
Characterization of the Toxigenic Potential of Bacillus cereus sensu lato Isolated from Raw Berries and Their Products

Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Isolates
2.2. DNA Extraction
2.3. Primers and Multiplex PCR Reaction
2.4. Cytotoxic Activity
2.5. Statistical Analysis
3. Results and Discussion
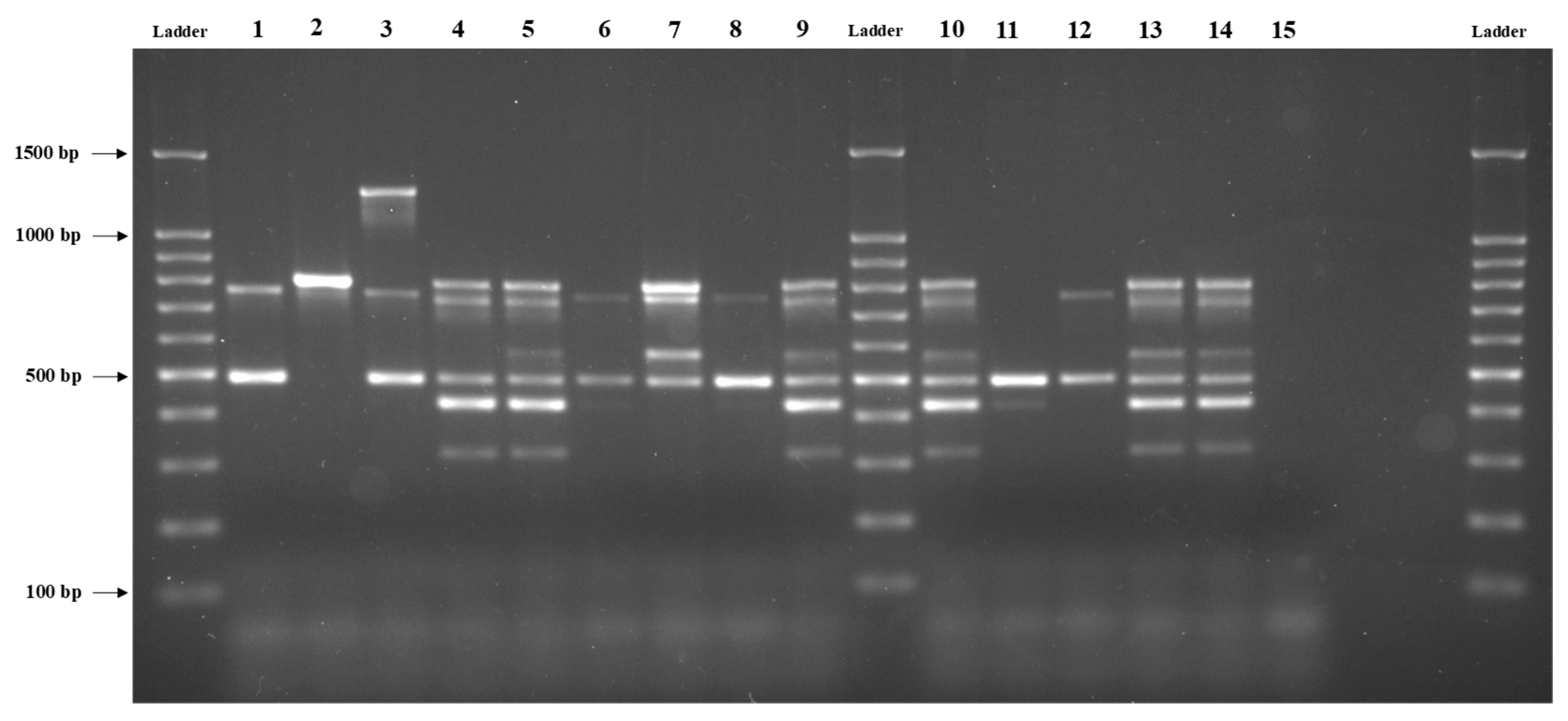
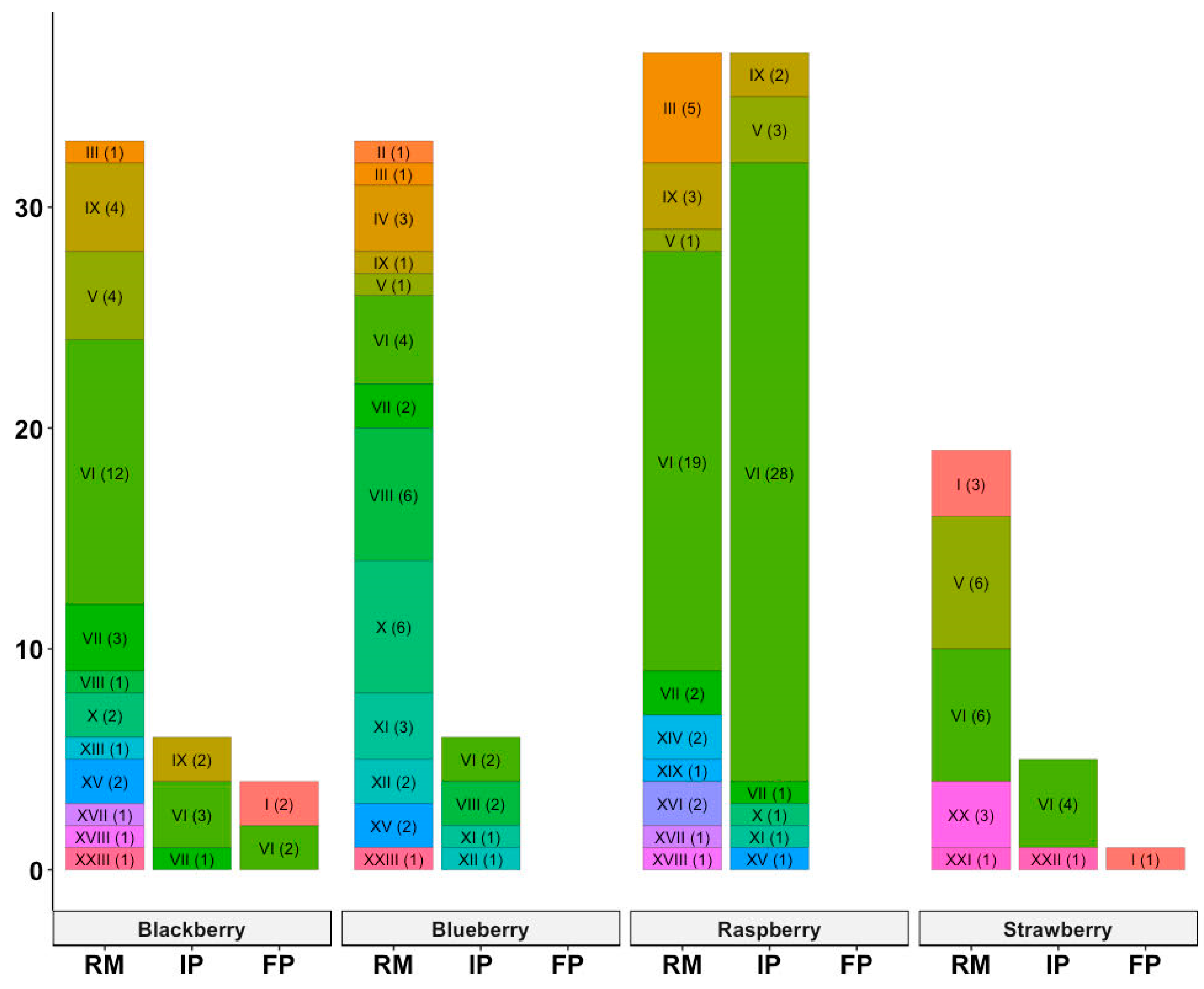
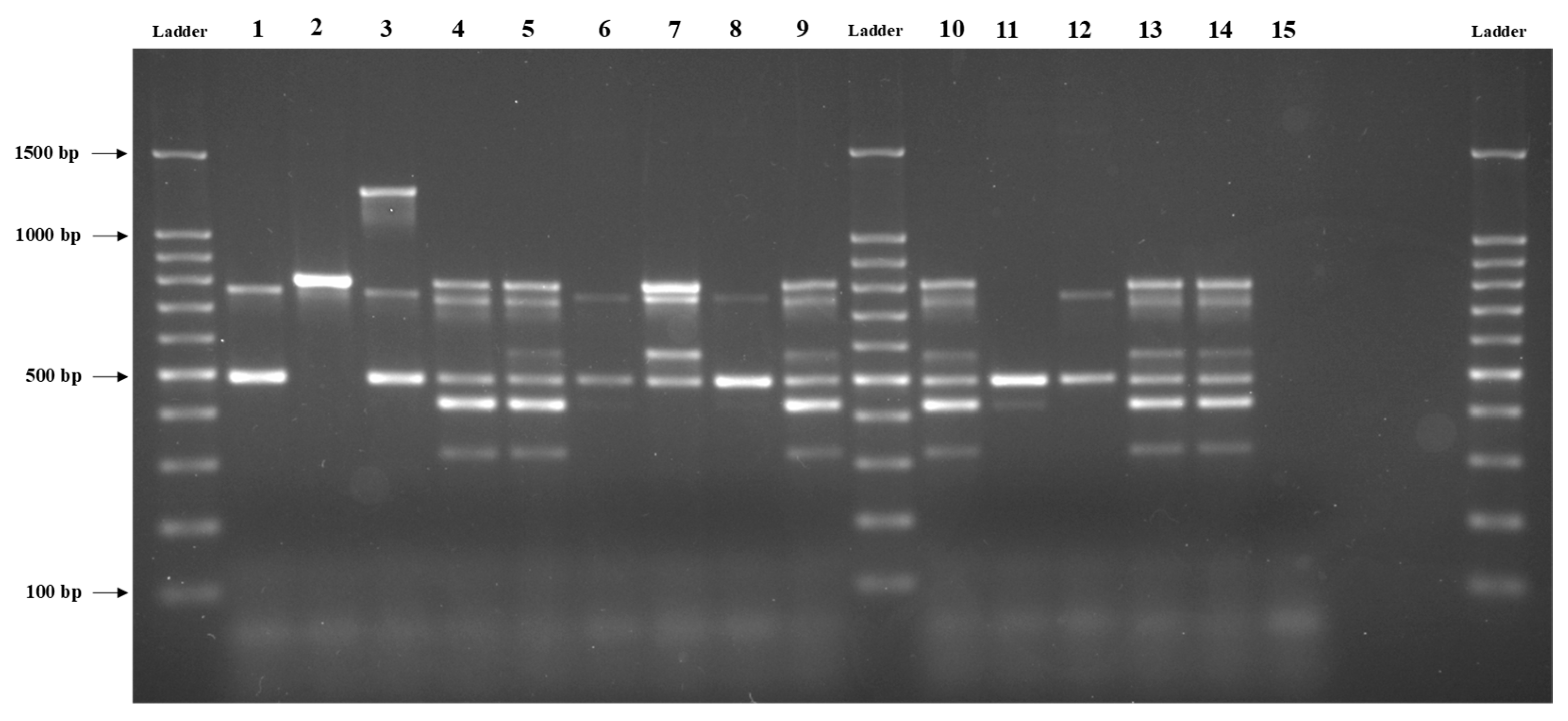
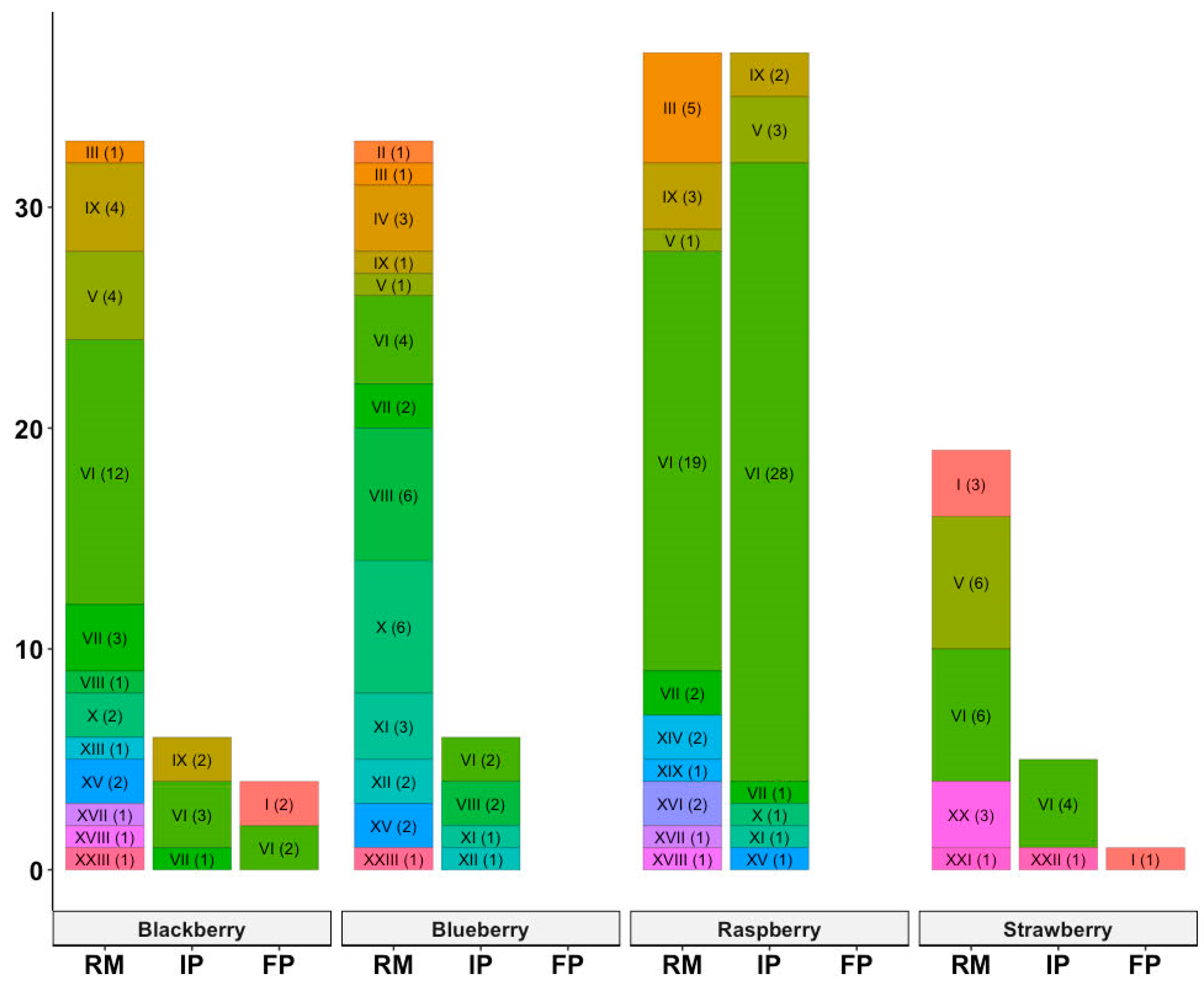
3.1. Distribution of Virulence Genes among B. cereus Isolates
3.2. Cytotoxic Activity of B. cereus Isolates
4. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Macieira, A.; Barbosa, J.; Teixeira, P. Food Safety in Local farming of fruits and vegetables. Int. J. Environ. Res. Public Health 2021, 18, 9733. [Google Scholar] [CrossRef]
- European Food Safety Authority; European Centre for Disease Prevention and Control. The European Union One Health 2021 Zoonoses Report. EFSA J. 2022, 20, e07666. [Google Scholar] [CrossRef]
- Palumbo, M.; Harris, L.J.; Danyluk, M.D. Outbreaks of foodborne illness associated with common berries, 1983 through May 2013. 2013. Available online: http://edis.ifas.ufl.edu/fs232 (accessed on 27 February 2023).
- Ceuppens, S.; Rajkovic, A.; Heyndrickx, M.; Tsilia, V.; Van De Wiele, T.; Boon, N.; Uyttendaele, M. Regulation of toxin production by Bacillus cereus and its food safety implications. Crit. Rev. Microbiol. 2011, 37, 188–213. [Google Scholar] [CrossRef] [PubMed]
- Logan, N.A.; Carman, J.A.; Melling, J.; Berkeley, R.C.W. Identification of Bacillus anthracis by API tests. J. Med. Microbiol. 1985, 20, 75–85. [Google Scholar] [CrossRef]
- Guinebretière, M.-H.; Auger, S.; Galleron, N.; Contzen, M.; De Sarrau, B.; De Buyser, M.-L.; Lamberet, G.; Fagerlund, A.; Granum, P.E.; Lereclus, D.; et al. Bacillus cytotoxicus sp. nov. is a novel thermotolerant species of the Bacillus cereus Group occasionally associated with food poisoning. Int. J. Syst. Evol. Microbiol. 2013, 63, 31–40. [Google Scholar] [CrossRef] [PubMed]
- Lechner, S.; Mayr, R.; Francis, K.P.; Pruss, B.M.; Kaplan, T.; Wießner-Gunkel, E.; Stewart, G.S.A.B.; Scherer, S. Bacillus weihenstephanensis sp. nov. is a new psychrotolerant species of the Bacillus cereus group. Int. J. Syst. Evol. Microbiol. 1998, 48, 1373–1382. [Google Scholar] [CrossRef]
- Nakamura, L.K. Bacillus pseudomycoides sp. nov. Int. J. Syst. Evol. Microbiol. 1998, 48, 1031–1035. [Google Scholar] [CrossRef]
- Jiménez, G.; Urdiain, M.; Cifuentes, A.; López-López, A.; Blanch, A.R.; Tamames, J.; Kämpfer, P.; Kolstø, A.-B.; Ramón, D.; Martínez, J.F.; et al. Description of Bacillus toyonensis sp. nov., a novel species of the Bacillus cereus group, and pairwise genome comparisons of the species of the group by means of ANI calculations. Syst. Appl. Microbiol. 2013, 36, 383–391. [Google Scholar] [CrossRef]
- Miller, R.A.; Beno, S.M.; Kent, D.J.; Carroll, L.M.; Martin, N.H.; Boor, K.J.; Kovac, J. Bacillus wiedmannii sp. nov., a psychrotolerant and cytotoxic Bacillus cereus group species isolated from dairy foods and dairy environments. Int. J. Syst. Evol. Microbiol. 2016, 66, 4744–4753. [Google Scholar] [CrossRef]
- Rasko, D.A.; Altherr, M.R.; Han, C.S.; Ravel, J. Genomics of the Bacillus cereus group of organisms. FEMS Microbiol. Rev. 2005, 29, 303–329. [Google Scholar] [CrossRef]
- Amor, M.G.-B.; Siala, M.; Zayani, M.; Grosset, N.; Smaoui, S.; Messadi-Akrout, F.; Baron, F.; Jan, S.; Gautier, M.; Gdoura, R. Isolation, identification, prevalence, and genetic diversity of Bacillus cereus group bacteria from different foodstuffs in Tunisia. Front. Microbiol. 2018, 9, 447. [Google Scholar] [CrossRef]
- Carroll, L.M.; Wiedmann, M.; Kovac, J. Proposal of a taxonomic nomenclature for the Bacillus cereus group which reconciles genomic definitions of bacterial species with clinical and industrial phenotypes. Mbio 2020, 11, e00034-20. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.; Liu, J.; Yin, Z.; Zhang, L. Phylogenetic and protein prediction analysis reveals the taxonomically diverse distribution of virulence factors in Bacillus cereus strains. PLoS ONE 2022, 17, e0262974. [Google Scholar] [CrossRef] [PubMed]
- Oliveira, M.; Rodrigues, C.M.; Teixeira, P. Microbiological quality of raw berries and their products: A focus on foodborne pathogens. Heliyon 2019, 5, e02992. [Google Scholar] [CrossRef] [PubMed]
- Park, K.M.; Jeong, M.; Park, K.J.; Koo, M. Prevalence, enterotoxin genes, and antibiotic resistance of Bacillus cereus isolated from raw vegetables in Korea. J. Food Prot. 2018, 81, 1590–1597. [Google Scholar] [CrossRef]
- Tango, C.N.; Wei, S.; Khan, I.; Hussain, M.S.; Kounkeu, P.F.N.; Park, J.H.; Kim, S.; Oh, D.H. microbiological quality and safety of fresh fruits and vegetables at retail levels in Korea. J. Food Sci. 2018, 83, 386–392. [Google Scholar] [CrossRef]
- Faille, C.; Membre, J.M.; Kubaczka, M.; Gavini, F. Altered Ability of Bacillus cereus Spores to grow under unfavorable conditions (presence of nisin, low temperature, acidic ph, presence of nacl) following heat treatment during sporulation. J. Food Prot. 2002, 65, 1930–1936. [Google Scholar] [CrossRef]
- Kotiranta, A.; Lounatmaa, K.; Haapasalo, M. Epidemiology and pathogenesis of Bacillus cereus infections. Microbes Infect. 2000, 2, 189–198. [Google Scholar] [CrossRef]
- Ehling-Schulz, M.; Fricker, M.; Scherer, S. Bacillus cereus, the causative agent of an emetic type of food-borne illness. Mol. Nutr. Food Res. 2004, 48, 479–487. [Google Scholar] [CrossRef]
- Schoeni, J.L.; Wong, A.C.L. Bacillus cereus food poisoning and its toxins. J. Food Prot. 2005, 68, 636–648. [Google Scholar] [CrossRef]
- Stenfors Arnesen, L.P.; Fagerlund, A.; Granum, P.E. From soil to gut: Bacillus cereus and its food poisoning toxins. FEMS Microbiol. Rev. 2008, 32, 579–606. [Google Scholar] [CrossRef] [PubMed]
- Wijnands, L.M.; Pielaat, A.; Dufrenne, J.B.; Zwietering, M.H.; van Leusden, F.M. Modelling the number of viable vegetative cells of Bacillus cereus passing through the stomach. J. Appl. Microbiol. 2009, 106, 258–267. [Google Scholar] [CrossRef]
- Dietrich, R.; Jessberger, N.; Ehling-Schulz, M.; Märtlbauer, E.; Granum, P.E. The food poisoning toxins of Bacillus cereus. Toxins 2021, 13, 98. [Google Scholar] [CrossRef] [PubMed]
- Ehling-Schulz, M.; Guinebretiere, M.-H.; Monthán, A.; Berge, O.; Fricker, M.; Svensson, B. Toxin gene profiling of enterotoxic and emetic Bacillus cereus. FEMS Microbiol. Lett. 2006, 260, 232–240. [Google Scholar] [CrossRef] [PubMed]
- Fagerlund, A.; Ween, O.; Lund, T.; Hardy, S.P.; Granum, P.E. Genetic and functional analysis of the cytK family of genes in Bacillus cereus. Microbiology 2004, 150, 2689–2697. [Google Scholar] [CrossRef]
- Yang, S.; Wang, Y.; Liu, Y.; Jia, K.; Zhang, Z.; Dong, Q. Cereulide and emetic Bacillus cereus: Characterizations, impacts and public precautions. Foods 2023, 12, 833. [Google Scholar] [CrossRef]
- Ehling-Schulz, M.; Vukov, N.; Schulz, A.; Shaheen, R.; Andersson, M.; Märtlbauer, E.; Scherer, S. identification and partial characterization of the nonribosomal peptide synthetase gene responsible for cereulide production in emetic Bacillus cereus. Appl. Environ. Microbiol. 2005, 71, 105–113. [Google Scholar] [CrossRef]
- Ehling-Schulz, M.; Frenzel, E.; Gohar, M. Food–bacteria interplay: Pathometabolism of emetic Bacillus cereus. Front. Microbiol. 2015, 6, 704. [Google Scholar] [CrossRef]
- Park, K.M.; Kim, H.J.; Park, K.J.; Koo, M. Susceptibility of emetic and enterotoxigenic Bacillus cereus grown at high temperature to disinfectants. Heliyon 2023, 9, e16863. [Google Scholar] [CrossRef]
- Bennett, S.D.; Walsh, K.A.; Gould, L.H. foodborne disease outbreaks caused by Bacillus cereus, Clostridium perfringens, and Staphylococcus aureus—United States, 1998–2008. Clin. Infect. Dis. 2013, 57, 425–433. [Google Scholar] [CrossRef]
- Beattie, S.H.; Williams, A.G. Detection of toxins. In Encyclopedia of Food Microbiology, 1st ed.; Robinson, R.K., Batt, C.A., Patel, P.D., Eds.; Academic Press: San Diego, CA, USA, 2000; pp. 141–158. [Google Scholar]
- Granum, P.E.; Lund, T. Bacillus cereus and its food poisoning toxins. FEMS Microbiol. Lett. 1997, 157, 223–228. [Google Scholar] [CrossRef]
- Jeßberger, N.; Dietrich, R.; Bock, S.; Didier, A.; Märtlbauer, E. Bacillus cereus enterotoxins act as major virulence factors and exhibit distinct cytotoxicity to different human cell lines. Toxicon 2014, 77, 49–57. [Google Scholar] [CrossRef] [PubMed]
- Ehling-Schulz, M.; Svensson, B.; Guinebretiere, M.-H.; Lindbäck, T.; Andersson, M.; Schulz, A.; Fricker, M.; Christiansson, A.; Granum, P.E.; Märtlbauer, E.; et al. Emetic toxin formation of Bacillus cereus is restricted to a single evolutionary lineage of closely related strains. Microbiology 2005, 151, 183–197. [Google Scholar] [CrossRef]
- Cui, Y.; Liu, Y.; Liu, X.; Xia, X.; Ding, S.; Zhu, K. Evaluation of the Toxicity and Toxicokinetics of Cereulide from an Emetic Bacillus cereus Strain of Milk Origin. Toxins 2016, 8, 156. [Google Scholar] [CrossRef] [PubMed]
- Forghani, F.; Kim, J.-B.; Oh, D.-H. Enterotoxigenic profiling of emetic toxin- and enterotoxin-producing Bacillus cereus, isolated from food, environmental, and clinical samples by Multiplex PCR. J. Food Sci. 2014, 79, M2288–M2293. [Google Scholar] [CrossRef] [PubMed]
- Guinebretière, M.-H.; Broussolle, V.; Nguyen-The, C. Enterotoxigenic profiles of food-poisoning and food-borne Bacillus cereus strains. J. Clin. Microbiol. 2002, 40, 3053–3056. [Google Scholar] [CrossRef]
- Kim, J.-M.; Forghani, F.; Kim, J.-B.; Park, Y.-B.; Park, M.-S.; Wang, J.; Park, J.H.; Oh, D.-H. Improved multiplex PCR assay for simultaneous detection of Bacillus cereus emetic and enterotoxic strains. Food Sci. Biotechnol. 2012, 21, 1439–1444. [Google Scholar] [CrossRef]
- Kim, M.-J.; Han, J.-K.; Park, J.-S.; Lee, J.-S.; Lee, S.-H.; Cho, J.-I.; Kim, K.-S. Various Enterotoxin and Other Virulence Factor Genes Widespread Among Bacillus cereus and Bacillus thuringiensis Strains. J. Microbiol. Biotechnol. 2015, 25, 872–879. [Google Scholar] [CrossRef]
- Amor, M.G.-B.; Jan, S.; Baron, F.; Grosset, N.; Culot, A.; Gdoura, R.; Gautier, M.; Techer, C. Toxigenic potential and antimicrobial susceptibility of Bacillus cereus group bacteria isolated from Tunisian foodstuffs. BMC Microbiol. 2019, 19, 196. [Google Scholar] [CrossRef]
- Fiedler, G.; Schneider, C.; Igbinosa, E.O.; Kabisch, J.; Brinks, E.; Becker, B.; Stoll, D.A.; Cho, G.-S.; Huch, M.; Franz, C.M.A.P. Antibiotics resistance and toxin profiles of Bacillus cereus-group isolates from fresh vegetables from German retail markets. BMC Microbiol. 2019, 19, 250. [Google Scholar] [CrossRef]
- Sánchez-Chica, J.; Correa, M.M.; Aceves-Diez, A.E.; Castañeda-Sandoval, L.M. Enterotoxin Gene Distribution and Genotypes of Bacillus cereus sensu lato Isolated from Cassava Starch. Toxins 2021, 13, 131. [Google Scholar] [CrossRef] [PubMed]
- Rajalingam, N.; Jung, J.; Seo, S.-M.; Jin, H.-S.; Kim, B.-E.; Jeong, M.-I.; Kim, D.; Ryu, J.-G.; Ryu, K.-Y.; Oh, K.K. Prevalence, distribution, enterotoxin profiles, antimicrobial resistance, and genetic diversity of Bacillus cereus group isolates from lettuce farms in Korea. Front. Microbiol. 2022, 13, 906040. [Google Scholar] [CrossRef]
- Chaves, J.Q.; Pires, E.S.; Vivoni, A.M. Genetic diversity, antimicrobial resistance and toxigenic profiles of Bacillus cereus isolated from food in Brazil over three decades. Int. J. Food Microbiol. 2011, 147, 12–16. [Google Scholar] [CrossRef] [PubMed]
- Guo, H.; Yu, P.; Yu, S.; Wang, J.; Zhang, J.; Zhang, Y.; Liao, X.; Wu, S.; Ye, Q.; Yang, X.; et al. Incidence, toxin gene profiling, antimicrobial susceptibility, and genetic diversity of Bacillus cereus isolated from quick-frozen food in China. LWT 2021, 140, 110824. [Google Scholar] [CrossRef]
- Kim, B.; Bang, J.; Kim, H.; Kim, Y.; Kim, B.-S.; Beuchat, L.R.; Ryu, J. Bacillus cereus and Bacillus thuringiensis spores in Korean rice: Prevalence and toxin production as affected by production area and degree of milling. Food Microbiol. 2014, 42, 89–94. [Google Scholar] [CrossRef]
- Ngamwongsatit, P.; Buasri, W.; Pianariyanon, P.; Pulsrikarn, C.; Ohba, M.; Assavanig, A.; Panbangred, W. Broad distribution of enterotoxin genes (hblCDA, nheABC, cytK, and entFM) among Bacillus thuringiensis and Bacillus cereus as shown by novel primers. Int. J. Food Microbiol. 2008, 121, 352–356. [Google Scholar] [CrossRef]
- Sánchez-Chica, J.; Correa, M.M.; Aceves-Diez, A.E.; Castañeda-Sandoval, L.M. Genetic and toxigenic diversity of Bacillus cereus group isolated from powdered foods. J. Food Sci. Technol. 2021, 58, 1892–1899. [Google Scholar] [CrossRef] [PubMed]
- Hansen, B.M.; Hendriksen, N.B. Detection of Enterotoxic Bacillus cereus and Bacillus thuringiensis Strains by PCR Analysis. Appl. Environ. Microbiol. 2001, 67, 185–189. [Google Scholar] [CrossRef]
- Thaenthanee, S.; Wong, A.C.L.; Panbangred, W. Phenotypic and genotypic comparisons reveal a broad distribution and heterogeneity of hemolysin BL genes among Bacillus cereus isolates. Int. J. Food Microbiol. 2005, 105, 203–212. [Google Scholar] [CrossRef]
- Tewari, A.; Singh, S.P.; Singh, R. Incidence and enterotoxigenic profile of Bacillus cereus in meat and meat products of Uttarakhand, India. J. Food Sci. Technol. 2015, 52, 1796–1801. [Google Scholar] [CrossRef]
- Castiaux, V.; Laloux, L.; Schneider, Y.-J.; Mahillon, J. Screening of cytotoxic B. cereus on differentiated Caco-2 cells and in co-culture with mucus-secreting (HT29-MTX) cells. Toxins 2016, 8, 320. [Google Scholar] [CrossRef]
- Lee, N.; Kim, M.-D.; Chang, H.-J.; Choi, S.-W.; Chun, H.S. Genetic diversity, antimicrobial resistance, toxin gene profiles, and toxin production ability of Bacillus cereus isolates from doenjang, a Korean fermented soybean paste. J. Food Saf. 2017, 37, e12363. [Google Scholar] [CrossRef]
- Wang, P.; Zhao, X.; Qu, T.; Liang, L.; Ji, Q.; Chen, Y. Insight into Bacillus cereus associated with infant foods in Beijing. Foods 2022, 11, 719. [Google Scholar] [CrossRef] [PubMed]
- Kindle, P.; Etter, D.; Stephan, R.; Johler, S. Population structure and toxin gene profiles of Bacillus cereus sensu lato isolated from flour products. FEMS Microbiol. Lett. 2019, 366, fnz240. [Google Scholar] [CrossRef]
- Aragon-Alegro, L.C.; Palcich, G.; Lopes, G.V.; Ribeiro, V.B.; Landgraf, M.; Destro, M.T. Enterotoxigenic and genetic profiles of Bacillus cereus strains of food origin in Brazil. J. Food Prot. 2008, 71, 2115–2118. [Google Scholar] [CrossRef]
- Chon, J.-W.; Kim, J.-H.; Lee, S.-J.; Hyeon, J.-Y.; Seo, K.-H. Toxin profile, antibiotic resistance, and phenotypic and molecular characterization of Bacillus cereus in Sunsik. Food Microbiol. 2012, 32, 217–222. [Google Scholar] [CrossRef]
- Agata, N.; Ohta, M.; Yokoyama, K. Production of Bacillus cereus emetic toxin (cereulide) in various foods. Int. J. Food Microbiol. 2002, 73, 23–27. [Google Scholar] [CrossRef] [PubMed]
- López, A.C.; Minnaard, J.; Pérez, P.F.; Alippi, A.M. A case of intoxication due to a highly cytotoxic Bacillus cereus strain isolated from cooked chicken. Food Microbiol. 2015, 46, 195–199. [Google Scholar] [CrossRef] [PubMed]
- Messelhäusser, U.; Frenzel, E.; Blöchinger, C.; Zucker, R.; Kämpf, P.; Ehling-Schulz, M. Emetic Bacillus cereus are more volatile than thought: Recent foodborne outbreaks and prevalence studies in Bavaria (2007–2013). BioMed Res. Int. 2014, 2014, 465603. [Google Scholar] [CrossRef]
- Jan, S.; Brunet, N.; Techer, C.; Le Maréchal, C.; Koné, A.; Grosset, N.; Cochet, M.; Gillard, A.; Gautier, M.; Puterflam, J.; et al. Biodiversity of psychrotrophic bacteria of the Bacillus cereus group collected on farm and in egg product industry. Food Microbiol. 2011, 28, 261–265. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
| Strawberries | Blueberries | Raspberries | Blackberries | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| RM | IP | FP | RM | IP | FP | RM | IP | FP | RM | IP | FP |
| 19 | 5 | 1 | 33 | 6 | 0 | 37 | 37 | 0 | 33 | 6 | 4 |
| Target Gene | Primer | Primer Sequence (5′-3′) | Amplicon Size (bp) | Reference |
|---|---|---|---|---|
| nheA | nheA 344 S | TACGCTAAGGAGGGGCA | 480 | [26] |
| nheA 843 A | GTTTTTATTGCTTCATCGGCT | |||
| nheB | nheB 1500 S | CTATCAGCACTTATGGCAG | 754 | [26] |
| nheB 2269 A | ACTCCTAGCGGTGTTCC | |||
| nheC | nheC 2820 S | CGGTAGTGATTGCTGGG | 564 | [26] |
| nheC 3401 A | CAGCATTCGTACTTGCCAA | |||
| hblA | HBLA1 | GTGCAGATGTTGATGCCGAT | 301 | [26] |
| HBLA2 | ATGCCACTGCGTGGACATAT | |||
| hblC | L2A | AATGGTCATCGGAACTCTAT | 731 | [26] |
| L2B | CTCGCTGTTCTGCTGTTAAT | |||
| hblD | L1A | AATCAAGAGCTGTCACGAAT | 411 | [26] |
| L1B | CACCAATTGACCATGCTAAT | |||
| cytK | CK-F-1859 | ACAGATATCGG(GT)CAAAATGC | 809 | [38] |
| CK-R-2668 | TCCAACCCAGTT(AT)(GC)CAGTTC | |||
| ces | cesF1 | GGTGACACATTATCATATAAGGTG | 1271 | [35] |
| cesR2 | GTAAGCGAACCTGTCTGTAACAACA |
| Target Gene | Amplification Conditions | Expected Fragment Length bp | |
|---|---|---|---|
| Cycles | Conditions | ||
| hblA, hblC, hblD, nheA, nheB, nheC, cytK, ces | 1 | Start cycle: 95 °C for 3 min | Between 300 and 1270 |
| 35 | Denaturation: 94 °C for 30 s | ||
| Annealing: 58 °C for 45 s | |||
| Extension: 72 °C for 1.50 min | |||
| 1 | Final extension: 72 °C for 5 min | ||
| ∞ | Storage: 4 °C | ||
| Toxin Profile Number | Virulence Genes | Number of Isolates (%) |
|---|---|---|
| I | ces nheA nheB | 6 (3.3) |
| II | cytK nheC nheA | 1 (0.6) |
| III | hblD hblA | 7 (3.9) |
| IV | nheA | 3 (1.7) |
| V | nheB hblC nheA hblD hblA | 15 (8.3) |
| VI | nheB hblC nheC nheA hblD hblA | 80 (44.2) |
| VII | nheB hblD hblA | 9 (5.0) |
| VIII | nheB nheA | 9 (5.0) |
| IX | nheB nheA hblD hblA | 12 (6.6) |
| X | nheA hblD | 9 (5.0) |
| XI | nheB hblC nheC nheA | 5 (2.8) |
| XII | nheB nheA hblD | 3 (1.7) |
| XIII | hblC nheA hblD hblA | 1 (0.6) |
| XIV | nheA hblD hblA | 2 (1.1) |
| XV | hlbD hblC hblA | 5 (2.8) |
| XVI | nheB hblC nheA hblD | 2 (1.1) |
| XVII | hblD | 2 (1.1) |
| XVIII | nheB hblD | 2 (1.1) |
| XIX | cytK | 1 (0.6) |
| XX | nheB nheC nheA hblD hblA | 3 (1.7) |
| XXI | nheB hblC hblD hblA | 1 (0.6) |
| XXII | nheB hblC nheA | 1 (0.6) |
| XXIII | hblC nheA hblD | 2 (1.1) |
| Virulence Genes | Number (%) of B. cereus | ||||
|---|---|---|---|---|---|
| Total (n = 181) | Blackberries (n = 43) | Raspberries (n = 74) | Strawberries (n = 25) | Blueberries (n = 39) | |
| ces | 6 (3.3) | 2 (4.6) | 0 | 4 (16) | 0 |
| hblA | 135 (74.6) | 35 (81.4) | 67 (90.5) | 20 (80) | 13 (33.3) |
| hblC | 112 (61.9) | 25 (58.1) | 55 (74.3) | 18 (72) | 14 (35.9) |
| hblD | 155 (85.6) | 40 (93.0) | 72 (97.3) | 20 (80) | 23 (59) |
| nheA | 154 (85.1) | 34 (79.1) | 62 (83.8) | 24 (96) | 34 (87.2) |
| nheB | 148 (81.8) | 35 (81.4) | 63 (85.1) | 25 (100) | 25 (64.1) |
| nheC | 89 (49.2) | 17 (39.5) | 48 (64.9) | 13 (52) | 11 (28.2) |
| cytK | 2 (1.1) | 0 | 1 (1.3) | 0 | 1 (2.6) |
| Cytotoxic Activity (%) * | ||
|---|---|---|
| Bacteria Isolates | 18 h | 5 Days |
| Bacillus cereus subsp. cytotoxicus NVH 391-98 | 64.4 ± 8.0 | 73.7 ± 12.0 |
| BactoFlavor ScarLet | 8.5 ± 0.5 | 1.7 ± 0.3 |
| B. cereus isolate profile II | 68.2 ± 11.2 | 66.4 ± 5.3 |
| B. cereus isolate profile XIX | 64.5 ± 9.1 | 64.0 ± 2.3 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Oliveira, M.; Carvalho, M.; Teixeira, P. Characterization of the Toxigenic Potential of Bacillus cereus sensu lato Isolated from Raw Berries and Their Products. Foods 2023, 12, 4021. https://doi.org/10.3390/foods12214021
Oliveira M, Carvalho M, Teixeira P. Characterization of the Toxigenic Potential of Bacillus cereus sensu lato Isolated from Raw Berries and Their Products. Foods. 2023; 12(21):4021. https://doi.org/10.3390/foods12214021
Chicago/Turabian StyleOliveira, Márcia, Marta Carvalho, and Paula Teixeira. 2023. "Characterization of the Toxigenic Potential of Bacillus cereus sensu lato Isolated from Raw Berries and Their Products" Foods 12, no. 21: 4021. https://doi.org/10.3390/foods12214021
APA StyleOliveira, M., Carvalho, M., & Teixeira, P. (2023). Characterization of the Toxigenic Potential of Bacillus cereus sensu lato Isolated from Raw Berries and Their Products. Foods, 12(21), 4021. https://doi.org/10.3390/foods12214021