

The Effect of Multistage Refinement on the Bio-Physico-Chemical Properties and Gel-Forming Ability of Fish Protein Isolates from Mackerel (Rastrelliger kanagurta)




Abstract
:1. Introduction
2. Materials and Methods
2.1. Fish Sample Preparation
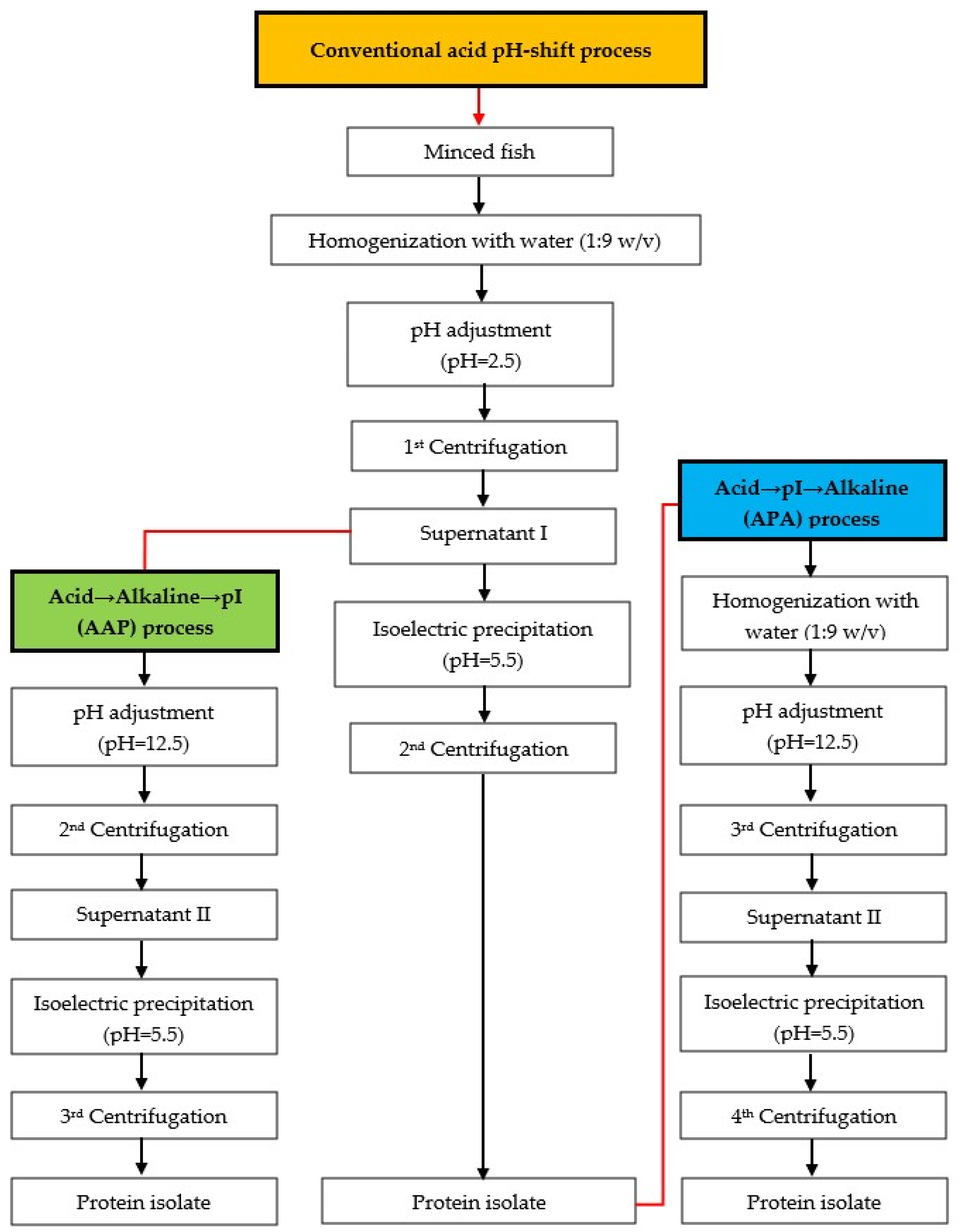
2.2. Protein Isolation Using Conventional and Modified Acid pH-Shift Processes
2.3. Characterization of Protein Isolates and Untreated Mince
2.3.1. Determination of Ca2+-ATPase Activity, Reactive Sulfhydryl (SH) Group, and Protein Surface Hydrophobicity
2.3.2. Determination of TCA-Soluble Peptide
2.3.3. Determination of Total Lipid Content and Thiobarbituric Acid Reactive Substances (TBARS)
2.3.4. Determination of Fishy Odor
2.3.5. Determination of Total Heme Protein Content, Myoglobin Absorption Spectra, and Color Characteristics
2.3.6. Sodium Dodecyl Sulfate-Polyacrylamide Gel Electrophoresis (SDS-PAGE)
2.4. Gelation
2.4.1. Dynamic Rheological Analysis
2.4.2. Gel Preparation
2.5. Gel Characterization of Protein Isolates and Untreated Mince
2.5.1. Breaking Force, Deformation, Gel Strength, and Texture Profile Analysis (TPA)
2.5.2. Expelled Moisture
2.5.3. Fourier Transform Infrared (FTIR) Spectra
2.5.4. Scanning Electron Microscopic (SEM) Images
2.6. Statistical Analysis
3. Results and Discussion
3.1. Bio-Physico-Chemical Properties of Protein Isolates
3.1.1. Biochemical Properties
3.1.2. TCA-Soluble Peptides
3.1.3. Lipid Reduction, Oxidation, and Fishy Odor Analysis
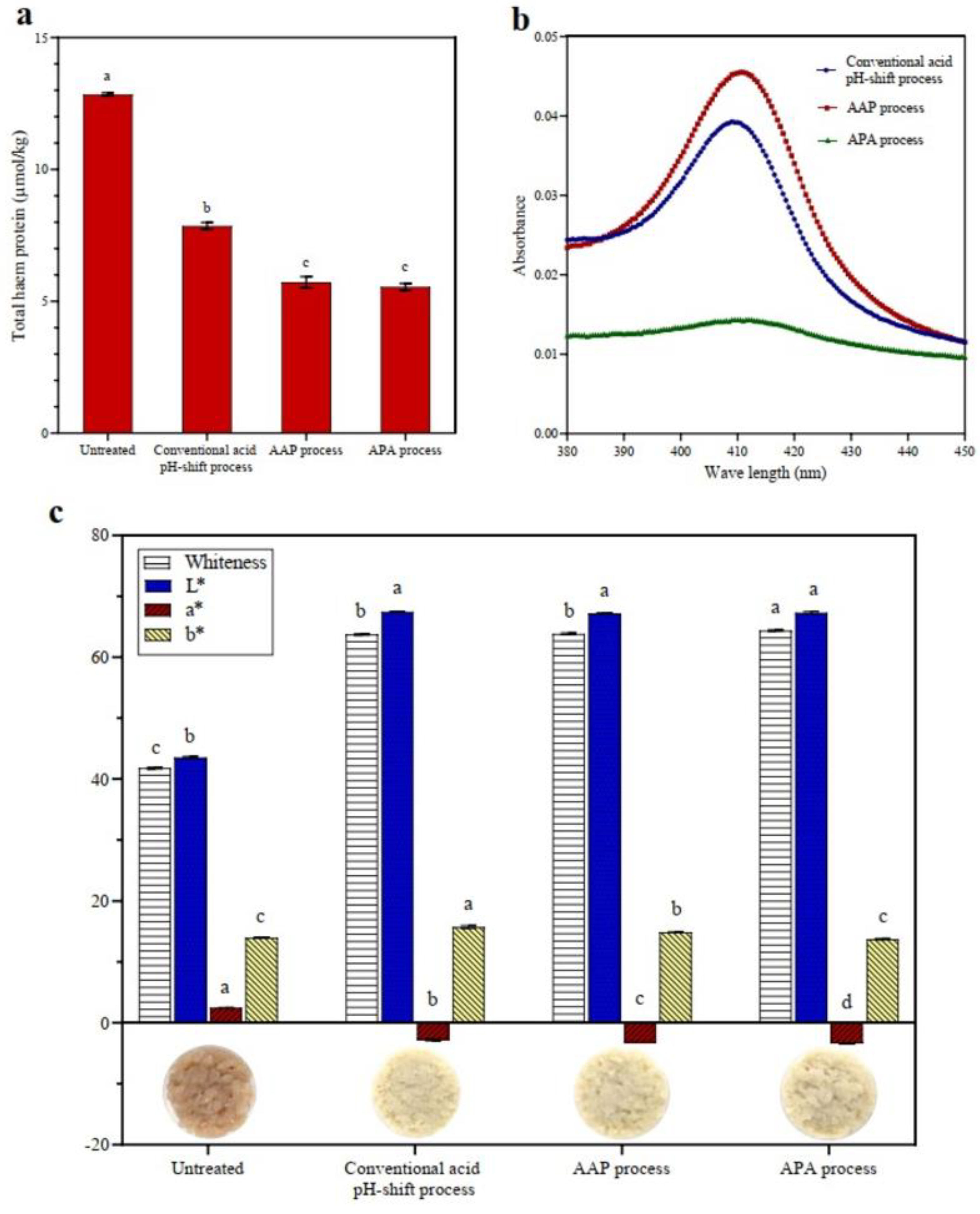
3.1.4. Heme Pigment Removal and Color Characteristics
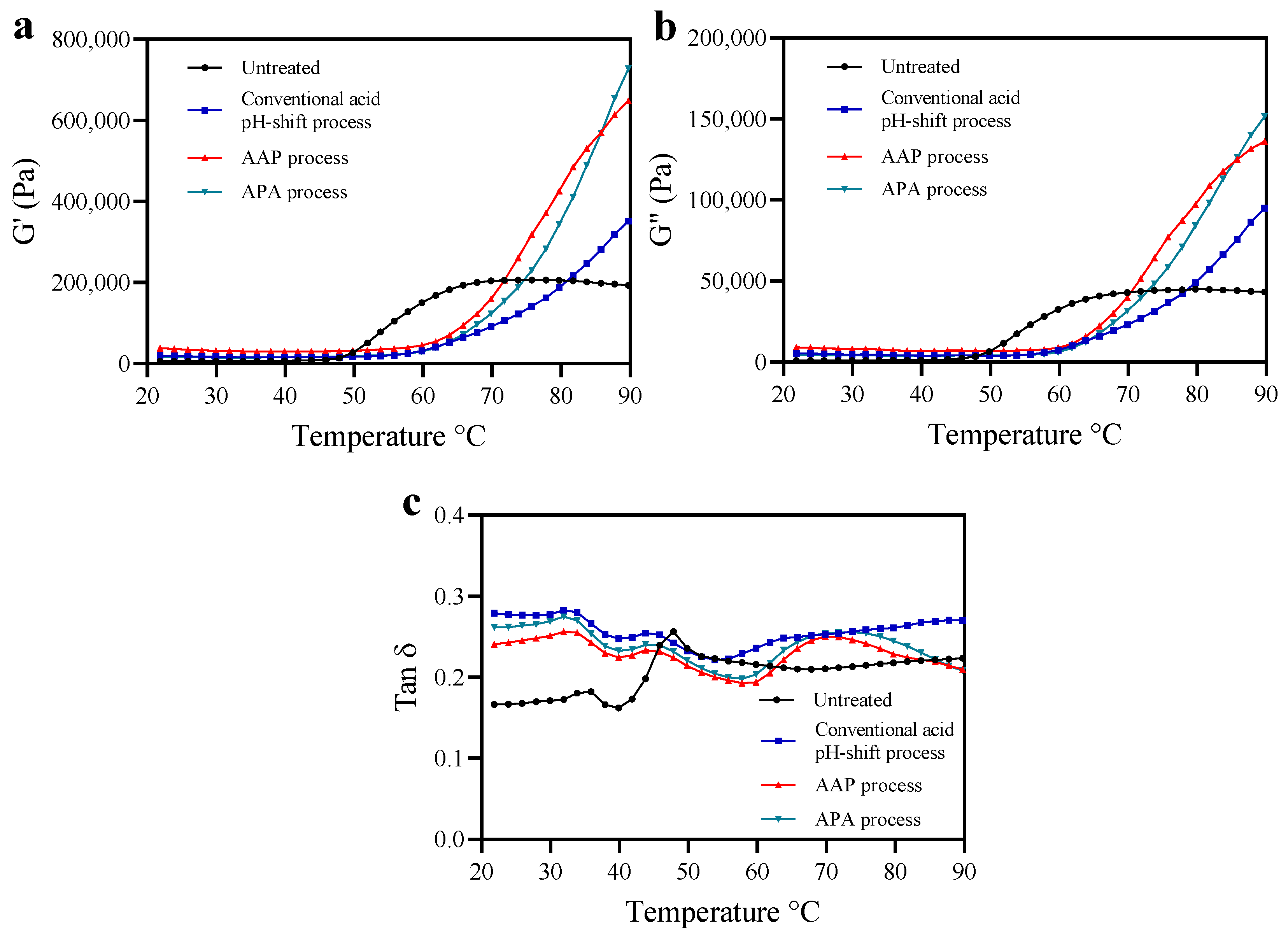
3.2. Oscillatory Dynamic Rheology
3.3. Gel Characteristics
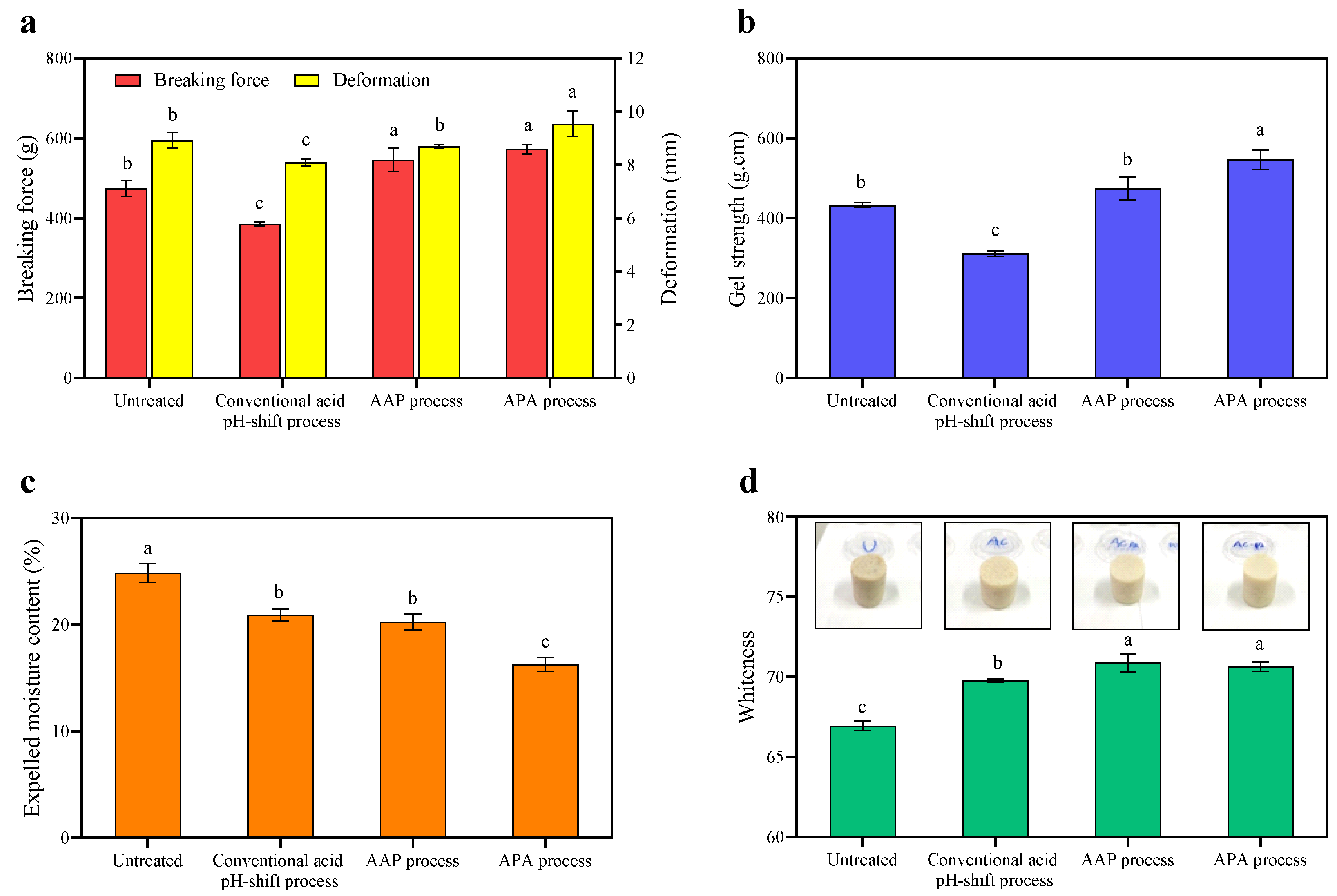
3.3.1. Breaking Force, Deformation, Gel Strength, and TPA
3.3.2. Expelled Moisture and Color Properties
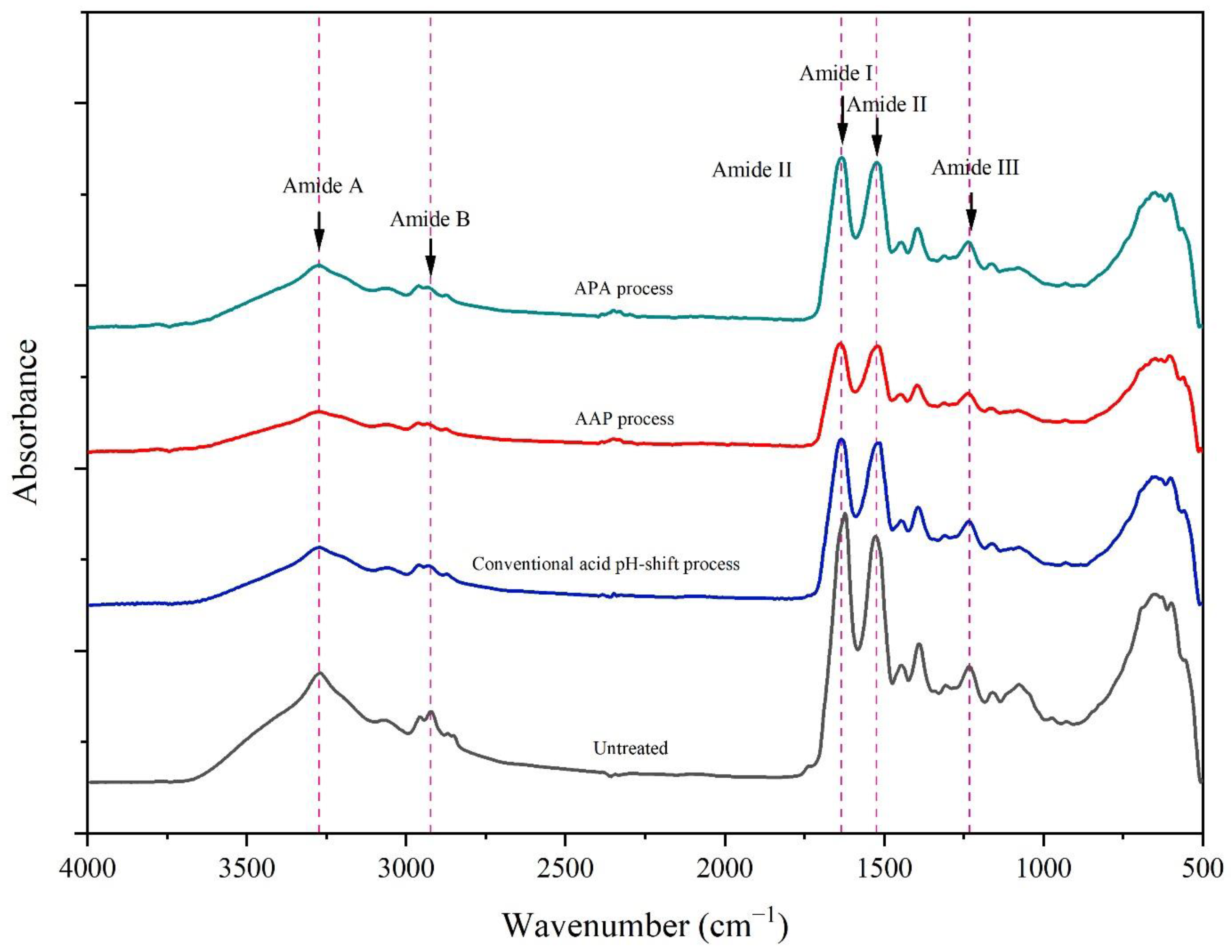
3.3.3. FTIR Spectra
3.3.4. Gel Microstructure
3.4. SDS-PAGE Patterns
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Zhong, C.; Feng, Y.; Xu, Y. Production of Fish Analogues from Plant Proteins: Potential Strategies, Challenges, and Outlook. Foods 2023, 12, 614. [Google Scholar] [CrossRef] [PubMed]
- Kakko, T.; Aitta, E.; Laaksonen, O.; Tolvanen, P.; Jokela, L.; Salmi, T.; Damerau, A.; Yang, B. Baltic herring (Clupea harengus membras) protein isolate produced using the pH-shift process and its application in food models. Food Res. Int. 2022, 158, 111578. [Google Scholar] [CrossRef]
- FAO. The State of World Fisheries and Aquaculture: Towards Blue Transformation; FAO: Rome, Italy, 2022. [Google Scholar] [CrossRef]
- Surasani, V.K.R. Acid and alkaline solubilization (pH shift) process: A better approach for the utilization of fish processing waste and by-products. Environ. Sci. Pollut. Res. 2018, 25, 18345–18363. [Google Scholar] [CrossRef] [PubMed]
- Department of Fisheries. Fisheries statistics of Thailand; Fishery Statistics Analysis and Research Group, Fisheries Development Policy and Strategy Division, Department of Fisheries: Bangkok, Thailand, 2020. [Google Scholar]
- Somjid, P.; Panpipat, W.; Cheong, L.Z.; Chaijan, M. Comparative effect of cricket protein powder and soy protein isolate on gel properties of Indian mackerel surimi. Foods 2022, 11, 3445. [Google Scholar] [CrossRef] [PubMed]
- Chaijan, M.; Benjakul, S.; Visessanguan, W.; Faustman, C. Characteristics and gel properties of muscles from sardine (Sardinella gibbosa) and mackerel (Rastrelliger kanagurta) caught in Thailand. Food Res. Int. 2004, 37, 1021–1030. [Google Scholar] [CrossRef]
- Zhang, K.; Li, N.; Wang, Z.; Feng, D.; Liu, X.; Zhou, D.; Li, D. Recent advances in the color of aquatic products: Evaluation methods, discoloration mechanism, and protection technologies. Food Chem. 2023, 434, 137495. [Google Scholar] [CrossRef]
- Phetsang, H.; Panpipat, W.; Undeland, I.; Panya, A.; Phonsatta, N.; Chaijan, M. Comparative quality and volatilomic characterisation of unwashed mince, surimi, and pH-shift-processed protein isolates from farm-raised hybrid catfish (Clarias macrocephalus × Clarias gariepinus). Food Chem. 2021, 364, 130365. [Google Scholar] [CrossRef]
- Tang, Z.X.; Ying, R.F.; Shi, L.E. Physicochemical and functional characteristics of proteins treated by a pH-shift process: A review. Int. J. Food Sci. Technol. 2021, 56, 515–529. [Google Scholar] [CrossRef]
- Kristinsson, H.G.; Ingadottir, B. Recovery and properties of muscle proteins extracted from tilapia (Oreochromis niloticus) light muscle by pH shift processing. J. Food Sci. 2006, 71, 132–141. [Google Scholar] [CrossRef]
- Matak, K.E.; Tahergorabi, R.; Jaczynski, J. A review: Protein isolates recovered by isoelectric solubilization/precipitation processing from muscle food by-products as a component of nutraceutical foods. Food Res. Int. 2015, 77, 697–703. [Google Scholar] [CrossRef]
- Abdollahi, M.; Axelsson, J.; Carlsson, N.G.; Nylund, G.M.; Albers, E.; Undeland, I. Effect of stabilization method and freeze/thaw-aided precipitation on structural and functional properties of proteins recovered from brown seaweed (Saccharina latissima). Food Hydrocoll. 2019, 96, 140–150. [Google Scholar] [CrossRef]
- Hinchcliffe, J.; Carlsson, N.G.; Jönsson, E.; Sundell, K.; Undeland, I. Aquafeed ingredient production from herring (Clupea harengus) by-products using pH-shift processing: Effect from by-product combinations, protein solubilization-pH and centrifugation force. Anim. Feed Sci. Technol. 2019, 247, 273–284. [Google Scholar] [CrossRef]
- Zhou, Y.; Liu, J.J.H.; Kang, Y.; Cui, H.; Yang, H. Effects of acid and alkaline treatments on physicochemical and rheological properties of tilapia surimi prepared by pH shift method during cold storage. Food Res. Int. 2021, 145, 110424. [Google Scholar] [CrossRef]
- Tian, Y.; Wang, W.; Yuan, C.; Zhang, L.; Liu, J.; Liu, J. Nutritional and digestive properties of protein isolates extracted from the muscle of the common carp using pH-shift processing. J. Food Process. Preserv. 2017, 41, 12847. [Google Scholar] [CrossRef] [PubMed]
- Tahergorabi, R.; Beamer, S.K.; Matak, K.E.; Jaczynski, J. Functional food products made from fish protein isolate recovered with isoelectric solubilization/precipitation. LWT 2012, 48, 89–95. [Google Scholar] [CrossRef]
- Pires, C.; Batista, I.; Fradinho, P.; Costa, S. Utilization of alkaline-recovered proteins from cape hake by-products in the preparation of frankfurter-type fish sausages. J. Aquat. Food Prod. Technol. 2009, 18, 170–190. [Google Scholar] [CrossRef]
- Shaviklo, G.R.; Arason, S.; Thorkelsson, G.; Sveinsdottir, K.; Martinsdottir, E. Sensory attributes of haddock balls affected by added fish protein isolate and frozen storage. J. Sens. Stud. 2010, 25, 316–331. [Google Scholar] [CrossRef]
- Shaviklo, A.R.; Moradinezhad, N.; Abolghasemi, S.J.; Motamedzadegan, A.; Kamali-Damavandi, N.; Rafipour, F. Product optimization of fish burger containing tuna protein isolates for better sensory quality and frozen storage stability. Turk. J. Fish. Aquat. Sci. 2016, 16, 923–933. [Google Scholar] [CrossRef] [PubMed]
- Nolsøe, H.; Undeland, I. The acid and alkaline solubilization process for the isolation of muscle proteins: State of the art. Food Bioprocess Technol. 2009, 2, 1–27. [Google Scholar] [CrossRef]
- Zhou, Y.; Yang, H. Effects of calcium ion on gel properties and gelation of tilapia (Oreochromis niloticus) protein isolates processed with pH shift method. Food Chem. 2019, 277, 327–335. [Google Scholar] [CrossRef]
- Panpipat, W.; Chaijan, M. Functional properties of pH-shifted protein isolates from bigeye snapper (Priacanthus tayenus) head by-product. Int. J. Food Prop. 2017, 20, 596–610. [Google Scholar] [CrossRef]
- Yongsawatdigul, J.; Park, J.W. Effects of alkali and acid solubilization on gelation characteristics of rockfish muscle proteins. J. Food Sci. 2004, 69, 499–505. [Google Scholar] [CrossRef]
- Zhang, J.; Abdollahi, M.; Alminger, M.; Undeland, I. Cross-processing herring and salmon co-products with agricultural and marine side-streams or seaweeds produces protein isolates more stable towards lipid oxidation. Food Chem. 2022, 382, 132314. [Google Scholar] [CrossRef] [PubMed]
- Abdollahi, M.; Olofsson, E.; Zhang, J.; Alminger, M.; Undeland, I. Minimizing lipid oxidation during pH-shift processing of fish by-products by cross-processing with lingonberry press cake, shrimp shells or brown seaweed. Food Chem. 2020, 327, 127078. [Google Scholar] [CrossRef] [PubMed]
- Abdollahi, M.; Marmon, S.; Chaijan, M.; Undeland, I. Tuning the pH-shift protein-isolation method for maximum hemoglobin-removal from blood rich fish muscle. Food Chem. 2016, 212, 213–224. [Google Scholar] [CrossRef]
- Marmon, S.K.; Undeland, I. Protein isolation from gutted herring (Clupea harengus) using pH-shift processes. J. Agric. Food Chem. 2010, 58, 10480–10486. [Google Scholar] [CrossRef]
- Benjakul, S.; Seymour, T.A.; Morrissey, M.T.; An, H. Physicochemical changes in Pacific whiting muscle proteins during iced storage. J. Food Sci. 1997, 62, 729–733. [Google Scholar] [CrossRef]
- Ellman, G.L. Tissue sulfhydryl groups. Arch. Biochem. Biophys. 1959, 82, 70–77. [Google Scholar] [CrossRef]
- Chelh, I.; Gatellier, P.; Santé-Lhoutellier, V. A simplified procedure for myofibril hydrophobicity determination. Meat Sci. 2006, 74, 681–683. [Google Scholar] [CrossRef]
- Panpipat, W.; Chaijan, M. Biochemical and physicochemical characteristics of protein isolates from bigeye snapper (Priacanthus tayenus) head by-product using pH shift method. Turk. J. Fish. Aquat. Sci. 2016, 16, 041–050. [Google Scholar]
- Bligh, E.G.; Dyer, W.J. A rapid method of total lipid extraction and purification. Can. J. Biochem. Physiol. 1959, 37, 911–917. [Google Scholar] [CrossRef]
- Panpipat, W.; Cheong, L.Z.; Chaijan, M. Impact of lecithin incorporation on gel properties of bigeye snapper (Priacanthus tayenus) surimi. Int. J. Food Sci. Technol. 2021, 56, 2481–2491. [Google Scholar] [CrossRef]
- Chaijan, M.; Undeland, I. Development of a new method for determination of total heme protein in fish muscle. Food Chem. 2015, 173, 1133–1141. [Google Scholar] [CrossRef] [PubMed]
- Somjid, P.; Panpipat, W.; Cheong, L.Z.; Chaijan, M. Reduced washing cycle for sustainable mackerel (Rastrelliger kanagurta) surimi production: Evaluation of bio-physico-chemical, rheological, and gel-forming properties. Foods 2021, 10, 2717. [Google Scholar] [CrossRef] [PubMed]
- Laemmli, U.K. Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature 1970, 227, 680–685. [Google Scholar] [CrossRef] [PubMed]
- Somjid, P.; Panpipat, W.; Chaijan, M. Carbonated water as a novel washing medium for mackerel (Auxis thazard) surimi production. J. Food Sci. Technol. 2017, 54, 3979–3988. [Google Scholar] [CrossRef]
- Tao, L.; Tian, L.; Zhang, X.; Huang, X.; Long, H.; Chang, F.; Li, T.; Li, S. Effects of γ-polyglutamic acid on the physicochemical properties and microstructure of grass carp (Ctenopharyngodon idellus) surimi during frozen storage. LWT 2020, 134, 109960. [Google Scholar] [CrossRef]
- Kim, Y.; Park, J.A.E.; Choi, Y. New approaches for the effective recovery of fish proteins and their physicochemical characteristics. Fish. Sci. 2003, 69, 1231–1239. [Google Scholar] [CrossRef]
- Zhu, X.; Zhu, M.; He, D.; Li, X.; Shi, L.; Wang, L.; Xu, J.; Zheng, Y.; Yin, T. Cryoprotective roles of carbox-ymethyl chitosan during the frozen storage of surimi: Protein structures, gel behaviors and edible qualities. Foods 2022, 11, 356. [Google Scholar] [CrossRef]
- Abdollahi, M.; Rezaei, M.; Jafarpour, A.; Undeland, I. Dynamic rheological, microstructural and physicochemical properties of blend fish protein recovered from kilka (Clupeonella cultriventris) and silver carp (Hypophthalmichthys molitrix) by the pH-shift process or washing-based technology. Food Chem. 2017, 229, 695–709. [Google Scholar] [CrossRef]
- Zhang, Y.; Chang, S.K. Microbial transglutaminase cross-linking enhances the textural and rheological properties of the surimi-like gels made from alkali-extracted protein isolate from catfish byproducts and the role of disulfide bonds in gelling. Foods 2023, 12, 2029. [Google Scholar] [CrossRef] [PubMed]
- Kristinsson, H.; Theodore, A.E.; Demir, N.; Ingadottir, B. A comparative study between acid- and alkali-aided processing and surimi processing for the recovery of proteins from Channel catfish muscle. J. Food Sci. 2005, 70, 298–306. [Google Scholar] [CrossRef]
- Thiansilakul, Y.; Benjakul, S.; Richards, M.P. Changes in heme proteins and lipids associated with off-odour of seabass (Lates calcarifer) and red tilapia (Oreochromis mossambicus × O. niloticus) during iced storage. Food Chem. 2010, 121, 1109–1119. [Google Scholar] [CrossRef]
- Murthy, L.N.; Phadke, G.G.; Jeyakumari, A.; Ravishankar, C.N. Effect of added calcium and heat setting on gel forming and functional properties of Sardinella fimbriata surimi. J. Food Sci. Technol. 2021, 58, 427–436. [Google Scholar] [CrossRef]
- Abdollahi, M.; Undeland, I. Physicochemical and gel-forming properties of protein isolated from salmon, cod and herring by-products using the pH-shift method. LWT 2019, 101, 678–684. [Google Scholar] [CrossRef]
- Chaijan, M.; Panpipat, W.; Benjakul, S. Physicochemical and gelling properties of short-bodied mackerel (Rastrelliger brachysoma) protein isolate prepared using alkaline-aided process. Food Bioprod. Process. 2010, 88, 174–180. [Google Scholar] [CrossRef]
- Surasani, V.K.R.; Tyagi, A.; Kudre, T. Recovery of proteins from rohu processing waste using pH shift method: Characterization of isolates. J. Aquat. Food Prod. 2017, 26, 356–365. [Google Scholar] [CrossRef]
- Zhou, X.; Chen, T.; Lin, H.; Chen, H.; Liu, J.; Lyu, F.; Ding, Y. Physicochemical properties and microstructure of surimi treated with egg white modified by tea polyphenols. Food Hydrocoll. 2019, 90, 82–89. [Google Scholar] [CrossRef]
- Yuan, L.; Yu, J.; Mu, J.; Shi, T.; Sun, Q.; Jin, W.; Gao, R. Effects of deacetylation of konjac glucomannan on the physico-chemical properties of surimi gels from silver carp (Hypophthalmichthys molitrix). RSC Adv. 2019, 9, 19828–19836. [Google Scholar] [CrossRef] [PubMed]
- Taherian, A.R.; Mondor, M.; Labranche, J.; Drolet, H.; Ippersiel, D.; Lamarche, F. Comparative study of functional properties of commercial and membrane processed yellow pea protein isolates. Food Res. Int. 2011, 44, 2505–2514. [Google Scholar] [CrossRef]
- Raghavan, S.; Kristinsson, H.G. Conformational and rheological changes in catfish myosin during alkali-induced unfolding and refolding. Food Chem. 2008, 107, 385–398. [Google Scholar] [CrossRef]
- Thawornchinsombut, S.; Park, J.W. Effect of NaCl on gelation characteristics of acid-and alkali-treated pacific whiting fish protein isolates. J. Food Biochem. 2017, 31, 427–455. [Google Scholar] [CrossRef]
- Priyadarshini, B.; Xavier, K.M.; Nayak, B.B.; Dhanapal, K.; Balange, A.K. Instrumental quality attributes of single washed surimi gels of tilapia: Effect of different washing media. LWT 2017, 86, 385–392. [Google Scholar] [CrossRef]
- Mozhaev, V.V.; Heremans, K.; Frank, J.; Masson, P.; Balny, C. High pressure effects on protein structure and function. Proteins Struct. Funct. Genet. 1996, 24, 81–91. [Google Scholar] [CrossRef]
- Sun, L.C.; Kaneko, K.; Okazaki, E.; Cao, M.J.; Ohwaki, H.; Weng, W.Y.; Osako, K. Comparative study of proteins recovered from whole North Pacific krill Euphausia pacifica by acidic and alkaline treatment during isoelectric solubilization/precipitation. Fish. Sci. 2013, 79, 537–546. [Google Scholar] [CrossRef]
- Azadian, M.; Moosavi-Nasab, M.; Abedi, E. Comparison of functional properties and SDS-PAGE patterns between fish protein isolate and surimi produced from silver carp. Eur. Food Res. Technol. 2012, 235, 83–90. [Google Scholar] [CrossRef]
- Kristinsson, H.G.; Liang, Y. Effect of pH-shift processing and surimi processing on Atlantic croaker (Micropogonias undulates) muscle proteins. J. Food Sci. 2006, 71, 304–312. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameter | Untreated | Conventional Acid pH-Shift Process | Modified Acid pH-Shift Process | |
|---|---|---|---|---|
| AAP Process | APA Process | |||
| Ca2+-ATPase activity (µmol/mg protein/min) | 2.10 ± 0.02 a | 1.48 ± 0.01 b | 1.53 ± 0.12 b | 1.00 ± 0.04 c |
| Reactive SH content (mol/108 g protein) | 5.35 ± 0.07 a | 2.87 ± 0.03 b | 2.58 ± 0.08 c | 2.57 ± 0.14 c |
| Surface hydrophobicity (BPB bound (µg)) | 51.95 ± 4.73 d | 105.49 ± 7.07 c | 115.10 ± 0.58 bc | 121.15 ± 2.54 a |
| TCA-soluble peptide content (µmol tyrosine/g sample) | 1.59 ± 0.27 a | 0.53 ± 0.05 b | 0.32 ± 0.07 c | 0.27 ± 0.06 c |
| Lipid reduction (%) | - | 80.82 ± 1.53 b | 82.32 ± 0.49 ab | 83.45 ± 1.13 a |
| TBARS (mg/kg) | 9.10 ± 0.38 a | 4.59 ± 0.09 b | 2.75 ± 0.13 c | 2.97 ± 0.10 c |
| Fishy odor score | 7.55 ± 2.56 a | 4.51 ± 1.45 b | 2.63 ± 1.39 c | 2.94 ± 1.65 c |
| Parameter | Untreated | Conventional Acid pH-Shift Process | Modified Acid pH-Shift Process | |
|---|---|---|---|---|
| AAP Process | APA Process | |||
| Hardness (N) | 19.28 ± 0.62 b | 18.36 ± 1.03 c | 19.98 ± 0.27 b | 27.57 ± 1.15 a |
| Springiness (cm) | 7.44 ± 0.02 b | 7.19 ± 0.04 d | 7.32 ± 0.05 c | 7.54 ± 0.04 a |
| Cohesiveness | 0.44 ± 0.03 b | 0.46 ± 0.02 b | 0.55 ± 0.00 a | 0.48 ± 0.01 b |
| Gumminess (N) | 8.41 ± 0.39 c | 8.39 ± 0.06 c | 10.96 ± 0.17 b | 13.33 ± 0.42 a |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Somjid, P.; Chaijan, M.; Rawdkuen, S.; Grossmann, L.; Panpipat, W. The Effect of Multistage Refinement on the Bio-Physico-Chemical Properties and Gel-Forming Ability of Fish Protein Isolates from Mackerel (Rastrelliger kanagurta). Foods 2023, 12, 3894. https://doi.org/10.3390/foods12213894
Somjid P, Chaijan M, Rawdkuen S, Grossmann L, Panpipat W. The Effect of Multistage Refinement on the Bio-Physico-Chemical Properties and Gel-Forming Ability of Fish Protein Isolates from Mackerel (Rastrelliger kanagurta). Foods. 2023; 12(21):3894. https://doi.org/10.3390/foods12213894
Chicago/Turabian StyleSomjid, Panumas, Manat Chaijan, Saroat Rawdkuen, Lutz Grossmann, and Worawan Panpipat. 2023. "The Effect of Multistage Refinement on the Bio-Physico-Chemical Properties and Gel-Forming Ability of Fish Protein Isolates from Mackerel (Rastrelliger kanagurta)" Foods 12, no. 21: 3894. https://doi.org/10.3390/foods12213894
APA StyleSomjid, P., Chaijan, M., Rawdkuen, S., Grossmann, L., & Panpipat, W. (2023). The Effect of Multistage Refinement on the Bio-Physico-Chemical Properties and Gel-Forming Ability of Fish Protein Isolates from Mackerel (Rastrelliger kanagurta). Foods, 12(21), 3894. https://doi.org/10.3390/foods12213894