scholar] [CrossRef]
- Ahn-Jarvis, J.; Clinton, S.K.; Riedl, K.M.; Vodovotz, Y.; Schwartz, S.J. Impact of food matrix on isoflavone metabolism and cardiovascular biomarkers in adults with hypercholesterolemia. Food Funct. 2012, 3, 1051–1058. [Google Scholar] [CrossRef] [PubMed]
- Marciani, L.; Pritchard, S.E.; Hellier-Woods, C.; Costigan, C.; Hoad, C.L.; Gowland, P.A.; Spiller, R.C. Delayed gastric emptying and reduced postprandial small bowel water content of equicaloric whole meal bread versus rice meals in healthy subjects: Novel MRI insights. Eur. J. Clin. Nutr. 2013, 67, 754–758. [Google Scholar] [CrossRef] [PubMed]
- Setchell, K.D.; Brown, N.M.; Lydeking-Olsen, E. The clinical importance of the metabolite equol—A clue to the effectiveness of soy and its isoflavones. J. Nutr. 2002, 132, 3577–3584. [Google Scholar] [CrossRef] [PubMed]
- Sánchez-Calvo, J.M.; Rodríguez-Iglesias, M.A.; Molinillo, J.M.; Macías, F.A. Soy isoflavones and their relationship with microflora: Beneficial effects on human health in equol producers. Phytochem. Rev. 2013, 12, 979–1000. [Google Scholar] [CrossRef]
- Mayo, B.; Vázquez, L.; Flórez, A.B. Equol: A bacterial metabolite from the daidzein isoflavone and its presumed beneficial health effects. Nutrients 2019, 11, 2231. [Google Scholar] [CrossRef]
- Rowland, I.R.; Wiseman, H.; Sanders, T.A.; Adlercreutz, H.; Bowey, E.A. Interindividual variation in metabolism of soy isoflavones and lignans: Influence of habitual diet on equol production by the gut microflora. Nutr. Cancer 2000, 36, 27–32. [Google Scholar] [CrossRef]
- Akaza, H.; Miyanaga, N.; Takashima, N.; Naito, S.; Hirao, Y.; Tsukamoto, T.; Fujioka, T.; Mori, M.; Kim, W.J.; Song, J.M.; et al. Comparisons of percent equol producers between prostate cancer patients and controls: Case-controlled studies of isoflavones in Japanese, Korean and American residents. Jpn. J. Clin. Oncol. 2004, 34, 86–89. [Google Scholar] [CrossRef]
- Iino, C.; Shimoyama, T.; Iino, K.; Yokoyama, Y.; Chinda, D.; Sakuraba, H.; Fukuda, S.; Nakaji, S. Daidzein intake is associated with equol producing status through an increase in the intestinal bacteria responsible for equol production. Nutrients 2019, 11, 433. [Google Scholar] [CrossRef]
- Yoshikata, R.; Myint, K.Z.; Ohta, H.; Ishigaki, Y. Inter-relationship between diet, lifestyle habits, gut microflora, and the equol-producer phenotype: Baseline findings from a placebo-controlled intervention trial. Menopause 2019, 26, 273–285. [Google Scholar] [CrossRef]
- Wells, C.L.; Jechorek, R.P.; Kinneberg, K.M.; Debol, S.M.; Erlandsen, S.L. The isoflavone genistein inhibits internalization of enteric bacteria by cultured Caco-2 and HT-29 enterocytes. J. Nutr. 1999, 129, 634–640. [Google Scholar] [CrossRef]
- Abron, J.D.; Singh, N.P.; Price, R.L.; Nagarkatti, M.; Nagarkatti, P.S.; Singh, U.P. Genistein induces macrophage polarization and systemic cytokine to ameliorate experimental colitis. PLoS ONE 2018, 13, e0199631. [Google Scholar] [CrossRef]
- Sato, Y.; Itagaki, S.; Oikawa, S.; Ogura, J.; Kobayashi, M.; Hirano, T.; Sugawara, M.; Iseki, K. Protective effect of soy isoflavone genistein on ischemia-reperfusion in the rat small intestine. Biol. Pharm. Bull. 2011, 34, 1448–1454. [Google Scholar] [CrossRef]
- Wu, Z.Y.; Sang, L.X.; Chang, B. Isoflavones and inflammatory bowel disease. World J. Clin. Cases 2020, 8, 2081–2091. [Google Scholar] [CrossRef]
- Jalili, M.; Vahedi, H.; Janani, L.; Poustchi, H.; Malekzadeh, R.; Hekmatdoost, A. Soy isoflavones supplementation for patients with irritable bowel syndrome: A randomized double blind clinical trial. Middle East J. Dig. Dis. 2015, 7, 170–176. [Google Scholar]
- Jalili, M.; Vahedi, H.; Poustchi, H.; Hekmatdoost, A. Soy isoflavones and cholecalciferol reduce inflammation, and gut permeability, without any effect on antioxidant capacity in irritable bowel syndrome: A randomized clinical trial. Clin. Nutr. ESPEN 2019, 34, 50–54. [Google Scholar] [CrossRef]
- Ahn-Jarvis, J.; Lombardo, E.; Cruz-Monserrate, Z.; Badi, N.; Crowe, O.; Kaul, S.; Komar, H.; Krishna, S.G.; Lesinski, G.B.; Mace, T.A.; et al. Reduction of inflammation in chronic pancreatitis using a soy bread intervention: A feasibility study. Pancreatology 2020, 20, 852–859. [Google Scholar] [CrossRef]
- Ahn-Jarvis, J.H.; Clinton, S.K.; Grainger, E.M.; Riedl, K.M.; Schwartz, S.J.; Lee, M.L.T.; Cruz-Cano, R.; Young, G.S.; Lesinski, G.B.; Vodovotz, Y. Isoflavone Pharmacokinetics and Metabolism after Consumption of a Standardized Soy and Soy–Almond Bread in Men with Asymptomatic Prostate Cancer. Cancer Prev. Res. 2015, 8, 1045–1054. [Google Scholar] [CrossRef]
- Ahn-Jarvis, J.H.; Riedl, K.M.; Schwartz, S.J.; Vodovotz, Y. Design and selection of soy breads used for evaluating isoflavone bioavailability in clinical trials. J. Agric. Food Chem. 2013, 61, 3111–3120. [Google Scholar] [CrossRef]
- Grainger, E.M.; Moran, N.E.; Francis, D.M.; Schwartz, S.J.; Wan, L.; Thomas-Ahner, J.; Kopec, R.E.; Riedl, K.M.; Young, G.S.; Abaza, R.; et al. A novel tomato-soy juice induces a dose-response increase in urinary and plasma phytochemical biomarkers in men with prostate cancer. J. Nutr. 2019, 149, 26–35. [Google Scholar] [CrossRef]
- Dhingra, S.; Jood, S. Organoleptic and nutritional evaluation of wheat breads supplemented with soybean and barley flour. Food Chem. 2002, 77, 479–488. [Google Scholar] [CrossRef]
- Ahn-Jarvis, J.H.; Schwartz, S.J.; Vodovotz, Y. Optimizing isoflavone-rich food delivery systems for human clinical trials. In Isoflavones: Chemistry, Analysis, Function and Effects; Preedy, V., Ed.; Royal Society of Chemistry Publishing: Cambridge, UK, 2012; pp. 399–422. [Google Scholar] [CrossRef]
- Ratnawati, L.; Desnilasari, D.; Surahman, D.N.; Kumalasari, R. Evaluation of physicochemical, functional and pasting properties of soybean, mung bean and red kidney bean flour as ingredient in biscuit. IOP Conf. Ser. Earth Environ. Sci. 2019, 251, 012026. [Google Scholar] [CrossRef]
- Jin, M.; Shen, M.H.; Jin, M.H.; Jin, A.H.; Yin, X.Z.; Quan, J.S. Hypoglycemic property of soy isoflavones from hypocotyl in Goto-Kakizaki diabetic rats. J. Clin. Biochem. Nutr. 2018, 62, 148–154. [Google Scholar] [CrossRef] [PubMed]
- Ronis, M.J.; Chen, Y.; Badeaux, J.; Badger, T.M. Dietary soy protein isolate attenuates metabolic syndrome in rats via effects on PPAR, LXR, and SREBP signaling. J. Nutr. 2009, 139, 1431–1438. [Google Scholar] [CrossRef] [PubMed]
- Mezei, O.; Banz, W.J.; Steger, R.W.; Peluso, M.R.; Winters, T.A.; Shay, N. Soy isoflavones exert antidiabetic and hypolipidemic effects through the PPAR pathways in obese Zucker rats and murine RAW 264.7 cells. J. Nutr. 2003, 133, 1238–1243. [Google Scholar] [CrossRef] [PubMed]
- Jayagopal, V.; Albertazzi, P.; Kilpatrick, E.S.; Howarth, E.M.; Jennings, P.E.; Hepburn, D.A.; Atkin, S.L. Beneficial effects of soy phytoestrogen intake in postmenopausal women with type 2 diabetes. Diabetes Care 2002, 25, 1709–1714. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.M.; Chen, Y.M.; Ho, S.C. Effects of soy intake on glycemic control: A meta-analysis of randomized controlled trials. Am. J. Clin. Nutr. 2011, 93, 1092–1101. [Google Scholar] [CrossRef]
- Roberts, K.M.; Golian, P.; Nahikian-Nelms, M.; Hinton, A.; Madril, P.; Basch, K.; Conwell, D.; Hart, P.A. Does the Healthy Eating Index and Mediterranean Diet Score identify the nutritional adequacy of dietary patterns in chronic pancreatitis? Dig. Dis. Sci. 2019, 64, 2318–2326. [Google Scholar] [CrossRef]
- Adlercreutz, H.; Honjo, H.; Higashi, A.; Fotsis, T.; Hämäläinen, E.; Hasegawa, T.; Okada, H. Urinary excretion of lignans and isoflavonoid phytoestrogens in Japanese men and women consuming a traditional Japanese diet. Am. J. Clin. Nutr. 1991, 54, 1093–1100. [Google Scholar] [CrossRef]
- Setchell, K.D.; Clerici, C. Equol: History, chemistry, and formation. J. Nutr. 2010, 140, 1355S–1362S. [Google Scholar] [CrossRef]
- Gu, L.; House, S.E.; Prior, R.L.; Fang, N.; Ronis, M.J.; Clarkson, T.B.; Wilson, M.E.; Badger, T.M. Metabolic phenotype of isoflavones differ among female rats, pigs, monkeys, and women. J. Nutr. 2006, 136, 1215–1221. [Google Scholar] [CrossRef]
- Roberts, K.M.; Hart, P.A.; Duggan, S. Revisiting Dietary Approaches in the Management of Chronic Pancreatitis. Curr. Treat. Options Gastroenterol. 2022, 20, 605–623. [Google Scholar] [CrossRef]
- Holscher, H.D.; Caporaso, J.G.; Hooda, S.; Brulc, J.M.; Fahey Jr, G.C.; Swanson, K.S. Fiber supplementation influences phylogenetic structure and functional capacity of the human intestinal microbiome: Follow-up of a randomized controlled trial. Am. J. Clin. Nutr. 2015, 101, 55–64. [Google Scholar] [CrossRef]
- Lynch, S.V.; Pedersen, O. The human intestinal microbiome in health and disease. N. Engl. J. Med. 2016, 375, 2369–2379. [Google Scholar] [CrossRef]
- Salminen, S.; Bouley, C.; Boutron, M.C.; Cummings, J.H.; Franck, A.; Gibson, G.R.; Isolauri, E.; Moreau, M.C.; Roberfroid, M.; Rowland, I. Functional food science and gastrointestinal physiology and function. Br. J. Nutr. 1998, 80, S147–S171. [Google Scholar] [CrossRef]
- Bulgari, R.; Franzoni, G.; Ferrante, A. Biostimulants application in horticultural crops under abiotic stress conditions. Agronomy 2019, 9, 306. [Google Scholar] [CrossRef]
- Newman, R.; Waterland, N.; Moon, Y.; Tou, J.C. Selenium biofortification of agricultural crops and effects on plant nutrients and bioactive compounds important for human health and disease prevention–a review. Plant Foods Hum. Nutr. 2019, 74, 449–460. [Google Scholar] [CrossRef]
- Kumar, S.; Pandey, G. Biofortification of pulses and legumes to enhance nutrition. Heliyon 2020, 6, e03682. [Google Scholar] [CrossRef]
- Tuncel, A.; Corbin, K.R.; Ahn-Jarvis, J.; Harris, S.; Hawkins, E.; Smedley, M.A.; Harwood, W.; Warren, F.J.; Patron, N.J.; Smith, A.M. Cas9-mediated mutagenesis of potato starch-branching enzymes generates a range of tuber starch phenotypes. Plant Biotechnol. J. 2019, 17, 2259–2271. [Google Scholar] [CrossRef]
- Parada, J.; Santos, J.L. Interactions between starch, lipids, and proteins in foods: Microstructure control for glycemic response modulation. Crit. Rev. Food Sci. Nutr. 2016, 56, 2362–2369. [Google Scholar] [CrossRef]
- Hart, P.A.; Andersen, D.K.; Lyons, E.; Cote, G.A.; Cruz-Monserrate, Z.; Dworkin, R.H.; Elmunzer, B.J.; Fogel, E.L.; Forsmark, C.E.; Gilron, I.; et al. Clinical Trials in Pancreatitis: Opportunities and Challenges in the Design and Conduct of Patient-Focused Clinical Trials in Recurrent Acute and Chronic Pancreatitis: Summary of a National Institute of Diabetes and Digestive and Kidney Diseases Workshop. Pancreas 2022, 51, 715–722. [Google Scholar] [CrossRef]
- Mace, T.A.; Ware, M.B.; King, S.A.; Loftus, S.; Farren, M.R.; McMichael, E.; Scoville, S.; Geraghty, C.; Young, G.; Carson III, W.E.; et al. Soy isoflavones and their metabolites modulate cytokine-induced natural killer cell function. Sci. Rep. 2019, 9, 5068. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Nutrition Parameters | |
|---|---|
| Serving Size (g) | 92.4 ± 2.3 |
| Nutrient Composition 1 | |
| Energy (kcal) | 220.8 |
| Fat (g) | 3.4 |
| Total Carbohydrates (g) | 37.8 |
| Dietary Fiber (g) | 3.7 |
| Protein (g) | 15.2 |
| Total Isoflavones (Mean ± SD, mg) 2 | 34.38 ± 0.21 |
| Daidzein (mg) | 1.44 ± 0.10 |
| Daidzin (mg) | 3.81 ± 0.23 |
| 6″-O-Acetyldaidzin (mg) | 0.54 ± 0.03 |
| 6″-O-Malonyldaidzin (mg) | 6.48 ± 0.39 |
| Genistein (mg) | 1.84 ± 0.14 |
| Genistin (mg) | 6.92 ± 0.40 |
| 6″-O-Acetylgenistin (mg) | 0.67 ± 0.03 |
| 6″-O-Malonylgenistin (mg) | 11.67 ± 0.63 |
| Glycitein (mg) | 0.10 ± 0.01 |
| Glycitin (mg) | 0.77 ± 0.04 |
| 6″-O-Acetylglycitin (mg) | Trivial 3 |
| 6″-O-Malonylglycitin (mg) | 0.12 ± 0.01 |
| Participant Information | Dose Escalating Phase (Phase 1) | MTD 1 (Phase 2) | ||
|---|---|---|---|---|
| 1 Slice/Day (n = 3) | 2 Slice/Day (n = 3) | 3 Slice/Day (n = 3) 2 | 3 Slice/Day (n = 1) | |
| Clinical Features | ||||
| Body Mass Index (kg/m2,mean ± SD) | 23.4 ± 5.2 | 28.4 ± 1.6 | 26.5 ± 3.9 | 24.6 |
| Lipase (PERT) with meals (%) | 66 | 100 | 66 | yes |
| Lipase dose range (1000 units/meal) | 24 and 72 | 24 to 48 | 24 and 72 | 24 |
| Diabetes mellitus (%) | 33 | 100 | 66 | no |
| Pancreatic calcification/calculi (%) | 100 | 100 | 100 | yes |
| Main Pancreatic Duct Dilatation (%) | 66 | 66 | 100 | no |
| Clinical Laboratory Values (mean ± SD) 3 | ||||
| Creatinine (mg/dL) | ||||
| Pre-intervention | 1.03 ± 0.06 | 1.17 ± 0.02 | 0.94 ± 0.09 | 1.49 |
| Post-intervention | 0.09 ± 0.06 | 1.24 ± 0.05 | 0.89 ± 0.07 | 1.49 |
| Blood glucose (mg/dL) | ||||
| Pre-intervention | 104 ± 6 | 172 ± 52 | 236 ± 141 | 82 |
| Post-intervention | 113 ± 8 | 139 ± 24 | 194 ± 79 | 89 |
| Alanine aminotransferase (IU/L) | ||||
| Pre-intervention | 16 ± 5 | 22 ± 10 | 17 ± 2 | 12 |
| Post-intervention | 18 ± 4 | 26 ± 10 | 17 ± 3 | 16 |
| Aspartate aminotransferase (IU/L) | ||||
| Pre-intervention | 16 ± 1 | 18 ± 4 | 17 ± 1 | 19 |
| Post-intervention | 22 ± 4 | 19 ± 4 | 18 ± 3 | 21 |
| Total bilirubin (mg/dL) | ||||
| Pre-intervention | 0.6 ± 0.1 | 0.4 ± 0.1 | 0.7 ± 0.1 | 0.4 |
| Post-intervention | 0.5 ± 0.1 | 0.4 ± 0.1 | 0.7 ± 0.1 | 0.4 |
| Albumin (g/dL) | ||||
| Pre-intervention | 4.3 ± 0.2 | 4.3 ± 0.3 | 4.1 ± 0.2 | 3.6 |
| Post-intervention | 4.3 ± 0.2 | 4.3 ± 0.3 | 4.1 ± 0.2 | 3.6 |
| Alkaline phosphatase (IU/L) | ||||
| Pre-intervention | 74 ± 12 | 110 ± 25 | 101 ± 45 | 98 |
| Post-intervention | 70 ± 7 | 109 ± 16 | 108 ± 34 | 101 |
| Daily Nutrient Intake (mean ± SEM) 4 | ||||
| Total Energy (kcal) | 2300 ± 288 | 2095 ± 240 | 1517 ± 221 | 1592 ± 158 |
| Total fat (g) | 95.0 ± 16.8 | 75.2 ± 11.7 | 65.7 ± 14.4 | 49.0 ± 18.6 |
| Cholesterol (mg) | 391.4 ± 109.5 | 174.3 ± 33.2 | 216.9 ± 64.5 | 81.6 ± 71.1 |
| Total carbohydrate (g) | 278.1 ± 32.2 | 288.8 ± 38.5 | 191.5 ± 31.8 | 252.7 ± 46.3 |
| Dietary fiber (g) | 21.4 ± 5.6 | 18.4 ± 2.9 | 25.6 ± 5.5 | 24.9 ± 11.3 |
| Total protein(g) 5 | 96.7 ± 13.4 a | 74.6 ± 8.3 ab | 48.2 ± 9.6 b | 50.1 ± 12.1 |
| Animal protein (g) 5 | 74.7 ± 11.3 a | 46.4 ± 9.6 ab | 28.6 ± 7.7 b | 18.5 ± 12.6 |
| Vegetable protein (g) | 22.0 ± 3.6 | 28.2 ± 3.3 | 19.6 ± 3.4 | 29.1 ± 8.2 |
| Total water (g) | 3296 ± 936 | 2149 ± 517 | 2083 ± 514 | 2143 ± 189 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ahn-Jarvis, J.H.; Sosh, D.; Lombardo, E.; Lesinski, G.B.; Conwell, D.L.; Hart, P.A.; Vodovotz, Y. Short-Term Soy Bread Intervention Leads to a Dose-Response Increase in Urinary Isoflavone Metabolites and Satiety in Chronic Pancreatitis. Foods 2023, 12, 1762. https://doi.org/10.3390/foods12091762
Ahn-Jarvis JH, Sosh D, Lombardo E, Lesinski GB, Conwell DL, Hart PA, Vodovotz Y. Short-Term Soy Bread Intervention Leads to a Dose-Response Increase in Urinary Isoflavone Metabolites and Satiety in Chronic Pancreatitis. Foods. 2023; 12(9):1762. https://doi.org/10.3390/foods12091762
Chicago/Turabian StyleAhn-Jarvis, Jennifer H., Daniel Sosh, Erin Lombardo, Gregory B. Lesinski, Darwin L. Conwell, Phil A. Hart, and Yael Vodovotz. 2023. "Short-Term Soy Bread Intervention Leads to a Dose-Response Increase in Urinary Isoflavone Metabolites and Satiety in Chronic Pancreatitis" Foods 12, no. 9: 1762. https://doi.org/10.3390/foods12091762
APA StyleAhn-Jarvis, J. H., Sosh, D., Lombardo, E., Lesinski, G. B., Conwell, D. L., Hart, P. A., & Vodovotz, Y. (2023). Short-Term Soy Bread Intervention Leads to a Dose-Response Increase in Urinary Isoflavone Metabolites and Satiety in Chronic Pancreatitis. Foods, 12(9), 1762. https://doi.org/10.3390/foods12091762