
Changes of Bioactive Components and Antioxidant Capacity of Pear Ferment in Simulated Gastrointestinal Digestion In Vitro

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Plant Materials and Strains
2.2. Standards, Reagents, and Other Chemicals
2.3. Methods
2.3.1. Strain Activation and Culture
2.3.2. Preparation of ‘Xuehua’ Pear Ferment
2.3.3. Simulated Gastric and Intestinal Fluid Digestion In Vitro
2.3.4. Determination of TPC
2.3.5. Determination of TFC
2.3.6. Protease Activity
2.3.7. Superoxide Dismutase (SOD) Activity
2.3.8. Antioxidant Capacity
Determination of DPPH-RSA
Determination of ·OH-RSA
Total Reduction Capacity
2.3.9. Phenolic Components Analyses
2.3.10. Organic Acids Analyses
2.4. Statistical Analysis
3. Results
3.1. Effect of Simulated Digestion on TPC In Vitro
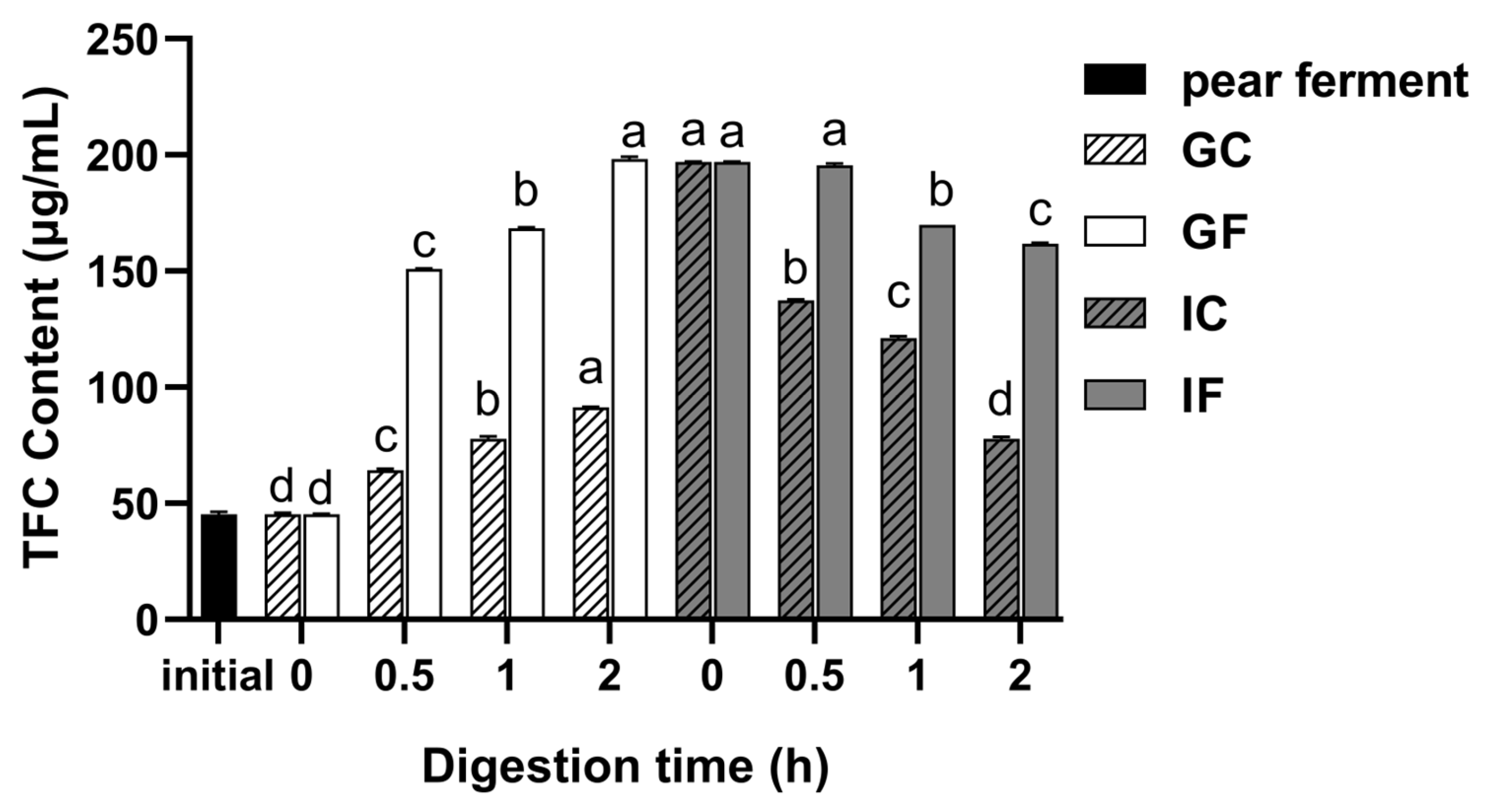
3.2. Effect of Simulated Digestion on TFC In Vitro
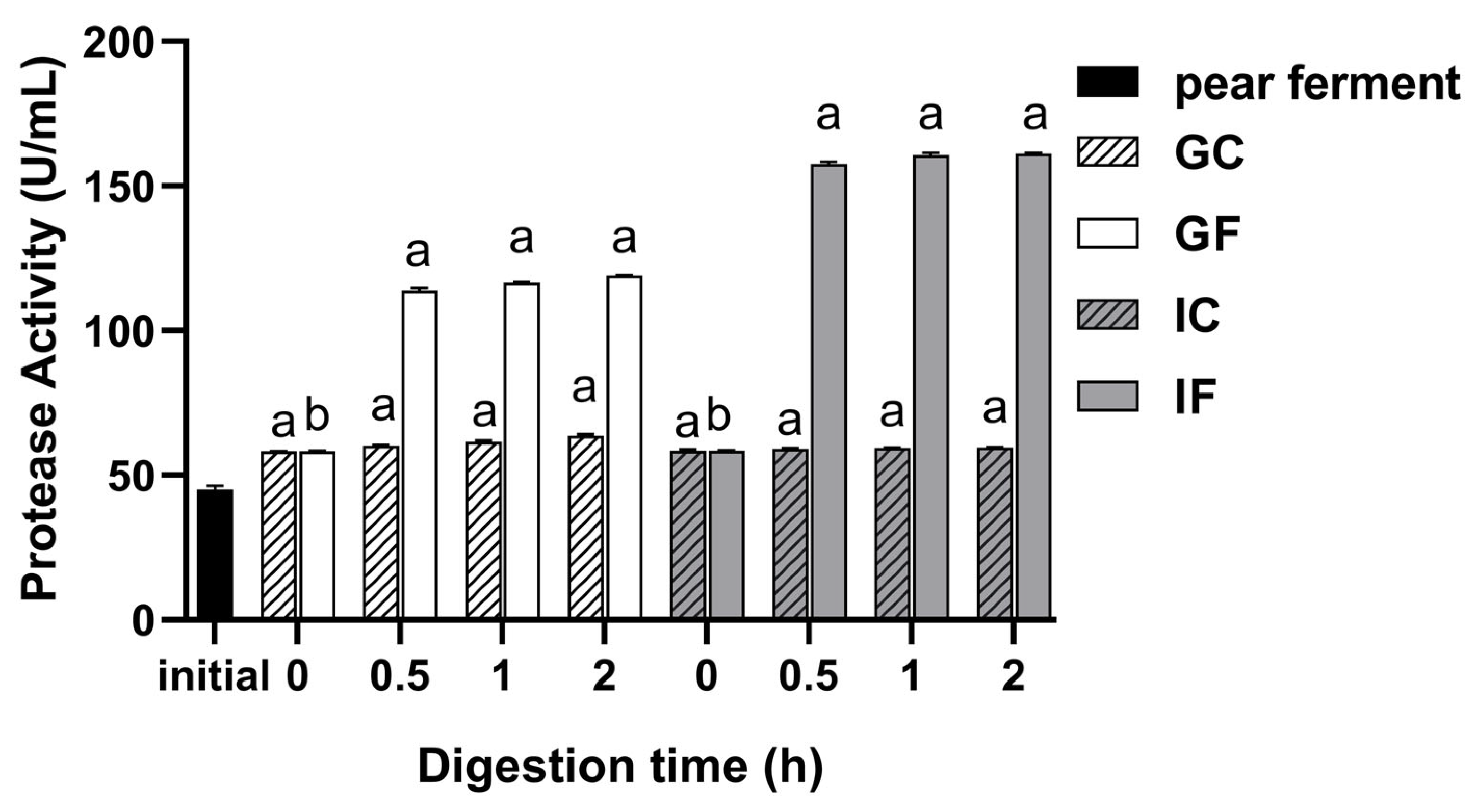
3.3. Effect of Simulated Digestion on Protease Activity In Vitro
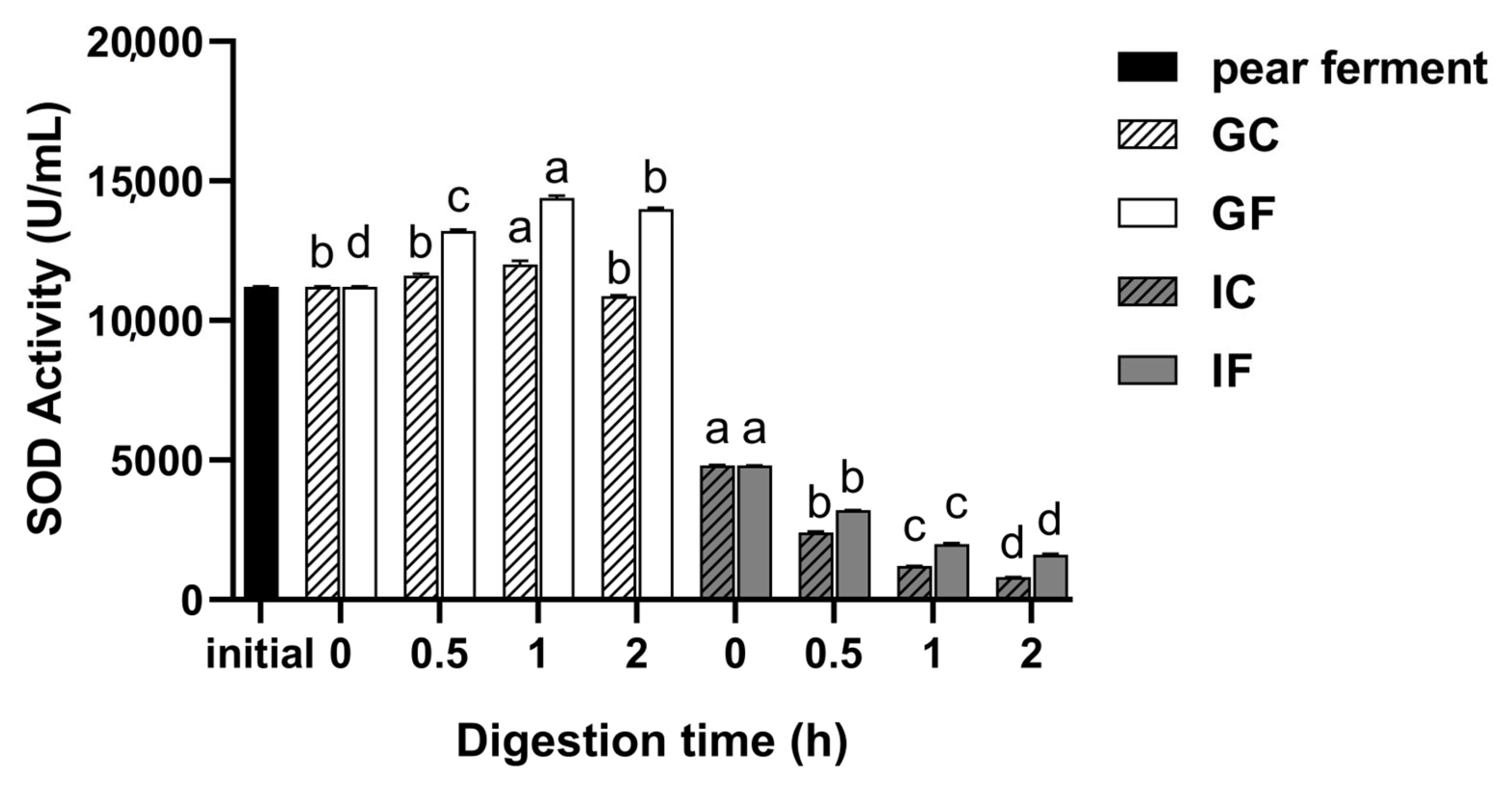
3.4. Effect of Simulated Digestion on SOD Activity In Vitro
3.5. Effect of Simulated Digestion on Antioxidant Capacity In Vitro
3.5.1. Changes in DPPH-RSA during Simulated Digestion In Vitro
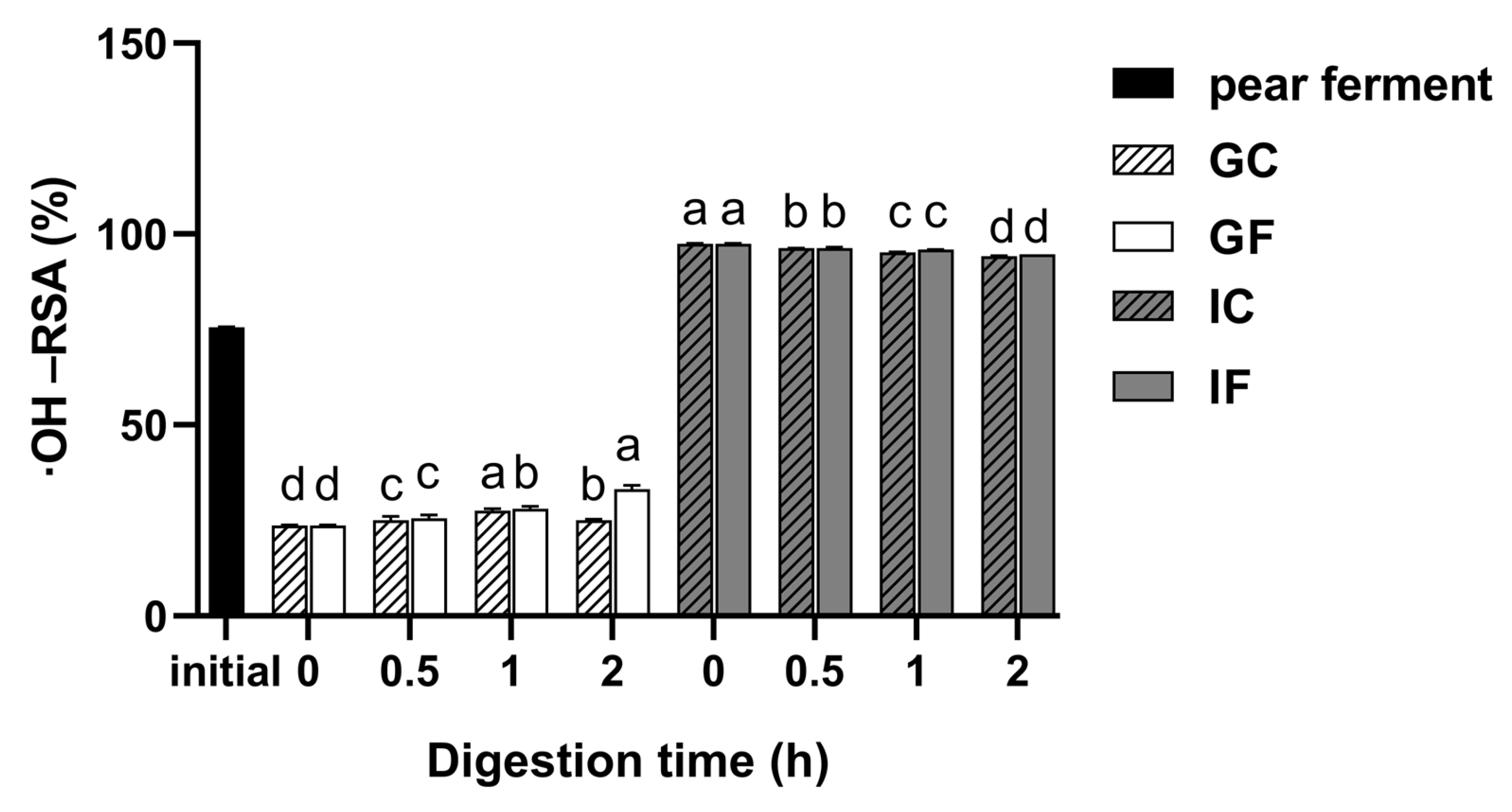
3.5.2. Changes in ·OH-RSA during Simulated Digestion In Vitro
3.5.3. Changes in the Capacity of Total Reducing during Simulated Digestion In Vitro
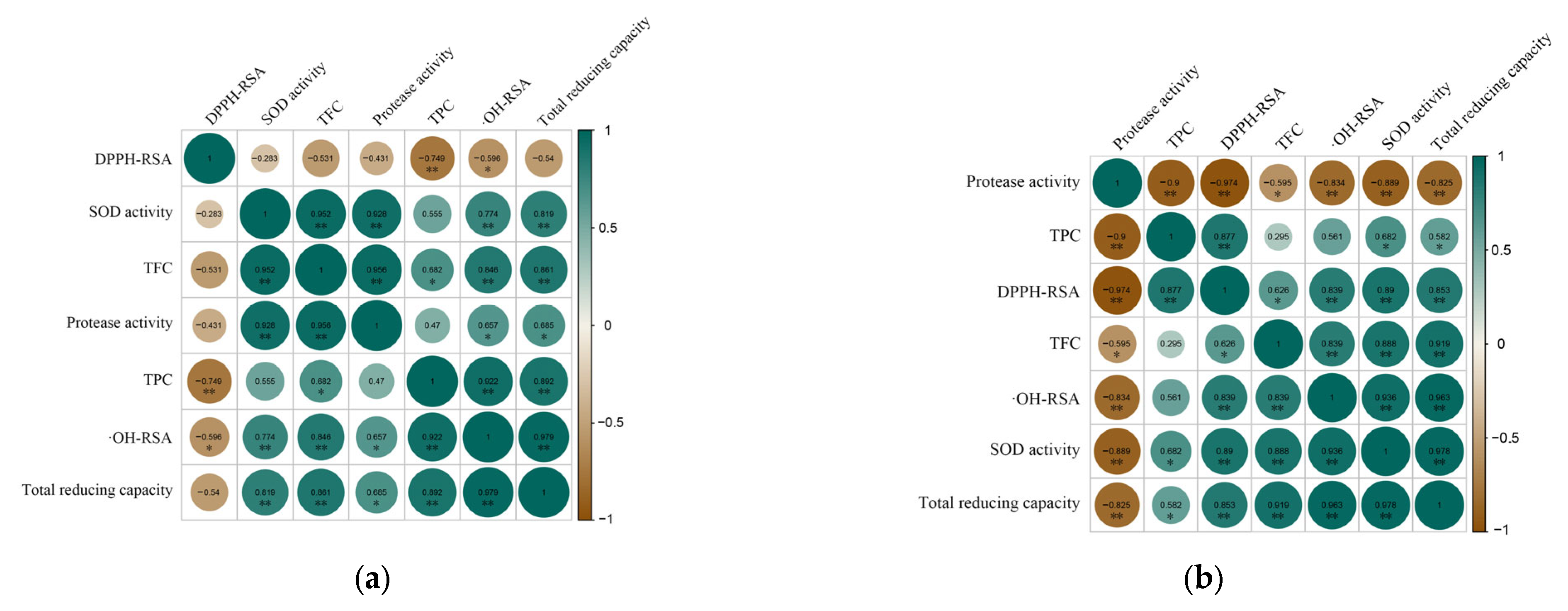
3.5.4. Correlation Analysis between Bioactive Ingredients and Antioxidant Activity
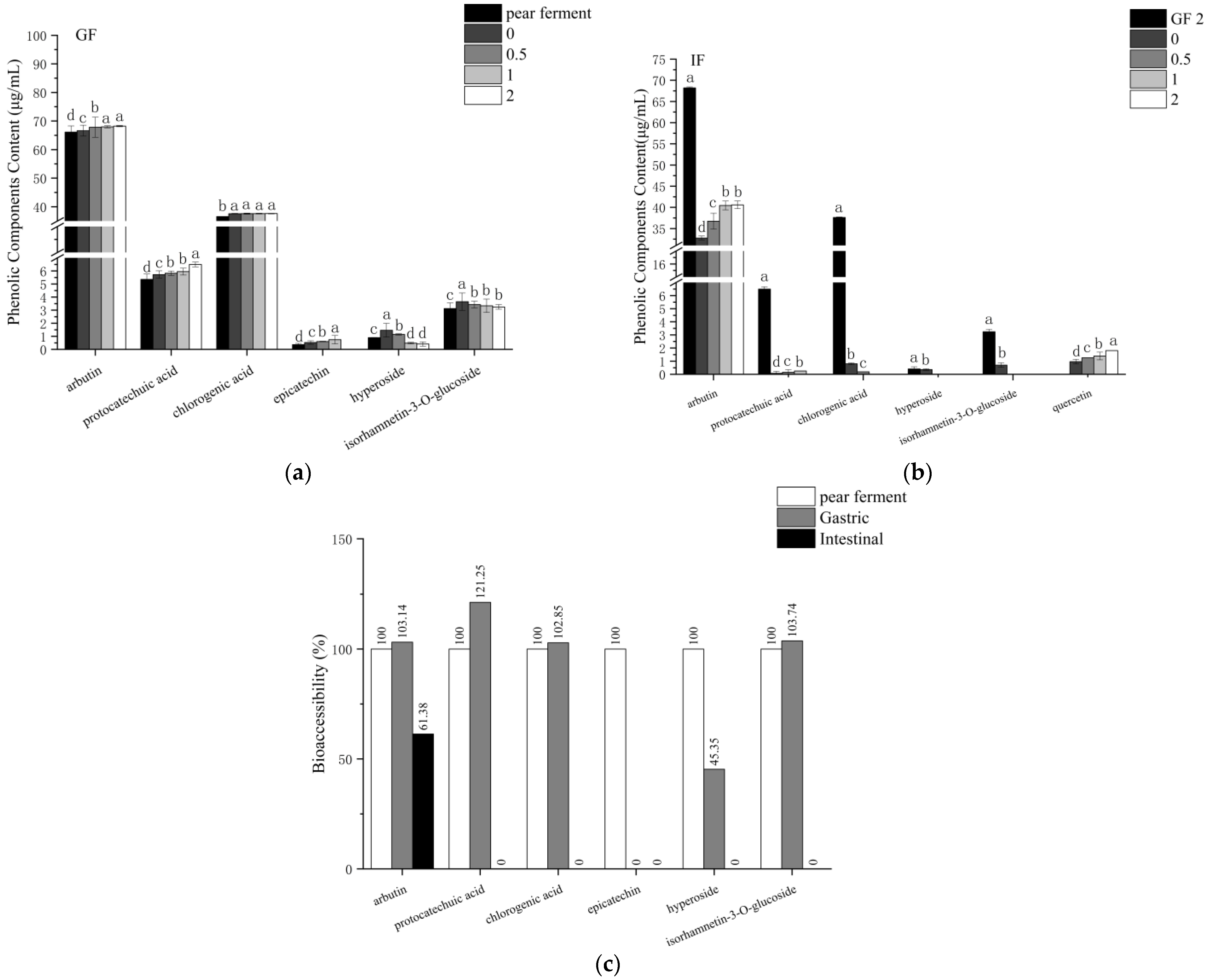
3.6. Effect of Simulated Digestion on Phenolic Components In Vitro
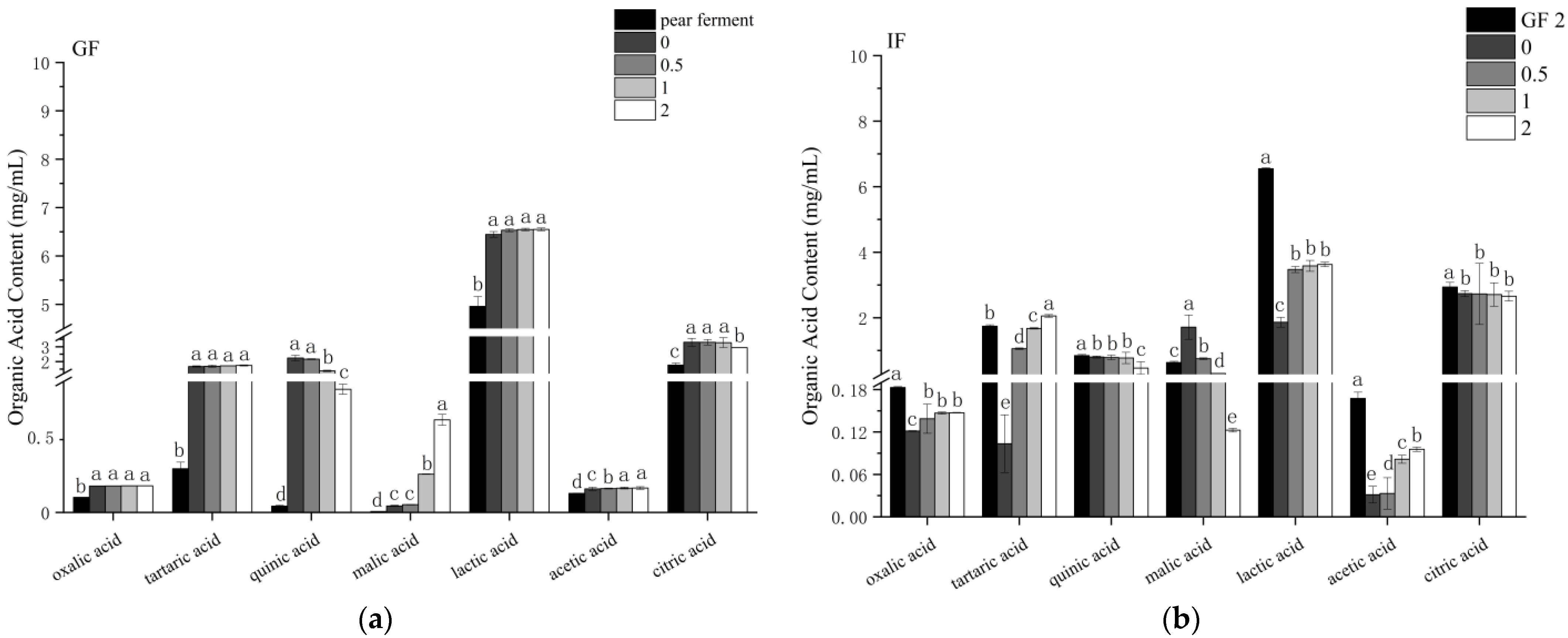
3.7. Effect of Simulated Digestion on Organic Acids In Vitro
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Dai, J.; Sha, R.; Wang, Z.; Cui, Y.; Fang, S.; Mao, J. Edible plant Jiaosu: Manufacturing, bioactive compounds, potential health benefits, and safety aspects. J. Sci. Food Agric. 2020, 100, 5313–5323. [Google Scholar] [CrossRef] [PubMed]
- Feng, Y.; Min, Z.; Arun, S.M.; Gao, Z. Recent research process of fermented plant extract: A review. Trends Food Sci. Technol. 2017, 65, 40–48. [Google Scholar] [CrossRef]
- He, Y.; Liu, G.; Xia, C.; Chen, J.; Zhao, J.; Li, X.; Deng, J.; Wang, X.; Xiang, Z.; Zeng, P. Laxative effect of mulberry ferment on two models of constipated mice. J. Funct. Foods 2022, 90, 104971. [Google Scholar] [CrossRef]
- Kuwaki, S.; Tanaka, H. Studies on the Plant-Based Extract Fermented by Lactic Acid Bacteria and Yeast. In Bulletin of Faculty of Health and Welfare Sciences; Okayama Prefectural University: Okayama, Japan, 2012. [Google Scholar]
- Lin, Y.; Lin, H.; Chen, Y.; Yang, D.; Li, C. Hepatoprotective effects of naturally fermented noni juice against thioacetamide-induced liver fibrosis in rats. J. Chin. Med. Assoc. 2017, 80, 212–221. [Google Scholar] [CrossRef] [PubMed]
- Chiu, H.; Chen, Y.; Lu, Y.; Han, Y.; Shen, Y.; Kamesh, V.; Wang, C. Regulatory efficacy of fermented plant extract on the intestinal microflora and lipid profile in mildly hypercholesterolemic individuals. J. Food Drug Anal. 2017, 25, 819–827. [Google Scholar] [CrossRef]
- Zulkawi, N.; Ng, K.; Zamberi, R.; Yeap, S.; Satharasinghe, D.; Jaganath, I.; Jamaluddin, A.; Tan, S.; Ho, W.; Alitheen, N.; et al. In vitro characterization and in vivo toxicity, antioxidant and immunomodulatory effect of fermented foods; Xeniji™. BMC Complement. Altern. Med. 2017, 17, 344. [Google Scholar] [CrossRef] [PubMed]
- Su, L.; Su, G. Effects of puerarin fermented liquid on reducing blood pressure of spontaneously hypertensive rats. J. Food Saf. Qual. 2016, 7, 3924–3928. [Google Scholar]
- Meng, F.; Chen, T.; Ma, D.; Wang, X.; Zhao, X.; Tian, P.; Wang, H.; Hai, Z.; Shen, L.; Tang, X.; et al. Reclamation of herb residues using probiotics and their therapeutic effect on diarrhea. Mediat. Inflamm. 2017, 2017, 4265898. [Google Scholar] [CrossRef]
- Marina, D.; Joan, Q.; Magdalena, R. Gamma-aminobutyric acid as a bioactive compound in foods: A review. J. Funct. Foods 2014, 10, 407–420. [Google Scholar] [CrossRef]
- Patricia, G.; Aranzazu, M.; Antonio, S.; Alberto, F. Phenolic-compound-extraction systems for fruit and vegetable samples. Molecules 2010, 15, 8813. [Google Scholar] [CrossRef]
- Beatriz, A.; Sergio, G.; María, B.S.; Luis, A.B.; Blanca, G.; Francisca, V.; Rosa, M.A. Polyphenolic contents in citrus fruit juices: Authenticity assessment. Eur. Food Res. Technol. Z. Lebensm. Unters. Forschung. A 2014, 238, 803–818. [Google Scholar] [CrossRef]
- Jiao, X.; Li, B.; Zhang, Q.; Gao, N.; Zhang, X.; Meng, X. Effect of in vitro-simulated gastrointestinal digestion on the stability and antioxidant activity of blueberry polyphenols and their cellular antioxidant activity towards HepG2 cells. Int. J. Food Sci. Technol. 2018, 53, 61–71. [Google Scholar] [CrossRef]
- Davide, T.; Elena, V.; Davide, B.; Angela, C. In vitro bio-accessibility and antioxidant activity of grape polyphenols. Food Chem. 2009, 120, 599–606. [Google Scholar] [CrossRef]
- Ding, Y.; Ma, Y.; Li, S.; Xi, X.; Sun, J.; Liu, Y.; Mou, J. Effect of inoculated fermentation on the bioaccessibility and antioxidant activity of polyphenols from grape jiaosu. Food Sci. 2022, 43, 108–116. [Google Scholar] [CrossRef]
- Mashitoa, F.M.; Manhivi, V.E.; Akinola, S.A.; Garcia, C.; Remize, F.; Shoko, T.; Sivakumar, D. Changes in phenolics and antioxidant capacity during fermentation and simulated in vitro digestion of mango puree fermented with different lactic acid bacteria. J. Food Process. Preserv. 2021, 45, e15937. [Google Scholar] [CrossRef]
- Valero-Cases, E.; Nuncio-Jáuregui, N.; Frutos, M.J. Influence of fermentation with different lactic acid bacteria and in vitro digestion on the biotransformation of phenolic compounds in fermented pomegranate juices. J. Agric. Food Chem. 2017, 65, 6488–6496. [Google Scholar] [CrossRef]
- Mashitoa, F.M.; Akinola, S.A.; Manhevi, V.E.; Garcia, C.; Remize, F.; Slabbert, R.M.; Sivakumar, D. Influence of fermentation of pasteurised papaya puree with different lactic acid bacterial strains on quality and bioaccessibility of phenolic compounds during in vitro digestion. Foods 2021, 10, 962. [Google Scholar] [CrossRef]
- Song, Y.; Lee, S.; Cho, J.; Moon, C.; Seo, D.; Jung, W. Expression and degradation patterns of chitinase purified from Xuehuali (Pyrus bretschneiderilia) pollen. Int. J. Biol. Macromol. 2018, 107, 446–452. [Google Scholar] [CrossRef]
- Sarkar, D.; Ankolekar, C.; Pinto, M.; Shetty, K. Dietary functional benefits of bartlett and starkrimson pears for potential management of hyperglycemia, hypertension and ulcer bacteria helicobacter pylori while supporting beneficial probiotic bacterial response. Food Res. Int. 2015, 69, 80–90. [Google Scholar] [CrossRef]
- Yan, Z.; Zhong, Y.; Duan, Y.; Chen, Q.; Li, F. Antioxidant mechanism of tea polyphenols and its impact on health benefits. Anim. Nutr. 2020, 6, 115–123. [Google Scholar] [CrossRef]
- Bae, J.Y.; Seo, Y.H.; Oh, S.W. Antibacterial activities of polyphenols against foodborne pathogens and their application as antibacterial agents. Food Sci. Biotechnol. 2022, 31, 985–997. [Google Scholar] [CrossRef] [PubMed]
- He, J.; Cheng, Y.; Guan, J.; Ge, W.; Zhao, Z. Changes of chlorogenic acid content and its synthesis-associated genes expression in Xuehua pear fruit during development. J. Integr. Agric. 2017, 16, 471–477. [Google Scholar] [CrossRef]
- QB/T 5323-2018; Plant Jiaosu. China Light Industry Press: Beijing, China, 2018; pp. 1–3.
- Gerardi, C.; Durante, M.; Tufariello, M.; Grieco, F.; Giovinazzo, G. Effects of time and temperature on stability of bioactive molecules, color and volatile compounds during storage of grape pomace flour. Appl. Sci. 2022, 12, 3956. [Google Scholar] [CrossRef]
- Chen, S.; Li, X.; Liu, X.; Wang, N.; An, Q.; Ye, X.; Zhao, Z.; Zhao, M.; Han, Y.; Ouyang, K.; et al. Investigation of chemical composition, antioxidant activity, and the effects of alfalfa flavonoids on growth performance. Oxidative Med. Cell. Longev. 2020, 2020, 8569237. [Google Scholar] [CrossRef]
- GB/T 23527-2009; Determination of Protease Activity. China Standard Publishing House: Beijing, China, 2009; pp. 6–8.
- GB/T 5009.171-2003; Determination of Superoxide Dismutase (SOD) Activity in Health Food. China Standard Press: Beijing, China, 2003; pp. 413–414.
- Romanet, R.; Sarhane, Z.; Bahut, F.; Uhl, J.; Schmitt-Kopplin, P.; Nikolantonaki, M.; Gougeon, R. Exploring the chemical space of white wine antioxidant capacity: A combined DPPH, EPR and FT-ICR-MS study. Food Chem. 2021, 355, 129566. [Google Scholar] [CrossRef]
- Zacarías-García, J.; Rey, F.; Gil, J.; Rodrigo, M.J.; Zacarías, L. Antioxidant capacity in fruit of citrus cultivars with marked differences in pulp coloration: Contribution of carotenoids and vitamin C. Food Sci. Technol. Int. 2020, 27, 1082013220944018. [Google Scholar] [CrossRef]
- Liyana-Pathirana, C.M.; Shahidi, F.; Alasalvar, C. Antioxidant activity of cherry laurel fruit (Laurocerasus officinalis Roem.) and its concentrated juice. Food Chem. 2005, 99, 121–128. [Google Scholar] [CrossRef]
- Su, D.; Luo, M.; Liu, H.; Qi, X.; Zeng, Q.; He, S.; Fen, S.; Zhang, J. The effect of simulated digestion on the composition of phenolic compounds and antioxidant activities in lychee pulp of different cultivars. Int. J. Food Sci. Technol. 2019, 54, 3042–3050. [Google Scholar] [CrossRef]
- Gutiérrez-Grijalva, Erick.; Zamudio-Sosa, Victor.; Contreras-Angulo, L.; Leyva-López, N.; Heredia, J. Bioaccessibility of phenolic compounds from mistletoe infusions and effect of in vitro digestion on its antioxidant and pancreatic lipase inhibitory activity. Foods 2022, 11, 3319. [Google Scholar] [CrossRef]
- Wojdyło, A.; Samoticha, J.; Chmielewska, J. The influence of different strains of oenococcus oeni malolactic bacteria on profile of organic acids and phenolic compounds of red wine cultivars rondo and regent growing in a cold region. J. Food Sci. 2020, 85, 1070–1081. [Google Scholar] [CrossRef]
- Kim, S.; Nam, D. Reliability, Accuracy, and Use Frequency of evaluation methods for amount of tongue coating. Chin. J. Integr. Med. 2019, 25, 378–385. [Google Scholar] [CrossRef] [PubMed]
- Sana, U.; Chang, G.H. Evaluation of Total Flavonoid, Total phenolic contents, and antioxidant activity of strychnobiflavone. Indones. J. Chem. 2020, 20, 716–721. [Google Scholar] [CrossRef]
- Yee, P.L.; Sook, F.P.; Mashitah, M.Y.; Jolius, G. Correlation between the total phenolic, total flavonoid and antioxidant content of phaleriamacrocarpa extract. Int. J. Recent Technol. Eng. (IJRTE) 2019, 8, 91–96. [Google Scholar] [CrossRef]
- Bouayed, J.; Hoffmann, L.; Bohn, T. Total phenolics, flavonoids, anthocyanins and antioxidant activity following simulated gastro-intestinal digestion and dialysis of apple varieties: Bioaccessibility and potential uptake. Food Chem. 2011, 128, 14–21. [Google Scholar] [CrossRef] [PubMed]
- Cong, Y.; Peng, M.; Liu, D.; Sun, H.; Tang, X. Changes in total polyphenols, total flavonoids and antioxidant activity of citrus during in vitro gastrointestinal digestion process. Food Sci. 2016, 37, 96–103. [Google Scholar] [CrossRef]
- Li, C.; Peng, Y.; Tang, W.; Li, T.; Gatasheh, M.; Rasheed, R.A.; Fu, J.; He, J.; Wang, W.; Shen, Y.; et al. Antioxidant, anti-lipidemic, hypoglycemic and antiproliferative effects of phenolics from Cortex Mori Radicis. Arab. J. Chem. 2022, 15, 103824. [Google Scholar] [CrossRef]
- Ogah, O.; Watkins, C.S.; Ubi, B.E.; Oraguzie, N.C. Phenolic compounds in Rosaceae fruit and nut crops. J. Agric. Food Chem. 2014, 62, 9369–9386. [Google Scholar] [CrossRef]
- Qin, W.; Ketnawa, S.; Ogawa, Y. Effect of digestive enzymes and pH on variation of bioavailability of green tea during simulated in vitro gastrointestinal digestion. Food Sci. Hum. Wellness 2022, 11, 669–675. [Google Scholar] [CrossRef]
- Wang, L.; Manson, J.E.; Gaziano, J.M.; Buring, J.E.; Sesso, H.D. Fruit and vegetable intake and the risk of hypertension in middle-aged and older women. Am. J. Hypertens. 2012, 25, 180–189. [Google Scholar] [CrossRef]
- Wang, Y.; Guo, Y.; Wang, Y.; Zhao, L.; Wu, X.; Liao, X. Analyses of phenolic composition and antioxidant activities of nfc apple juices from different cultivars. J. Chinses Inst. Food Sci. Technol. 2020, 20, 74–83. [Google Scholar] [CrossRef]
- Engin, C.; Mohammad, C.; Selin, A.; Ebru, T.A.; Erdem, Y. Effect of in vitro gastrointestinal digestion on the bioavailability of phenolic components and the antioxidant potentials of some Turkish fruit wines. Food Res. Int. 2015, 78, 209–215. [Google Scholar] [CrossRef]
- Chen, X.; Li, M.; Xu, Q.; Xiong, C.; Sun, Y. Changes in polyphenol contents and antioxidant activity in hawthorn (Crataegus pinnatifida Bunge) during simulated gastrointestinal digestion. Food Sci. 2019, 40, 31–37. [Google Scholar] [CrossRef]
- Ryan, L.; Prescott, S.L. Stability of the antioxidant capacity of twenty-five commercially available fruit juices subjected to an in vitro digestion. Int. J. Food Sci. Technol. 2010, 45, 1191–1197. [Google Scholar] [CrossRef]
- Peter, C.W.; Aisling, M.; Lisa, R. Stability of the total antioxidant capacity and total polyphenol content of 23 commercially available vegetable juices before and after in vitro digestion measured by FRAP, DPPH, ABTS and Folin–Ciocalteu methods. Food Res. Int. 2011, 44, 217–224. [Google Scholar] [CrossRef]
- Jara-Palacios, M.J.; Gonçalves, S.; Hernanz, D.; Heredia, F.J.; Romano, A. Effects of in vitro gastrointestinal digestion on phenolic compounds and antioxidant activity of different white winemaking byproducts extracts. Food Res. Int. 2018, 109, 433–439. [Google Scholar] [CrossRef] [PubMed]
- Dou, Z.; Chen, C.; Fu, X.; Liu, R. A dynamic view on the chemical composition and bioactive properties of mulberry fruit using an in vitro digestion and fermentation model. Food Funct. 2022, 13, 4142–4157. [Google Scholar] [CrossRef] [PubMed]
- Tenore, G.C.; Campiglia, P.; Giannetti, D.; Novellino, E. Simulated gastrointestinal digestion, intestinal permeation and plasma protein interaction of white, green, and black tea polyphenols. Food Chem. 2015, 169, 320–326. [Google Scholar] [CrossRef]
- Zhai, F.; Wang, Q.; Han, J. Nutritional components and antioxidant properties of seven kinds of cereals fermented by the basidiomycete Agaricus blazei. J. Cereal Sci. 2015, 65, 202–208. [Google Scholar] [CrossRef]
- Huang, H.; Korivi, M.; Tsai, C.; Yang, J.; Tsai, Y. Supplementation of Lactobacillus plantarum K68 and fruit-vegetable ferment along with high fat-fructose diet attenuates metabolic syndrome in rats with insulin resistance. Evid. -Based Complement. Altern. Med. eCAM 2013, 2013, 943020. [Google Scholar] [CrossRef]
- Marikovsky, M.; Ziv, V.; Nevo, N.; Harris-Cerruti, C.; Mahler, O. Cu/Zn superoxide dismutase plays important role in immune response. J. Immunol. 2003, 170, 2993–3001. [Google Scholar] [CrossRef]
- Ismy, J.; Sugandi, S.; Rachmadi, D.; Hardjowijoto, S.; Mustafa, A. The effect of exogenous Superoxide Dismutase (SOD) on caspase-3 activation and apoptosis induction in Pc-3 prostate cancer cells. Res. Rep. Urol. 2020, 12, 503–508. [Google Scholar] [CrossRef] [PubMed]
- Sun, S. The Research of Technology and Functional Active Ingredients for Litchi Juice Mixed Strains Fermentation. Master’s Thesis, South China Agricultural University, Guangzhou, China, 2016. [Google Scholar]
- Zhou, L.; Xu, Q.; Zhang, Y.; Zhou, Z.; Guan, W.; Li, Y. Purification, Characterization and in vitro anthelmintic activity of a neutral metalloprotease from laccocephalum mylittae. Chin. J. Chem. Eng. 2010, 18, 122–128. [Google Scholar] [CrossRef]
- Wu, X.; Wang, W.; Zhu, T.; Liang, T.; Lu, F.; He, W.; Zhang, H.; Liu, Z.; He, S.; Gao, K.; et al. Phenylpropanoid glycoside inhibition of pepsin, trypsin and α-chymotrypsin enzyme activity in Kudingcha leaves from Ligustrum purpurascens. Food Res. Int. 2013, 54, 1376–1382. [Google Scholar] [CrossRef]
- Schrijver, R.; Ollevier, F. Protein digestion in juvenile turbot (Scophthalmus maximus) and effects of dietary administration of Vibrio proteolyticus. Aquaculture 2000, 186, 107–116. [Google Scholar] [CrossRef]
- Zeng, L.; Li, C.; Peng, H.; Yu, Y.; Zhou, F.; Fu, G. Stability of SOD in yogurt to simulated gastrointestinal digestive fluid. Food Sci. 2014, 35, 274–278. [Google Scholar] [CrossRef]
- Liao, X.; Guo, L.; Ye, Z.; Qiu, L.; Gu, F.; Lin, J. Use of autochthonous lactic acid bacteria starters to ferment mango juice for promoting its probiotic roles. Prep. Biochem. Biotechnol. 2016, 46, 399–405. [Google Scholar] [CrossRef]
- Anitha, S.; Krishnan, S.; Senthilkumar, K.; Sasirekha, V. A comparative investigation on the scavenging of 2,2-diphenyl-1-picrylhydrazyl radical by the natural antioxidants (+) catechin and (-) epicatechin. J. Mol. Struct. 2021, 1242, 130805. [Google Scholar] [CrossRef]
- Xia, L.; Fang, L.; Pao, L.; Yin, S.; Wanru, G. Investigation of the interaction for three citrus flavonoids and α-amylase by surface plasmon resonance. Food Res. Int. 2017, 97, 1–6. [Google Scholar] [CrossRef]
- Su, D.; Liu, H.; Zeng, Q.; Qi, X.; Yao, X.; Zhang, J. Changes in the phenolic contents and antioxidant activities of citrus peels from different cultivars after in vitro digestion. Int. J. Food Sci. Technol. 2017, 52, 2471–2478. [Google Scholar] [CrossRef]
- Spinella, M. Phenolics from walnut (Juglans regia L.) kernels: Antioxidant activity and interactions with proteins. Food Chem. 2007, 107, 607–612. [Google Scholar] [CrossRef]
- Zhang, Q.; Rui, X.; Li, W.; Chen, X.; Jiang, M.; Dong, M. Anti-swarming and -biofilm activities of rose phenolic extract during simulated in vitro gastrointestinal digestion. Food Control 2016, 64, 189–195. [Google Scholar] [CrossRef]
- Anna, R.C.B.; Leonardo, M.S.M.; Paula, L.G.M.; Sascha, H.; Veridiana, V.R. Lactobacillus fermentation of jussara pulp leads to the enzymatic conversion of anthocyanins increasing antioxidant activity. J. Food Compos. Anal. 2018, 69, 162–170. [Google Scholar] [CrossRef]
- Song, H.K.; Ahn, J.K.; Kim, K.H.; Lee, S.J.; Baek, J.Y.; Chung, I.M. Assessment of antioxidative capacity in relation to seed trails of rice varieties. Korean J. Crop Sci. 2006, 51, 544–553. [Google Scholar]
- Pavan, V.; Renata, A.S.S.; Glaucia, M.P. The effect of in vitro digestion on the antioxidant activity of fruit extracts (Carica papaya, Artocarpus heterophillus and Annona marcgravii). LWT-Food Sci. Technol. 2014, 59, 1247–1251. [Google Scholar] [CrossRef]
- Huang, S.; Ma, Y.; Zhang, C.; Cai, S.; Pang, M. Bioaccessibility and antioxidant activity of phenolics in native and fermented Prinsepia utilis Royle seed during a simulated gastrointestinal digestion in vitro. J. Funct. Foods 2017, 37, 354–362. [Google Scholar] [CrossRef]
- Wang, G.; Chen, C.; He, G.; Liu, S.; Lv, Y. Antioxidant capacity of mango fermented beverage in simulated digestion in vitro. Food Sci. Technol. 2021, 46, 110–115. [Google Scholar] [CrossRef]
- Tsao, R.; Yang, R.; Xie, S.; Sockovie, E.; Khanizadeh, S. Which polyphenolic compounds contribute to the total antioxidant activities of apple? J. Agric. Food Chem. 2005, 53, 4989–4995. [Google Scholar] [CrossRef]
- Mao, X.; Zhang, Y.; Liu, Y. Study on extraction technology and antioxidant activity of proanthocyanins from grape branches and tendrils. Storage Process 2018, 18, 82–89. [Google Scholar] [CrossRef]
- Xie, Y.; Kosińska, A.; Xu, H.; Andlauer, W. Milk enhances intestinal absorption of green tea catechins in in vitro digestion/Caco-2 cells model. Food Res. Int. 2013, 53, 793–800. [Google Scholar] [CrossRef]
- Jiang, J.; Zhang, Z.; Zhao, J.; Liu, Y. The effect of non-covalent interaction of chlorogenic acid with whey protein and casein on physicochemical and radical-scavenging activity of in vitro protein digests. Food Chem. 2018, 268, 334–341. [Google Scholar] [CrossRef]
- Lutfun, N.; Afaf, A.; Anil, K.; Sarker, S.D. Arbutin: Occurrence in plants, and its potential as an anticancer agent. Molecules 2022, 27, 8786. [Google Scholar] [CrossRef]
- Moghadam, D.; Zarei, R.; Vakili, S.; Ghojoghi, R.; Zarezade, V.; Veisi, A.; Sabaghan, M.; Azadbakht, O.; Behrouj, H. The effect of natural polyphenols resveratrol, gallic acid, and kuromanin chloride on human telomerase reverse transcriptase (hTERT) expression in HepG2 hepatocellular carcinoma: Role of SIRT1/Nrf2 signaling pathway and oxidative stress. Mol. Biol. Rep. 2022, 50, 77–84. [Google Scholar] [CrossRef] [PubMed]
- Weize, R.; Liu, Z.; Li, X.; Shen, J.; Liu, Y.; Li, Y. Total coumarin content and antioxidant activity in different parts of obazema, a yi medicine. Med. Plant 2019, 10, 39–44. [Google Scholar] [CrossRef]
- Yuan, M.; Cui, X.; Ye, F.; Zhao, G. Effects of steam explosion pretreatment on polyphenol composition, surface structure and antioxidant activity of apple pomace. Food Ferment. Ind. 2022, 48, 179–185. [Google Scholar] [CrossRef]
- Kahle, K.; Kempf, M.; Schreier, P.; Scheppach, W.; Schrenk, D.; Kautenburger, T.; Hecker, D.; Huemmer, W.; Ackermann, M.; Richling, E. Intestinal transit and systemic metabolism of apple polyphenols. Eur. J. Nutr. 2011, 50, 507–522. [Google Scholar] [CrossRef]
- Sabboh-Jourdan, H.; Valla, F.; Epriliati, I.; Gidley, M.J. Organic acid bioavailability from banana and sweet potato using an in vitro digestion and Caco-2 cell model. Eur. J. Nutr. 2011, 50, 31–40. [Google Scholar] [CrossRef]










Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, X.; Li, Y.; Li, Y.; Zhao, J.; Cheng, Y.; Wang, Y.; Guan, J. Changes of Bioactive Components and Antioxidant Capacity of Pear Ferment in Simulated Gastrointestinal Digestion In Vitro. Foods 2023, 12, 1211. https://doi.org/10.3390/foods12061211
Zhang X, Li Y, Li Y, Zhao J, Cheng Y, Wang Y, Guan J. Changes of Bioactive Components and Antioxidant Capacity of Pear Ferment in Simulated Gastrointestinal Digestion In Vitro. Foods. 2023; 12(6):1211. https://doi.org/10.3390/foods12061211
Chicago/Turabian StyleZhang, Xiaoying, Yiming Li, Yue Li, Jiangli Zhao, Yudou Cheng, Yongxia Wang, and Junfeng Guan. 2023. "Changes of Bioactive Components and Antioxidant Capacity of Pear Ferment in Simulated Gastrointestinal Digestion In Vitro" Foods 12, no. 6: 1211. https://doi.org/10.3390/foods12061211
APA StyleZhang, X., Li, Y., Li, Y., Zhao, J., Cheng, Y., Wang, Y., & Guan, J. (2023). Changes of Bioactive Components and Antioxidant Capacity of Pear Ferment in Simulated Gastrointestinal Digestion In Vitro. Foods, 12(6), 1211. https://doi.org/10.3390/foods12061211