
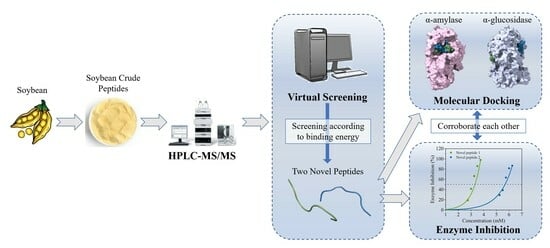
Virtual Screening Technology for Two Novel Peptides in Soybean as Inhibitors of α-Amylase and α-Glucosidase



Abstract
:
1. Introduction
2. Materials and Methods
2.1. Chemicals
2.2. Methods
2.2.1. α-Amylase Inhibition Assay In Vitro
2.2.2. α-Glucosidase Inhibition Assay In Vitro
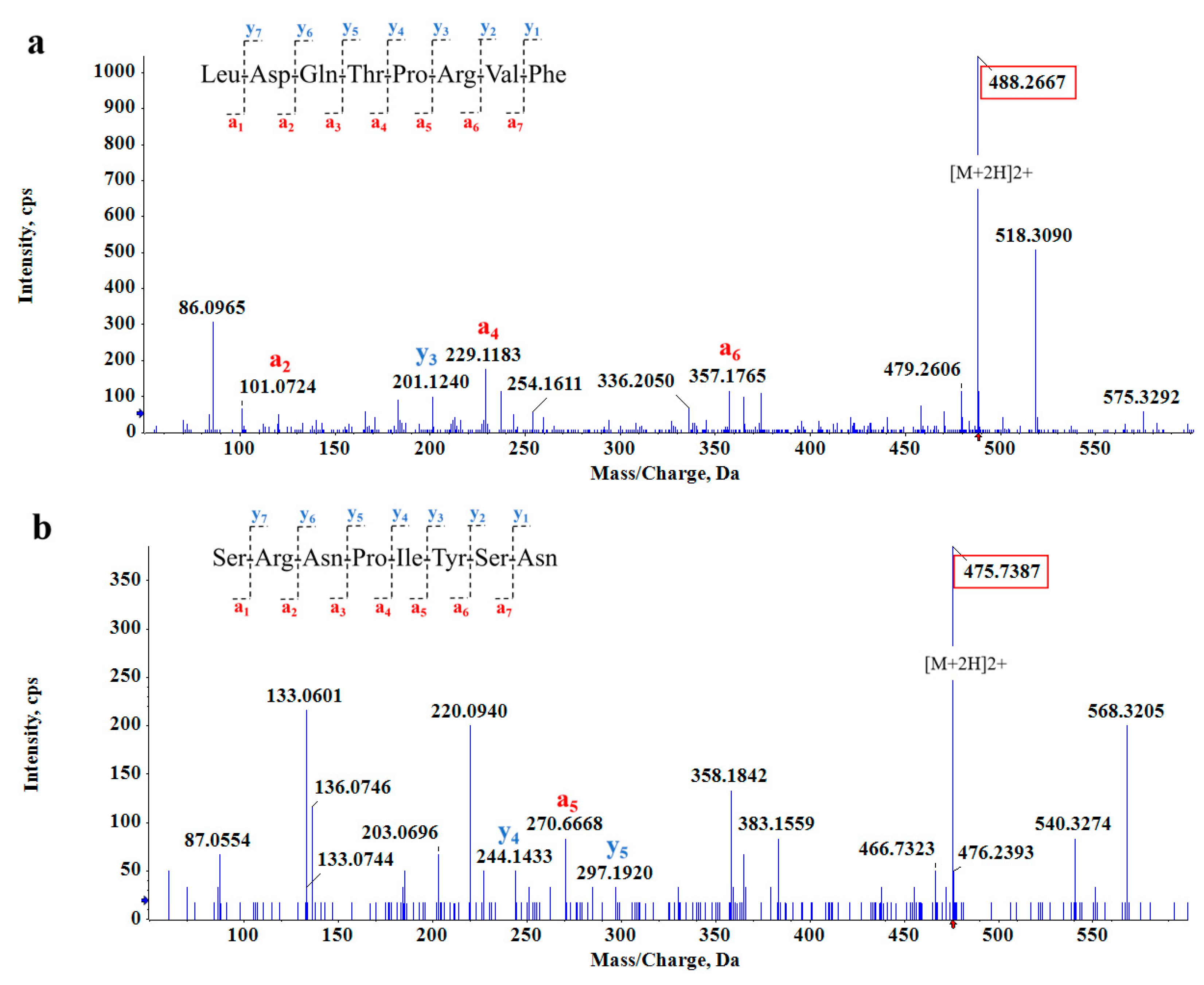
2.2.3. Structural Identification of Soybean Peptides (SPs)
2.2.4. Virtual Screening
2.2.5. Molecular Docking
2.2.6. Prediction of Physicochemical Properties
2.2.7. Statistical Analysis
3. Results
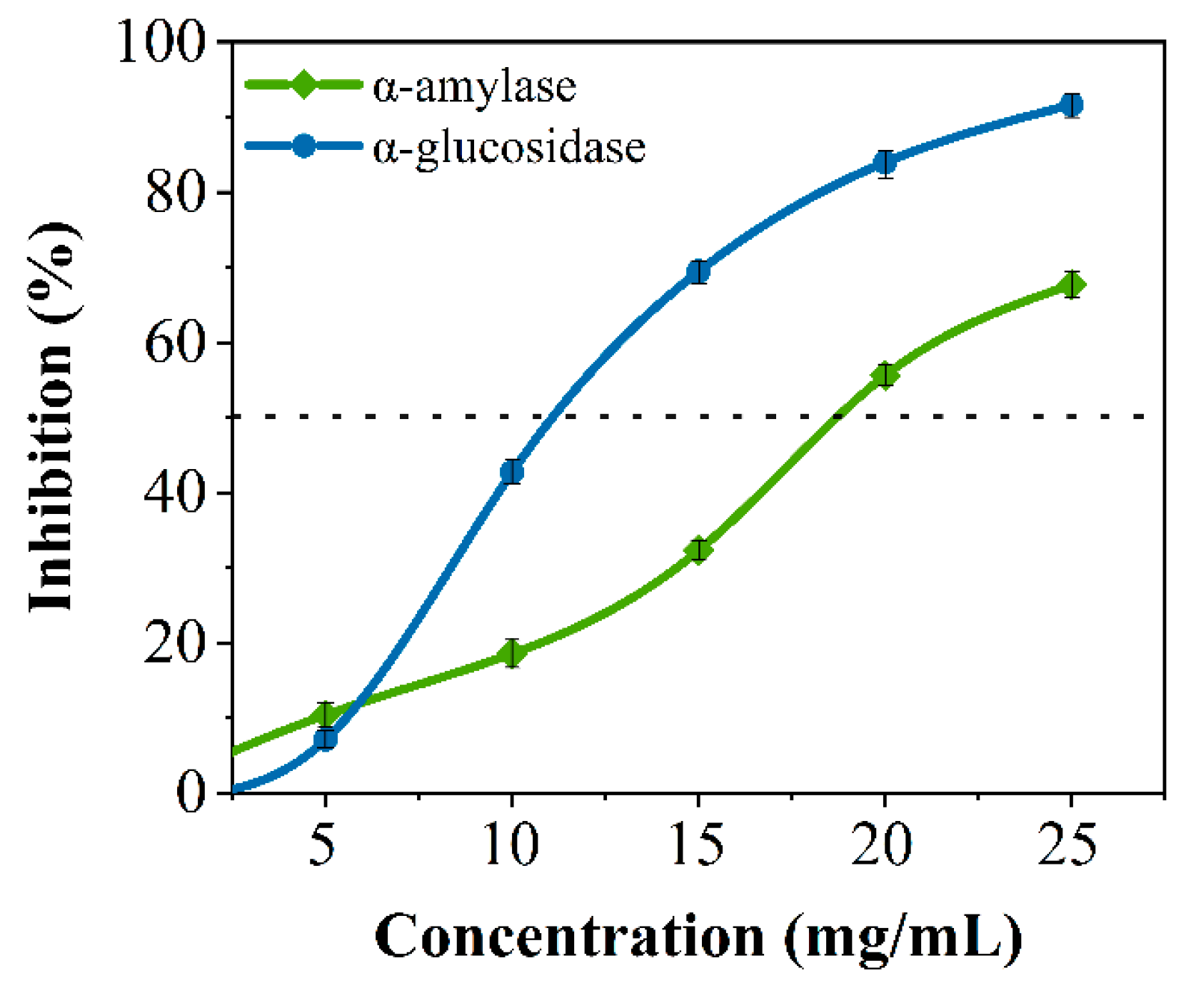
3.1. Inhibitory Effects of Crude SPs for α-Amylase and α-Glucosidase In Vitro
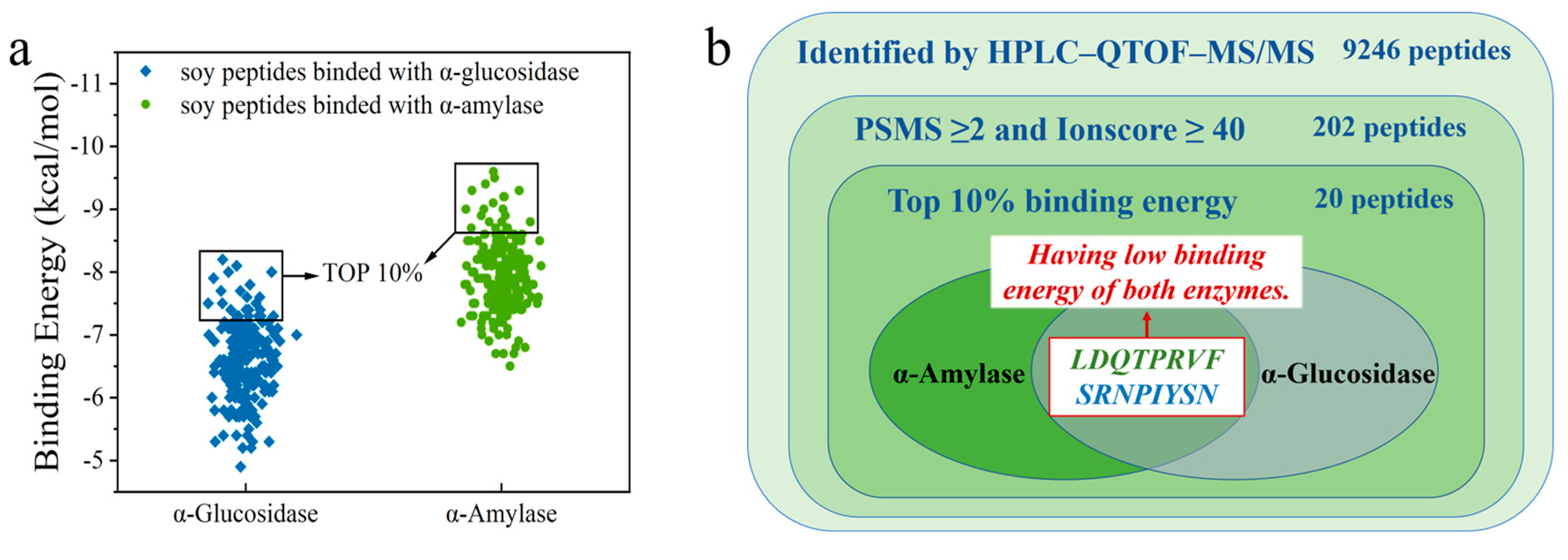
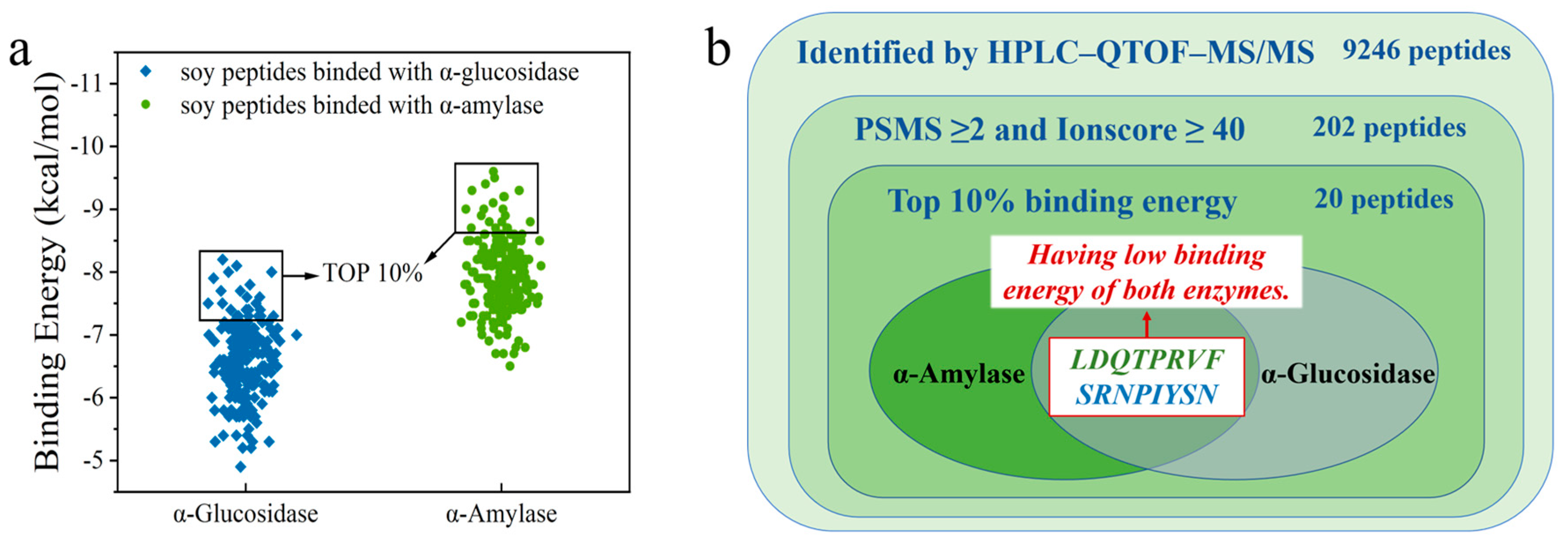
3.2. Peptide Fragment Screening with α-Amylase and α-Glucosidase Inhibitory Activity
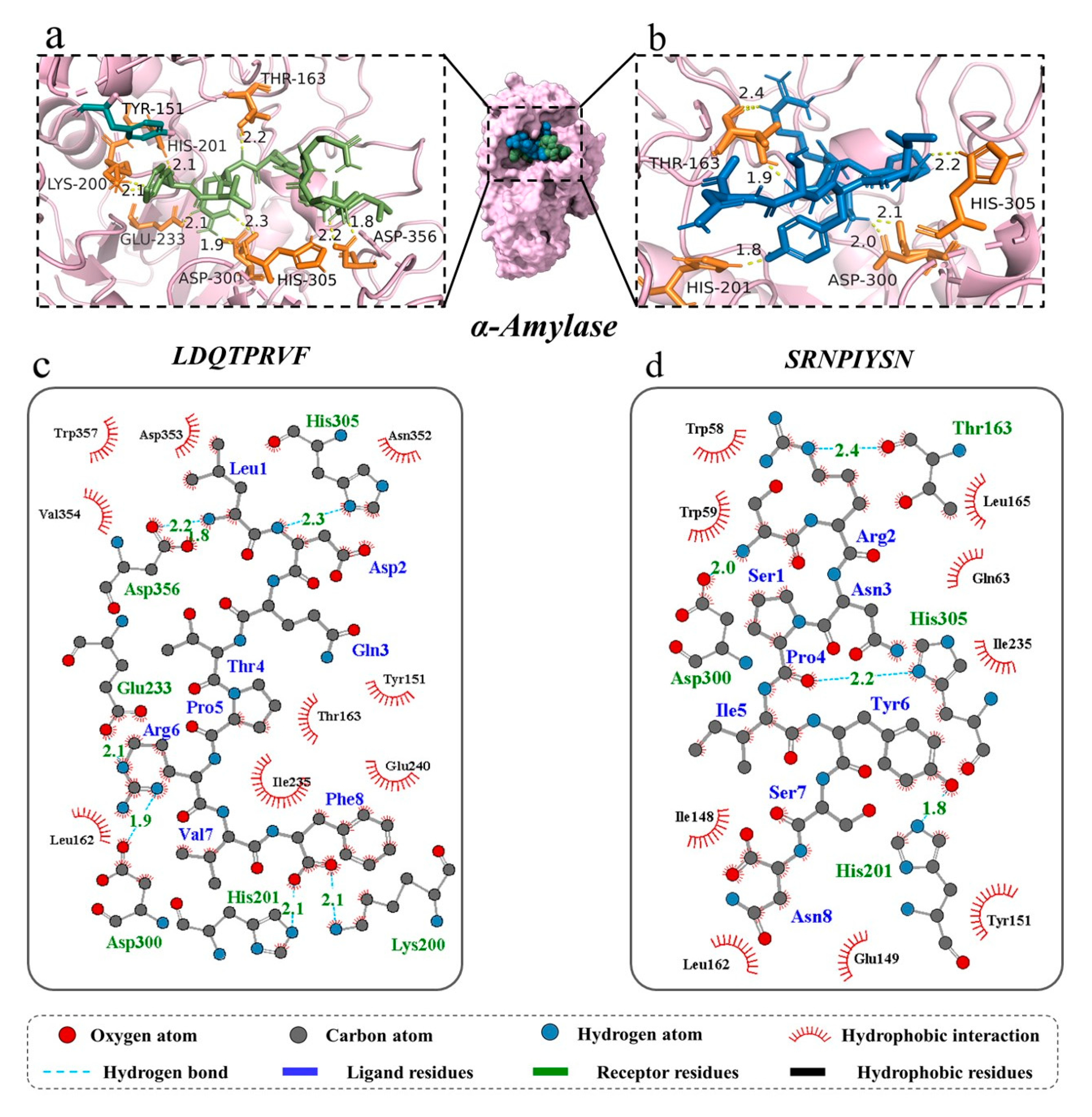
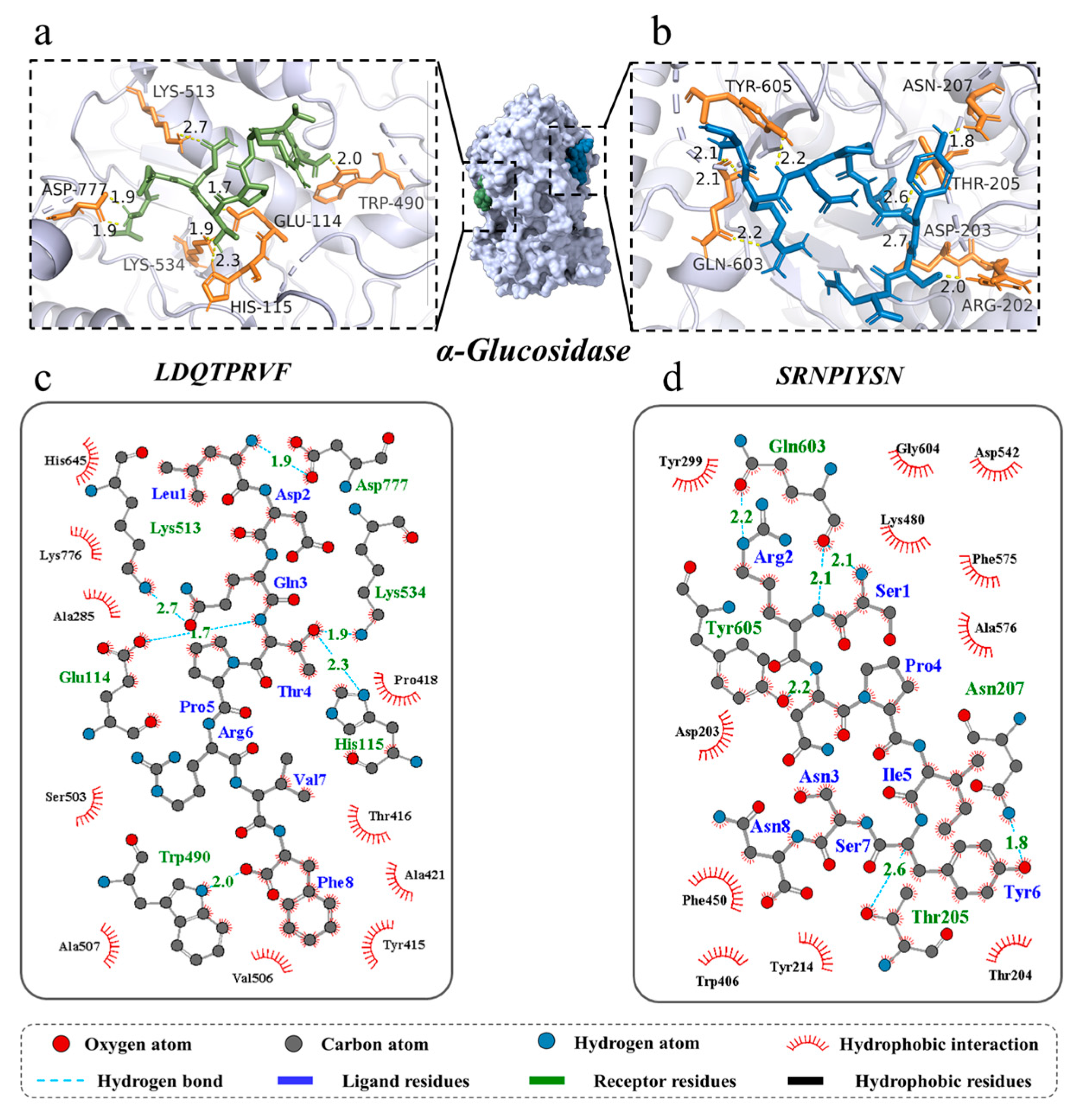
3.3. Molecular Docking for Screened Peptides
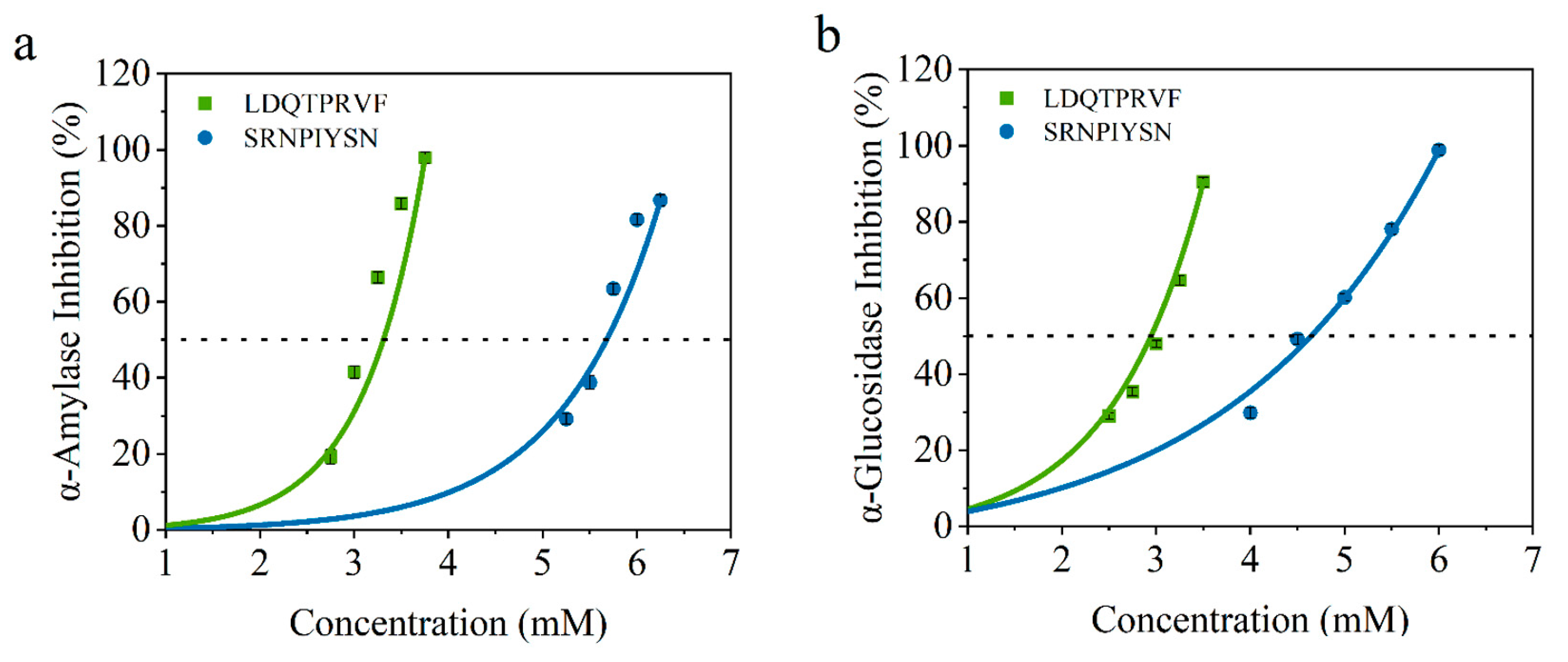
3.4. Inhibitory Activity Verification for Screened Peptides In Vitro
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Laakso, M.; Kuusisto, J. Insulin resistance and hyperglycaemia in cardiovascular disease development. Nat. Rev. Endocrinol. 2014, 10, 293–302. [Google Scholar] [CrossRef] [PubMed]
- Zheng, Y.; Ley, S.H.; Hu, F.B. Global aetiology and epidemiology of type 2 diabetes mellitus and its complications. Nat. Rev. Endocrinol. 2018, 14, 88–98. [Google Scholar] [CrossRef] [PubMed]
- Lin, Y.; Sun, Z. Current views on type 2 diabetes. J. Endocrinol. 2010, 204, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.-P.; He, H.; Lu, Y.-H. Four Flavonoid Compounds from Phyllostachys edulis Leaf Extract Retard the Digestion of Starch and Its Working Mechanisms. J. Agric. Food Chem. 2014, 62, 7760–7770. [Google Scholar] [CrossRef] [PubMed]
- Podsędek, A.; Majewska, I.; Kucharska, A.Z. Inhibitory Potential of Red Cabbage against Digestive Enzymes Linked to Obesity and Type 2 Diabetes. J. Agric. Food Chem. 2017, 65, 7192–7199. [Google Scholar] [CrossRef] [PubMed]
- Acquah, C.; Dzuvor, C.K.O.; Tosh, S.; Agyei, D. Anti-diabetic effects of bioactive peptides: Recent advances and clinical implications. Crit. Rev. Food Sci. Nutr. 2022, 62, 2158–2171. [Google Scholar] [CrossRef] [PubMed]
- Yilmazer-Musa, M.; Griffith, A.M.; Michels, A.J.; Schneider, E.; Frei, B. Grape Seed and Tea Extracts and Catechin 3-Gallates Are Potent Inhibitors of alpha-Amylase and alpha-Glucosidase Activity. J. Agric. Food Chem. 2012, 60, 8924–8929. [Google Scholar] [CrossRef] [PubMed]
- Singh, B.P.; Vij, S.; Hati, S. Functional significance of bioactive peptides derived from soybean. Peptides 2014, 54, 171–179. [Google Scholar] [CrossRef]
- Duffuler, P.; Bhullar, K.S.; Zani, S.C.d.C.; Wu, J. Bioactive Peptides: From Basic Research to Clinical Trials and Commercialization. J. Agric. Food Chem. 2022, 70, 3585–3595. [Google Scholar] [CrossRef]
- Coscueta, E.R.; Campos, D.A.; Osório, H.; Nerli, B.B.; Pintado, M. Enzymatic soy protein hydrolysis: A tool for bio functional food ingredient production. Food Chem. X 2019, 1, 7. [Google Scholar] [CrossRef]
- Admassu, H.; Gasmalla, M.A.; Yang, R.; Zhao, W. Identification of Bioactive Peptides with alpha-Amylase Inhibitory Potential from Enzymatic Protein Hy-drolysates of Red Seaweed (Porphyra spp). J. Agric. Food Chem. 2018, 66, 4872–4882. [Google Scholar] [CrossRef] [PubMed]
- Zhu, J.; Chen, X.; Luo, J.; Liu, Y.; Wang, B.; Liang, Z.; Li, L. Insight into the binding modes and mechanisms of inhibition between soybean-peptides and α-amylase based on spectrofluorimetry and kinetic analysis. LWT-Food Sci. Technol. 2021, 142, 110977. [Google Scholar] [CrossRef]
- Zhou, M.; Ren, G.; Zhang, B.; Ma, F.; Fan, J.; Qiu, Z. Screening and identification of a novel antidiabetic peptide from collagen hydrolysates of Chinese giant sal-amander skin: Network pharmacology, inhibition kinetics and protection of IR-HepG2 cells. Food Funct. 2022, 13, 3329–3342. [Google Scholar] [CrossRef]
- Salami, M.; Motahar, S.F.S.; Ariaeenejad, S.; Mamaghani, A.S.A.; Kavousi, K.; Moosavi-Movahedi, A.A.; Salekdeh, G.H. The novel homologue of the human alpha-glucosidase inhibited by the non-germinated and germinated quinoa protein hydrolysates after in vitro gastrointestinal digestion. J. Food Biochem. 2022, 46, e14030. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Wu, F.; He, Z.; Fang, X.; Liu, X. Optimization and Molecular Mechanism of Novel α-Glucosidase Inhibitory Peptides Derived from Camellia Seed Cake through Enzymatic Hydrolysis. Foods 2023, 12, 393. [Google Scholar] [CrossRef] [PubMed]
- Esfandi, R.; Seidu, I.; Willmore, W.; Tsopmo, A. Antioxidant, pancreatic lipase, and alpha-amylase inhibitory properties of oat bran hydrolyzed proteins and peptides. J. Food Biochem. 2022, 46, e13762. [Google Scholar] [CrossRef] [PubMed]
- Liu, W.; Li, H.; Wen, Y.; Liu, Y.; Wang, J.; Sun, B. Molecular Mechanism for the α-Glucosidase Inhibitory Effect of Wheat Germ Peptides. J. Agric. Food Chem. 2021, 69, 15231–15239. [Google Scholar] [CrossRef] [PubMed]
- Sun, L.; Wang, Y.; Miao, M. Inhibition of α-amylase by polyphenolic compounds: Substrate digestion, binding interactions and nutritional intervention. Trends Food Sci. Technol. 2020, 104, 190–207. [Google Scholar] [CrossRef]
- Yu, Z.; Yin, Y.; Zhao, W.; Liu, J.; Chen, F. Anti-diabetic activity peptides from albumin against alpha-glucosidase and alpha-amylase. Food Chem. 2012, 135, 2078–2085. [Google Scholar] [CrossRef]
- Sanjukta, S.; Rai, A.K. Production of bioactive peptides during soybean fermentation and their potential health benefits. Trends Food Sci. Technol. 2016, 50, 1–10. [Google Scholar] [CrossRef]
- Ku, Y.-S.; Cheng, S.-S.; Gerhardt, A.; Cheung, M.-Y.; Contador, C.A.; Poon, L.-Y.W.; Lam, H.-M. Secretory Peptides as Bullets: Effector Peptides from Pathogens against Antimicrobial Peptides from Soybean. Int. J. Mol. Sci. 2020, 21, 9294. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Zhang, Y.; Zou, Y.; Li, L.; Yan, J.; Chen, S.; Zhang, S.; Zhu, J. Heat-induced amorphous aggregates assembly of soy protein modulate in vitro digestibility of potato starch. Int. J. Biol. Macromol. 2023, 227, 222–230. [Google Scholar] [CrossRef] [PubMed]
- Jiang, M.; Yan, H.; He, R.; Ma, Y. Purification and a molecular docking study of α-glucosidase-inhibitory peptides from a soybean protein hydrolysate with ultrasonic pretreatment. Eur. Food Res. Technol. 2018, 244, 1995–2005. [Google Scholar] [CrossRef]
- Wang, R.; Zhao, H.; Pan, X.; Orfila, C.; Lu, W.; Ma, Y. Preparation of bioactive peptides with antidiabetic, antihypertensive, and antioxidant activities and identification of α-glucosidase inhibitory peptides from soy protein. Food Sci. Nutr. 2019, 7, 1848–1856. [Google Scholar] [CrossRef] [PubMed]
- Sangilimuthu, A.Y.; Sivaraman, T.; Chandrasekaran, R.; Sundaram, K.M.; Ekambaram, G. Screening chemical inhibitors for alpha-amylase from leaves extracts of Murraya koenigii (Linn.) and Aegle marmelos L. J. Complement. Integr. Med. 2020, 18, 51–57. [Google Scholar] [CrossRef] [PubMed]
- Ren, L.; Qin, X.; Cao, X.; Wang, L.; Bai, F.; Bai, G.; Shen, Y. Structural insight into substrate specificity of human intestinal maltase-glucoamylase. Protein Cell 2011, 2, 827–836. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Wu, D.; Liu, J.; Li, G.; Zhang, Z.; Chen, C.; Zhang, L.; Li, J. Characterization of xanthine oxidase inhibitory activities of phenols from pickled radish with molecular simulation. Food Chem. X 2022, 14, 10. [Google Scholar] [CrossRef] [PubMed]
- Alnuaimi, A.; Ajayi, F.F.; Hamdi, M.; Mudgil, P.; Kamal, H.; Gan, C.Y.; Maqsood, S. A comparative analysis of anti-lipidemic potential of soybean (Glycine max) protein hydrolysates obtained from different ripening stages: Identification, and molecular interaction mechanisms of novel bioactive peptides. Food Chem. 2023, 402, 134192. [Google Scholar] [CrossRef]
- Ninomiya, K.; Yamaguchi, Y.; Shinmachi, F.; Kumagai, H.; Kumagai, H. Suppression of postprandial blood glucose elevation by buckwheat (Fagpopyrum esculentum) albumin hydrolysate and identification of the peptide responsible to the function. Food Sci. Hum. Wellness 2022, 11, 992–998. [Google Scholar] [CrossRef]
- Siddiqui, M.R.; AlOthman, Z.A.; Rahman, N. Analytical techniques in pharmaceutical analysis: A review. Arab. J. Chem. 2017, 10, S1409–S1421. [Google Scholar] [CrossRef]
- Shackleton, C. Clinical steroid mass spectrometry: A 45-year history culminating in HPLC–MS/MS becoming an essential tool for patient diagnosis. J. Steroid Biochem. Mol. Biol. 2010, 121, 481–490. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; He, S.; Rui, X.; Simpson, B.K. Interactions of C. frondosa-derived inhibitory peptides against angiotensin I-converting enzyme (ACE), alpha-amylase and lipase. Food Chem. 2022, 367, 130695. [Google Scholar] [CrossRef] [PubMed]
- Daliri, E.B.; Oh, D.H.; Lee, B.H. Bioactive Peptides. Foods 2017, 6, 32. [Google Scholar] [CrossRef] [PubMed]
- Galili, G.; Amir, R.; Fernie, A.R. The Regulation of Essential Amino Acid Synthesis and Accumulation in Plants. Annu. Rev. Plant Biol. 2016, 67, 153–178. [Google Scholar] [CrossRef] [PubMed]
- Lu, Y.; Zhou, W.; Feng, Y.; Li, Y.; Liu, K.; Liu, L.; Lin, D.; He, Z.; Wu, X. Acteoside and Acyl-Migrated Acteoside, Compounds in Chinese Kudingcha Tea, Inhibit alpha-Amylase In Vitro. J. Med. Food 2017, 20, 577–585. [Google Scholar] [CrossRef]
- Goldbach, L.; Vermeulen, B.J.A.; Caner, S.; Liu, M.; Tysoe, C.; van Gijzel, L.; Yoshisada, R.; Trellet, M.; van Ingen, H.; Brayer, G.D.; et al. Folding Then Binding vs. Folding Through Binding in Macrocyclic Peptide Inhibitors of Human Pancreatic α-Amylase. ACS Chem. Biol. 2019, 14, 1751–1759. [Google Scholar] [CrossRef]
- Sim, L.; Quezada-Calvillo, R.; Sterchi, E.E.; Nichols, B.L.; Rose, D.R. Human Intestinal Maltase–Glucoamylase: Crystal Structure of the N-Terminal Catalytic Subunit and Basis of Inhibition and Substrate Specificity. J. Mol. Biol. 2008, 375, 782–792. [Google Scholar] [CrossRef]
- Manivasagan, P.; Venkatesan, J.; Sivakumar, K.; Kim, S.K. Actinobacterial enzyme inhibitors—A review. Crit. Rev. Microbiol. 2013, 41, 261–272. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| PEPTIDES | LDQTPRVF | SRNPIYSN |
|---|---|---|
| Calculated mass (Da) | 975.10 | 950.01 |
| Observed mass (Da) | 974.90 | 949.80 |
| Isoelectric point | pH 6.64 | pH 9.57 |
| Grand average of hydropathicity (GRAVY) | −0.375 | −1.438 |
| Toxicity | Non-toxic | Non-toxic |
| Extinction coefficient (M−1 cm−1) | 0 | 1280 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tang, X.; Chen, X.; Wang, H.; Yang, J.; Li, L.; Zhu, J.; Liu, Y. Virtual Screening Technology for Two Novel Peptides in Soybean as Inhibitors of α-Amylase and α-Glucosidase. Foods 2023, 12, 4387. https://doi.org/10.3390/foods12244387
Tang X, Chen X, Wang H, Yang J, Li L, Zhu J, Liu Y. Virtual Screening Technology for Two Novel Peptides in Soybean as Inhibitors of α-Amylase and α-Glucosidase. Foods. 2023; 12(24):4387. https://doi.org/10.3390/foods12244387
Chicago/Turabian StyleTang, Xiyao, Xu Chen, Hong Wang, Jinyi Yang, Lin Li, Jie Zhu, and Yujia Liu. 2023. "Virtual Screening Technology for Two Novel Peptides in Soybean as Inhibitors of α-Amylase and α-Glucosidase" Foods 12, no. 24: 4387. https://doi.org/10.3390/foods12244387
APA StyleTang, X., Chen, X., Wang, H., Yang, J., Li, L., Zhu, J., & Liu, Y. (2023). Virtual Screening Technology for Two Novel Peptides in Soybean as Inhibitors of α-Amylase and α-Glucosidase. Foods, 12(24), 4387. https://doi.org/10.3390/foods12244387