
Virtual Screening of Soybean Protein Isolate-Binding Phytochemicals and Interaction Characterization

Abstract
1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Virtual Screening of SPI-Binding Phytochemicals
2.3. Validation of SPI-Phytochemicals Binding by Fluorescence Quenching Assays
2.4. Molecular Dynamics Simulation
2.5. Statistical Analysis
3. Results and Discussion
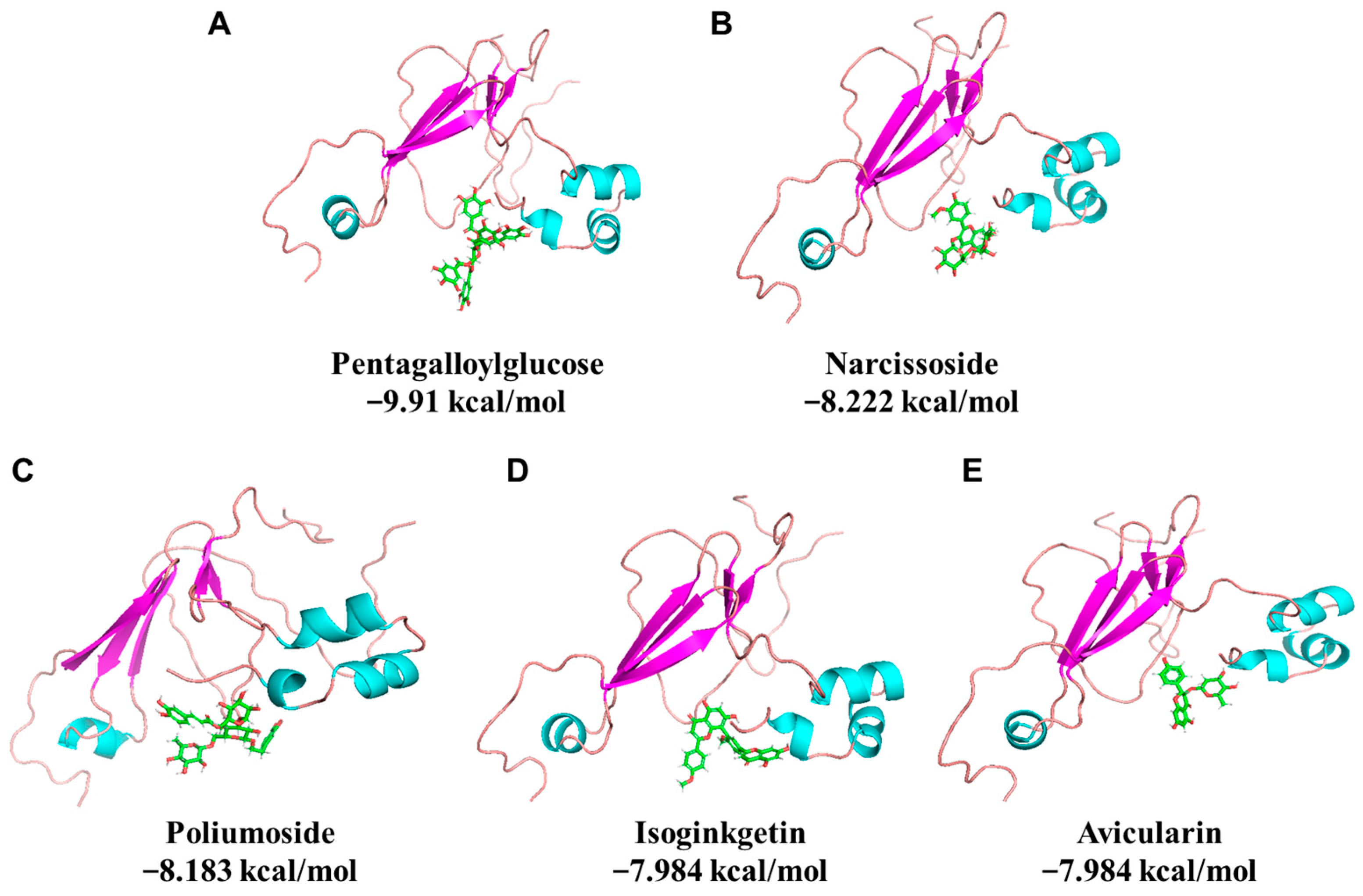
3.1. Virtual Screening of Phytochemicals against Soybean Protein
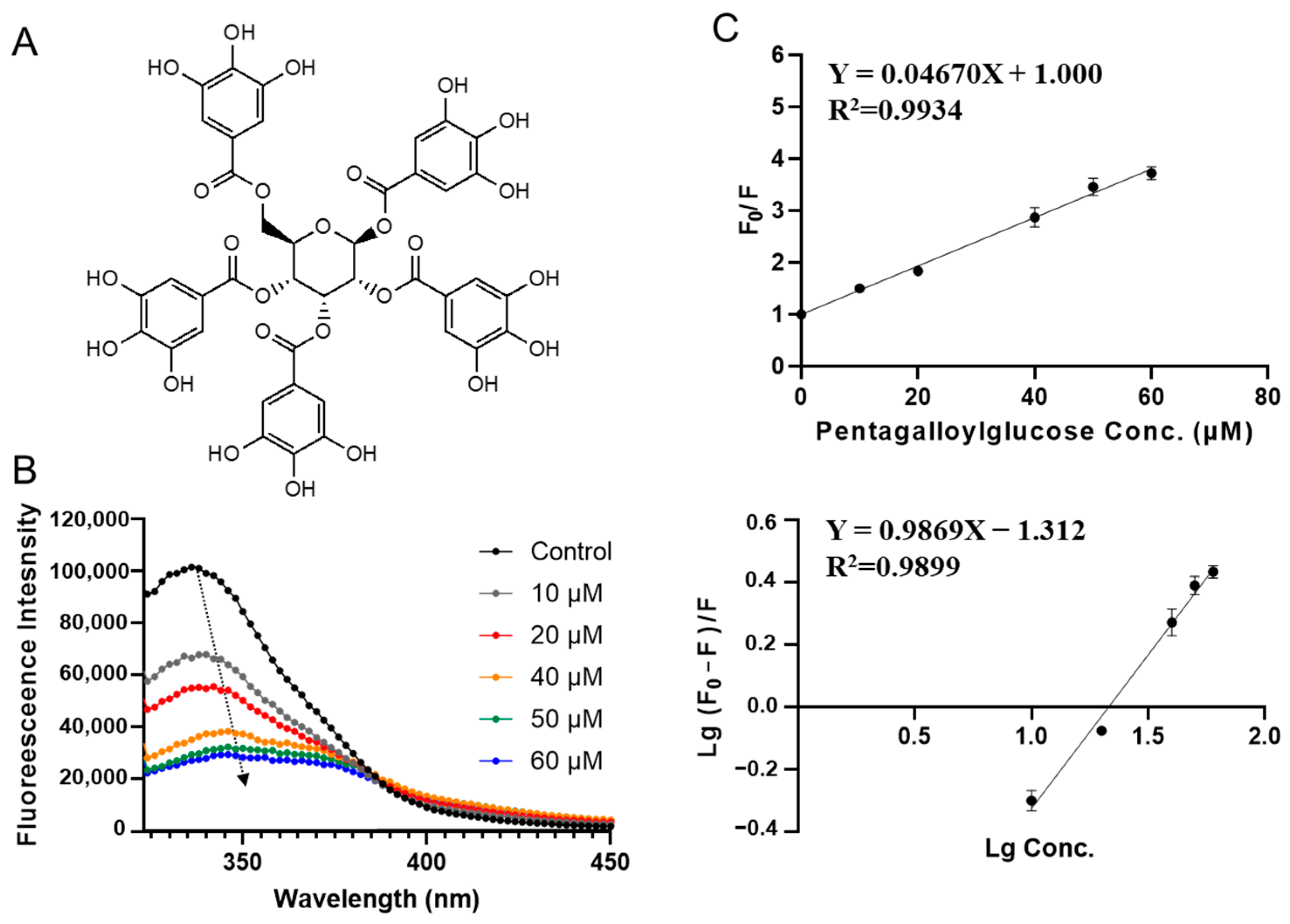
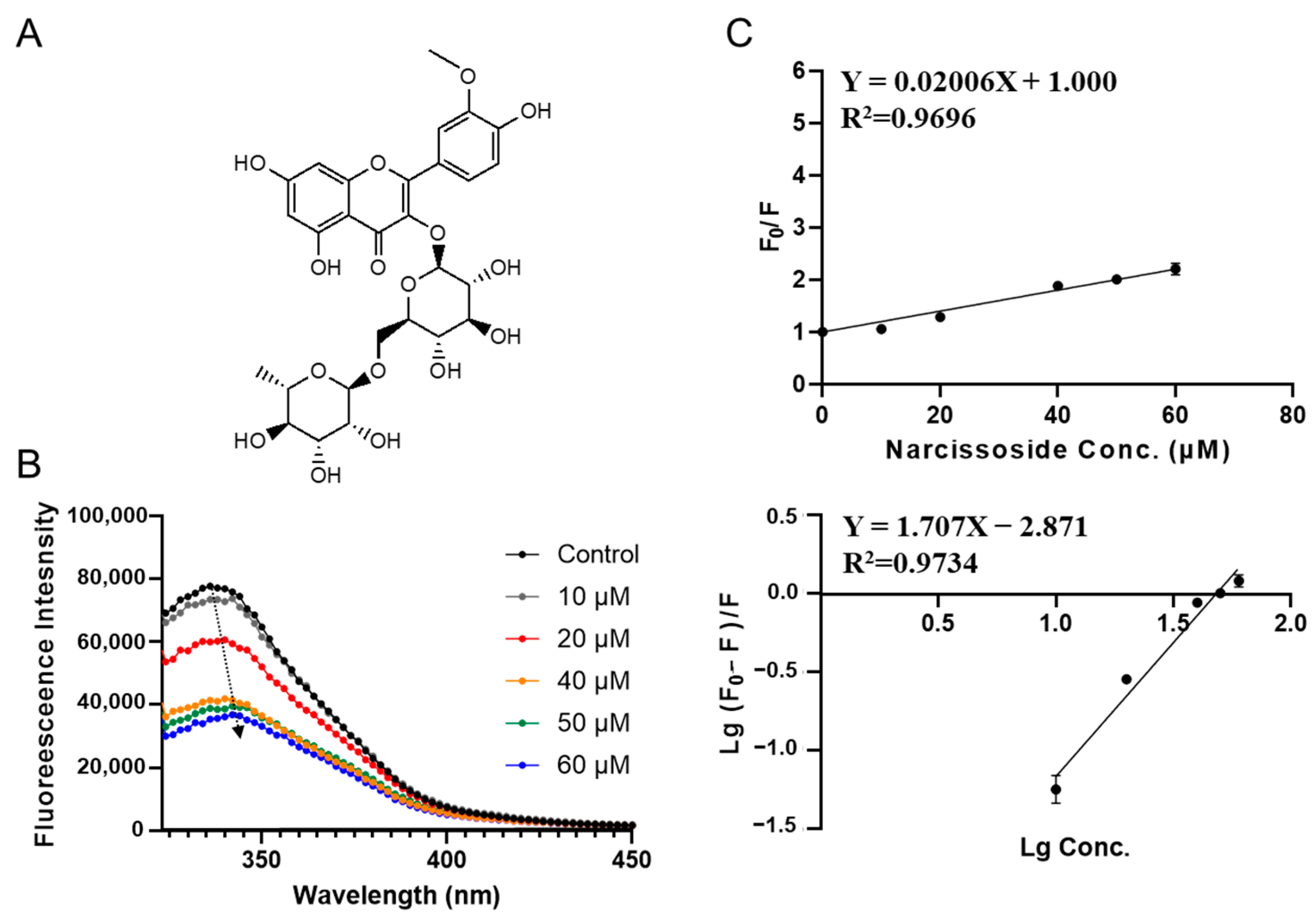
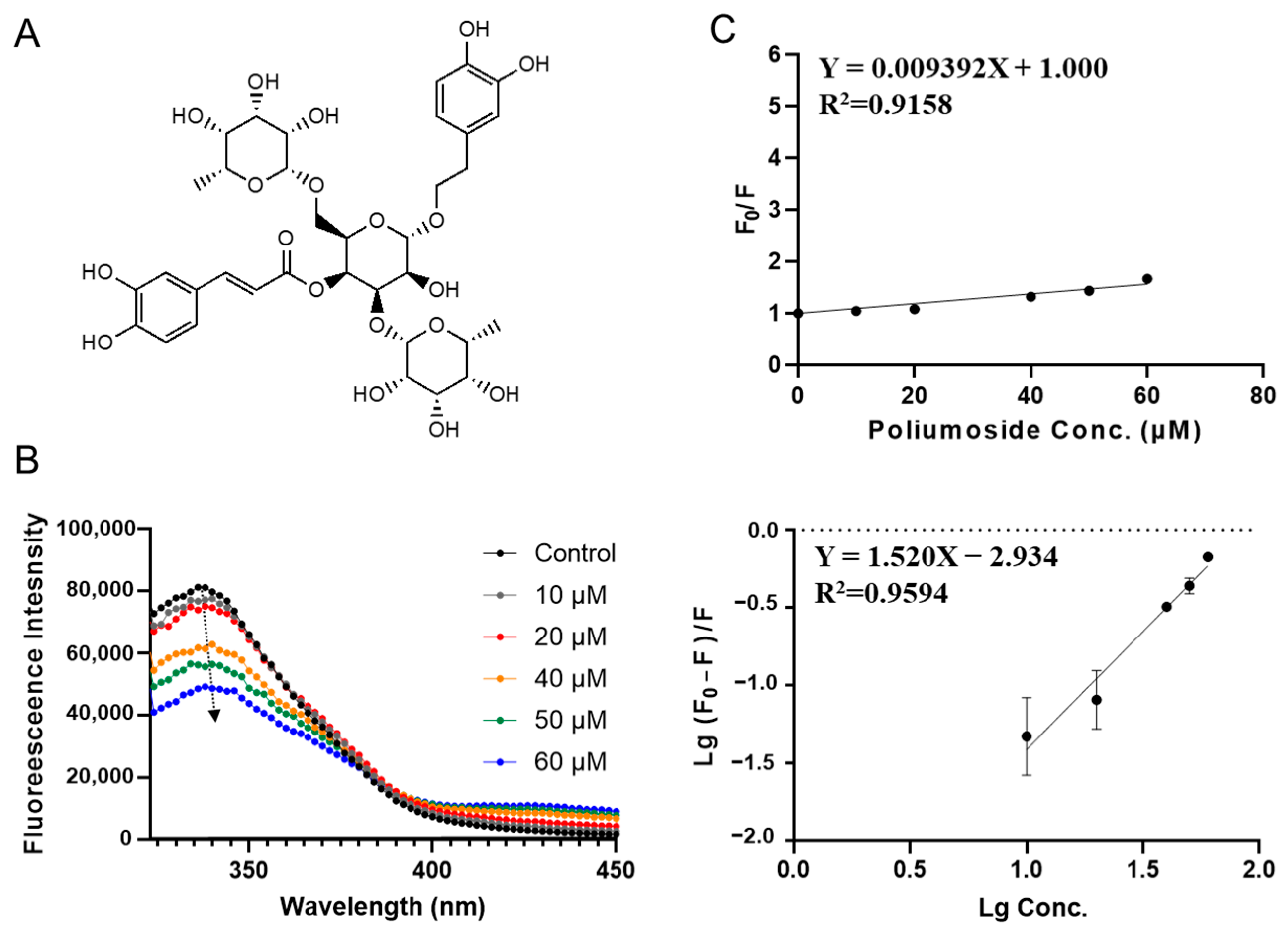
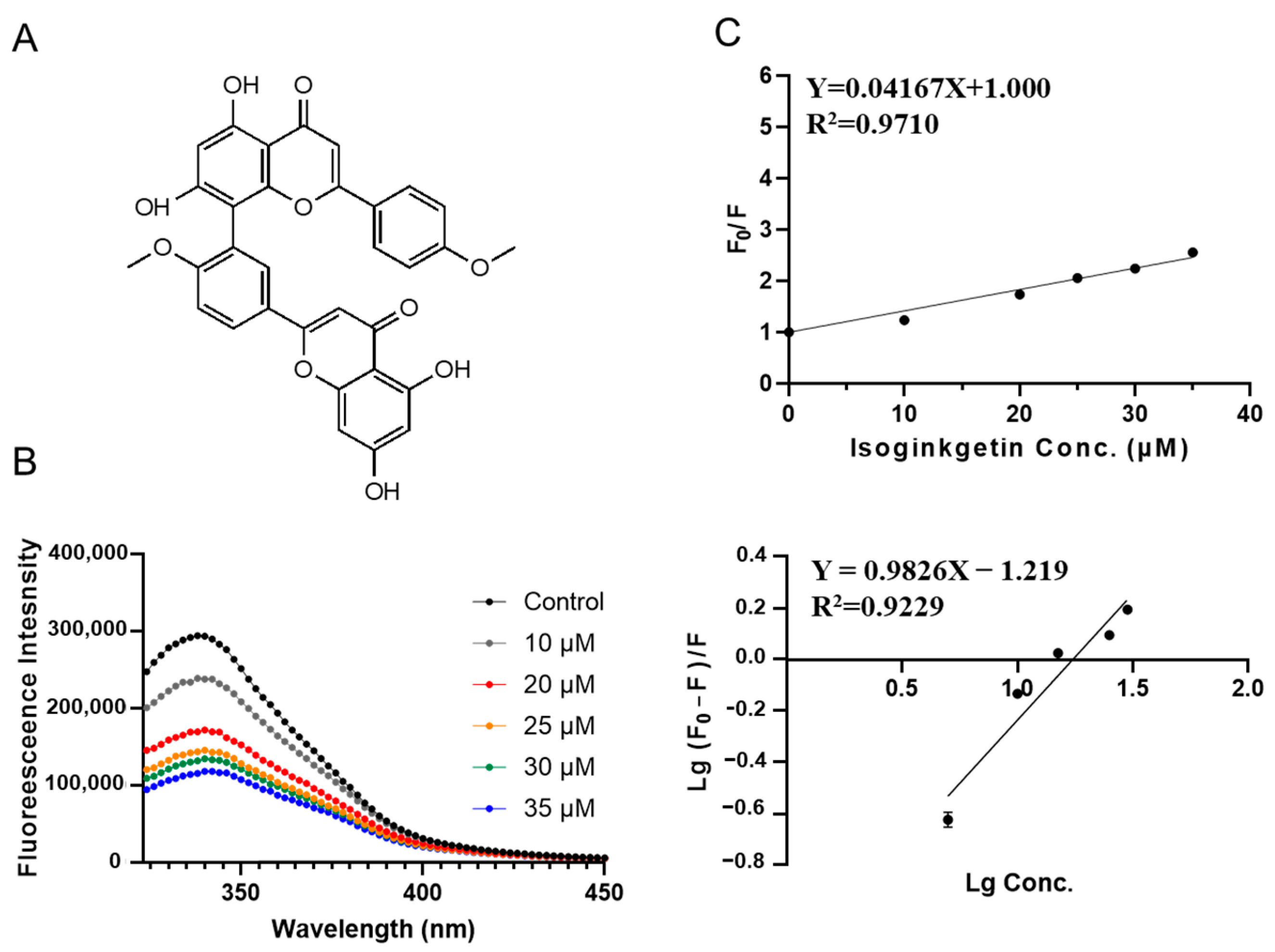
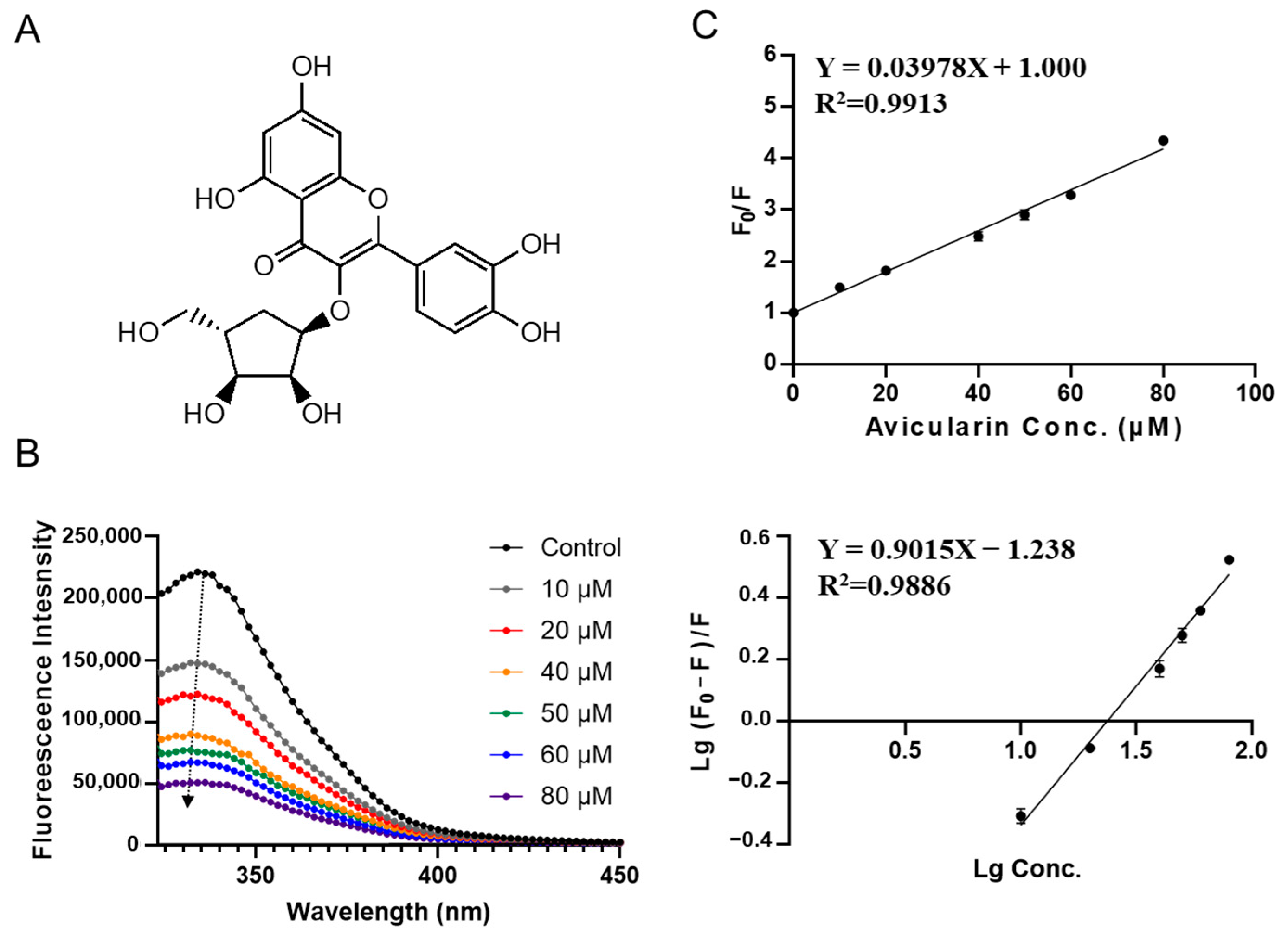
3.2. Characterization of SPI-Phytochemical Interaction by Fluorescence Quenching Assays
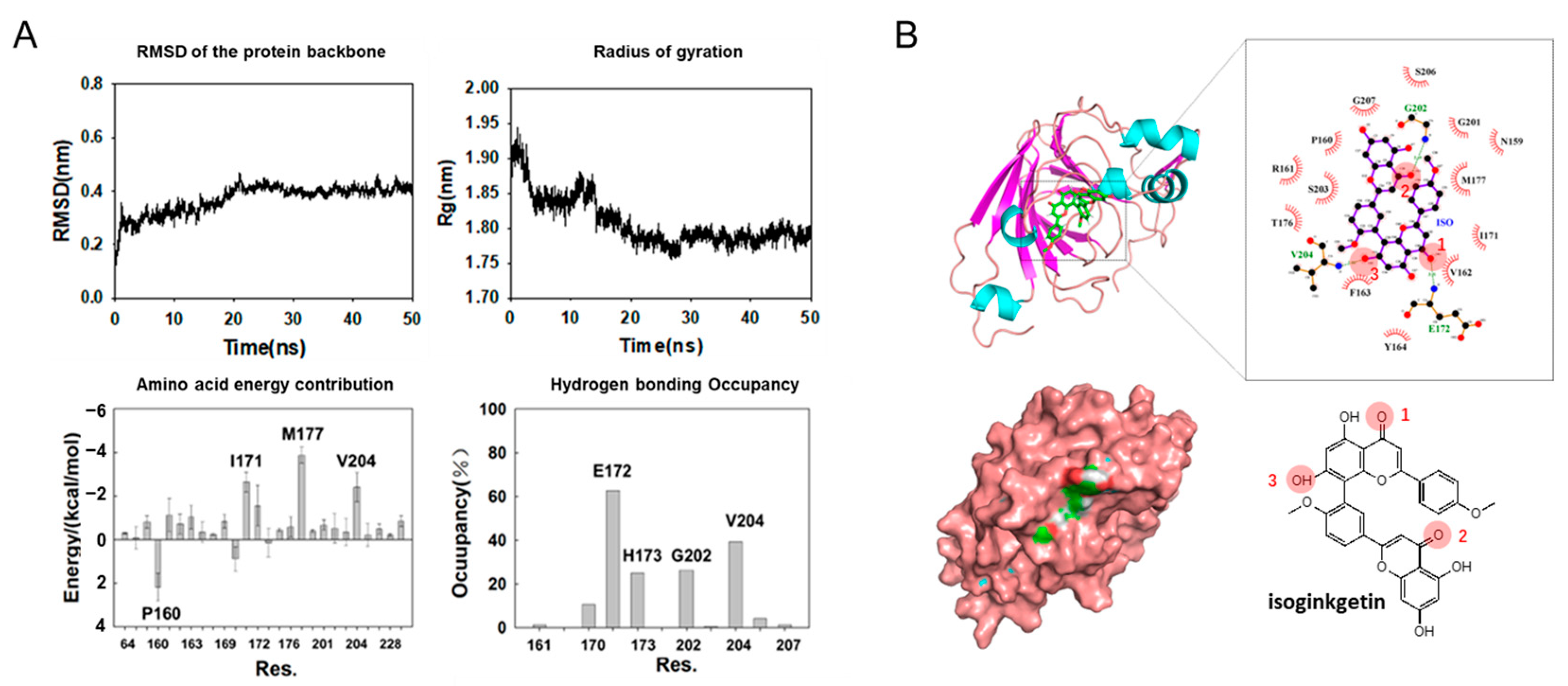
3.3. Molecular Dynamics Simulation on SPI-isoginkgetin interaction
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Sirtori, C.R.; Eberini, I.; Arnoldi, A. Hypocholesterolaemic effects of soya proteins: Results of recent studies are predictable from the Anderson meta-analysis data. Br. J. Nutr. 2007, 97, 816–822. [Google Scholar] [CrossRef]
- An, C.; Kuda, T.; Yazaki, T.; Takahashi, H.; Kimura, B. Caecal fermentation, putrefaction and microbiotas in rats fed milk casein, soy protein or fish meal. Appl. Microbiol. Biotechnol. 2013, 98, 2779–2787. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Li, H.; Xiao, L. Immune Modulation of Soybean Protein and Soybean Oligopeptides on Epidermal Trauma Infection in Aged Mice with Negative Nitrogen Balance. Food Sci. 2018, 39, 145–151. [Google Scholar] [CrossRef]
- Saz, J.M.; Marina, M.L. High performance liquid chromatography and capillary electrophoresis in the analysis of soybean proteins and peptides in foodstuffs. J. Sep. Sci. 2007, 30, 431–451. [Google Scholar] [CrossRef] [PubMed]
- Ao, L.; Liu, P.; Wu, A.; Zhao, J.; Hu, X. Characterization of Soybean Protein Isolate-Food Polyphenol Interaction via Virtual Screening and Experimental Studies. Foods 2021, 10, 2813. [Google Scholar] [CrossRef]
- You, Y.; Yang, L.; Chen, H.; Xiong, L.; Yang, F. Effects of (−)-Epigallocatechin-3-gallate on the Functional and Structural Properties of Soybean Protein Isolate. J. Agric. Food Chem. 2021, 69, 2306–2315. [Google Scholar] [CrossRef]
- Vesic, J.; Stambolic, I.; Apostolovic, D.; Milcic, M.; Stanic-Vucinic, D.; Velickovic, T.C. Complexes of green tea polyphenol, epigalocatechin-3-gallate, and 2S albumins of peanut. Food Chem. 2015, 185, 309–317. [Google Scholar] [CrossRef]
- Meng, Y.; Li, C. Conformational changes and functional properties of whey protein isolate-polyphenol complexes formed by non-covalent interaction. Food Chem. 2021, 364, 129622. [Google Scholar] [CrossRef]
- Cheng, J.; Zhu, M.; Liu, X. Insight into the conformational and functional properties of myofibrillar protein modified by mulberry polyphenols. Food Chem. 2019, 308, 125592. [Google Scholar] [CrossRef]
- Ye, J.; Deng, L.; Wang, Y.; McClements, D.J.; Luo, S.; Liu, C. Impact of rutin on the foaming properties of soybean protein: Formation and characterization of flavonoid-protein complexes. Food Chem. 2021, 362, 130238. [Google Scholar] [CrossRef]
- Jia, Z.; Zheng, M.; Tao, F.; Chen, W.; Huang, G.; Jiang, J. Effect of covalent modification by (−)-epigallocatechin-3-gallate on physicochemical and functional properties of whey protein isolate. LWT 2016, 66, 305–310. [Google Scholar] [CrossRef]
- Yildirim-Elikoglu, S.; Erdem, Y.K. Interactions between milk proteins and polyphenols: Binding mechanisms, related changes, and the future trends in the dairy industry. Food Rev. Int. 2017, 34, 665–697. [Google Scholar] [CrossRef]
- Su, J.; Guo, Q.; Chen, Y.; Mao, L.; Gao, Y.; Yuan, F. Utilization of β-lactoglobulin- (−)-Epigallocatechin- 3-gallate(EGCG) composite colloidal nanoparticles as stabilizers for lutein pickering emulsion. Food Hydrocoll. 2019, 98, 105293. [Google Scholar] [CrossRef]
- Xu, J.; Guo, S.; Li, X.; Jiang, S.; Zhong, X.; Zheng, Z. Gel properties of transglutaminase-induced soy protein isolate–polyphenol complex: Influence of epigallocatechin-3-gallate. J. Sci. Food Agric. 2021, 101, 3870–3879. [Google Scholar] [CrossRef] [PubMed]
- Hassan, S.; Azim, S. A Therapeutic Connection between Dietary Phytochemicals and ATP Synthase. Curr. Med. Chem. 2017, 24, 3894–3906. [Google Scholar] [CrossRef]
- Shahidi, F.; Pan, Y. Influence of food matrix and food processing on the chemical interaction and bioaccessibility of dietary phytochemicals: A review. Crit. Rev. Food Sci. Nutr. 2021, 62, 6421–6445. [Google Scholar] [CrossRef] [PubMed]
- Zembyla, M.; Murray, B.S.; Radford, S.J.; Sarkar, A. Water-in-oil Pickering emulsions stabilized by an interfacial complex of water-insoluble polyphenol crystals and protein. J. Colloid Interface Sci. 2019, 548, 88–99. [Google Scholar] [CrossRef]
- Le Bourvellec, C.; Renard, C. Interactions between Polyphenols and Macromolecules: Quantification Methods and Mechanisms. Crit. Rev. Food Sci. Nutr. 2012, 52, 213–248. [Google Scholar] [CrossRef]
- Müller, L.; Caris-Veyrat, C.; Lowe, G.; Böhm, V. Lycopene and Its Antioxidant Role in the Prevention of Cardiovascular Diseases—A Critical Review. Crit. Rev. Food Sci. Nutr. 2014, 56, 1868–1879. [Google Scholar] [CrossRef]
- Panigrahi, S.K.; Mishra, A.K. Inner filter effect in fluorescence spectroscopy: As a problem and as a solution. J. Photochem. Photobiol. C Photochem. Rev. 2019, 41, 100318. [Google Scholar] [CrossRef]
- Van Der Spoel, D.; Lindahl, E.; Hess, B.; Groenhof, G.; Mark, A.E.; Berendsen, H.J.C. GROMACS: Fast, flexible, and free. J. Comput. Chem. 2005, 26, 1701–1718. [Google Scholar] [CrossRef] [PubMed]
- Neese, F. Software update: The ORCA program system—Version 5.0. WIREs Comput. Mol. Sci. 2022, 12, e1606. [Google Scholar] [CrossRef]
- Lu, T.; Chen, F. Multiwfn: A multifunctional wavefunction analyzer. J. Comput. Chem. 2012, 33, 580–592. [Google Scholar] [CrossRef] [PubMed]
- Lu, T. Sobtop, Version [1.0(dev3)]. Available online: https://sobereva.com/soft/Sobtop (accessed on 26 March 2022).
- Liang, L.; Duan, W.; Zhang, J.; Huang, Y.; Zhang, Y.; Sun, B. Characterization and molecular docking study of taste peptides from chicken soup by sensory analysis combined with nano-LC-Q-TOF-MS/MS. Food Chem. 2022, 383, 132455. [Google Scholar] [CrossRef] [PubMed]
- Valdés-Tresanco, M.S.; Valiente, P.A.; Moreno, E. gmx_MMPBSA: A New Tool to Perform End-State Free Energy Calculations with GROMACS. J. Chem. Theory Comput. 2021, 17, 6281–6291. [Google Scholar] [CrossRef] [PubMed]
- Yoon, S.; Kim, D.; Roh, K.M.; Ahn, D.; Kang, H.J.; Chung, S.J. Identification of Vaccinia -H1 Related Phosphatase as an Anticancer Target for 1,2,3,4,6- O -Pentagalloylglucose. Chem. Biodivers. 2019, 17, e1900414. [Google Scholar] [CrossRef]
- Liu, T.; Cao, L.; Zhang, T.; Fu, H. Molecular docking studies, anti-Alzheimer’s disease, antidiabetic, and anti-acute myeloid leukemia potentials of narcissoside. Arch. Physiol. Biochem. 2020, 19, 1–11. [Google Scholar] [CrossRef]
- Deng, R.; Xu, Y.; Feng, F.; Liu, W. Identification of poliumoside metabolites in rat feces by high performance liquid chromatography coupled with quadrupole time-of-flight tandem mass spectrometry. J. Chromatogr. B 2014, 969, 285–296. [Google Scholar] [CrossRef]
- Yu, S.Y.; Lee, I.-S.; Jung, S.-H.; Lee, Y.M.; Lee, Y.-R.; Kim, J.-H.; Sun, H.; Kim, J.S. Caffeoylated Phenylpropanoid Glycosides from Brandisia hancei Inhibit Advanced Glycation End Product Formation and Aldose Reductase in Vitro and Vessel Dilation in Larval Zebrafish in Vivo. Planta Medica 2013, 79, 1705–1709. [Google Scholar] [CrossRef]
- He, Z.-D.; Lau, K.-M.; Xu, H.-X.; Li, P.-C.; But, P.P.-H. Antioxidant activity of phenylethanoid glycosides from Brandisia hancei. J. Ethnopharmacol. 2000, 71, 483–486. [Google Scholar] [CrossRef]
- Wu, A.; Yang, Z.; Huang, Y.; Yuan, H.; Lin, C.; Wang, T.; Zhao, Z.; Zhou, Y.; Zhu, C. Natural phenylethanoid glycosides isolated from Callicarpa kwangtungensis suppressed lipopolysaccharide-mediated inflammatory response via activating Keap1/Nrf2/HO-1 pathway in RAW 264.7 macrophages cell. J. Ethnopharmacol. 2020, 258, 112857. [Google Scholar] [CrossRef] [PubMed]
- Li, P.; Zhang, F.; Li, Y.; Zhang, C.; Yang, Z.; Zhang, Y.; Song, C. Isoginkgetin treatment attenuated lipopolysaccharide-induced monoamine neurotransmitter deficiency and depression-like behaviors through downregulating p38/NF-κB signaling pathway and suppressing microglia-induced apoptosis. J. Psychopharmacol. 2021, 35, 1285–1299. [Google Scholar] [CrossRef] [PubMed]
- Vo, V.A.; Lee, J.-W.; Chang, J.-E.; Kim, J.-Y.; Kim, N.-H.; Lee, H.J.; Kim, S.-S.; Chun, W.; Kwon, Y.-S. Avicularin Inhibits Lipopolysaccharide-Induced Inflammatory Response by Suppressing ERK Phosphorylation in RAW 264.7 Macrophages. Biomol. Ther. 2012, 20, 532–537. [Google Scholar] [CrossRef] [PubMed]
- Amadi, J.A.; Amadi, P.; Njoku, U.C.; Onitchi, C.L. Lettuce-avicularin treatment reverses insulin resistance through stimulation of glycolytic kinases and insulin signaling molecules. Iran. J. Basic Med. Sci. 2021, 24, 232–239. [Google Scholar] [CrossRef] [PubMed]
- Liang, L.; Zhou, C.; Zhang, J.; Huang, Y.; Zhao, J.; Sun, B.; Zhang, Y. Characteristics of umami peptides identified from por-cine bone soup and molecular docking to the taste receptor T1R1/T1R3. Food. Chem. 2022, 387, 132870. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compounds | CAS | Molecular Weight (Da) | Docking Energy (kcal/mol) | Ranking |
|---|---|---|---|---|
| Pentagalloylglucose | 14937-32-7 | 940.7 | −9.91 | 1 |
| Narcissoside | 604-80-8 | 624.5 | −8.222 | 2 |
| Poliumoside | 94079-81-9 | 770.7 | −8.183 | 3 |
| Isoginkgetin | 548-19-6 | 566.5 | −7.984 | 4 |
| Avicularin | 572-30-5 | 434.3 | −7.984 | 5 |
| Isoliensinine | 6817-41-0 | 610.7 | −7.938 | 6 |
| Isoquercitrin | 482-35-9 | 464.4 | −7.913 | 7 |
| Thonningianin A | 271579-11-4 | 874.7 | −7.823 | 8 |
| 2′-O-galloylhyperin | 53209-27-1 | 616.5 | −7.749 | 9 |
| Forsythoside B | 81525-13-5 | 756.7 | −7.743 | 10 |
| Crotonoside | 1818-71-9 | 283.24 | −7.727 | 11 |
| Gnetol | 86361-55-9 | 244.24 | −7.7 | 12 |
| Oxyresveratrol | 29700-22-9 | 244.24 | −7.682 | 13 |
| (-)-swainsonine | 136997-64-3 | 712.7 | −7.639 | 14 |
| Amentoflavone | 1617-53-4 | 538.5 | −7.584 | 15 |
| Brazilin | 474-07-7 | 286.28 | −7.538 | 16 |
| Liquiritin Apioside | 74639-14-8 | 550.5 | −7.491 | 17 |
| Rhodiosin | 86831-54-1 | 610.5 | −7.489 | 18 |
| Chelidonine | 476-32-4 | 353.4 | −7.452 | 19 |
| Rhapontigenin | 500-65-2 | 258.269 | −7.442 | 20 |
| Compounds | Ksv (×106 L·mol−1) | Ka (×106 L·mol−1) | n |
|---|---|---|---|
| Pentagalloylglucose | 0.047 ± 0.001 a | 0.049 ± 0.010 a | 1.00 ± 0.04 b |
| Narcissoside | 0.020 ± 0.001 c | 0.0013 ± 0.0004 b | 1.71 ± 0.16 a |
| Poliumoside | 0.0094 ± 0.0010 d | 0.0012 ± 0.0006 b | 1.52 ± 0.18 a |
| Isoginkgetin | 0.042 ± 0.002 b | 0.060 ± 0.020 a | 0.98 ± 0.12 b |
| Avicularin | 0.040 ± 0.001 b | 0.058 ± 0.010 a | 0.90 ± 0.05 b |
| Title 1 | Title 2 |
|---|---|
| Van der Waals | −64.15 |
| Electrostatic | −8.13 |
| Polar solvation | 42.46 |
| Nonpolar solvation | −4.82 |
| Ggas | −72.28 |
| Gsolv | 37.64 |
| Total | −34.64 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, P.; Wu, A.; Song, Y.; Zhao, J. Virtual Screening of Soybean Protein Isolate-Binding Phytochemicals and Interaction Characterization. Foods 2023, 12, 272. https://doi.org/10.3390/foods12020272
Liu P, Wu A, Song Y, Zhao J. Virtual Screening of Soybean Protein Isolate-Binding Phytochemicals and Interaction Characterization. Foods. 2023; 12(2):272. https://doi.org/10.3390/foods12020272
Chicago/Turabian StyleLiu, Panhang, Annan Wu, Yi Song, and Jing Zhao. 2023. "Virtual Screening of Soybean Protein Isolate-Binding Phytochemicals and Interaction Characterization" Foods 12, no. 2: 272. https://doi.org/10.3390/foods12020272
APA StyleLiu, P., Wu, A., Song, Y., & Zhao, J. (2023). Virtual Screening of Soybean Protein Isolate-Binding Phytochemicals and Interaction Characterization. Foods, 12(2), 272. https://doi.org/10.3390/foods12020272