
Integrative Analysis of Proteomics and Transcriptomics of Longissimus dorsi with Different Feeding Systems in Yaks
 , ,
, ,
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals, Feeding Regimes, and Weight Determination
2.2. Tissue Collection and Measurements
2.3. RNA-Seq Analysis
2.4. Protein Digestion and TMT Labeling
2.5. Multi-Omics Joint-Level Analysis (Transcriptome and Proteome)
2.6. Target Analysis Using Parallel Reaction Monitoring
2.7. Statistical Analyses
3. Results
3.1. Characterization of Growth Performance and Meat and Carcass Traits
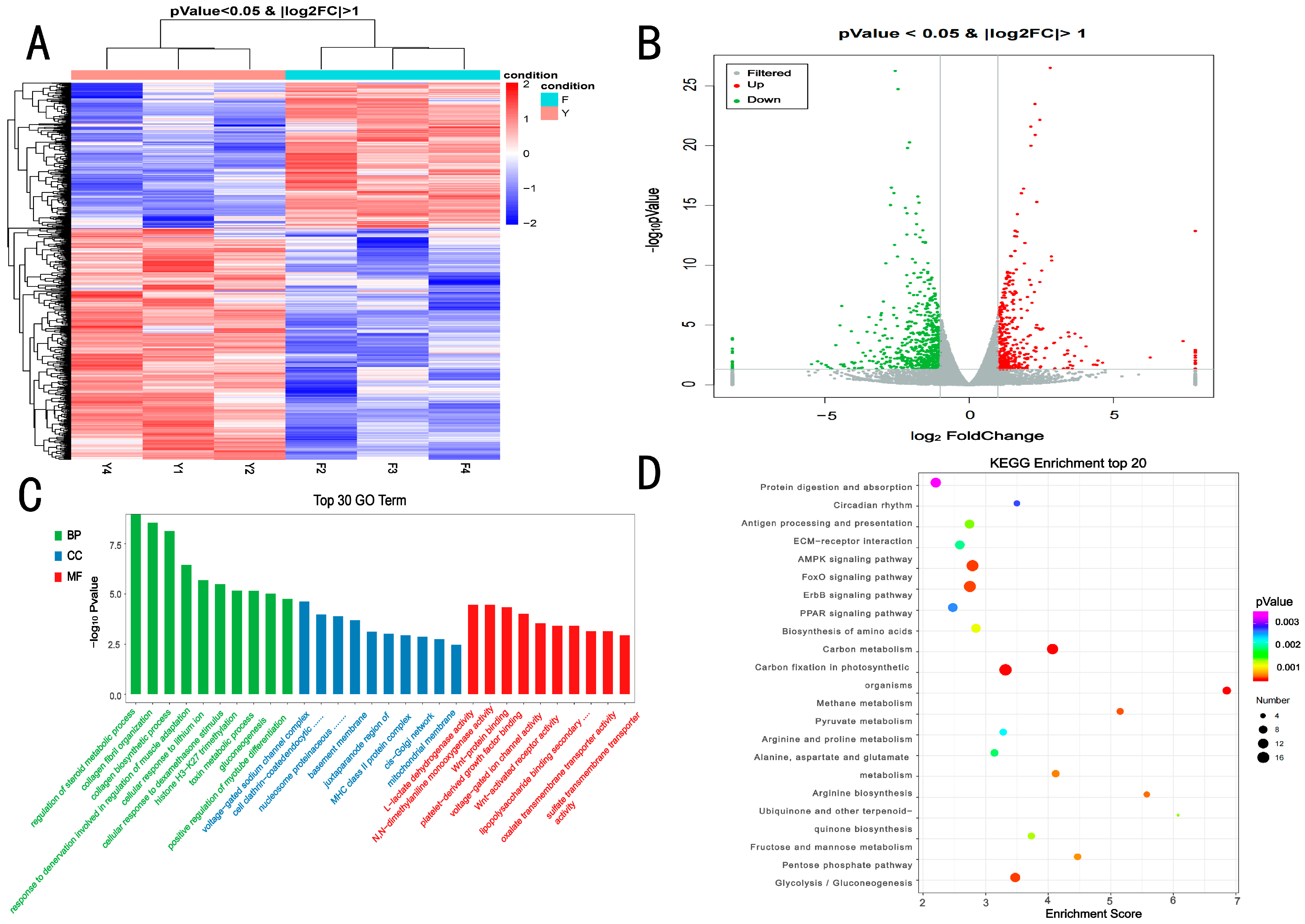
3.2. Overall Statistics for Transcriptomic Analysis
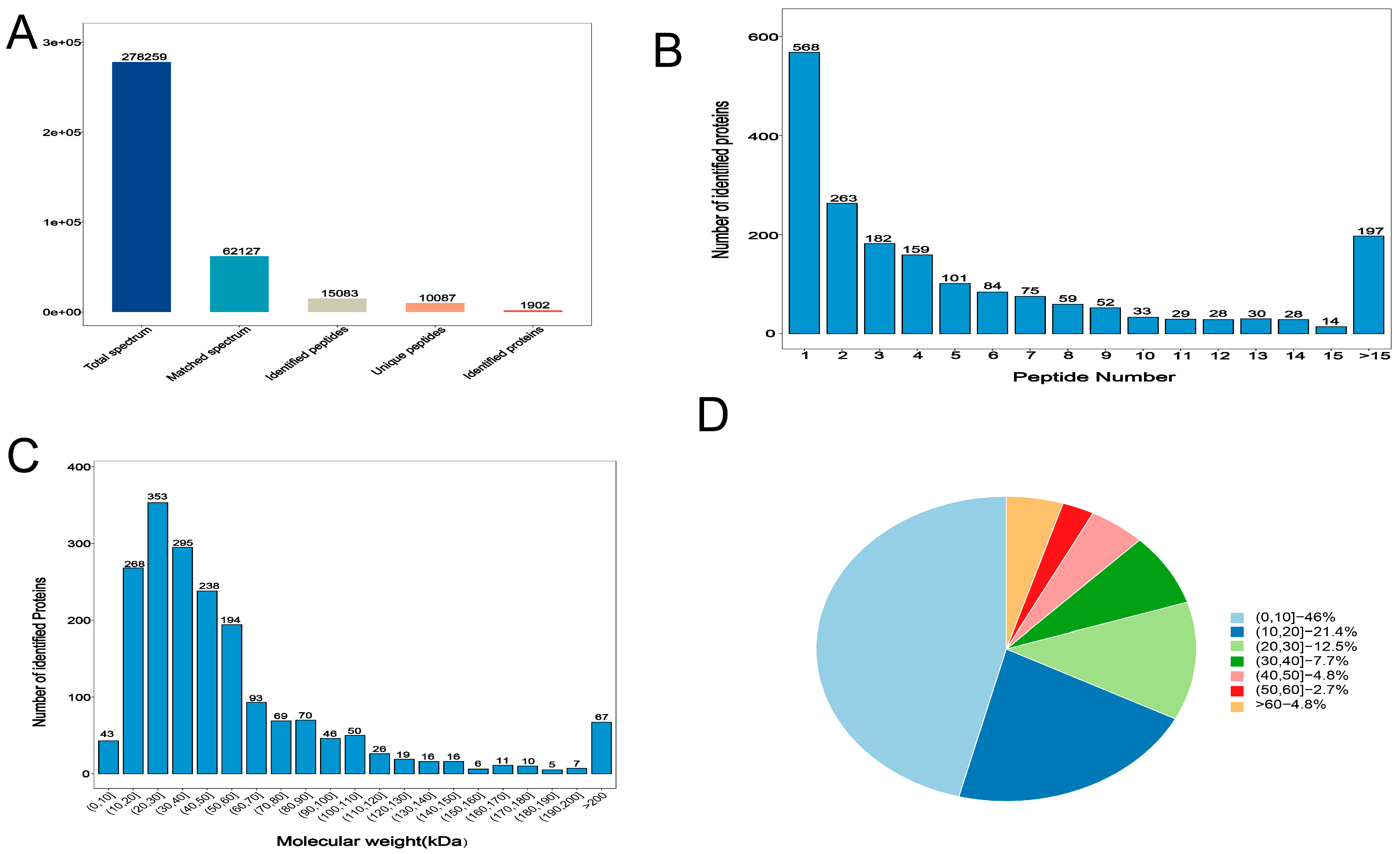
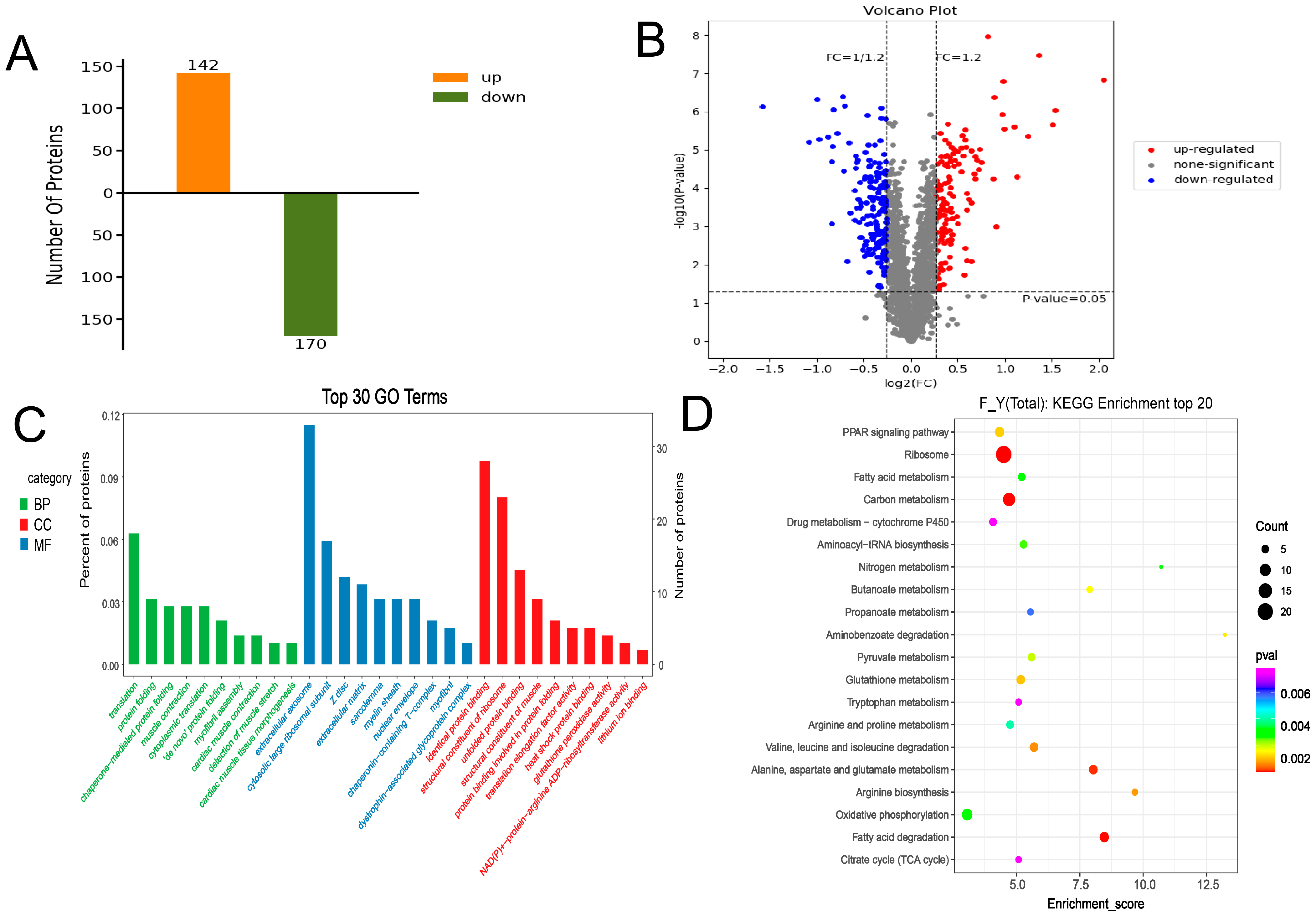
3.3. Overall Statistics for Proteomics Analysis
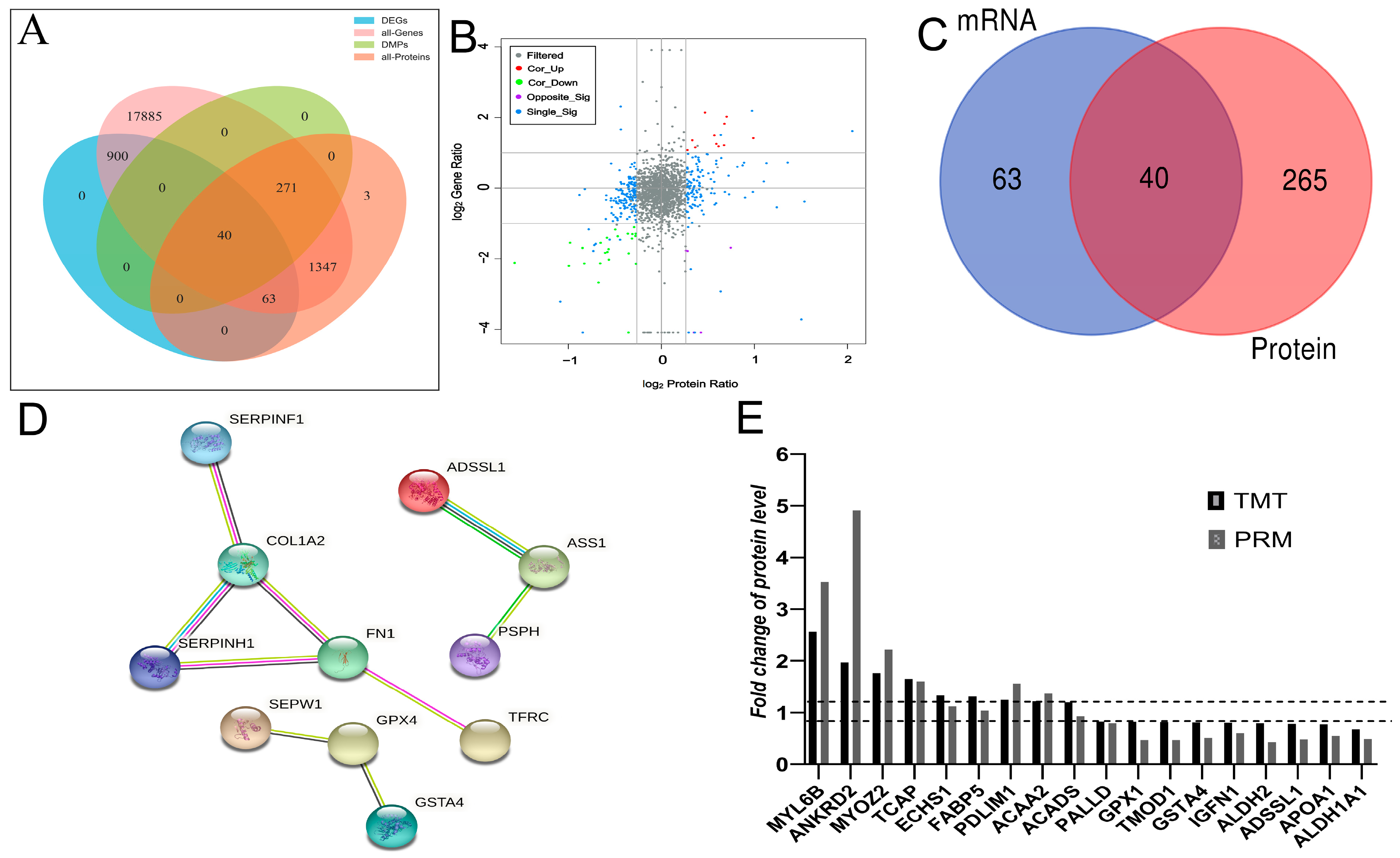
3.4. Correlation between Transcriptome and Proteome
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| TMT | Tandem Mass Tag |
| LD | Longissimus dorsi |
| DEGs | Differentially expressed genes |
| DAPs | Differentially abundant proteins |
| PRM | Parallel reaction monitoring |
| FA | Fatty acid |
| HPLC | High-performance liquid chromatography |
| FC | Fold change |
| KEGG | Kyoto Encyclopedia of Genes and Genomes |
| GO | Gene Ontology |
| PPI | Protein-protein interaction |
| LC-MS/MS | Liquid chromatography-tandem mass spectrometry |
| IMF | Intramuscular fat |
| FABP | FA-binding protein |
| MYH | Myosin heavy chain |
| PGAM2 | Phosphoglycerate mutase 2 |
| COL1A2 | Collagen type I alpha 2 chain |
| PPAR | Peroxisome proliferator-activated receptor |
| FoxO | Forkhead box, sub-group O |
| FDR | False discovery rate |
References
- Hu, C.; Ding, L.; Jiang, C.; Ma, C.; Liu, B.; Li, D.; Degen, A.A. Effects of Management, Dietary Intake, and Genotype on Rumen Morphology, Fermentation, and Microbiota, and on Meat Quality in Yaks and Cattle. Front. Nutr. 2021, 8, 755255. [Google Scholar] [CrossRef] [PubMed]
- Miao, F.; Guo, Z.; Xue, R.; Wang, X.; Shen, Y. Effects of grazing and precipitation on herbage biomass, herbage nutritive value, and yak performance in an alpine meadow on the Qinghai-Tibetan Plateau. PLoS ONE 2015, 10, e0127275. [Google Scholar] [CrossRef]
- Coles, C.A. Adipokines in Healthy Skeletal Muscle and Metabolic Disease. Adv. Exp. Med. Biol. 2016, 900, 133–160. [Google Scholar] [CrossRef]
- Liu, X.H.; Harlow, L.; Graham, Z.A.; Bauman, W.A.; Cardozo, C. Spinal Cord Injury Leads to Hyperoxidation and Nitrosylation of Skeletal Muscle Ryanodine Receptor-1 Associated with Upregulation of Nicotinamide Adenine Dinucleotide Phosphate Oxidase 4. J. Neurotrauma 2017, 34, 2069–2074. [Google Scholar] [CrossRef]
- Aldenderfer, M. Peopling the Tibetan plateau: Insights from archaeology. High Alt. Med. Biol. 2011, 12, 141–147. [Google Scholar] [CrossRef] [PubMed]
- Amitani, M.; Asakawa, A.; Amitani, H.; Inui, A. Control of food intake and muscle wasting in cachexia. Int. J. Biochem. Cell Biol. 2013, 45, 2179–2185. [Google Scholar] [CrossRef] [PubMed]
- Du, M.; Yang, C.; Liang, Z.; Zhang, J.; Yang, Y.; Ahmad, A.A.; Yan, P.; Ding, X. Dietary Energy Levels Affect Carbohydrate Metabolism-Related Bacteria and Improve Meat Quality in the Longissimus Thoracis Muscle of Yak (Bos grunniens). Front. Vet. Sci. 2021, 8, 718036. [Google Scholar] [CrossRef]
- Ye, C.; Zeng, X.; Zhu, J.; Liu, Y.; Ye, Q.; Qiao, S.; Zeng, X. Dietary N-Carbamylglutamate Supplementation in a Reduced Protein Diet Affects Carcass Traits and the Profile of Muscle Amino Acids and Fatty Acids in Finishing Pigs. J. Agric. Food Chem. 2017, 65, 5751–5758. [Google Scholar] [CrossRef]
- da Costa, M.J.; Huertas, S.M.; Gallo, C.; Dalla Costa, O.A. Strategies to promote farm animal welfare in Latin America and their effects on carcass and meat quality traits. Meat. Sci. 2012, 92, 221–226. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Wang, Y.; Hussain, T.; Dai, C.; Li, J.; Huang, P.; Li, Y.; Ding, X.; Huang, J.; Ji, F.; et al. Effects of dietary energy level on growth performance, blood parameters and meat quality in fattening male Hu lambs. J. Anim. Physiol. Anim. Nutr. 2020, 104, 418–430. [Google Scholar] [CrossRef]
- Cimmino, R.; Barone, C.M.A.; Claps, S.; Varricchio, E.; Rufrano, D.; Caroprese, M.; Albenzio, M.; De Palo, P.; Campanile, G.; Neglia, G. Effects of dietary supplementation with polyphenols on meat quality in Saanen goat kids. BMC Vet. Res. 2018, 14, 181. [Google Scholar] [CrossRef]
- Dieters, L.S.E.; Meale, S.J.; Quigley, S.P.; Hoffman, L.C. Meat quality characteristics of lot-fed Australian Rangeland goats are unaffected by live weight at slaughter. Meat. Sci. 2021, 175, 108437. [Google Scholar] [CrossRef]
- Liu, Y.-X.; Ma, X.-M.; Xiong, L.; Wu, X.-Y.; Liang, C.-N.; Bao, P.-J.; Yu, Q.-L.; Yan, P. Effects of Intensive Fattening with Total Mixed Rations on Carcass Characteristics, Meat Quality, and Meat Chemical Composition of Yak and Mechanism Based on Serum and Transcriptomic Profiles. Front. Vet. Sci. 2021, 7, 599418. [Google Scholar] [CrossRef]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A flexible trimmer for Illumina sequence data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef] [PubMed]
- Daehwan, K.; Ben, L.; Salzberg, S.L. HISAT: A fast spliced aligner with low memory requirements. Nat. Methods 2015, 12, 357–360. [Google Scholar] [CrossRef]
- Trapnell, C. Transcript assembly and quantification by RNA-Seq reveals unannotated transcripts and isoform switching during cell differentiation. Nat Biotechnol. Nat. Biotechnol. 2010, 28, 511–515. [Google Scholar] [CrossRef]
- Anders, S. Analysing RNA-Seq data with the DESeq package. Mol. Biol. 2010, 43, 1–17. [Google Scholar]
- MacLean, B.; Tomazela, D.M.; Shulman, N.; Chambers, M.; Finney, G.L.; Frewen, B.; Kern, R.; Tabb, D.L.; Liebler, D.C.; MacCoss, M.J. Skyline: An open source document editor for creating and analyzing targeted proteomics experiments. Bioinformatics 2010, 26, 966–968. [Google Scholar] [CrossRef] [PubMed]
- Ding, B.A.; Ma, S.Q.; Li, Z.R.; Li, X.L.; Madigosky, S.R. Seasonal changes of rumen and intestine morphology of the Qinghai yak (Bos grunniens). Vet. World 2018, 11, 1135–1138. [Google Scholar] [CrossRef] [PubMed]
- Listrat, A.; Lebret, B.; Louveau, I.; Astruc, T.; Bonnet, M.; Lefaucheur, L.; Picard, B.; Bugeon, J. How Muscle Structure and Composition Influence Meat and Flesh Quality. Sci. World J. 2016, 2016, 3182746. [Google Scholar] [CrossRef]
- Geletu, U.S.; Usmael, M.A.; Mummed, Y.Y.; Ibrahim, A.M. Quality of Cattle Meat and Its Compositional Constituents. Vet. Med. Int. 2021, 2021, 7340495. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Wang, Z.; Hu, R.; Peng, Q.; Xue, B.; Wang, L. Comparison of carcass characteristics and meat quality between Simmental crossbred cattle, cattle-yaks and Xuanhan yellow cattle. J. Sci. Food Agric. 2021, 101, 3927–3932. [Google Scholar] [CrossRef] [PubMed]
- Abril, M.; Campo, M.M.; Onenç, A.; Sañudo, C.; Albertí, P.; Negueruela, A.I. Beef colour evolution as a function of ultimate pH. Meat. Sci. 2001, 58, 69–78. [Google Scholar] [CrossRef] [PubMed]
- Neath, K.E.; Del Barrio, A.N.; Lapitan, R.M.; Herrera, J.R.; Cruz, L.C.; Fujihara, T.; Muroya, S.; Chikuni, K.; Hirabayashi, M.; Kanai, Y. Difference in tenderness and pH decline between water buffalo meat and beef during postmortem aging. Meat. Sci. 2007, 75, 499–505. [Google Scholar] [CrossRef]
- La, Y.; Tang, J.; Guo, X.; Zhang, L.; Gan, S.; Zhang, X.; Zhang, J.; Hu, W. Proteomic analysis of sheep uterus reveals its role in prolificacy. J. Proteom. 2019, 210, 103526. [Google Scholar] [CrossRef]
- Tree, J.J.; Gerdes, K.; Tollervey, D. Transcriptome-Wide Analysis of Protein-RNA and RNA-RNA Interactions in Pathogenic Bacteria. Methods Enzym. 2018, 612, 467–488. [Google Scholar] [CrossRef]
- Murtaza, G.; Khan, A.K.; Rashid, R.; Muneer, S.; Hasan, S.M.F.; Chen, J. FOXO Transcriptional Factors and Long-Term Living. Oxid. Med. Cell Longev. 2017, 2017, 3494289. [Google Scholar] [CrossRef]
- Kamei, Y.; Miura, S.; Suzuki, M.; Kai, Y.; Mizukami, J.; Taniguchi, T.; Mochida, K.; Hata, T.; Matsuda, J.; Aburatani, H.; et al. Skeletal muscle FOXO1 (FKHR) transgenic mice have less skeletal muscle mass, down-regulated Type I (slow twitch/red muscle) fiber genes, and impaired glycemic control. J. Biol. Chem. 2004, 279, 41114–41123. [Google Scholar] [CrossRef]
- Wang, Y.Z.; Huang, Y.N.; Sun, K.Y.; Qi, J.H.; Xiang, L. Leptin gene transfer regulates fibromuscular development and lipid deposition in muscles via SIRT1, FOXO3a and PGC-1α in mice in vivo. Int. J. Mol. Med. 2011, 28, 617–623. [Google Scholar] [CrossRef]
- Kang, Y.K.; Choi, Y.M.; Lee, S.H.; Choe, J.H.; Hong, K.C.; Kim, B.C. Effects of myosin heavy chain isoforms on meat quality, fatty acid composition, and sensory evaluation in Berkshire pigs. Meat. Sci. 2011, 89, 384–389. [Google Scholar] [CrossRef]
- Ryu, Y.C.; Kim, B.C. The relationship between muscle fiber characteristics, postmortem metabolic rate, and meat quality of pig longissimus dorsi muscle. Meat. Sci. 2005, 71, 351–357. [Google Scholar] [CrossRef] [PubMed]
- Fothergill-Gilmore, L.A.; Watson, H.C. The phosphoglycerate mutases. Adv. Enzymol. Relat. Areas Mol. Biol. 1989, 62, 227–313. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Wei, Z.; Liu, L.; Cheng, Z.; Lin, Y.; Ji, F.; Gong, W. Crystal structure of human B-type phosphoglycerate mutase bound with citrate. Biochem. Biophys. Res. Commun. 2005, 331, 1207–1215. [Google Scholar] [CrossRef] [PubMed]
- Qiu, H.; Zhao, S.; Xu, X.; Yerle, M.; Liu, B. Assignment and expression patterns of porcine muscle-specific isoform of phosphoglycerate mutase gene. J. Genet Genom. 2008, 35, 257–260. [Google Scholar] [CrossRef]
- Fontanesi, L.; Davoli, R.; Nanni Costa, L.; Beretti, F.; Scotti, E.; Tazzoli, M.; Tassone, F.; Colombo, M.; Buttazzoni, L.; Russo, V. Investigation of candidate genes for glycolytic potential of porcine skeletal muscle: Association with meat quality and production traits in Italian Large White pigs. Meat. Sci. 2008, 80, 780–787. [Google Scholar] [CrossRef]
- He, K.; Wang, Q.; Wang, Z.; Pan, Y. Association study between gene polymorphisms in PPAR signaling pathway and porcine meat quality traits. Mamm. Genome 2013, 24, 322–331. [Google Scholar] [CrossRef]
- Yu, X.H.; Zheng, X.L.; Tang, C.K. Peroxisome Proliferator-Activated Receptor α in Lipid Metabolism and Atherosclerosis. Adv. Clin. Chem. 2015, 71, 171–203. [Google Scholar] [CrossRef]
- Green, S.; Wahli, W. Peroxisome proliferator-activated receptors: Finding the orphan a home. Mol. Cell Endocrinol. 1994, 100, 149–153. [Google Scholar] [CrossRef]
- Li, J.; Henry, E.; Wang, L.; Delelis, O.; Wang, H.; Simon, F.; Tauc, P.; Brochon, J.C.; Zhao, Y.; Deprez, E. Comparative study of the fatty acid binding process of a new FABP from Cherax quadricarinatus by fluorescence intensity, lifetime and anisotropy. PLoS ONE 2012, 7, e51079. [Google Scholar] [CrossRef]
- Huang, X.; Zhou, Y.; Sun, Y.; Wang, Q. Intestinal fatty acid binding protein: A rising therapeutic target in lipid metabolism. Prog. Lipid Res. 2022, 87, 101178. [Google Scholar] [CrossRef]
- Ishizawa, R.; Masuda, K.; Sakata, S.; Nakatani, A. Effects of different fatty acid chain lengths on fatty acid oxidation-related protein expression levels in rat skeletal muscles. J. Oleo Sci. 2015, 64, 415–421. [Google Scholar] [CrossRef] [PubMed]
- Lim, D.; Chai, H.H.; Lee, S.H.; Cho, Y.M.; Choi, J.W.; Kim, N.K. Gene Expression Patterns Associated with Peroxisome Proliferator-activated Receptor (PPAR) Signaling in the Longissimus dorsi of Hanwoo (Korean Cattle). Asian-Australas J. Anim. Sci. 2015, 28, 1075–1083. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Wang, J.; Yang, D.D.; Liu, Z.L.; Zeng, Y.Q.; Chen, W. Expression of lipid metabolism genes provides new insights into intramuscular fat deposition in Laiwu pigs. Asian-Australas J. Anim. Sci. 2020, 33, 390–397. [Google Scholar] [CrossRef]
- Wang, Y.; Sakaguchi, M.; Sabit, H.; Tamai, S.; Ichinose, T.; Tanaka, S.; Kinoshita, M.; Uchida, Y.; Ohtsuki, S.; Nakada, M. COL1A2 inhibition suppresses glioblastoma cell proliferation and invasion. J. Neurosurg. 2022, 1, 1–10. [Google Scholar] [CrossRef]
- Gelse, K.; Pöschl, E.; Aigner, T. Collagens—Structure, function, and biosynthesis. Adv. Drug Deliv. Rev. 2003, 55, 1531–1546. [Google Scholar] [CrossRef] [PubMed]
- Tang, X.; Xu, G.; Dai, H.; Xu, P.; Zhang, C.; Gu, R. Differences in muscle cellularity and flesh quality between wild and farmed Coilia nasus (Engraulidae). J. Sci. Food Agric. 2012, 92, 1504–1510. [Google Scholar] [CrossRef]
- Sato, K.; Yoshinaka, R.; Itoh, Y.; Sato, M. Molecular species of collagen in the intramuscular connective tissue of fish. Comp. Biochem. Physiol. Part B Comp. Biochem. 1989, 92, 87–91. [Google Scholar] [CrossRef]
- Daiwen, C.; Keying, Z.; Zuyu, H. The mechanism for developing pork quality traits. Sichuan Nongye Daxue Xuebao 2002, 20, 60–66. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Item | Content (%) |
|---|---|
| Corn | 48.53 |
| Wheat Bran | 6.22 |
| Rapeseed Cake | 7.25 |
| Baking Soda | 0.25 |
| Dairy Salt | 0.25 |
| 4% Premix Compound | 1.5 |
| Oat Hay | 22 |
| Alfalfa Hays | 14 |
| Total | 100 |
| Carcass Characteristics | Group G | Group HF | p-Value |
|---|---|---|---|
| Initial body weight/kg | 208.33 ± 12.53 | 215.67 ± 21.83 | 0.6549 |
| Final body weight/kg | 306.83 ± 24.34 B | 377.17 ± 15.77 A | 0.0010 |
| Average daily gain/kg | 0.52 ± 0.12 B | 0.86 ± 0.15 A | 0.0017 |
| Carcass weight/kg | 124.28 ± 8.77 B | 208.04 ± 8.75 A | <0.001 |
| Net meat weight/kg | 94.73 ± 4.55 B | 176.23 ± 10.61 A | <0.001 |
| Bone weight/kg | 31.08 ± 1.96 b | 27.17 ± 2.46 a | 0.012 |
| Slaughter rate/% | 42.49 ± 10.24 B | 55.26 ± 14.25 A | <0.001 |
| Meat-bone ratio | 3.46 ± 0.09 B | 5.68 ± 0.29 A | <0.001 |
| Eye muscle area/cm2 | 57.89 ± 8.83 A | 76.64 ± 10.45 B | <0.001 |
| Meat Quality | Group G | Group HF | p-Value |
|---|---|---|---|
| L* 45 min | 6.32 ± 0.80 | 6.98 ± 1.27 | 0.305 |
| a* 45 min | 28.12 ± 2.32 | 29.29 ± 2.49 | 0.418 |
| b* 45 min | 5.67 ± 1.04 | 6.42 ± 1.38 | 0.311 |
| L * 24 h | 7.61 ± 0.99 | 8.86 ± 1.20 | 0.054 |
| a* 24h | 32.68 ± 0.91 | 33.23 ± 1.19 | 0.385 |
| b* 24 h | 9.94 ± 1.18 | 8.62 ± 0.81 | 0.047 |
| pH 45 min | 6.89 ± 0.32 A | 6.34 ± 0.24 B | 0.007 |
| pH 24 h | 5.49 ± 0.25 B | 6.03 ± 0.28 A | 0.005 |
| Cooking loss (%) | 17.82 ± 3.84 B | 27.21 ± 1.70 A | <0.001 |
| Drip loss (%) | 15.06 ± 2.30 | 17.27 ± 3.12 | 0.192 |
| Shear force (N) | 17.70 ± 1.51 A | 14.69 ± 2.35 B | 0.0025 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ma, X.; Guo, X.; La, Y.; Wu, X.; Chu, M.; Bao, P.; Yan, P.; Liang, C. Integrative Analysis of Proteomics and Transcriptomics of Longissimus dorsi with Different Feeding Systems in Yaks. Foods 2023, 12, 257. https://doi.org/10.3390/foods12020257
Ma X, Guo X, La Y, Wu X, Chu M, Bao P, Yan P, Liang C. Integrative Analysis of Proteomics and Transcriptomics of Longissimus dorsi with Different Feeding Systems in Yaks. Foods. 2023; 12(2):257. https://doi.org/10.3390/foods12020257
Chicago/Turabian StyleMa, Xiaoming, Xian Guo, Yongfu La, Xiaoyun Wu, Min Chu, Pengjia Bao, Ping Yan, and Chunnian Liang. 2023. "Integrative Analysis of Proteomics and Transcriptomics of Longissimus dorsi with Different Feeding Systems in Yaks" Foods 12, no. 2: 257. https://doi.org/10.3390/foods12020257
APA StyleMa, X., Guo, X., La, Y., Wu, X., Chu, M., Bao, P., Yan, P., & Liang, C. (2023). Integrative Analysis of Proteomics and Transcriptomics of Longissimus dorsi with Different Feeding Systems in Yaks. Foods, 12(2), 257. https://doi.org/10.3390/foods12020257