
Use of Human Intestinal Enteroids for Recovery of Infectious Human Norovirus from Berries and Lettuce
Abstract
:
1. Introduction
2. Materials and Methods
2.1. Human Intestinal Enteroids
2.2. Viruses
2.3. TCID50 Assays
2.4. Virus Recovery from Foods
2.5. RNA Extraction and Purification
2.6. Detection by Real-Time RT-qPCR
2.7. Statistical Analysis
3. Results
3.1. Screening of NoV Fecal Filtrate Replication with J2 HIEs
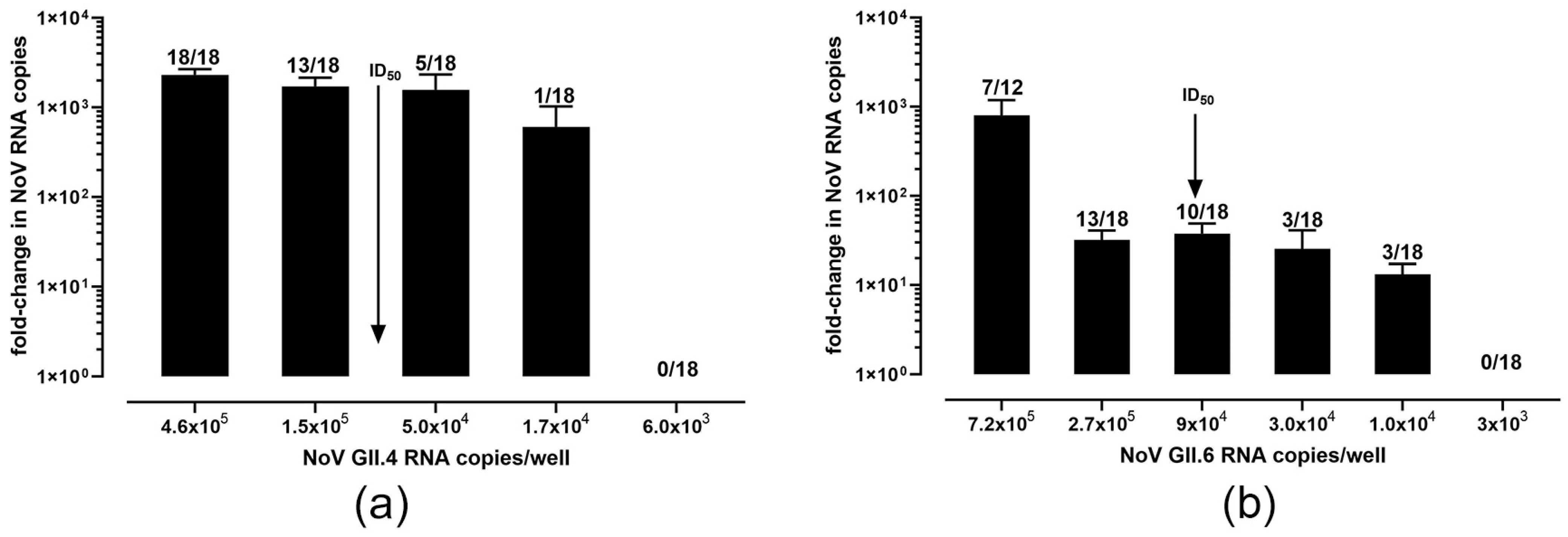
3.2. Characterization of Culture Conditions for GII.4[P16] and GII.6[P7] Genotypes
3.3. Infection of J2 HIEs with Virus Recovered from Frozen Raspberries
3.4. Infection of J2 HIEs with Virus Recovered from Frozen Strawberries
3.5. Infection of J2 HIEs with Virus Recovered from Fresh Lettuce
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bányai, K.; Estes, M.K.; Martella, V.; Parashar, U.D. Viral gastroenteritis. Lancet 2018, 392, 175–186. [Google Scholar] [CrossRef] [PubMed]
- Burke, R.M.; Mattison, C.P.; Pindyck, T.; Dahl, R.M.; Rudd, J.; Bi, D.; Curns, A.T.; Parashar, U.; Hall, A.J. Burden of Norovirus in the United States, as Estimated Based on Administrative Data: Updates for Medically Attended Illness and Mortality, 2001–2015. Clin. Infect. Dis. 2021, 73, e1–e8. [Google Scholar] [CrossRef] [PubMed]
- Carter, M.J. Enterically infecting viruses: Pathogenicity, transmission and significance for food and waterborne infection. J. Appl. Microbiol. 2005, 98, 1354–1380. [Google Scholar] [CrossRef] [PubMed]
- Seitz, S.R.; Leon, J.S.; Schwab, K.J.; Lyon, G.M.; Dowd, M.; McDaniels, M.; Abdulhafid, G.; Fernandez, M.L.; Lindesmith, L.C.; Baric, R.S.; et al. Norovirus infectivity in humans and persistence in water. Appl. Environ. Microbiol. 2011, 77, 6884–6888. [Google Scholar] [CrossRef] [PubMed]
- Lou, F.; DiCaprio, E.; Li, X.; Dai, X.; Ma, Y.; Hughes, J.; Chen, H.; Kingsley, D.H.; Li, J. Variable High-Pressure-Processing Sensitivities for Genogroup II Human Noroviruses. Appl. Environ. Microbiol. 2016, 82, 6037–6045. [Google Scholar] [CrossRef] [PubMed]
- Ettayebi, K.; Crawford, S.E.; Murakami, K.; Broughman, J.R.; Karandikar, U.; Tenge, V.R.; Neill, F.H.; Blutt, S.E.; Zeng, X.L.; Qu, L.; et al. Replication of human noroviruses in stem cell-derived human enteroids. Science 2016, 353, 1387–1393. [Google Scholar] [CrossRef]
- Bozkurt, H.; Phan-Thien, K.Y.; van Ogtrop, F.; Bell, T.; McConchie, R. Outbreaks, occurrence, and control of norovirus and hepatitis a virus contamination in berries: A review. Crit. Rev. Food Sci. Nutr. 2021, 61, 116–138. [Google Scholar] [CrossRef]
- Mäde, D.; Trübner, K.; Neubert, E.; Höhne, M.; Johne, R. Detection and typing of norovirus from frozen strawberries involved in a large-scale gastroenteritis outbreak in Germany. Food Environ. Virol. 2013, 5, 162–168. [Google Scholar] [CrossRef]
- Fraisse, A.; Coudray-Meunier, C.; Martin-Latil, S.; Hennechart-Collette, C.; Delannoy, S.; Fach, P.; Perelle, S. Digital RT-PCR method for hepatitis A virus and norovirus quantification in soft berries. Int. J. Food Microbiol. 2017, 243, 36–45. [Google Scholar] [CrossRef]
- Kotwal, G.; Cannon, J.L. Environmental persistence and transfer of enteric viruses. Curr. Opin. Virol. 2014, 4, 37–43. [Google Scholar] [CrossRef]
- Matthews, J.E.; Dickey, B.W.; Miller, R.D.; Felzer, J.R.; Dawson, B.P.; Lee, A.S.; Rocks, J.J.; Kiel, J.; Montes, J.S.; Moe, C.L.; et al. The epidemiology of published norovirus outbreaks: A review of risk factors associated with attack rate and genogroup. Epidemiol. Infect. 2012, 140, 1161–1172. [Google Scholar] [CrossRef] [PubMed]
- Atmar, R.L.; Opekun, A.R.; Gilger, M.A.; Estes, M.K.; Crawford, S.E.; Neill, F.H.; Ramani, S.; Hill, H.; Ferreira, J.; Graham, D.Y. Determination of the 50% human infectious dose for Norwalk virus. J. Infect. Dis. 2014, 209, 1016–1022. [Google Scholar] [CrossRef] [PubMed]
- Duizer, E.; Schwab, K.J.; Neill, F.H.; Atmar, R.L.; Koopmans, M.P.G.; Estes, M.K. Laboratory efforts to cultivate noroviruses. J. Gen. Virol. 2004, 85, 79–87. [Google Scholar] [CrossRef] [PubMed]
- Herbst-Kralovetz, M.M.; Radtke, A.L.; Lay, M.K.; Hjelm, B.E.; Bolick, A.N.; Sarker, S.S.; Atmar, R.L.; Kingsley, D.H.; Arntzen, C.J.; Estes, M.K.; et al. Lack of norovirus replication and histo-blood group antigen expression in 3-dimensional intestinal epithelial cells. Emerg. Infect. Dis. 2013, 19, 431–438. [Google Scholar] [CrossRef]
- Chan, M.C.; Cheung, S.K.C.; Mohammad, K.N.; Chan, J.C.M.; Estes, M.K.; Chan, P.K.S. Use of Human Intestinal Enteroids to Detect Human Norovirus Infectivity. Emerg. Infect. Dis. 2019, 25, 1730–1735. [Google Scholar] [CrossRef]
- Costantini, V.; Morantz, E.K.; Browne, H.; Ettayebi, K.; Zeng, X.L.; Atmar, R.L.; Estes, M.K.; Vinjé, J. Human Norovirus Replication in Human Intestinal Enteroids as Model to Evaluate Virus Inactivation. Emerg. Infect. Dis. 2018, 24, 1453–1464. [Google Scholar] [CrossRef]
- Desdouits, M.; Polo, D.; Le Mennec, C.; Strubbia, S.; Zeng, X.L.; Ettayebi, K.; Atmar, R.L.; Estes, M.K.; Le Guyader, F.S. Use of Human Intestinal Enteroids to Evaluate Persistence of Infectious Human Norovirus in Seawater. Emerg. Infect. Dis. 2022, 28, 1475–1479. [Google Scholar] [CrossRef]
- Ettayebi, K.; Tenge, V.R.; Cortes-Penfield, N.W.; Crawford, S.E.; Neill, F.H.; Zeng, X.L.; Yu, X.; Ayyar, B.V.; Burrin, D.; Ramani, S.; et al. New Insights and Enhanced Human Norovirus Cultivation in Human Intestinal Enteroids. mSphere 2021, 6, e01136-20. [Google Scholar] [CrossRef]
- Ford-Siltz, L.A.; Wales, S.; Tohma, K.; Gao, Y.; Parra, G.I. Genotype-Specific Neutralization of Norovirus Is Mediated by Antibodies Against the Protruding Domain of the Major Capsid Protein. J. Infect. Dis. 2022, 225, 1205–1214. [Google Scholar] [CrossRef]
- Kennedy, L.C.; Costantini, V.P.; Huynh, K.A.; Loeb, S.K.; Jennings, W.C.; Lowry, S.; Mattioli, M.C.; Vinjé, J.; Boehm, A.B. Persistence of Human Norovirus (GII) in Surface Water: Decay Rate Constants and Inactivation Mechanisms. Environ. Sci. Technol. 2023, 57, 3671–3679. [Google Scholar] [CrossRef]
- Overbey, K.N.; Zachos, N.C.; Coulter, C.; Schwab, K.J. Optimizing Human Intestinal Enteroids for Environmental Monitoring of Human Norovirus. Food Environ. Virol. 2021, 13, 470–484. [Google Scholar] [CrossRef] [PubMed]
- Escudero-Abarca, B.I.; Goulter, R.M.; Bradshaw, J.; Faircloth, J.; Leslie, R.A.; Manuel, C.S.; Arbogast, J.W.; Jaykus, L.A. Efficacy of an alcohol-based surface disinfectant formulation against human norovirus. J. Appl. Microbiol. 2022, 132, 3590–3600. [Google Scholar] [CrossRef] [PubMed]
- Escudero-Abarca, B.I.; Goulter, R.M.; Manuel, C.S.; Leslie, R.A.; Green, K.; Arbogast, J.W.; Jaykus, L.A. Comparative Assessment of the Efficacy of Commercial Hand Sanitizers Against Human Norovirus Evaluated by an in vivo Fingerpad Method. Front. Microbiol. 2022, 13, 869087. [Google Scholar] [CrossRef] [PubMed]
- Ettayebi, K.; Salmen, W.; Imai, K.; Hagi, A.; Neill, F.H.; Atmar, R.L.; Prasad, B.V.V.; Estes, M.K. Antiviral Activity of Olanexidine-Containing Hand Rub against Human Noroviruses. mBio 2022, 13, e0284821. [Google Scholar] [CrossRef]
- Hayashi, T.; Yamaoka, Y.; Ito, A.; Kamaishi, T.; Sugiyama, R.; Estes, M.K.; Muramatsu, M.; Murakami, K. Evaluation of Heat Inactivation of Human Norovirus in Freshwater Clams Using Human Intestinal Enteroids. Viruses 2022, 14, 1014. [Google Scholar] [CrossRef]
- Yang, Z.; Vinjé, J.; Elkins, C.A.; Kulka, M. Complete Genome Sequence of Human Norovirus Strain GII.P7-GII.6 Detected in a Patient in the United States in 2014. Genome Announc. 2016, 4, e01211-16. [Google Scholar] [CrossRef]
- Lei, C.; Yang, J.; Hu, J.; Sun, X. On the Calculation of TCID(50) for Quantitation of Virus Infectivity. Virol. Sin. 2021, 36, 141–144. [Google Scholar] [CrossRef]
- Williams-Woods, J.; Rodriguez, R.; Marchant, J.; Swinford, A.G.; Burkhardt, W., III. Chapter 26—Concentration, Extraction and Detection of Enteric Viruses from Food. In Bacteriological Analytical Manual; Food and Drug Administration: Washington, DC, USA, 2022. Available online: https://www.fda.gov/media/160119/download (accessed on 21 November 2023).
- Yu, C.; Wales, S.Q.; Mammel, M.K.; Hida, K.; Kulka, M. Optimizing a custom tiling microarray for low input detection and identification of unamplified virus targets. J. Virol. Methods 2016, 234, 54–64. [Google Scholar] [CrossRef]
- Yang, Z.; Mammel, M.; Wales, S.Q. Near-Complete Genome Sequence of a Human Norovirus GII.1[Pg] Strain Associated with Acute Gastroenteritis, Determined Using Long-Read Sequencing. Microbiol. Resour. Announc. 2021, 10, e0040121. [Google Scholar] [CrossRef]
- Müller, L.; Rasmussen, L.D.; Jensen, T.; Schultz, A.C.; Kjelsø, C.; Barnadas, C.; Sigsgaard, K.; Larsen, A.R.; Widstrup Jensen, C.; Jeppesen, S.; et al. Series of Norovirus Outbreaks Caused by Consumption of Green Coral Lettuce, Denmark, April 2016. PLoS Curr. 2016, 8. [Google Scholar] [CrossRef]
- Greening, G.E.; Cannon, J.L. Human and Animal Viruses in Food (Including Taxonomy of Enteric Viruses). In Viruses in Foods; Springer: Cham, Switzerland, 2016; pp. 5–57. [Google Scholar] [CrossRef]
- Perrin, A.; Loutreul, J.; Boudaud, N.; Bertrand, I.; Gantzer, C. Rapid, simple and efficient method for detection of viral genomes on raspberries. J. Virol. Methods 2015, 224, 95–101. [Google Scholar] [CrossRef] [PubMed]
- Stals, A.; Van Coillie, E.; Uyttendaele, M. Viral genes everywhere: Public health implications of PCR-based testing of foods. Curr. Opin. Virol. 2013, 3, 69–73. [Google Scholar] [CrossRef] [PubMed]
- Bartsch, C.; Szabo, K.; Dinh-Thanh, M.; Schrader, C.; Trojnar, E.; Johne, R. Comparison and optimization of detection methods for noroviruses in frozen strawberries containing different amounts of RT-PCR inhibitors. Food Microbiol. 2016, 60, 124–130. [Google Scholar] [CrossRef] [PubMed]
- Schrader, C.; Schielke, A.; Ellerbroek, L.; Johne, R. PCR inhibitors—Occurrence, properties and removal. J. Appl. Microbiol. 2012, 113, 1014–1026. [Google Scholar] [CrossRef] [PubMed]
- Jones, M.K.; Watanabe, M.; Zhu, S.; Graves, C.L.; Keyes, L.R.; Grau, K.R.; Gonzalez-Hernandez, M.B.; Iovine, N.M.; Wobus, C.E.; Vinjé, J.; et al. Enteric bacteria promote human and mouse norovirus infection of B cells. Science 2014, 346, 755–759. [Google Scholar] [CrossRef] [PubMed]
- Karst, S.M. The influence of commensal bacteria on infection with enteric viruses. Nat. Rev. Microbiol. 2016, 14, 197–204. [Google Scholar] [CrossRef] [PubMed]
- Hagbom, M.; Lin, J.; Falkeborn, T.; Serrander, L.; Albert, J.; Nordgren, J.; Sharma, S. Replication in Human Intestinal Enteroids of Infectious Norovirus from Vomit Samples. Emerg. Infect. Dis. 2021, 27, 2212–2214. [Google Scholar] [CrossRef]
- Kirby, A.E.; Streby, A.; Moe, C.L. Vomiting as a Symptom and Transmission Risk in Norovirus Illness: Evidence from Human Challenge Studies. PLoS ONE 2016, 11, e0143759. [Google Scholar] [CrossRef]
- Gustafsson, R.K.; Engdahl, E.E.; Fogdell-Hahn, A. Development and validation of a Q-PCR based TCID50 method for human herpesvirus 6. Virol. J. 2012, 9, 311. [Google Scholar] [CrossRef]
- Duong, T.; McAllister, J.; Eldahan, K.; Wang, J.; Onishi, E.; Shen, K.; Schrock, R.; Gu, B.; Wang, P. Improvement of Precision in Recombinant Adeno-Associated Virus Infectious Titer Assay with Droplet Digital PCR as an Endpoint Measurement. Hum. Gene Ther. 2023, 34, 742–757. [Google Scholar] [CrossRef]
- Sato, S.; Hisaie, K.; Kurokawa, S.; Suzuki, A.; Sakon, N.; Uchida, Y.; Yuki, Y.; Kiyono, H. Human Norovirus Propagation in Human Induced Pluripotent Stem Cell-Derived Intestinal Epithelial Cells. Cell Mol. Gastroenterol. Hepatol. 2019, 7, 686–688.e685. [Google Scholar] [CrossRef] [PubMed]
- ISO 15216-2:2019; Microbiology of the Food Chain: Horizontal Method for Determination of Hepatitis a Virus and Norovirus Using Real-Time RT-PCR. Part 2: Method for Detection. ISO: Geneva, Switzerland, 2019.
- Butot, S.; Putallaz, T.; Amoroso, R.; Sánchez, G. Inactivation of enteric viruses in minimally processed berries and herbs. Appl. Environ. Microbiol. 2009, 75, 4155–4161. [Google Scholar] [CrossRef] [PubMed]
- Hewitt, J.; Greening, G.E. Survival and persistence of norovirus, hepatitis A virus, and feline calicivirus in marinated mussels. J. Food Prot. 2004, 67, 1743–1750. [Google Scholar] [CrossRef] [PubMed]
- Summa, M.; von Bonsdorff, C.H.; Maunula, L. Evaluation of four virus recovery methods for detecting noroviruses on fresh lettuce, sliced ham, and frozen raspberries. J. Virol. Methods 2012, 183, 154–160. [Google Scholar] [CrossRef] [PubMed]
- De Keuckelaere, A.; Li, D.; Deliens, B.; Stals, A.; Uyttendaele, M. Batch testing for noroviruses in frozen raspberries. Int. J. Food Microbiol. 2015, 192, 43–50. [Google Scholar] [CrossRef] [PubMed]
- Hennechart-Collette, C.; Dehan, O.; Laurentie, M.; Fraisse, A.; Martin-Latil, S.; Perelle, S. Detection of norovirus, hepatitis A and hepatitis E viruses in multicomponent foodstuffs. Int. J. Food Microbiol. 2021, 337, 108931. [Google Scholar] [CrossRef]
- Raymond, P.; Paul, S.; Perron, A.; Deschênes, L. Norovirus Extraction from Frozen Raspberries Using Magnetic Silica Beads. Food Environ. Virol. 2021, 13, 248–258. [Google Scholar] [CrossRef]
- Atmar, R.L.; Ramani, S.; Estes, M.K. Human noroviruses: Recent advances in a 50-year history. Curr. Opin. Infect. Dis. 2018, 31, 422–432. [Google Scholar] [CrossRef]
- Ludwig-Begall, L.F.; Mauroy, A.; Thiry, E. Noroviruses-The State of the Art, Nearly Fifty Years after Their Initial Discovery. Viruses 2021, 13, 1541. [Google Scholar] [CrossRef]
- Esseili, M.A.; Saif, L.J.; Farkas, T.; Wang, Q. Feline Calicivirus, Murine Norovirus, Porcine Sapovirus, and Tulane Virus Survival on Postharvest Lettuce. Appl. Environ. Microbiol. 2015, 81, 5085–5092. [Google Scholar] [CrossRef]
- Hida, K.; Papafragkou, E.; Kulka, M. Testing for Human Norovirus and Recovery of Process Control in Outbreak-Associated Produce Items. J. Food Prot. 2018, 81, 105–114. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
{kind=link}
| Positive (≥5-Fold Increase) | Negative (≤5-Fold Increase) | |||||
|---|---|---|---|---|---|---|
| Genotype: | RNA cp: | Fold-inc: | GII Genotype: | RNA cp: | GI Genotype: | RNA cp: |
| GII.4 Sydney (BCM) | 3.00 × 105 | 2.9 × 103 | GII.4 (2014) * | 2.90 × 105 | GI.6[P11] * | 1.02 × 106 |
| GII.4[P16] (011617) * | 1.98 × 105 | 1.4 × 104 | GII.4[P16] (1043 NVRL) | 9.50 × 104 | GI.3[P3] (#2) ^ | 3.83 × 107 |
| GII.4[P16] (1036 NVRL) | 4.00 × 105 | 1.4 × 103 | GII.4 NO/Sydney (16-78)+ | 2.60 × 106 | GI.3[P3] (#3) ^ | 6.36 × 106 |
| GII.4[P16] (1041 NVRL) | 4.70 × 104 | 2.1 × 103 | GII.6 (15-65)+ | 3.10 × 104 | GI.3[P3] (461-1) ^ | 5.06 × 103 |
| GII.4[P16] (1046 NVRL) | 8.00 × 105 | 5.0 × 100 | GII.6[P7] (2014 vomit) * | 5.40 × 104 | GI.3[P3] (461-2) ^ | 6.38 × 105 |
| GII.4 Sydney (13-39)+ | 5.60 × 105 | 2.4 × 103 | GII.6[P7] EP3 * | 1.80 × 103 | GI.3[P3] (461-3) ^ | 2.16 × 105 |
| GII.4 Sydney (15-59)+ | 2.60 × 105 | 1.6 × 102 | GII.6[P7] EP4 * | 1.40 × 104 | GI * | 3.80 × 106 |
| GII.6[P7] (2014 stool) * | 1.60 × 105 | 4.2 × 102 | GII.6[P7] EP5 * | 3.20 × 105 | GI (1048 NVRL) | 3.64 × 105 |
| GII.6 (14-55)+ | 1.70 × 105 | 1.9 × 101 | GII.6[P7] EP6 * | 6.60 × 105 | GI (1049 NVRL) | 3.32 × 104 |
| GII.6 * | 7.64 × 104 | 8.0 × 100 | GII.1 SD (2016) * | 7.20 × 105 | GI (1052 NVRL) | 5.75 × 102 |
| GII.1 JW (2016) * | 4.10 × 105 | 8.4 × 101 | GII.7 (vomit) * | 1.50 × 105 | GI (1054 NVRL) | 9.62 × 106 |
| GI (1257 NVRL) | 2.12 × 106 | 8.0 × 100 | GII.17 (118-2)^ | 9.92 × 104 | GI (1057 NVRL) | 1.07 × 105 |
| GII (1038 NVRL) | 2.65 × 103 | GI (1249 NVRL) | 1.39 × 105 | |||
| GII (1040 NVRL) | 1.42 × 105 | GI (1252 NVRL) | 8.98 × 103 | |||
| Weigh 50 g of food and inoculate with 100 µL of 10% stool suspension (~5 × 107 gc) |
| Let air dry for 1 h at RT |
| Elute virus with 0.1 M Tris-HCl-0.05 M glycine-1% beef extract buffer, pH 9.2 with Polyvinylpyrrolidone (PVP), and pectinase on a shaking platform (30 min at RT) |
| Clarify eluate with centrifugation at 10,000× g at 4 °C |
| Concentrate the virus by ultracentrifugation at 170,000× g for 1 h at 4 °C |
| Resuspend pellet in 600 µL media; sonicate and clarify particles at 4500 rpm/5 min |
| Infect enteroids (100 µL/well; 1 h adsorption; 3 washes) and collect samples at times 0 and 3 dpi |
| Extract RNA from cells and supernatants with TrizolQuantify virus recovery from foods and replication by RT-qPCR with in-house full-length transcripts |
| Produce Matrix | Sample | Dilution | Input | 0 hpi | 72 hpi | Fold Increase (Replication +/−) |
|---|---|---|---|---|---|---|
| Frozen Raspberries | GII.6[P7] | None | Udt | 1.3 × 102 * | 4.2 × 104 | 3.2 × 102 (+) |
| Udt | Udt | Udt | 0 (−) | |||
| Udt | 3.2 × 102 | 6.7 × 104 | 2.1 × 102 (+) | |||
| Udt | 3.9 × 102 | 4.2 × 106 | 1.1 × 104 (+) | |||
| Udt | 5.9 × 102 | 1.9 × 107 | 3.3 × 104 (+) | |||
| 3.4 × 102 | 1.2 × 103 | 1.4 × 107 | 1.2 × 104 (+) | |||
| GII.4[P16] | Udt | 1.3 × 102 * | 2.6 × 105 | 2.0 × 103 (+) | ||
| Udt | 1.3 × 102 * | 2.0 × 105 | 1.5 × 103 (+) | |||
| Udt | 6.1 × 102 | 1.0 × 108 | 1.6 × 105 (+) | |||
| 1.2 × 103 | 7.9 × 102 | 3.7 × 104 | 4.6 × 101 (+) | |||
| Udt | 2.7 × 103 | 1.1 × 108 | 3.9 × 104 (+) | |||
| Udt | 4.4 × 103 | 1.3 × 108 | 3.1 × 104 (+) | |||
| 1.6 × 105 | 6.4 × 103 | 6.0 × 107 | 9.4 × 103 (+) | |||
| 2.4 × 105 | 7.0 × 103 | 1.0 × 105 | 1.4 × 101 (+) | |||
| Frozen Strawberries | GII.6[P7] | none | Udt | 4.0 × 102 | 2.6 × 107 | 6.5 × 104 (+) |
| Udt | 1.5 × 103 | 1.4 × 104 | 9.5 × 100 (+) | |||
| 2.5 × 102 | 3.7 × 102 | 2.0 × 106 | 5.4 × 103 (+) | |||
| 7.1 × 102 | Udt | Udt | 0 (−) | |||
| 1:3 | 1.8 × 103 | 2.8 × 102 | Udt | 0 (−) | ||
| 1:10 | Udt | Udt | Udt | 0 (−) | ||
| Udt | 1.3 × 102 * | Udt | 0 (−) | |||
| Udt | 2.5 × 102 | 5.1 × 102 | 2.0 × 100 (−) | |||
| 5.1 × 101 | Udt | Udt | 0 (−) | |||
| 1:20 | Udt | 2.0 × 102 | 4.0 × 102 | 2.0 × 100 (−) | ||
| 4.7 × 103 | 8.8 × 102 | Udt | 0 (−) | |||
| GII.4[P16] | none | 7.4 × 102 | 1.3 × 102 * | 5.9 × 107 | 4.6 × 105 (+) | |
| 1.1 × 103 | 1.3 × 102 * | 7.8 × 106 | 6.1 × 104 (+) | |||
| 5.4 × 103 | 6.8 × 102 | 1.0 × 107 | 1.5 × 104 (+) | |||
| 6.9 × 104 | 4.1 × 102 | 5.6 × 105 | 1.4 × 103 (+) | |||
| 1:3 | 1.0 × 102 | 3.2 × 102 | 2.7 × 105 | 8.4 × 102 (+) | ||
| 4.0 × 103 | 2.8 × 102 | 1.6 × 107 | 5.7 × 104 (+) | |||
| 1:10 | 6.3 × 101 | 1.3 × 102 * | 1.7 × 107 | 1.3 × 105 (+) | ||
| 1.8 × 102 | 1.3 × 102 * | 1.4 × 106 | 1.1 × 104 (+) | |||
| 2.3 × 102 | 1.3 × 102 * | 1.1 × 102 | 9.0 × 10−1 (−) | |||
| 3.5 × 102 | Udt | Udt | 0 (−) | |||
| 1:20 | 4.8 × 101 | 1.5 × 102 | 1.1 × 101 | 7.1 × 10−2 (−) | ||
| 4.0 × 102 | 1.4 × 102 | 1.9 × 105 | 1.4 × 103 (+) | |||
| Fresh Lettuce | GII.6[P7] | None | 2.6 × 104 | 1.9 × 102 | 1.5 × 102 | 7.9 × 10−1 (−) |
| 3.3 × 104 | 6.1 × 102 | 5.6 × 102 | 9.3 × 10−1 (−) | |||
| 5.6 × 104 | 3.1 × 102 | Udt | 0 (−) | |||
| 9.7 × 104 | 9.6 × 102 | 3.4 × 103 | 3.5 × 100 (−) | |||
| 2.0 × 105 | 1.4 × 103 | 6.5 × 102 | 4.7 × 10−1 (−) | |||
| 3.7 × 105 | 5.7 × 102 | 1.3 × 102 | 2.3 × 10−1 (−) | |||
| 5.1 × 105 | 3.0 × 102 | 6.4 × 103 | 2.1 × 101 (+) | |||
| 1:10 | 2.0 × 104 | 2.2 × 102 | Udt | 0 (−) | ||
| 3.1 × 104 | 2.9 × 102 | Udt | 0 (−) | |||
| 4.0 × 104 | 4.1 × 102 | 1.5 × 102 | 3.6 × 10−1 (−) | |||
| 5.0 × 104 | 2.7 × 102 | 4.2 × 102 | 1.5 × 100 (−) | |||
| GII.4[P16] | None | 6.1 × 104 | 6.3 × 102 | 4.4 × 105 | 7.0 × 102 (+) | |
| 7.2 × 104 | 2.0 × 102 | 2.3 × 105 | 1.2 × 103 (+) | |||
| 8.0 × 104 | 2.7 × 102 | 7.4 × 103 | 2.8 × 101 (+) | |||
| 1.3 × 105 | 1.3 × 102 * | 6.6 × 104 | 5.2 × 102 (+) | |||
| 1.6 × 105 | 5.1 × 102 | 7.8 × 105 | 1.5 × 103 (+) | |||
| 3.1 × 105 | 3.6 × 102 | Udt | 0 (−) | |||
| 3.4 × 105 | 4.0 × 103 | 1.1 × 106 | 2.7 × 102(+) | |||
| 1.2 × 106 | 9.0 × 102 | 1.8 × 107 | 1.9 × 104 (+) | |||
| 1:10 | 1.4 × 104 | 1.3 × 102 * | 1.3 × 102 | 1.0 × 100 (−) | ||
| 1.6 × 104 | 1.3 × 102 * | Udt | 0 (−) | |||
| 2.2 × 104 | 1.3 × 102 * | 9.4 × 104 | 7.3 × 102 (+) | |||
| 1.2 × 105 | 1.3 × 102 | 5.2 × 105 | 3.9 × 103 (+) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wales, S.Q.; Kulka, M.; Keinard, B.; Ngo, D.; Papafragkou, E. Use of Human Intestinal Enteroids for Recovery of Infectious Human Norovirus from Berries and Lettuce. Foods 2023, 12, 4286. https://doi.org/10.3390/foods12234286
Wales SQ, Kulka M, Keinard B, Ngo D, Papafragkou E. Use of Human Intestinal Enteroids for Recovery of Infectious Human Norovirus from Berries and Lettuce. Foods. 2023; 12(23):4286. https://doi.org/10.3390/foods12234286
Chicago/Turabian StyleWales, Samantha Q., Michael Kulka, Brianna Keinard, Diana Ngo, and Efstathia Papafragkou. 2023. "Use of Human Intestinal Enteroids for Recovery of Infectious Human Norovirus from Berries and Lettuce" Foods 12, no. 23: 4286. https://doi.org/10.3390/foods12234286
APA StyleWales, S. Q., Kulka, M., Keinard, B., Ngo, D., & Papafragkou, E. (2023). Use of Human Intestinal Enteroids for Recovery of Infectious Human Norovirus from Berries and Lettuce. Foods, 12(23), 4286. https://doi.org/10.3390/foods12234286