
Bamboo Shoots Modulate Gut Microbiota, Eliminate Obesity in High-Fat-Diet-Fed Mice and Improve Lipid Metabolism



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Animals and Diets
2.3. Histological Analysis
2.4. Determination of Blood Biochemical Parameters in Serum
2.5. Glucose Tolerance Test
2.6. Determination of Insulin in Serum
2.7. Gut Microbiota Analysis
2.8. Quantification of Fecal Short-Chain Fatty Acids (SCFAs)
2.9. Determination of Metabolites in Feces
2.10. Statistical Analysis
3. Results
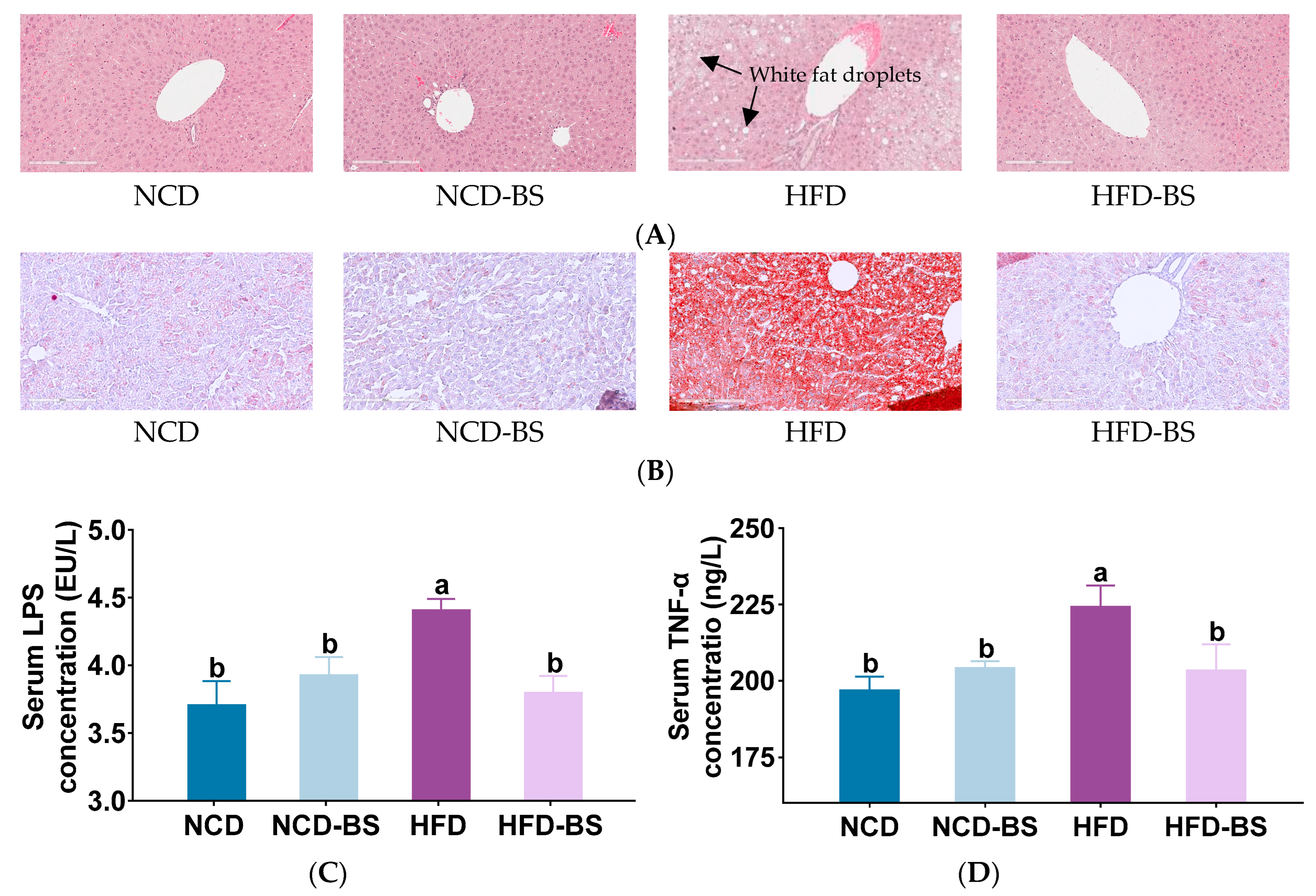
3.1. BS Alleviated Obesity Induced by HFD
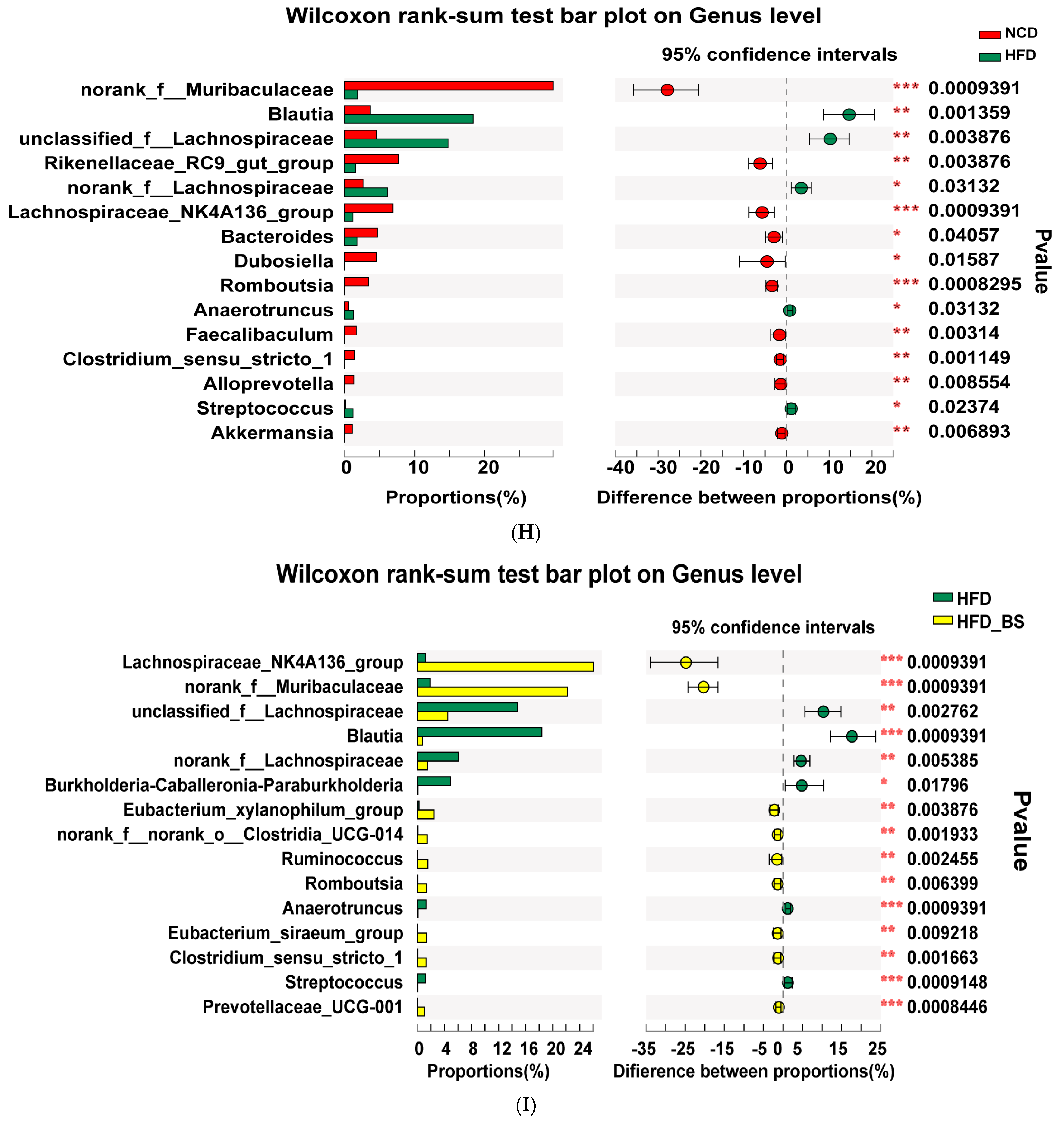
3.2. BS Improved Gut Dysbiosis in Obese Mice
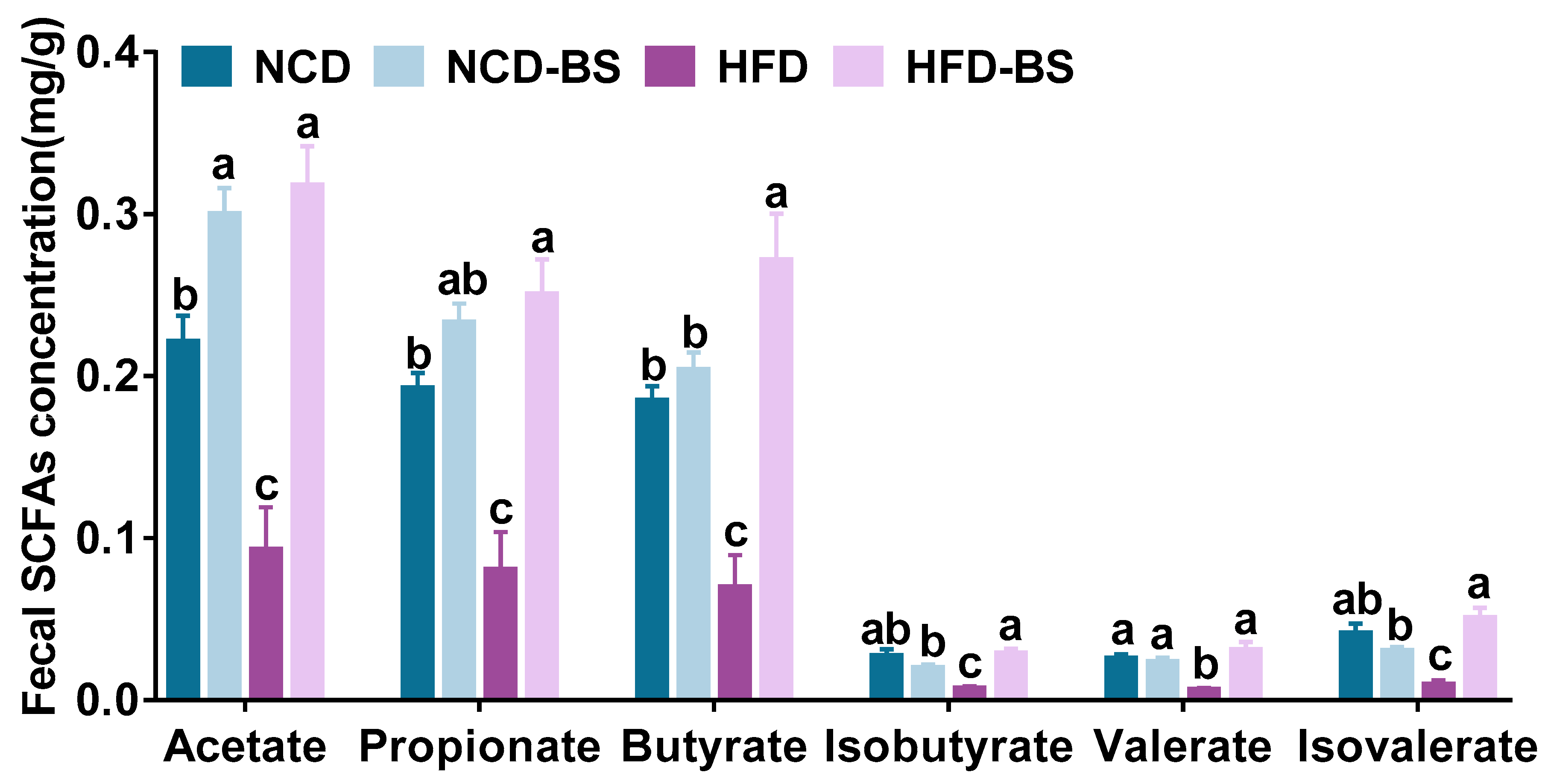
3.3. BS Promotes the Production of SCFAs
3.4. Effects of BS on Fecal Metabolites in High-Fat Diet-Fed Mice
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wang, P.; Li, D.; Ke, W.; Liang, D.; Hu, X.; Chen, F. Resveratrol-Induced Gut Microbiota Reduces Obesity in High-Fat Diet-Fed Mice. Int. J. Obes. 2020, 44, 213–225. [Google Scholar] [CrossRef] [PubMed]
- Carmody, R.N.; Gerber, G.K.; Luevano Jr, J.M.; Gatti, D.M.; Somes, L.; Svenson, K.L.; Turnbaugh, P.J. Diet Dominates Host Genotype in Shaping the Murine Gut Microbiota. Cell Host Microbe 2015, 17, 72–84. [Google Scholar] [CrossRef] [PubMed]
- Cook, K.S.; Min, H.Y.; Johnson, D.; Chaplinsky, R.J.; Flier, J.S.; Hunt, C.R. Adipsin: A Circulating Serine Protease Homolog Secreted by Adipose Tissue and Sciatic Nerve. Science 1987, 237, 402–405. [Google Scholar] [CrossRef]
- Reilly, S.M.; Saltiel, A.R. Adapting to Obesity with Adipose Tissue Inflammation. Nat. Rev. Endocrinol. 2017, 13, 633–643. [Google Scholar] [CrossRef]
- Rosen, E.D.; Spiegelman, B.M. What We Talk About When We Talk About Fat. Cell 2014, 156, 20–44. [Google Scholar] [CrossRef]
- Huo, Y.; Lu, X.; Wang, X.; Wang, X.; Chen, L.; Guo, H.; Zhang, M.; Li, Y. Bifidobacterium Animalis Subsp. Lactis A6 Alleviates Obesity Associated with Promoting Mitochondrial Biogenesis and Function of Adipose Tissue in Mice. Molecules 2020, 25, 1490. [Google Scholar] [CrossRef]
- Wang, X.; Zhao, B.; Sun, H.; You, H.; Qu, S. Effects of Sitagliptin on Intrahepatic Lipid Content in Patients with Non-Alcoholic Fatty Liver Disease. Front. Endocrinol. 2022, 13, 866189. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Q.; Fu, Y.; Zhang, F.; Wang, C.; Yang, X.; Bai, S.; Xue, Y.; Shen, Q. Heat-treated Adzuki Bean Protein Hydrolysates Reduce Obesity in Mice Fed a High-fat Diet via Remodeling Gut Microbiota and Improving Metabolic Function. Mol. Nutr. Food Res. 2022, 66, 2100907. [Google Scholar] [CrossRef]
- Backhed, F.; Manchester, J.K.; Semenkovich, C.F.; Gordon, J.I. Mechanisms Underlying the Resistance to Diet-Induced Obesity in Germ-Free Mice. Proc. Natl. Acad. Sci. USA 2007, 104, 979–984. [Google Scholar] [CrossRef]
- Clemente, J.C.; Ursell, L.K.; Parfrey, L.W.; Knight, R. The Impact of the Gut Microbiota on Human Health: An Integrative View. Cell 2012, 148, 1258–1270. [Google Scholar] [CrossRef]
- Nicolas, G.R.; Chang, P.V. Deciphering the Chemical Lexicon of Host-Gut Microbiota Interactions. Trends Pharmacol. Sci. 2019, 40, 430–445. [Google Scholar] [CrossRef]
- Denou, E.; Marcinko, K.; Surette, M.G.; Steinberg, G.R.; Schertzer, J.D. High-Intensity Exercise Training Increases the Diversity and Metabolic Capacity of the Mouse Distal Gut Microbiota during Diet-Induced Obesity. Am. J. Physiol. Metab. 2016, 310, E982–E993. [Google Scholar] [CrossRef]
- Heiss, C.N.; Olofsson, L.E. Gut Microbiota-Dependent Modulation of Energy Metabolism. J. Innate Immun. 2018, 10, 163–171. [Google Scholar] [CrossRef]
- Papathanasopoulos, A.; Camilleri, M. Dietary Fiber Supplements: Effects in Obesity and Metabolic Syndrome and Relationship to Gastrointestinal Functions. Gastroenterology 2010, 138, 65–72. [Google Scholar] [CrossRef] [PubMed]
- Sang, T.T.; Guo, C.J.; Guo, D.D.; Wu, J.J.; Wang, Y.J.; Wang, Y.; Chen, J.J.; Chen, C.J.; Wu, K.K.; Na, K.; et al. Suppression of Obesity and Inflammation by Polysaccharide from Sporoderm-Broken Spore of Ganoderma Lucidum via Gut Microbiota Regulation. Carbohydr. Polym. 2021, 256, 117594. [Google Scholar] [CrossRef]
- Kałużna-Czaplińska, J.; Gątarek, P.; Chartrand, M.S.; Dadar, M.; Bjørklund, G. Is There a Relationship between Intestinal Microbiota, Dietary Compounds, and Obesity? Trends Food Sci. Technol. 2017, 70, 105–113. [Google Scholar] [CrossRef]
- Singh, P.; Rathore, M.; Prakash, H.G. The Nutritional Facts of Bamboo Shoots Have a Potential and Prospects for Utilization as a Health Food: A Review. Asian J. Dairy Food Res. 2021, 40, 388–397. [Google Scholar] [CrossRef]
- Bal, L.M.; Kar, A.; Satya, S.; Naik, S.N. Kinetics of Colour Change of Bamboo Shoot Slices during Microwave Drying. Int. J. Food Sci. Technol. 2011, 46, 827–833. [Google Scholar] [CrossRef]
- Lu, B.; Cai, H.; Huang, W.; Wu, X.; Luo, Y.; Liu, L.; Ying, Z. Protective Effect of Bamboo Shoot Oil on Experimental Nonbacterial Prostatitis in Rats. Food Chem. 2011, 124, 1017–1023. [Google Scholar] [CrossRef]
- Zheng, Y.; Zhang, S.; Wang, Q.; Lu, X.; Lin, L.; Tian, Y.; Xiao, J.; Zheng, B. Characterization and Hypoglycemic Activity of a Beta-Pyran Polysaccharides from Bamboo Shoot (Leleba Oldhami Nakal) Shells. Carbohydr. Polym. Sci. Technol. Asp. Ind. Important Polysacch. 2016, 144, 438–446. [Google Scholar]
- Li, X.; Guo, J.; Ji, K.; Zhang, P. Bamboo Shoot Fiber Prevents Obesity in Mice by Modulating the Gut Microbiota. Sci. Rep. 2016, 6, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Hu, J.L.; Nie, S.P.; Min, F.F.; Xie, M.Y. Polysaccharide from Seeds of Plantago Asiatica L. Increases Short-Chain Fatty Acid Production and Fecal Moisture along with Lowering PH in Mouse Colon. J. Agric. Food Chem. 2012, 60, 11525–11532. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Fu, B.; Guo, J.; Ji, K.; Xu, Y.; Dahab, M.M.; Zhang, P. Bamboo Shoot Fiber Improves Insulin Sensitivity in High-Fat Diet-Fed Mice. J. Funct. Foods 2018, 49, 510–517. [Google Scholar] [CrossRef]
- Ren, Y.; Ma, Y.; Zhang, Z.; Qiu, L.; Zhai, H.; Gu, R.; Xie, Y. Total Alkaloids from Bamboo Shoots and Bamboo Shoot Shells of Pleioblastus Amarus (Keng) Keng f. and Their Anti-Inflammatory Activities. Molecules 2019, 24, 2699. [Google Scholar] [CrossRef]
- Arora, T.; Bäckhed, F. The Gut Microbiota and Metabolic Disease: Current Understanding and Future Perspectives. J. Intern. Med. 2016, 280, 339–349. [Google Scholar] [CrossRef] [PubMed]
- Van Gylswyk, N.O.; Van der Toorn, J. Eubacterium Uniforme Sp. Nov. and Eubacterium Xylanophilum Sp. Nov., Fiber-Digesting Bacteria from the Rumina of Sheep Fed Corn Stover. Int. J. Syst. Evol. Microbiol. 1985, 35, 323–326. [Google Scholar] [CrossRef]
- Kang, Y.; Cai, Y. Gut Microbiota and Obesity: Implications for Fecal Microbiota Transplantation Therapy. Hormones 2017, 16, 223–234. [Google Scholar] [CrossRef]
- Hou, D.; Zhao, Q.; Yousaf, L.; Khan, J.; Xue, Y.; Shen, Q. Consumption of Mung Bean (Vigna Radiata L.) Attenuates Obesity, Ameliorates Lipid Metabolic Disorders and Modifies the Gut Microbiota Composition in Mice Fed a High-Fat Diet. J. Funct. Foods 2020, 64, 103687. [Google Scholar] [CrossRef]
- Xia, F.; Xiang, S.; Chen, Z.; Song, L.; Li, Y.; Liao, Z.; Ge, B.; Zhou, B. The Probiotic Effects of AB23A on High-Fat-Diet-Induced Non-Alcoholic Fatty Liver Disease in Mice May Be Associated with Suppressing the Serum Levels of Lipopolysaccharides and Branched-Chain Amino Acids. Arch. Biochem. Biophys. 2021, 714, 109080. [Google Scholar] [CrossRef]
- Becker, N.; Kunath, J.; Loh, G.; Blaut, M. Human Intestinal Microbiota: Characterization of a Simplified and Stable Gnotobiotic Rat Model. Gut Microbes 2011, 2, 25–33. [Google Scholar] [CrossRef]
- Xu, S.; Dong, Y.; Shi, J.; Li, Z.; Che, L.; Lin, Y.; Li, J.; Feng, B.; Fang, Z.; Yong, Z. Responses of Vaginal Microbiota to Dietary Supplementation with Lysozyme and Its Relationship with Rectal Microbiota and Sow Performance from Late Gestation to Early Lactation. Animals 2021, 11, 593. [Google Scholar] [CrossRef] [PubMed]
- Haixia, W.; Ping, C.; Qingnan, Z.; Lin, Y.; Shan, T. Nutrition Components in 9 Provenances of Chimonobambusa Quadrangularis Shoots in Jiangxi Province. World Bamboo Ratt. 2018, 16, 3. [Google Scholar]
- JinLing, Y.; DengGao, X.; BingTang, H.; Guang, J.; ZhiQi, Z.; LiangJiang, H.; NaiXun, M. Study on Shoot Nutrition of Qiongzhuea Tumidinoda: A Rare and Protected Bamboo Species. For. Res. Beijing 2008, 21, 773–777. [Google Scholar]
- Boutard, M.; Cerisy, T.; Nogue, P.-Y.; Alberti, A.; Weissenbach, J.; Salanoubat, M.; Tolonen, A.C. Functional Diversity of Carbohydrate-Active Enzymes Enabling a Bacterium to Ferment Plant Biomass. PLoS Genet. 2014, 10, e1004773. [Google Scholar] [CrossRef]
- Li, H.; Liu, F.; Lu, J.; Shi, J.; Guan, J.; Yan, F.; Li, B.; Huo, G. Probiotic Mixture of Lactobacillus Plantarum Strains Improves Lipid Metabolism and Gut Microbiota Structure in High Fat Diet-Fed Mice. Front. Microbiol. 2020, 11, 512. [Google Scholar] [CrossRef]
- Smith, B.J.; Miller, R.A.; Ericsson, A.C.; Harrison, D.C.; Strong, R.; Schmidt, T.M. Changes in the Gut Microbiome and Fermentation Products Concurrent with Enhanced Longevity in Acarbose-Treated Mice. BMC Microbiol. 2019, 19, 1–16. [Google Scholar] [CrossRef]
- Ma, S.; You, Y.; Huang, L.; Long, S.; Zhang, J.; Guo, C.; Zhang, N.; Wu, X.; Xiao, Y.; Tan, H. Alterations in Gut Microbiota of Gestational Diabetes Patients during the First Trimester of Pregnancy. Front. Cell. Infect. Microbiol. 2020, 10, 58. [Google Scholar] [CrossRef] [PubMed]
- Tao, Y.; Wang, T.; Huang, C.; Lai, C.; Ling, Z.; Zhou, Y.; Yong, Q. Incomplete Degradation Products of Galactomannan from Sesbania Canabina Modulated the Caecal Microbial Community of Laying Hens. J. Anim. Sci. 2022, 100, skac087. [Google Scholar] [CrossRef]
- Hu, X.; Xie, Y.; Xiao, Y.; Zeng, W.; Gong, Z.; Du, J. Longitudinal Analysis of Fecal Microbiome and Metabolome during Renal Fibrotic Progression in a Unilateral Ureteral Obstruction Animal Model. Eur. J. Pharmacol. 2020, 886, 173555. [Google Scholar] [CrossRef]
- MOORE, W.E.C.; Johnson, J.L.; Holdeman, L. V Emendation of Bacteroidaceae and Butyrivibrio and Descriptions of Desulfomonas Gen. Nov. and Ten New Species in the Genera Desulfomonas, Butyrivibrio, Eubacterium, Clostridium, and Ruminococcus. Int. J. Syst. Evol. Microbiol. 1976, 26, 238–252. [Google Scholar] [CrossRef]
- Shen, F.; Zheng, R.-D.; Sun, X.-Q.; Ding, W.-J.; Wang, X.-Y.; Fan, J.-G. Gut Microbiota Dysbiosis in Patients with Non-Alcoholic Fatty Liver Disease. Hepatobiliary Pancreat. Dis. Int. 2017, 16, 375–381. [Google Scholar] [CrossRef]
- Kühn, B.; Brat, C.; Fettel, J.; Hellmuth, N.; Maucher, I.V.; Bulut, U.; Hock, K.J.; Grimmer, J.; Manolikakes, G.; Rühl, M. Anti-Inflammatory Nitro-Fatty Acids Suppress Tumor Growth by Triggering Mitochondrial Dysfunction and Activation of the Intrinsic Apoptotic Pathway in Colorectal Cancer Cells. Biochem. Pharmacol. 2018, 155, 48–60. [Google Scholar] [CrossRef] [PubMed]
- Yapi, A.P.; Kouadio, A.I. Antimicrobial and Antioxidant Activities of Palm Kernel Oils Extracted from Varieties Dura and Tenera of Oil Palm (Elaeis Guineensis Jacq.) and the Relationship to Their Chemical Composition. J. Food Stud. 2020, 9, 95–117. [Google Scholar] [CrossRef]
- Shan, K.; Qu, H.; Zhou, K.; Wang, L.; Zhu, C.; Chen, H.; Gu, Z.; Cui, J.; Fu, G.; Li, J. Distinct Gut Microbiota Induced by Different Fat-to-Sugar-Ratio High-Energy Diets Share Similar pro-Obesity Genetic and Metabolite Profiles in Prediabetic Mice. Msystems 2019, 4, e00219-19. [Google Scholar] [CrossRef] [PubMed]
- Montaigne, D.; Butruille, L.; Staels, B. PPAR Control of Metabolism and Cardiovascular Functions. Nat. Rev. Cardiol. 2021, 18, 809–823. [Google Scholar] [CrossRef] [PubMed]
- Dunny, E.; Evans, P. Stereocontrolled Synthesis of the PPAR-γ Agonist 10-Nitrolinoleic Acid. J. Org. Chem. 2010, 75, 5334–5336. [Google Scholar] [CrossRef] [PubMed]
- Chaudhari, S.N.; Harris, D.A.; Aliakbarian, H.; Luo, J.N.; Henke, M.T.; Subramaniam, R.; Vernon, A.H.; Tavakkoli, A.; Sheu, E.G.; Devlin, A.S. Bariatric Surgery Reveals a Gut-Restricted TGR5 Agonist with Anti-Diabetic Effects. Nat. Chem. Biol. 2021, 17, 20–29. [Google Scholar] [CrossRef]
- Zhou, X.-R.; Sun, C.-H.; Liu, J.-R.; Zhao, D. Dietary Conjugated Linoleic Acid Increases PPARγ Gene Expression in Adipose Tissue of Obese Rat, and Improves Insulin Resistance. Growth Horm. IGF Res. 2008, 18, 361–368. [Google Scholar] [CrossRef]









Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhou, X.; Pak, S.; Li, D.; Dong, L.; Chen, F.; Hu, X.; Ma, L. Bamboo Shoots Modulate Gut Microbiota, Eliminate Obesity in High-Fat-Diet-Fed Mice and Improve Lipid Metabolism. Foods 2023, 12, 1380. https://doi.org/10.3390/foods12071380
Zhou X, Pak S, Li D, Dong L, Chen F, Hu X, Ma L. Bamboo Shoots Modulate Gut Microbiota, Eliminate Obesity in High-Fat-Diet-Fed Mice and Improve Lipid Metabolism. Foods. 2023; 12(7):1380. https://doi.org/10.3390/foods12071380
Chicago/Turabian StyleZhou, Xiaolu, SolJu Pak, Daotong Li, Li Dong, Fang Chen, Xiaosong Hu, and Lingjun Ma. 2023. "Bamboo Shoots Modulate Gut Microbiota, Eliminate Obesity in High-Fat-Diet-Fed Mice and Improve Lipid Metabolism" Foods 12, no. 7: 1380. https://doi.org/10.3390/foods12071380
APA StyleZhou, X., Pak, S., Li, D., Dong, L., Chen, F., Hu, X., & Ma, L. (2023). Bamboo Shoots Modulate Gut Microbiota, Eliminate Obesity in High-Fat-Diet-Fed Mice and Improve Lipid Metabolism. Foods, 12(7), 1380. https://doi.org/10.3390/foods12071380