
Multi-Faceted Bioactivity Assessment of an Exopolysaccharide from Limosilactobacillus fermentum NCDC400: Antioxidant, Antibacterial, and Immunomodulatory Proficiencies
 , and
, and
Abstract
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Material and Methods
2.1. Production and Extraction of Exopolysaccharide
2.2. Determination of the Antioxidative Activity of EPS400
2.2.1. Preparation of EPS400 Solution
2.2.2. Determination of DPPH Scavenging Activity
2.2.3. Determination of ABTS Scavenging Activity
2.2.4. Determination of Hydroxyl Radical Scavenging Activity
2.2.5. Determination of Superoxide Anion Radical Scavenging Activity
2.2.6. Determination of Ferrous Ion Chelating Ability
2.3. Determination of Antimicrobial Activities of EPS400
2.4. Determination of Anti-Inflammatory Effects of EPS400 in Murine Peritoneal Macrophages
2.4.1. Isolation and Preparation of Murine Peritoneal Macrophage
2.4.2. Measurement of Cell Viability by MTT Assay
2.4.3. Measurement of Cytokine
2.4.4. Measurement of Nitrite Oxide
2.4.5. Phagocytosis Activity
2.5. Statistical Analysis
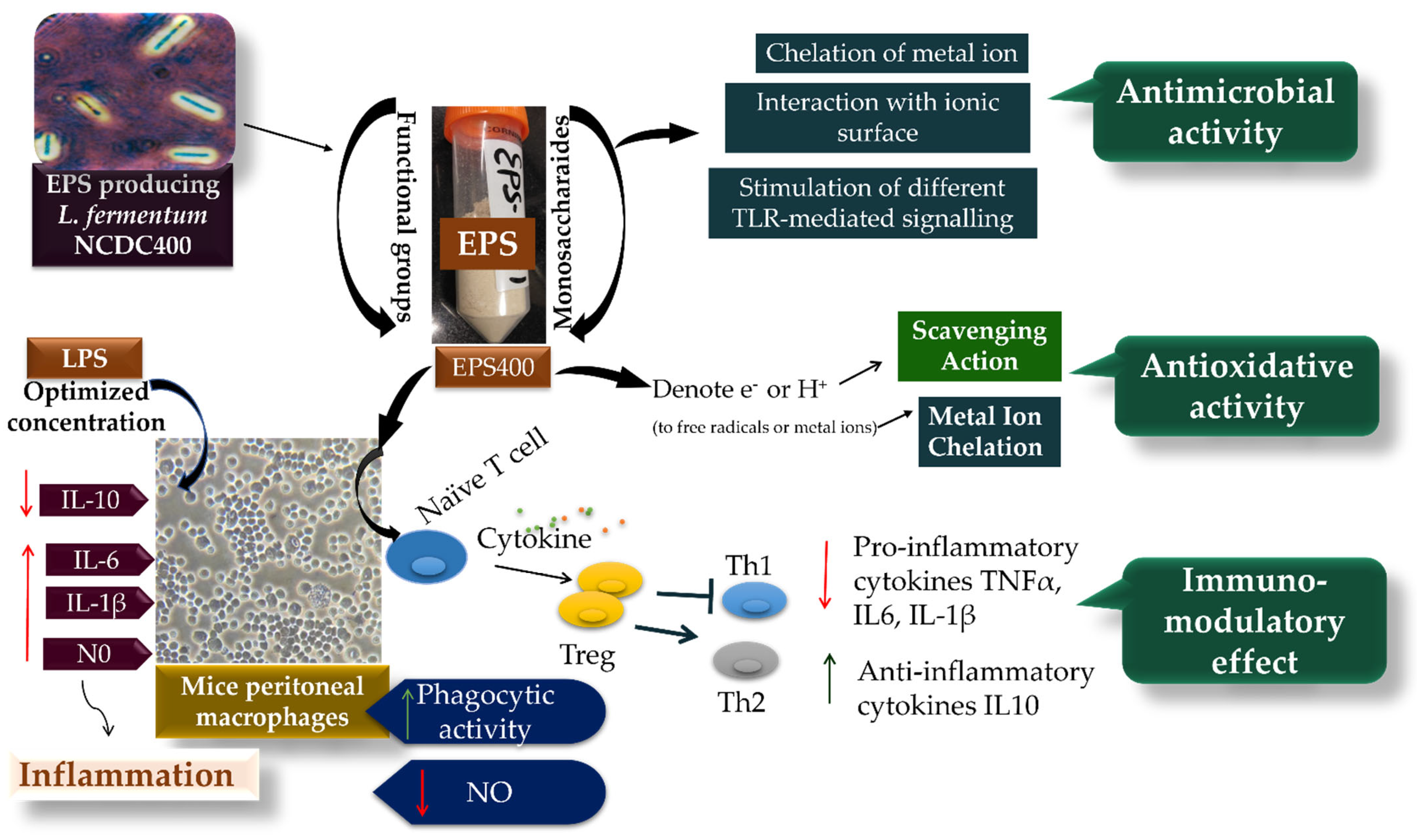
3. Results and Discussion
3.1. Yield of EPS400
3.2. Antioxidative Activities of EPS400
3.3. Antimicrobial Activity of EPS400
3.4. Immunomodulatory Potential of EPS400 in Murine Peritoneal Macrophages
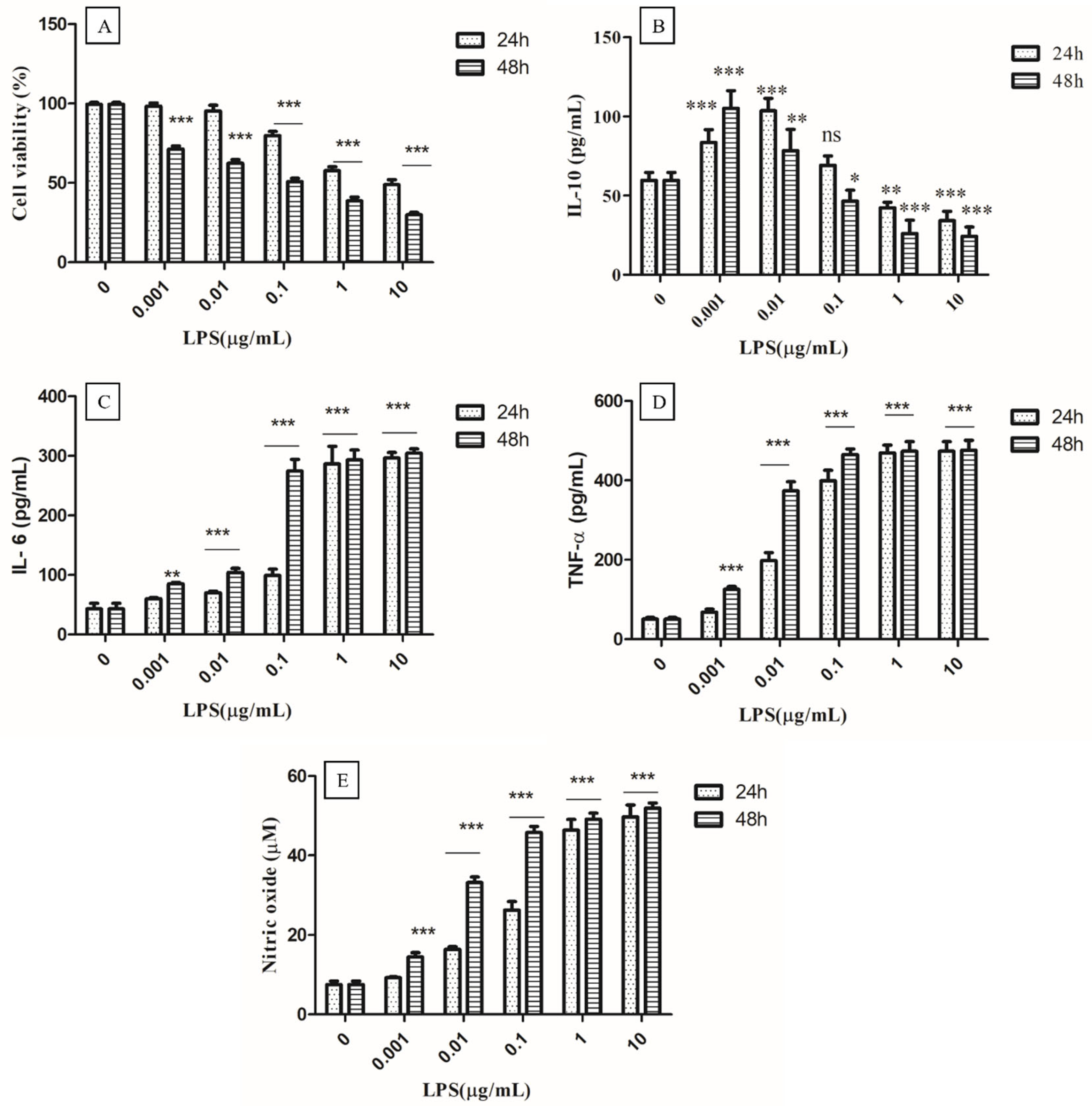
3.4.1. Optimization of Concentration of LPS
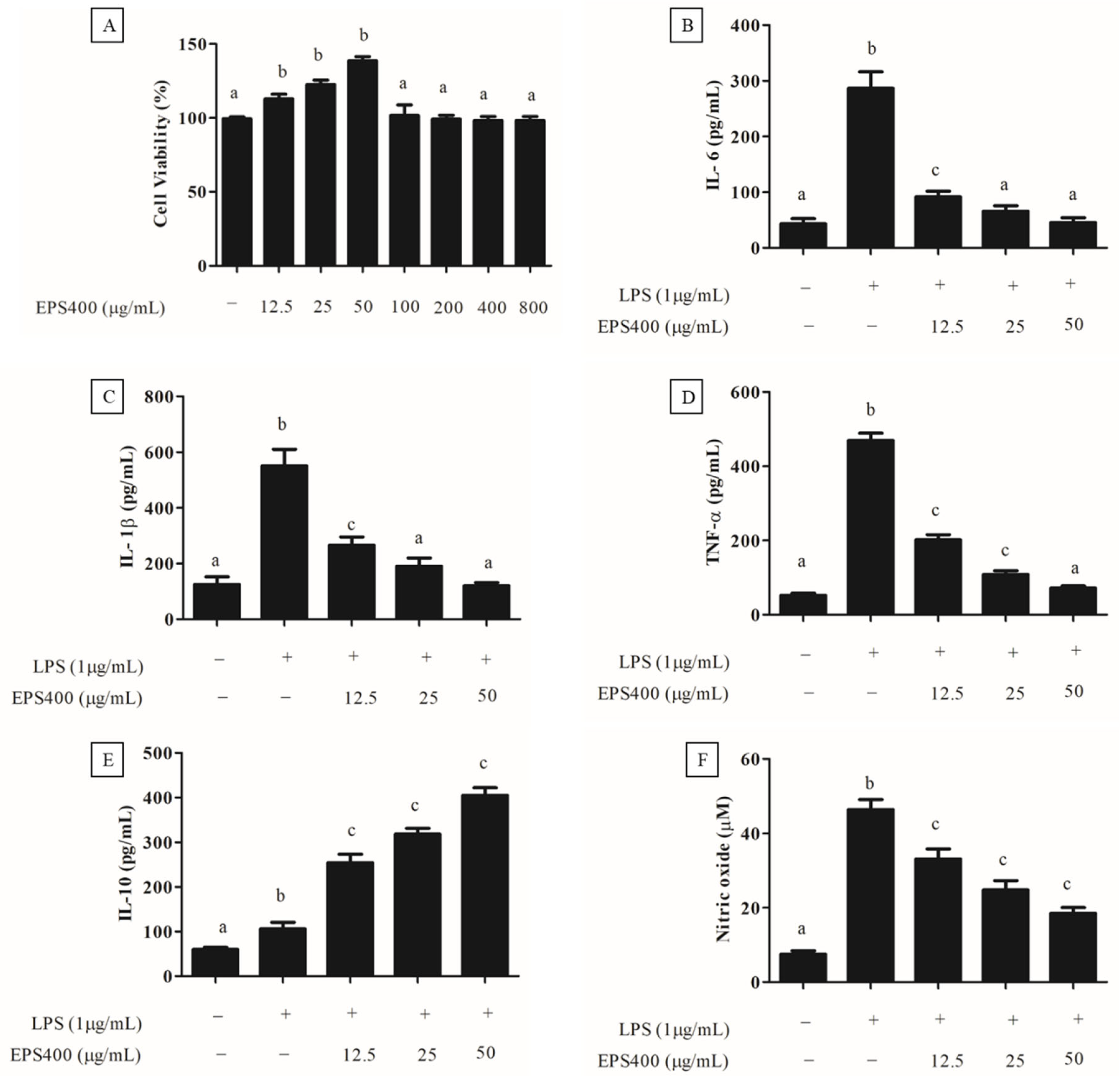
3.4.2. Effect of EPS400 on Cell Viability
3.4.3. Anti-Inflammatory Effect of EPS400 in Murine Peritoneal Macrophages
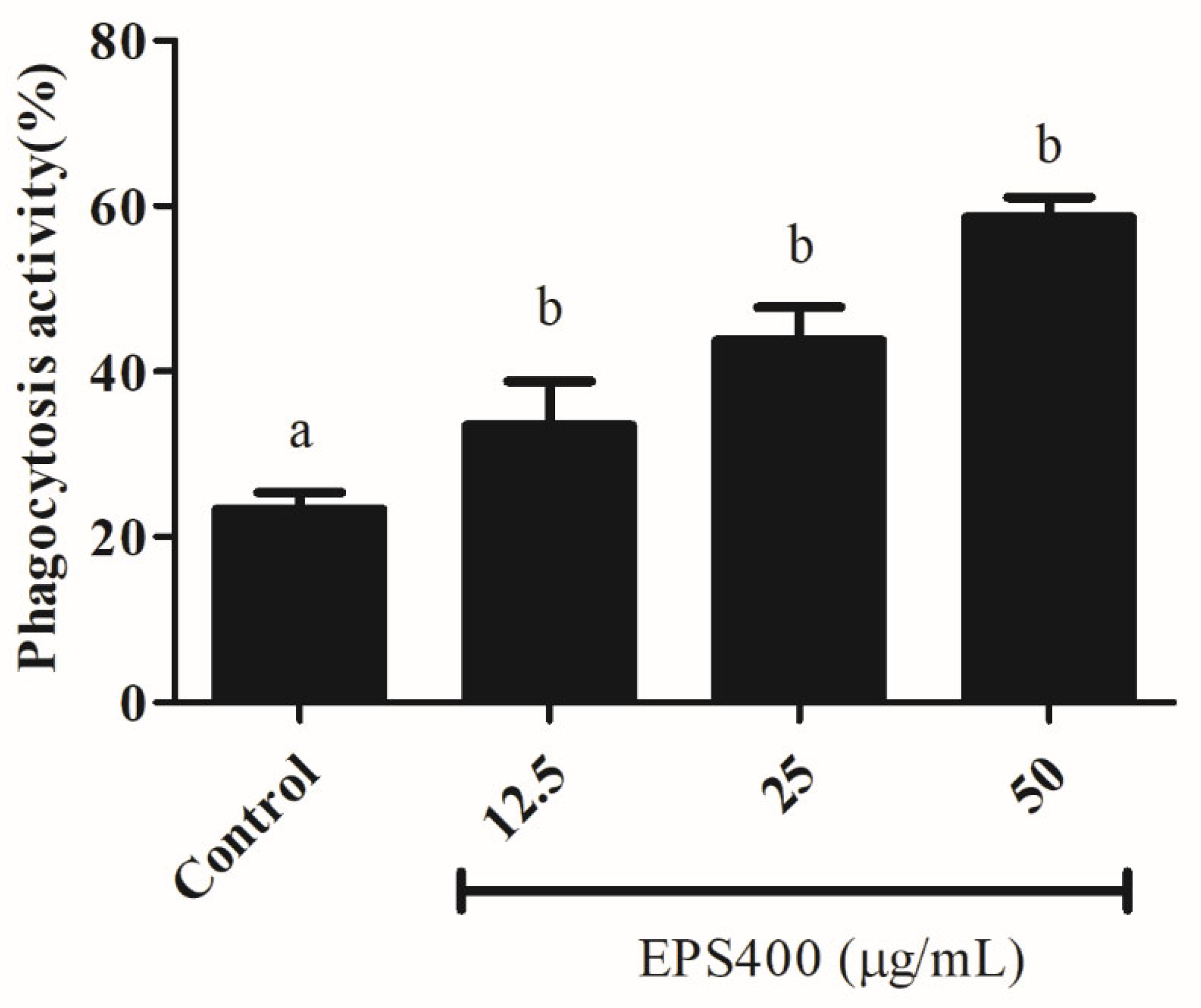
3.4.4. Effect of EPS400 on Phagocytosis Activity in Murine Peritoneal Macrophages
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Aslani, B.A.; Ghobadi, S. Studies on oxidants and antioxidants with a brief glance at their relevance to the immune system. Life Sci. 2016, 146, 163–173. [Google Scholar] [CrossRef] [PubMed]
- Singh, P.; Ali, S.A. Multifunctional role of S100 protein family in the immune system: An update. Cells 2022, 11, 2274. [Google Scholar] [CrossRef]
- Nani, A.; Murtaza, B.; Sayed Khan, A.; Khan, N.A.; Hichami, A. Antioxidant and anti-inflammatory potential of polyphenols contained in mediterranean diet in obesity: Molecular Mechanisms. Molecules 2021, 26, 985. [Google Scholar] [CrossRef]
- Ayyash, M.; Abu-Jdayil, B.; Itsaranuwat, P.; Galiwango, E.; Tamiello-Rosa, C.; Abdullah, H.; Hamed, F. Characterization, bioactivities, and rheological properties of exopolysaccharide produced by novel probiotic Lactobacillus plantarum C70 isolated from camel milk. Int. J. Biol. Macromol. 2020, 144, 938–946. [Google Scholar] [CrossRef] [PubMed]
- Gawande, K.; Kolhekar, M.; Kumari, M.; Kapila, S.; Sharma, P.; Ali, S.A.; Behare, P.V. Lactic acid bacteria based purified exopolysaccharide showed viscofying and hypercholesterolemic capabilites. Food Hydrocoll. Health 2021, 1, 100042. [Google Scholar] [CrossRef]
- Kumari, M.; Dasriya, V.L.; Nataraj, B.H.; Nagpal, R.; Behare, P.V. Lacticaseibacillus rhamnosus-Derived Exopolysaccharide Attenuates D-Galactose-Induced Oxidative Stress and Inflammatory Brain Injury and Modulates Gut Microbiota in a Mouse Model. Microorganisms 2022, 10, 2046. [Google Scholar] [CrossRef]
- Behare, P.V.; Singh, R.; Nagpal, R.; Rao, K.H. Exopolysaccharides producing Lactobacillus fermentum strain for enhancing rheological and sensory attributes of low-fat dahi. J. Food Sci. Technol. 2013, 50, 1228–1232. [Google Scholar] [CrossRef]
- Korcz, E.; Varga, L. Exopolysaccharides from lactic acid bacteria: Techno-functional application in the food industry. Trends Food Sci. 2021, 110, 375–384. [Google Scholar] [CrossRef]
- Oerlemans, M.M.; Akkerman, R.; Ferrari, M.; Walvoort, M.T.; de Vos, P. Benefits of bacteria-derived exopolysaccharides on gastrointestinal microbiota, immunity and health. J. Funct. Foods 2021, 76, 104289. [Google Scholar] [CrossRef]
- Wang, J.; Zhang, J.; Guo, H.; Cheng, Q.; Abbas, Z.; Tong, Y.; Zhang, R. Optimization of Exopolysaccharide Produced by Lactobacillus plantarum R301 and Its Antioxidant and Anti-Inflammatory Activities. Foods 2023, 12, 2481. [Google Scholar] [CrossRef]
- Azmal Ali, S.; Kumar, S.; Mohanty, A.K.; Behare, P. Draft genome sequence of Lactobacillus fermentum NCDC 400, isolated from a traditional Indian dairy product. Genome Announc. 2018, 6, e01492-17. [Google Scholar] [CrossRef] [PubMed]
- Nataraj, B.H.; Kumari, M.; Nagpal, R.; Ali, S.A.; Behare, P.V. Safety evaluation of indigenous probiotic Limosilactobacillus fermentum NCDC 400 using whole genome sequences and in vitro approaches. Food Biosci. 2023, 56, 103101. [Google Scholar] [CrossRef]
- Akram, M.; Ali, S.A.; Behare, P.; Kaul, G. Dietary intake of probiotic fermented milk benefits the gut and reproductive health in mice fed with an obesogenic diet. Food Funct. 2022, 13, 737–752. [Google Scholar] [CrossRef] [PubMed]
- Rani, K.; Ali, S.A.; Kaul, G.; Behare, P.V. Protective effect of probiotic and prebiotic fermented milk containing Lactobacillus fermentum against obesity-induced hepatic steatosis and inflammation. J. Food Biochem. 2022, 46, e14509. [Google Scholar] [CrossRef] [PubMed]
- Paulino do Nascimento, L.C.; Lacerda, D.C.; Ferreira, D.J.S.; de Souza, E.L.; de Brito Alves, J.L. Limosilactobacillus fermentum, current evidence on the antioxidant properties and opportunities to be exploited as a probiotic microorganism. Probiotics Antimicrob. 2022, 14, 960–979. [Google Scholar] [CrossRef]
- de Luna Freire, M.O.; do Nascimento LC, P.; de Oliveira, K.Á.R.; de Oliveira, A.M.; Napoleão, T.H.; Lima MD, S.; de Brito Alves, J.L. Effects of a mixed Limosilactobacillus fermentum formulation with claimed probiotic properties on cardiometabolic variables, biomarkers of inflammation and oxidative stress in male rats fed a high-fat diet. Foods 2021, 10, 2202. [Google Scholar] [CrossRef]
- Li, K.; Zhang, L.; Xue, J.; Yang, X.; Dong, X.; Sha, L.; He, L. Dietary Inulin Alleviates Diverse Stages of Type 2 Diabetes Mellitus via Anti-inflammation and Modulating Gut Microbiota in db/db mice. Food Funct. 2019, 10, 1915–1927. [Google Scholar] [CrossRef]
- El-Newary, S.A.; Ibrahim, A.Y.; Asker, M.S.; Mahmoud, M.G.; El Awady, M.E. Production, characterization and biological activities of acidic exopolysaccharide from marine Bacillus amyloliquefaciens 3MS 2017. Asian Pac. J. Trop. Med. 2017, 10, 652–662. [Google Scholar] [CrossRef]
- Kumari, M.; Patel, H.K.; Kokkiligadda, A.; Bhushan, B.; Tomar, S.K. Characterization of probiotic lactobacilli and development of fermented soymilk with improved technological properties. LWT 2022, 154, 112827. [Google Scholar] [CrossRef]
- Kapila, R.; Kapila, S.; Kapasiya, M.; Pandey, D.; Dang, A.; Saliganti, V. Comparative Evaluation of Oral Administration of Probiotic Lactobacilli-fermented Milks on Macrophage Function. Probiotics Antimicrob. Proteins 2012, 4, 173–179. [Google Scholar] [CrossRef]
- Strober, W. Trypan Blue Exclusion Test of Cell Viability. Curr. Protoc. Immunol. 1997, 21, A-3B. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Fang, X.; Wu, T.; Fang, L.; Liu, C.; Min, W. In vitro Immunomodulatory Effects of Acidic Exopolysaccharide Produced by Lactobacillus planetarium JLAU103 on RAW264. 7 Macrophages. Int. J. Biol. Macromol. 2020, 156, 1308–1315. [Google Scholar] [CrossRef] [PubMed]
- Srivastava, U.; Nataraj, B.H.; Kumari, M.; Kadyan, S.; Puniya, A.K.; Behare, P.V.; Nagpal, R. Antioxidant and immunomodulatory potency of Lacticaseibacillus rhamnosus NCDC24 fermented milk-derived peptides: A computationally guided in-vitro and ex-vivo investigation. Peptides 2022, 155, 170843. [Google Scholar] [CrossRef]
- Bhat, B.; Vaid, S.; Habib, B.; Bajaj, B.K. Design of experiments for enhanced production of bioactive exopolysaccharides from indigenous probiotic lactic acid bacteria. Indian J. Biochem. Biop. 2020, 57, 539–551. [Google Scholar]
- Zhang, J.; Zhao, X.; Jiang, Y.; Zhao, W.; Guo, T.; Cao, Y.; Yang, Z. Antioxidant status and gut microbiota change in an aging mouse model as influenced by exopolysaccharide produced by Lactobacillus plantarum YW11 isolated from Tibetan kefir. J. Dairy Sci. 2017, 100, 6025–6041. [Google Scholar] [CrossRef]
- Zhao, H.; Li, J.; Zhang, J.; Wang, X.; Hao, L.; Jia, L. Purification, in vitro Antioxidant and in vivo Anti-aging Activities of Exopolysaccharides by Agrocybe cylindracea. Int. J. Biol. Macromol. 2017, 102, 351–357. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.M.; Sun, X.Y.; Ouyang, J.M. Structural Characterization, Antioxidant Activity, and Biomedical Application of Astragalus Polysaccharide Degradation Products. Int. J. Polym. Sci. 2018, 2018, 5136185. [Google Scholar] [CrossRef]
- Andrew, M.; Jayaraman, G. Structural features of microbial exopolysaccharides in relation to their antioxidant activity. Carbohydr. Res. 2020, 487, 107881. [Google Scholar] [CrossRef]
- Li, S.; Huang, R.; Shah, N.P.; Tao, X.; Xiong, Y.; Wei, H. Antioxidant and antibacterial activities of exopolysaccharides from Bifidobacterium bifidum WBIN03 and Lactobacillus plantarum R315. J. Dairy Sci. 2014, 97, 7334–7343. [Google Scholar] [CrossRef]
- Mae, M.; Iyori, M.; Yasuda, M.; Mohammad Shamsul, H.; Kataoka, H.; Kiura, K.; Shibata, K.I. The diacylated lipopeptide FSL-1 enhances phagocytosis of bacteria by macrophages through a Toll-like receptor 2-mediated signalling pathway. FEMS Microbiol. Immunol. 2007, 49, 398–409. [Google Scholar] [CrossRef]
- Trabelsi, I.; Ktari, N.; Slima, S.B.; Triki, M.; Bardaa, S.; Mnif, H.; Salah, R.B. Evaluation of dermal wound healing activity and in vitro antibacterial and antioxidant activities of a new exopolysaccharide produced by Lactobacillus sp. Ca6. Int. J. Biol. Macromol. 2017, 103, 194–201. [Google Scholar] [CrossRef]
- Cao, H.; Sethumadhavan, K. Regulation of Cell Viability and Anti-inflammatory Tristetraprolin Family Gene Expression in Mouse Macrophages by Cottonseed extracts. Sci. Rep. 2020, 10, 775. [Google Scholar] [CrossRef] [PubMed]
- Jantaruk, P.; Roytrakul, S.; Sitthisak, S.; Kunthalert, D. Potential Role of an Antimicrobial Peptide, KLK in Inhibiting Lipopolysaccharide-induced Macrophage Inflammation. PLoS ONE 2017, 12, 183–852. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Xie, J.; Yang, Y.; Zhang, F.; Wang, S.; Wu, T.; Xie, M. Sulfated Cyclocarya paliurus Polysaccharides Markedly Attenuates Inflammation and Oxidative Damage in Lipopolysaccharide-treated Macrophage cells and Mice. Sci. Rep. 2017, 7, 40402. [Google Scholar] [CrossRef] [PubMed]
- Hidalgo-Cantabrana, C.; Sánchez, B.; Milani, C.; Ventura, M.; Margolles, A.; Ruas-Madiedo, P. Genomic overview and biological functions of exopolysaccharide biosynthesis in Bifidobacterium spp. Appl. Environ. Microbiol. 2014, 80, 9–18. [Google Scholar] [CrossRef]
- Roca, C.; Alves, V.D.; Freitas, F.; Reis, M.A. Exopolysaccharides Enriched in Rare Sugars: Bacterial sources, Production, and Applications. Front. Microbiol. 2015, 6, 288. [Google Scholar] [CrossRef]
- You, X.; Li, Z.; Ma, K.; Zhang, C.; Chen, X.; Wang, G.; Li, W. Structural characterization and immunomodulatory activity of an exopolysaccharide produced by Lactobacillus helveticus LZ-R-5. Carbohydr. Polym. 2020, 235, 115977. [Google Scholar] [CrossRef]
- Xu, W.; Zhao, M.; Fu, X.; Hou, J.; Wang, Y.; Shi, F.; Hu, S. Molecular mechanisms underlying macrophage immunomodulatory activity of Rubus chingii Hu polysaccharides. Int. J. Biol. Macromol. 2021, 185, 907–916. [Google Scholar] [CrossRef]
- Kaur, H.; Ali, S.A. Probiotics and gut microbiota: Mechanistic insights into gut immune homeostasis through TLR pathway regulation. Food Funct. 2022, 13, 7423–7447. [Google Scholar] [CrossRef]
- Kaur, H.; Kaur, G.; Ali, S.A. IL-33’s role in the gut immune system: A comprehensive review of its crosstalk and regulation. Life Sci. 2023, 327, 121868. [Google Scholar] [CrossRef]
- Kaur, H.; Ali, S.A.; Yan, F. Interactions between the gut microbiota-derived functional factors and intestinal epithelial cells–implication in the microbiota-host mutualism. Front. Immunol. 2022, 13, 1006081. [Google Scholar] [CrossRef] [PubMed]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kumari, M.; Haranahalli Nataraj, B.; Prasad, W.G.; Ali, S.A.; Behare, P.V. Multi-Faceted Bioactivity Assessment of an Exopolysaccharide from Limosilactobacillus fermentum NCDC400: Antioxidant, Antibacterial, and Immunomodulatory Proficiencies. Foods 2023, 12, 3595. https://doi.org/10.3390/foods12193595
Kumari M, Haranahalli Nataraj B, Prasad WG, Ali SA, Behare PV. Multi-Faceted Bioactivity Assessment of an Exopolysaccharide from Limosilactobacillus fermentum NCDC400: Antioxidant, Antibacterial, and Immunomodulatory Proficiencies. Foods. 2023; 12(19):3595. https://doi.org/10.3390/foods12193595
Chicago/Turabian StyleKumari, Manorama, Basavaprabhu Haranahalli Nataraj, Writdhama G. Prasad, Syed Azmal Ali, and Pradip V. Behare. 2023. "Multi-Faceted Bioactivity Assessment of an Exopolysaccharide from Limosilactobacillus fermentum NCDC400: Antioxidant, Antibacterial, and Immunomodulatory Proficiencies" Foods 12, no. 19: 3595. https://doi.org/10.3390/foods12193595
APA StyleKumari, M., Haranahalli Nataraj, B., Prasad, W. G., Ali, S. A., & Behare, P. V. (2023). Multi-Faceted Bioactivity Assessment of an Exopolysaccharide from Limosilactobacillus fermentum NCDC400: Antioxidant, Antibacterial, and Immunomodulatory Proficiencies. Foods, 12(19), 3595. https://doi.org/10.3390/foods12193595