
Acetic Acid Immersion Alleviates the Softening of Cooked Sagittaria sagittifolia L. Slices by Affecting Cell Wall Polysaccharides

Abstract

1. Introduction
2. Materials and Methods
2.1. Materials and Sample Preparation
2.2. Texture Measurement
2.3. Scanning Electron Microscopy (SEM)
2.4. Immunolabeling of Pectic Epitopes
2.5. Separation of Cell Wall Fractions
2.6. Fourier Transform Infrared (FT-IR) Spectroscopy of the AIR
2.7. Monosaccharide Composition Measurement in Fractions
2.8. Molecular Mass Distribution of Cell Wall Fractions
2.9. Thermogravimetry Analysis
2.10. Statistical Analysis
3. Results and Discussion
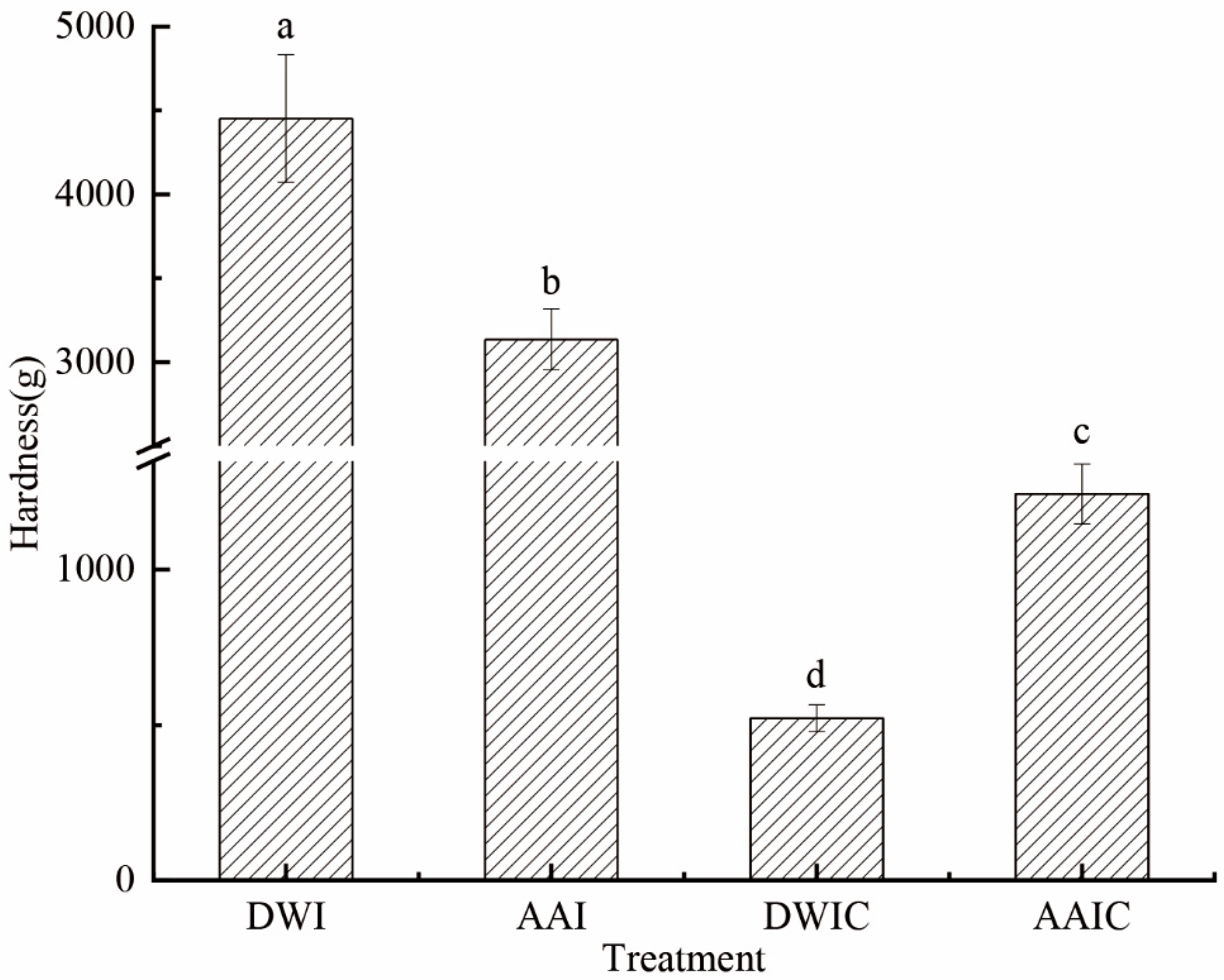
3.1. Effects of Acetic Acid Pretreatment on the Hardness of Fresh and Cooked Arrowhead Bulb Slices
3.2. Effects of Different Treatments on Cell Wall Microstructure of Arrowhead Bulb Slices
3.3. In Situ Immunolabeling of Pectic Epitopes
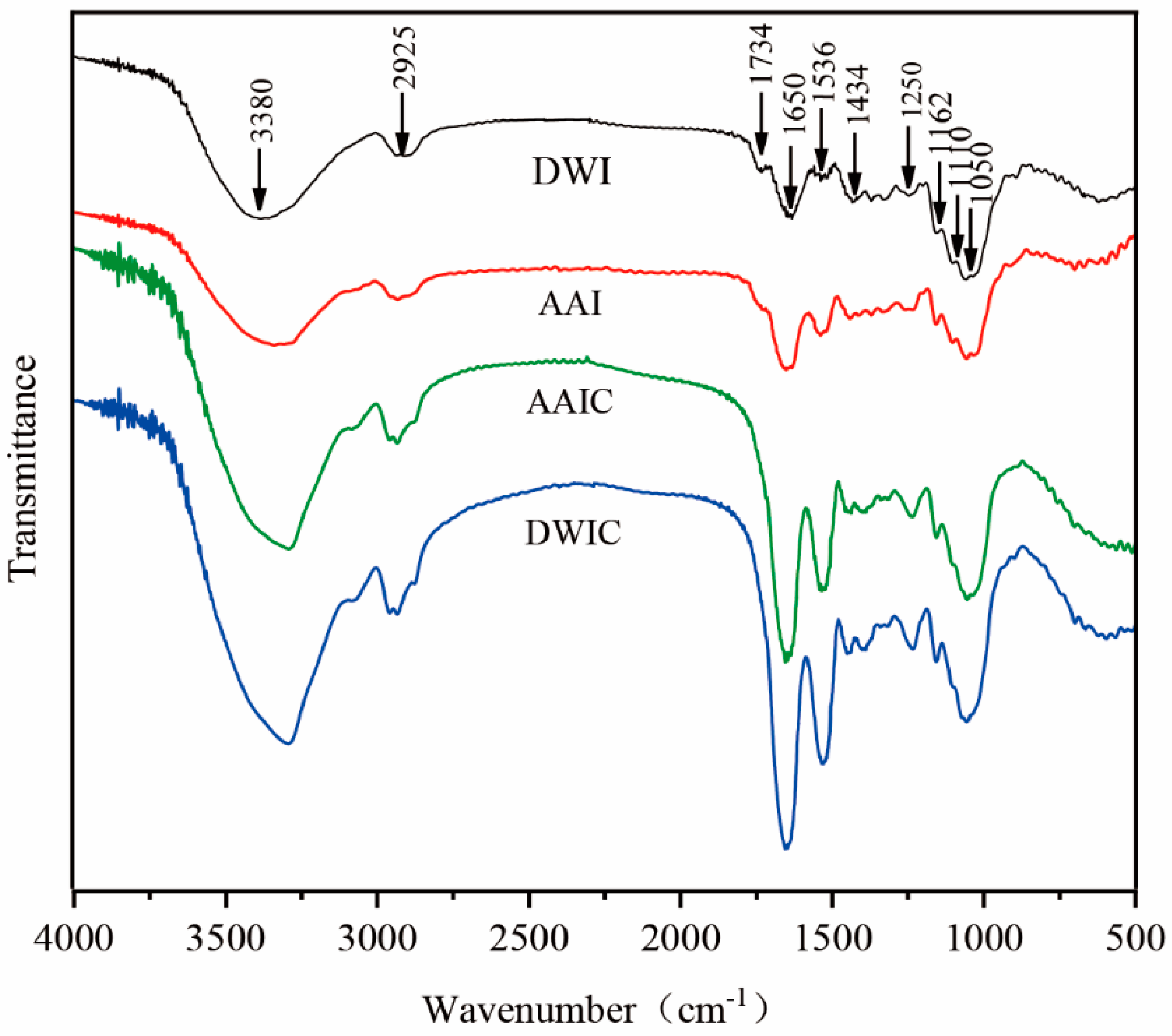
3.4. FT-IR
3.5. Composition and Sugar Ratio of Different Cell Wall Fractions
3.6. Molecular Weight Distribution
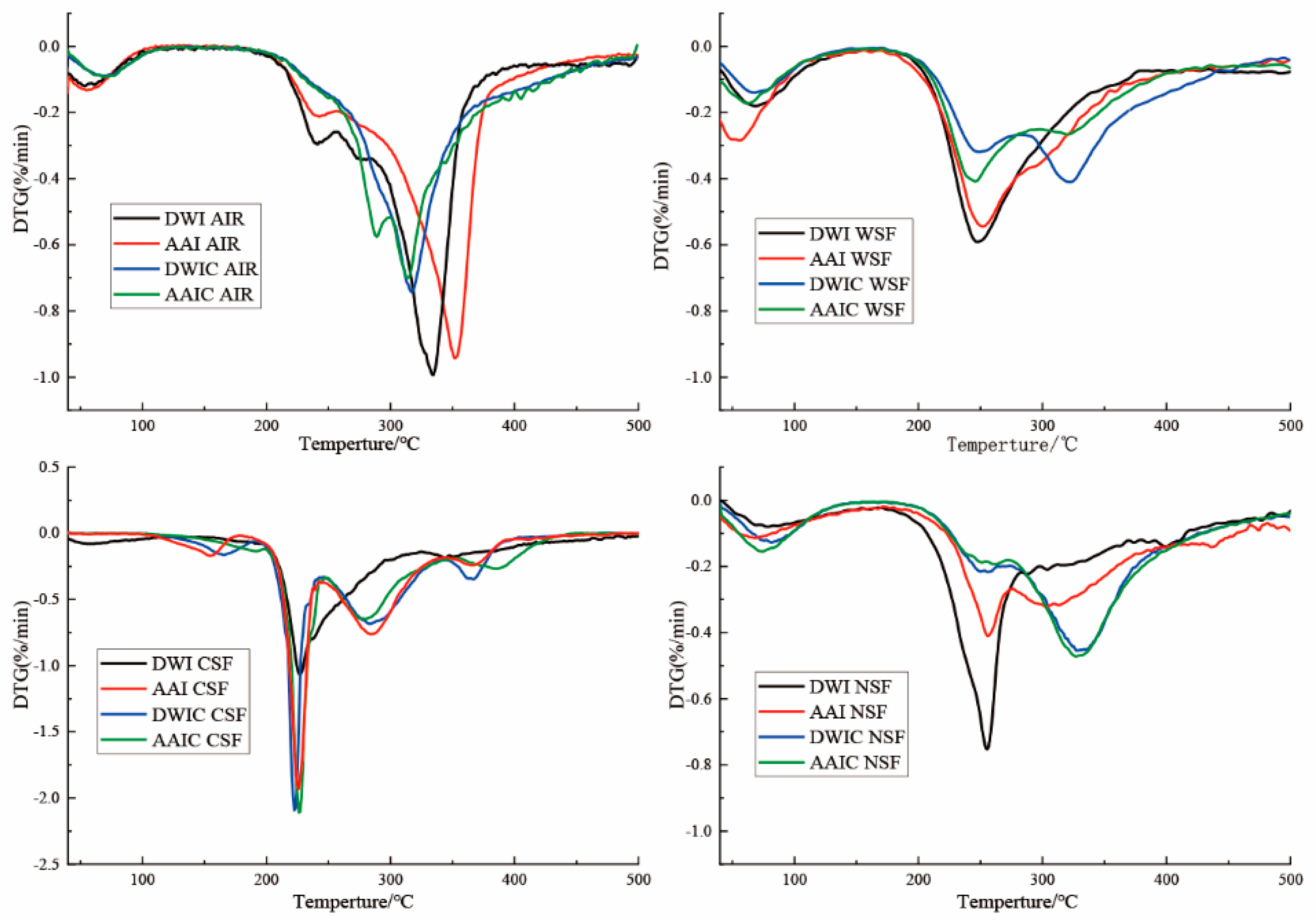
3.7. DTG Analysis
4. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
Abbreviations
| AAI | acetic acid immersion for 15 h |
| AAIC | cooked AAI |
| AIR | alcohol insoluble residue |
| Ara | arabinose |
| CLSM | confocal laser scanning microscopy |
| CSF | chelate-soluble fraction |
| DTG | derivative thermogravimetry |
| DWI | distilled water immersion for 15 h |
| DWIC | cooked DWI |
| FT-IR | Fourier transform infrared |
| Fuc | fucose |
| GalA | galacturonic acid |
| Gal | galactose |
| Glc | glucose |
| GPC | gel permeation chromatography |
| HF | hemicellulose fraction |
| HG | homogalacturonan |
| HPLC | high-performance liquid chromatography |
| MAbs | monoclonal antibodies |
| Man | mannose |
| NSF | sodium carbonate-soluble fraction |
| PMP | 1-phenyl-3-methyl-5-pyrazolone |
| RG-I | rhamnogalacturonan I |
| RG-II | rhamnogalacturonan II |
| Rha | rhamnose |
| SEM | scanning electron microscopy |
| TG | thermogravimetry |
| WSF | water-soluble fraction |
| Xyl | xylose |
References
- Feng, Y.; Juliet, I.C.; Qin, W.; Duan, Y.; Gao, X.; Liu, Q.; Zhang, H.; Ma, H. Rheology, In Vitro Digestion and Functional Properties of Sagittaria sagittifolia L. Resistant Starch as Affected by Different Preparation Methods. Starch-Stärke 2022, 74, 2100181. [Google Scholar] [CrossRef]
- Raza, H.; Ameer, K.; Ma, H.; Liang, Q.; Ren, X. Structural and physicochemical characterization of modified starch from arrowhead tuber (Sagittaria sagittifolia L.) using tri-frequency power ultrasound. Ultrason. Sonochemistry 2021, 80, 105826. [Google Scholar] [CrossRef] [PubMed]
- Paniagua, C.; Santiago-Doménech, N.; Kirby, A.R.; Gunning, A.P.; Morris, V.J.; Quesada, M.A.; Matas, A.J.; Mercado, J.A. Structural changes in cell wall pectins during strawberry fruit development. Plant Physiol. Biochem. 2017, 118, 55–63. [Google Scholar] [CrossRef]
- Frempong, K.E.B.; Chen, Y.; Wang, Z.; Xu, J.; Xu, X.; Cui, W.; Gong, H.; Peng, D.; Liang, L.; Meng, Y.; et al. Study on textural changes and pectin degradation of tarocco blood Orange during storage. Int. J. Food Prop. 2022, 25, 344–358. [Google Scholar] [CrossRef]
- Liu, X.; Renard, C.M.G.C.; Rolland-Sabaté, A.; Bureau, S.; Le Bourvellec, C. Modification of apple, beet and kiwifruit cell walls by boiling in acid conditions: Common and specific responses. Food Hydrocoll. 2021, 112, 106266. [Google Scholar] [CrossRef]
- Zhao, W.; Shehzad, H.; Yan, S.; Li, J.; Wang, Q. Acetic acid pretreatment improves the hardness of cooked potato slices. Food Chem. 2017, 228, 204–210. [Google Scholar] [CrossRef]
- Liu, G.; Liu, Y.; Yan, S.; Li, J. Acetic acid reducing the softening of lotus rhizome during heating by regulating the chelate-soluble polysaccharides. Carbohydr. Polym. 2020, 240, 116209. [Google Scholar] [CrossRef]
- Gong, W.; Shi, B.; Zeng, F.-K.; Dong, N.; Lei, Z.; Liu, J. Evaluation of cooking, nutritional, and quality characteristics of fresh-cut potato slice pretreated with acetic acid. J. Food Sci. 2021, 87, 427–437. [Google Scholar] [CrossRef]
- Moens, L.G.; Huang, W.; Van Loey, A.M.; Hendrickx, M.E.G. Effect of pulsed electric field and mild thermal processing on texture-related pectin properties to better understand carrot (Daucus carota) texture changes during subsequent cooking. Innov. Food Sci. Emerg. Technol. 2021, 70, 102700. [Google Scholar] [CrossRef]
- Houben, K.; Jolie, R.P.; Fraeye, I.; Van Loey, A.M.; Hendrickx, M.E. Comparative study of the cell wall composition of broccoli, carrot, and tomato: Structural characterization of the extractable pectins and hemicelluloses. Carbohydr. Res. 2011, 346, 1105–1111. [Google Scholar] [CrossRef]
- Ormerod, A.; Ralfs, J.; Jobling, S.; Gidley, M. The influence of starch swelling on the material properties of cooked potatoes. J. Mater. Sci. 2002, 37, 1667–1673. [Google Scholar] [CrossRef]
- Yagn, Z.; Duan, X.; Yang, J.; Wang, H.; Liu, F.; Xu, X.; Pan, S. Effects of high hydrostatic pressure and thermal treatment on texture properties of pickled kohlrabi. LWT—Food Sci. Technol. 2022, 157, 113078. [Google Scholar]
- Liu, Y.; Liu, J.; Liu, G.; Duan, R.; Sun, Y.; Li, J.; Yan, S.; Li, B. Sodium bicarbonate reduces the cooked hardness of lotus rhizome via side chain rearrangement and pectin degradation. Food Chem. 2021, 370, 130962. [Google Scholar] [CrossRef] [PubMed]
- Andersen, M.C.F.; Boos, I.; Marcus, S.E.; Kračun, S.K.; Rydahl, M.G.; Willats, W.G.T.; Knox, J.P.; Clausen, M.H. Characterization of the LM5 pectic galactan epitope with synthetic analogues of β-1,4-d-galactotetraose. Carbohydr. Res. 2016, 436, 36–40. [Google Scholar] [CrossRef]
- Ngolong Ngea, G.L.; Guillon, F.; Essia Ngang, J.J.; Bonnin, E.; Bouchet, B.; Saulnier, L. Modification of cell wall polysaccharides during retting of cassava roots. Food Chem. 2016, 213, 402–409. [Google Scholar] [CrossRef]
- Cornuault, V.; Buffetto, F.; Marcus, S.E.; Crépeau, M.-J.; Guillon, F.; Ralet, M.-C.; Knox, J.P. LM6-M: A high avidity rat monoclonal antibody to pectic α-1,5-L-arabinan. BioRxiv 2017. [Google Scholar] [CrossRef]
- Christiaens, S.; Van Buggenhout, S.; Vandevenne, E.; Jolie, R.; Van Loey, A.M.; Hendrickx, M.E. Towards a better understanding of the pectin structure–function relationship in broccoli during processing: Part II—Analyses with anti-pectin antibodies. Food Res. Int. 2011, 44, 2896–2906. [Google Scholar] [CrossRef]
- Kyomugasho, C.; Willemsen, K.L.D.D.; Christiaens, S.; Van Loey, A.M.; Hendrickx, M.E. Microscopic evidence for Ca2+ mediated pectin–pectin interactions in carrot-based suspensions. Food Chem. 2015, 188, 126–136. [Google Scholar] [CrossRef]
- Cornuault, V.; Posé, S.; Knox, J.P. Disentangling pectic homogalacturonan and rhamnogalacturonan-I polysaccharides: Evidence for sub-populations in fruit parenchyma systems. Food Chem. 2017, 246, 275–285. [Google Scholar] [CrossRef]
- Posé, S.; Marcus, S.E.; Knox, J.P. Differential metabolism of pectic galactan in tomato and strawberry fruit: Detection of the LM26 branched galactan epitope in ripe strawberry fruit. Physiol. Plant 2018, 164, 95–105. [Google Scholar] [CrossRef]
- Liu, J.; Wen, C.; Wang, M.; Wang, S.; Dong, N.; Lei, Z.; Lin, S.; Zhu, B. Enhancing the hardness of potato slices after boiling by combined treatment with lactic acid and calcium chloride: Mechanism and optimization. Food Chem. 2019, 308, 124832. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Chen, M.; Wen, C.; Zhou, J.; Gu, J.; Duan, Y.; Zhang, H.; Ren, X.; Ma, H. Structural characterization and immunostimulatory activity of a novel polysaccharide isolated with subcritical water from Sagittaria sagittifolia L. Int. J. Biol. Macromol. 2019, 133, 11–20. [Google Scholar] [CrossRef] [PubMed]
- Peng, J.; Bi, J.; Yi, J.; Wu, X.; Zhou, M.; Zhao, Y.; Liu, J.n. Characteristics of cell wall pectic polysaccharides affect textural properties of instant controlled pressure drop dried carrot chips derived from different tissue zone. Food Chem. 2019, 293, 358–367. [Google Scholar] [CrossRef]
- Zhang, J.; Wang, Z.-W. Thermal analysis and physiological behavior of cellulose/pectin complex from Canna edulis Ker by-product. Carbohydr. Polym. 2012, 87, 1153–1158. [Google Scholar] [CrossRef]
- Sun, Y.; Kang, X.; Chen, F.; Liao, X.; Hu, X. Mechanisms of carrot texture alteration induced by pure effect of high pressure processing. Innov. Food Sci. Emerg. Technol. 2018, 54, 260–269. [Google Scholar] [CrossRef]
- Greve, L.C.; Shackel, K.A.; Ahmadi, H.; McArdle, R.N.; Gohlke, J.R.; Labavitch, J.M. Impact of heating on carrot firmness: Contribution of cellular turgor. J. Agric. Food Chem. 1994, 42, 2896–2899. [Google Scholar] [CrossRef]
- Bordoloi, A.; Kaur, L.; Singh, J. Parenchyma cell microstructure and textural characteristics of raw and cooked potatoes. Food Chem. 2011, 133, 1092–1100. [Google Scholar] [CrossRef]
- Redgwell, R.J.; Fischer, M.; Kendal, E.; MacRae, E.A. Galactose loss and fruit ripening: High-molecular-weight arabinogalactans in the pectic polysaccharides of fruit cell walls. Planta 1997, 203, 174–181. [Google Scholar] [CrossRef]
- Ribas-Agustí, A.; Van Buggenhout, S.; Palmero, P.; Hendrickx, M.; Van Loey, A. Investigating the role of pectin in carrot cell wall changes during thermal processing: A microscopic approach. Innov. Food Sci. Emerg. Technol. 2013, 24, 113–120. [Google Scholar] [CrossRef]
- Nyman, E.M.G.L.; Svanberg, S.J.M. Modification of physicochemical properties of dietary fibre in carrots by mono- and divalent cations. Food Chem. 2002, 76, 273–280. [Google Scholar] [CrossRef]
- Imaizumi, T.; Szymańska-Chargot, M.; Pieczywek, P.M.; Chylińska, M.; Kozioł, A.; Ganczarenko, D.; Tanaka, F.; Uchino, T.; Zdunek, A. Evaluation of pectin nanostructure by atomic force microscopy in blanched carrot. LWT—Food Sci. Technol. 2017, 84, 658–667. [Google Scholar] [CrossRef]
- Gu, J.; Zhang, H.; Zhang, J.; Wen, C.; Ma, H.; Duan, Y.; He, Y. Preparation, characterization and bioactivity of polysaccharide fractions from Sagittaria sagittifolia L. Carbohydr. Polym. 2019, 229, 115355. [Google Scholar] [CrossRef] [PubMed]
- Reddy, J.P.; Rhim, J.-W. Extraction and Characterization of Cellulose Microfibers from Agricultural Wastes of Onion and Garlic. J. Nat. Fibers 2018, 15, 465–473. [Google Scholar] [CrossRef]
- Wang, W.; Ma, X.; Jiang, P.; Hu, L.; Zhi, Z.; Chen, J.; Ding, T.; Ye, X.; Liu, D. Characterization of pectin from grapefruit peel: A comparison of ultrasound-assisted and conventional heating extractions. Food Hydrocoll. 2016, 61, 730–739. [Google Scholar] [CrossRef]
- Lee, K.J.D.; Cornuault, V.; Manfield, I.W.; Ralet, M.-C.; Knox, J.P. Multi-scale spatial heterogeneity of pectic rhamnogalacturonan I (RG-I) structural features in tobacco seed endosperm cell walls. Plant J. 2013, 75, 1018–1027. [Google Scholar] [CrossRef]
- Basanta, M.F.; de Escalada Plá, M.F.; Stortz, C.A.; Rojas, A.M. Chemical and functional properties of cell wall polymers from two cherry varieties at two developmental stages. Carbohydr. Polym. 2012, 92, 830–841. [Google Scholar] [CrossRef]
- Broxterman, S.E.; Schols, H.A. Interactions between pectin and cellulose in primary plant cell walls. Carbohydr. Polym. 2018, 192, 263–272. [Google Scholar] [CrossRef]
- Lu, R.; Ma, Y.; Wang, X.; Zhao, X.; Liang, H.; Wang, D. Study of texture properties of ‘laba’ garlic in different color states and their change mechanisms. Int. J. Food Sci. Technol. 2021, 56, 4710–4721. [Google Scholar] [CrossRef]
- Wu, D.; Chen, S.; Ye, X.; Zheng, X.; Ahmadi, S.; Hu, W.; Yu, C.; Cheng, H.; Linhardt, R.J.; Chen, J. Enzyme-extracted raspberry pectin exhibits a high-branched structure and enhanced anti-inflammatory properties than hot acid-extracted pectin. Food Chem. 2022, 383, 132387. [Google Scholar] [CrossRef]
- Li, Q.; Yao, S.; Deng, L.; Zeng, K. Changes in biochemical properties and pectin nanostructures of juice sacs during the granulation process of pomelo fruit (Citrus grandis). Food Chem. 2021, 376, 131876. [Google Scholar] [CrossRef]
- Einhorn-Stoll, U.; Kunzek, H.; Dongowski, G. Thermal analysis of chemically and mechanically modified pectins. Food Hydrocoll. 2007, 21, 1101–1112. [Google Scholar] [CrossRef]
- Bryś, A.; Bryś, J.; Ostrowska-Ligęza, E.; Kaleta, A.; Górnicki, K.; Głowacki, S.; Koczoń, P. Wood biomass characterization by DSC or FT-IR spectroscopy. J. Therm. Anal. Calorim. 2016, 126, 27–35. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment | Sugar Content (mg/g) | Sugar Ratios | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Man | Rha | GalA | Glc | Gal | Xyl | Ara | Fuc | 1 | 2 | 3 | ||
| WSF | DWI | 0.88 ± 0.02 b | 8.81 ± 0.31 c | 75.05 ± 0.42 a | 3.22 ± 0.04 b | 17.41 ± 0.22 c | 2.99 ± 0.04 b | 17.30 ± 0.13 c | 0.84 ± 0.06 a | 1.583 ± 0.015 a | 0.117 ± 0.004 d | 3.941 ± 0.114 b |
| AAI | 0.50 ± 0.14 c | 11.90 ± 0.07 b | 72.07 ± 0.09 b | 3.52 ± 0.07 a | 22.97 ± 0.16 b | 4.79 ± 0.07 a | 22.50 ± 0.26 b | 0.80 ± 0.05 a | 1.143 ± 0.006 b | 0.165 ± 0.001 c | 3.819 ± 0.018 b | |
| DWIC | 1.02 ± 0.03 b | 6.42 ± 0.05 d | 21.91 ± 0.23 c | 1.47 ± 0.07 c | 13.15 ± 0.12 d | 0.74 ± 0.02 c | 11.72 ± 0.10 d | 0.76 ± 0.08 a | 0.667 ± 0.006 c | 0.293 ± 0.005 b | 3.876 ± 0.057 b | |
| AAIC | 4.61 ± 0.29 a | 15.28 ± 0.55 a | 17.74 ± 0.01 d | 3.12 ± 0.07 b | 32.65 ± 0.06 a | — | 37.32 ± 0.06 a | 0.35 ± 0.01 b | 0.210 ± 0.000 d | 0.863 ± 0.032 a | 4.580 ± 0.156 a | |
| CSF | DWI | 0.08 ± 0.01 c | 8.37 ± 0.03 a | 45.12 ± 0.40 a | 2.40 ± 0.02 b | 16.99 ± 0.33 a | 2.90 ± 0.21 a | 18.06 ± 0.05 a | 0.36 ± 0.31 | 0.967 ± 0.021 c | 0.186 ± 0.002 a | 4.184 ± 0.033 c |
| AAI | 0.20 ± 0.01 b | 0.11 ± 0.01 c | 3.91 ± 0.06 b | 3.86 ± 0.10 a | 0.11 ± 0.02 c | 0.23 ± 0.01 b | 0.39 ± 0.03 c | — | 4.637 ± 0.140 a | 0.029 ± 0.001 c | 4.378 ± 0.120 bc | |
| DWIC | 0.21 ± 0.01 a | 0.05 ± 0.01 d | 0.41 ± 0.11 d | 0.51 ± 0.14 d | — | 0.24 ± 0.01 b | 0.94 ± 0.01 b | — | 0.340 ± 0.092 d | 0.131 ± 0.030 b | 18.058 ± 1.060 a | |
| AAIC | — | 0.16 ± 0.01 b | 2.55 ± 0.07 c | 1.08 ± 0.14 c | 0.47 ± 0.06 b | 0.13 ± 0.06 b | 0.39 ± 0.08 c | — | 2.230 ± 0.202 b | 0.060 ± 0.001 d | 5.453 ± 0.593 b | |
| NSF | DWI | 0.94 ± 0.04 c | 6.97 ± 0.02 a | 29.44 ± 0.29 a | 1.04 ± 0.02 a | 18.13 ± 0.09 a | 1.81 ± 0.07 a | 20.42 ± 0.15 a | — | 0.620 ± 0.010 b | 0.237 ± 0.002 c | 5.536 ± 0.037 b |
| AAI | 0.73 ± 0.31 c | 3.06 ± 0.02 c | 15.26 ± 0.14 b | 0.63 ± 0.05 b | 7.67 ± 0.06 b | 0.92 ± 0.05 b | 10.25 ± 0.18 b | — | 0.697 ± 0.015 a | 0.200 ± 0.003 c | 5.860 ± 0.078 a | |
| DWIC | 4.19 ± 0.23 a | 3.20 ± 0.06 c | 1.80 ± 0.22 d | 0.98 ± 0.02 a | 2.25 ± 0.01 d | — | 2.36 ± 0.10 d | 1.05 ± 0.02 | 0.290 ± 0.070 c | 1.797 ± 0.193 a | 1.440 ± 0.024 d | |
| AAIC | 2.34 ± 0.14 b | 3.79 ± 0.27 b | 4.40 ± 0.06 c | 0.92 ± 0.17 a | 3.67 ± 0.54 c | 0.47 ± 0.02 c | 6.92 ± 0.40 c | — | 0.297 ± 0.021 c | 0.860 ± 0.070 b | 2.807 ± 0.365 c | |
| HF | DWI | 1.94 ± 0.18 c | 4.75 ± 0.04 c | 12.20 ± 0.04 a | 129.20 ± 0.63 d | 36.47 ± 0.20 b | 45.17 ± 0.17 b | 35.48 ± 0.22 b | 2.95 ± 0.06 b | — | — | — |
| AAI | 2.64 ± 0.06 b | 6.45 ± 0.06 a | 9.82 ± 2.29 b | 138.52 ± 1.19 c | 44.17 ± 0.30 a | 72.60 ± 0.62 a | 68.63 ± 0.59 a | 7.64 ± 0.16 a | — | — | — | |
| DWIC | 1.51 ± 0.06 d | 4.87 ± 0.02 b | 7.42 ± 0.13 c | 224.13 ± 0.39 a | 24.57 ± 0.07 c | 34.45 ± 0.14 c | 24.84 ± 0.02 d | 2.09 ± 0.07 c | — | — | — | |
| AAIC | 3.53 ± 0.05 a | 3.06 ± 0.02 d | 7.03 ± 0.68 c | 209.49 ± 0.22 b | 16.27 ± 0.03 d | 21.37 ± 0.09 d | 28.40 ± 0.07 c | 1.37 ± 0.06 d | — | — | — | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sun, Y.; Liu, Y.; Li, J.; Yan, S. Acetic Acid Immersion Alleviates the Softening of Cooked Sagittaria sagittifolia L. Slices by Affecting Cell Wall Polysaccharides. Foods 2023, 12, 506. https://doi.org/10.3390/foods12030506
Sun Y, Liu Y, Li J, Yan S. Acetic Acid Immersion Alleviates the Softening of Cooked Sagittaria sagittifolia L. Slices by Affecting Cell Wall Polysaccharides. Foods. 2023; 12(3):506. https://doi.org/10.3390/foods12030506
Chicago/Turabian StyleSun, Yangyang, Yanzhao Liu, Jie Li, and Shoulei Yan. 2023. "Acetic Acid Immersion Alleviates the Softening of Cooked Sagittaria sagittifolia L. Slices by Affecting Cell Wall Polysaccharides" Foods 12, no. 3: 506. https://doi.org/10.3390/foods12030506
APA StyleSun, Y., Liu, Y., Li, J., & Yan, S. (2023). Acetic Acid Immersion Alleviates the Softening of Cooked Sagittaria sagittifolia L. Slices by Affecting Cell Wall Polysaccharides. Foods, 12(3), 506. https://doi.org/10.3390/foods12030506