
The Differences in Protein Degradation and Sensitization Reduction of Mangoes between Juices and Pieces Fermentation

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Fermentation of Mango Fruits
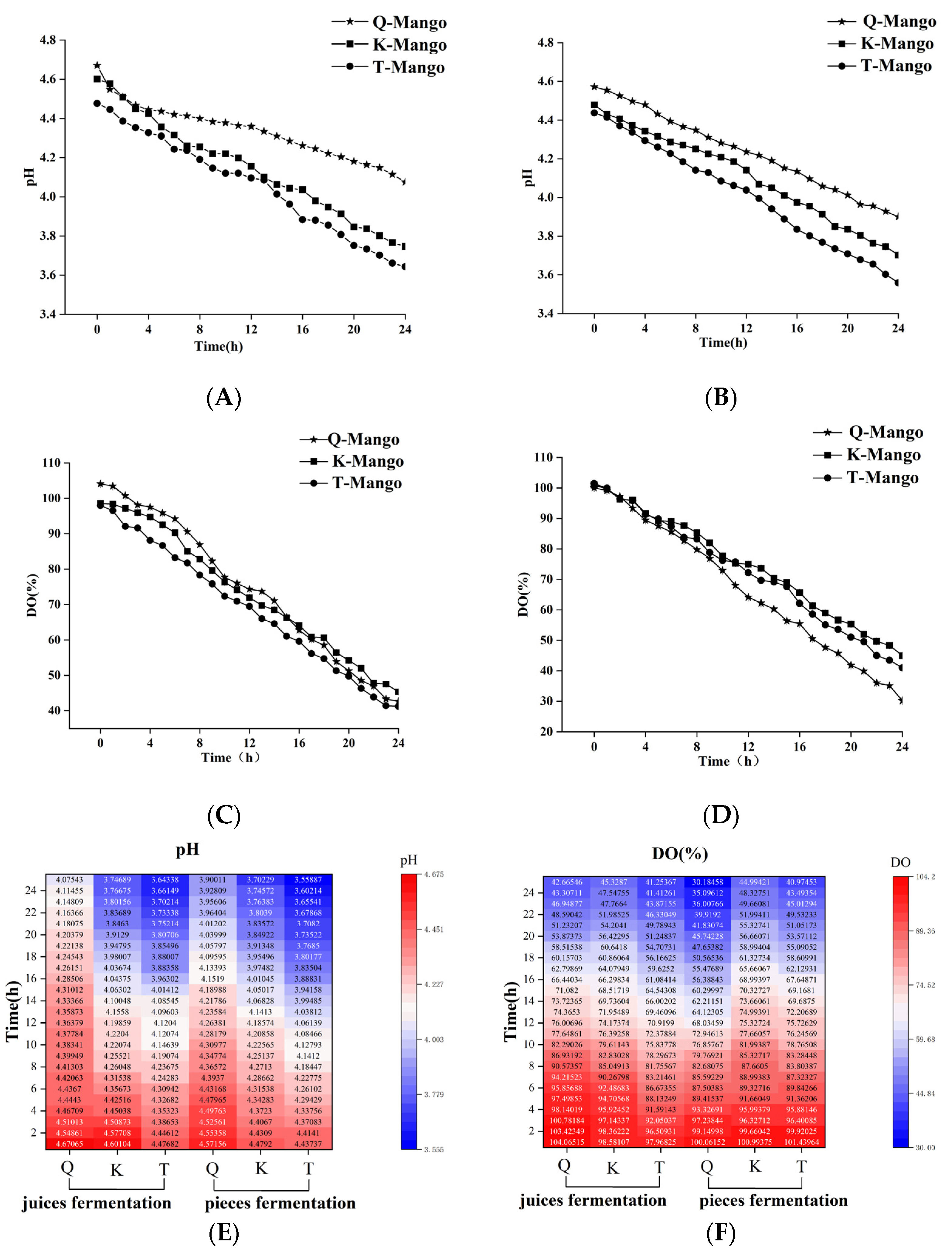
2.3. PH and Dissolved Oxygen (DO)
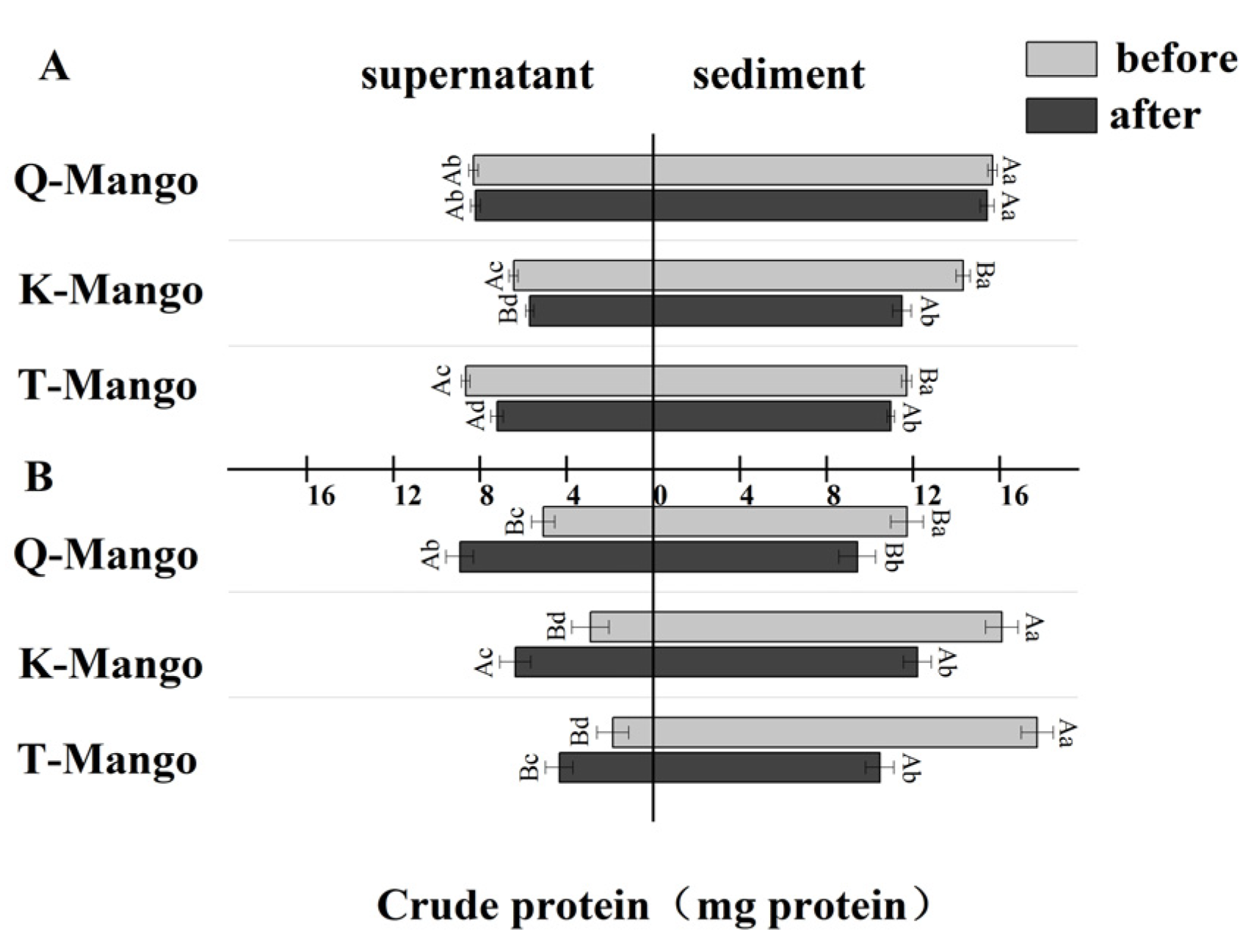
2.4. Crude Protein Content
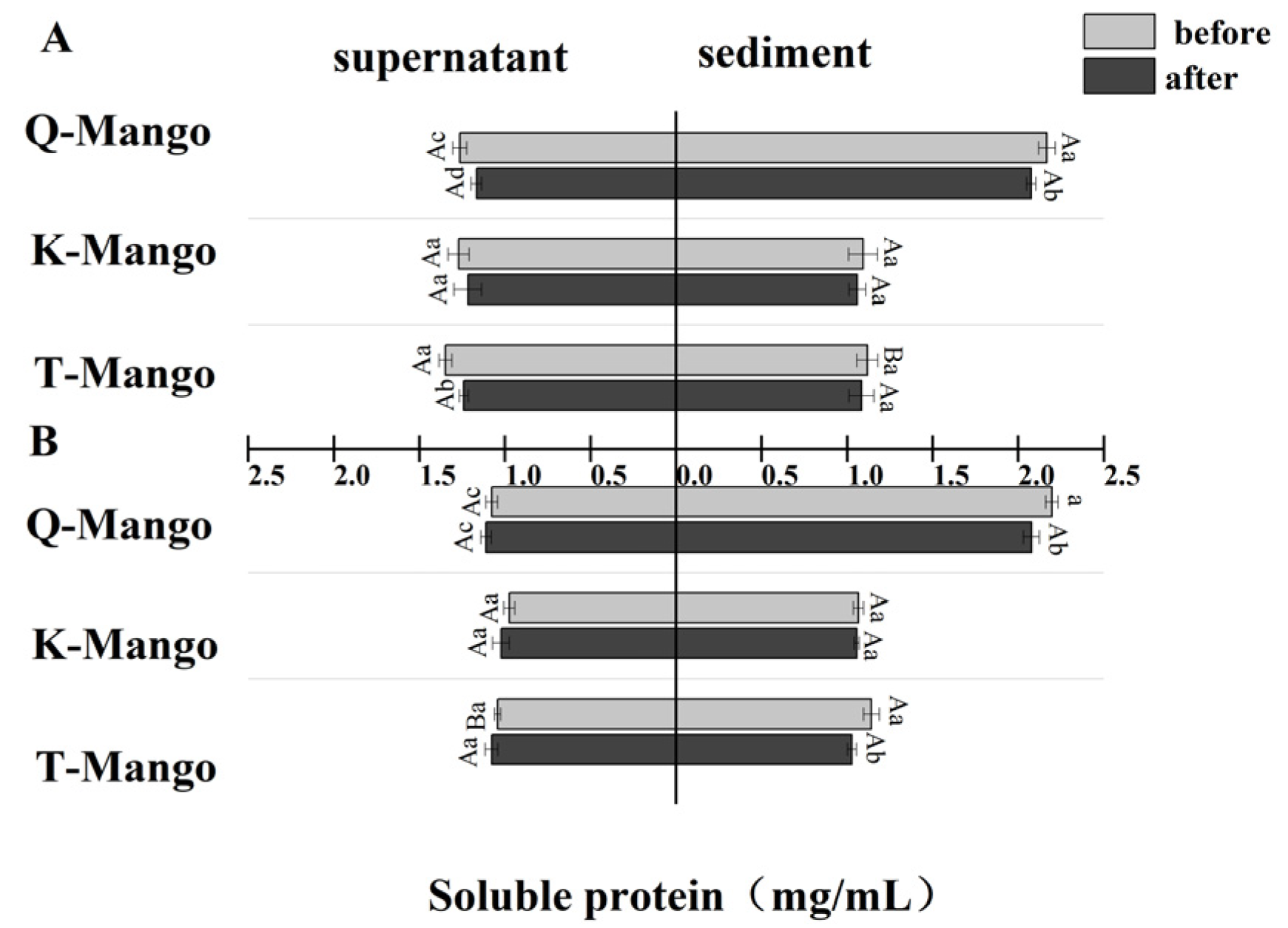
2.5. Soluble Protein Content
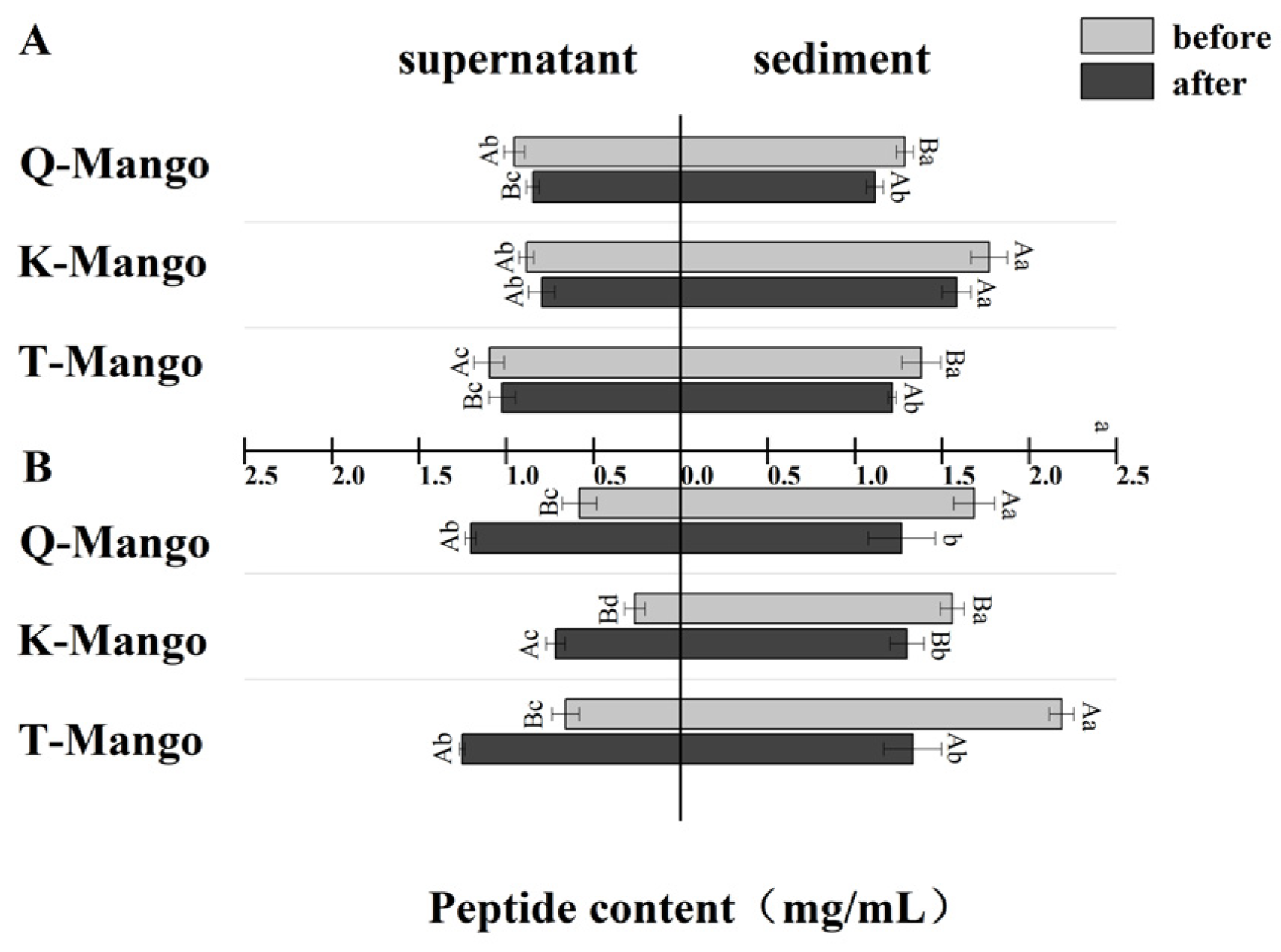
2.6. Peptide Content
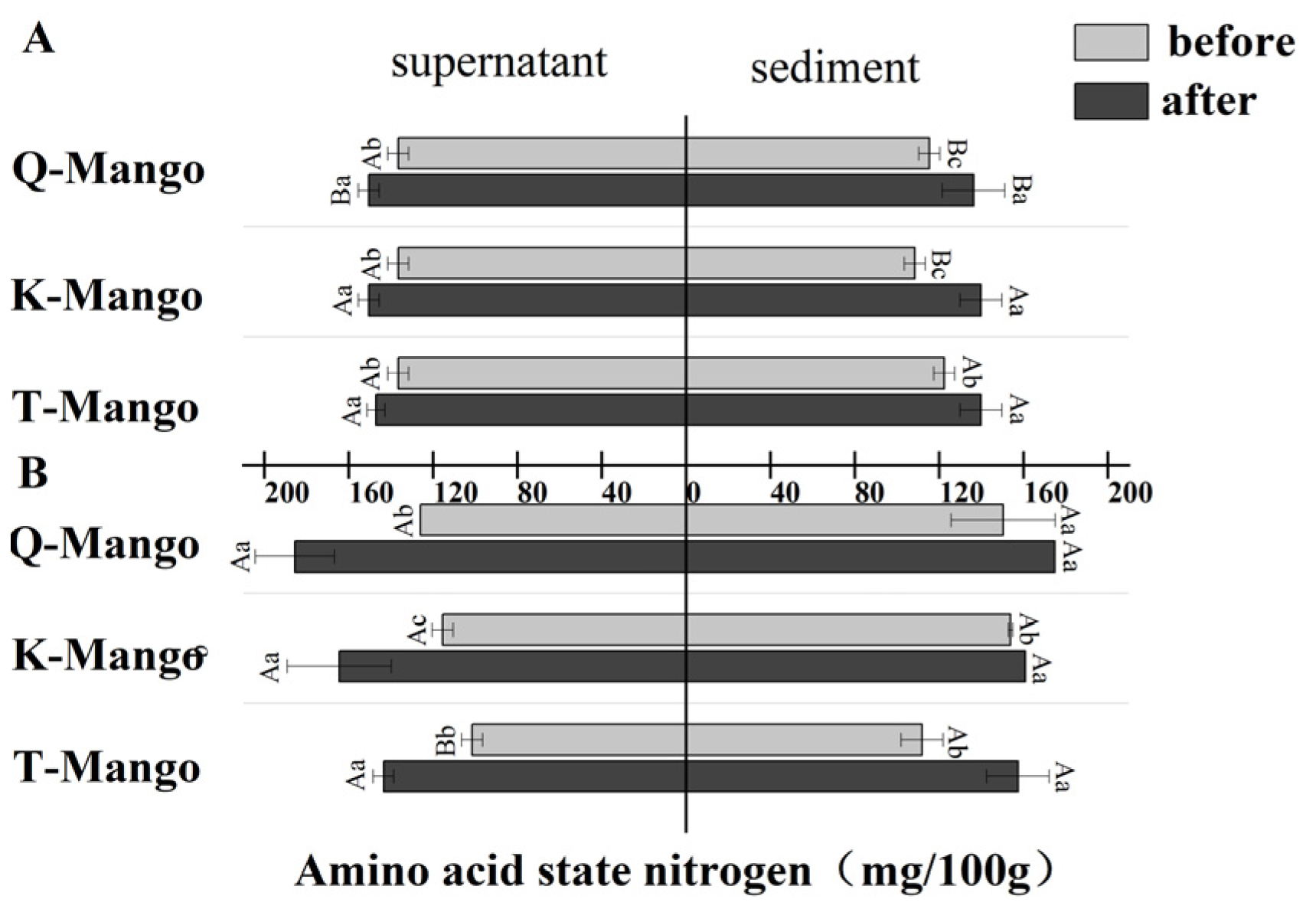
2.7. Amino Acid State Nitrogen
2.8. SDS-PAGE
2.9. IgE Immunoreactivity
2.10. Statistical Analyses
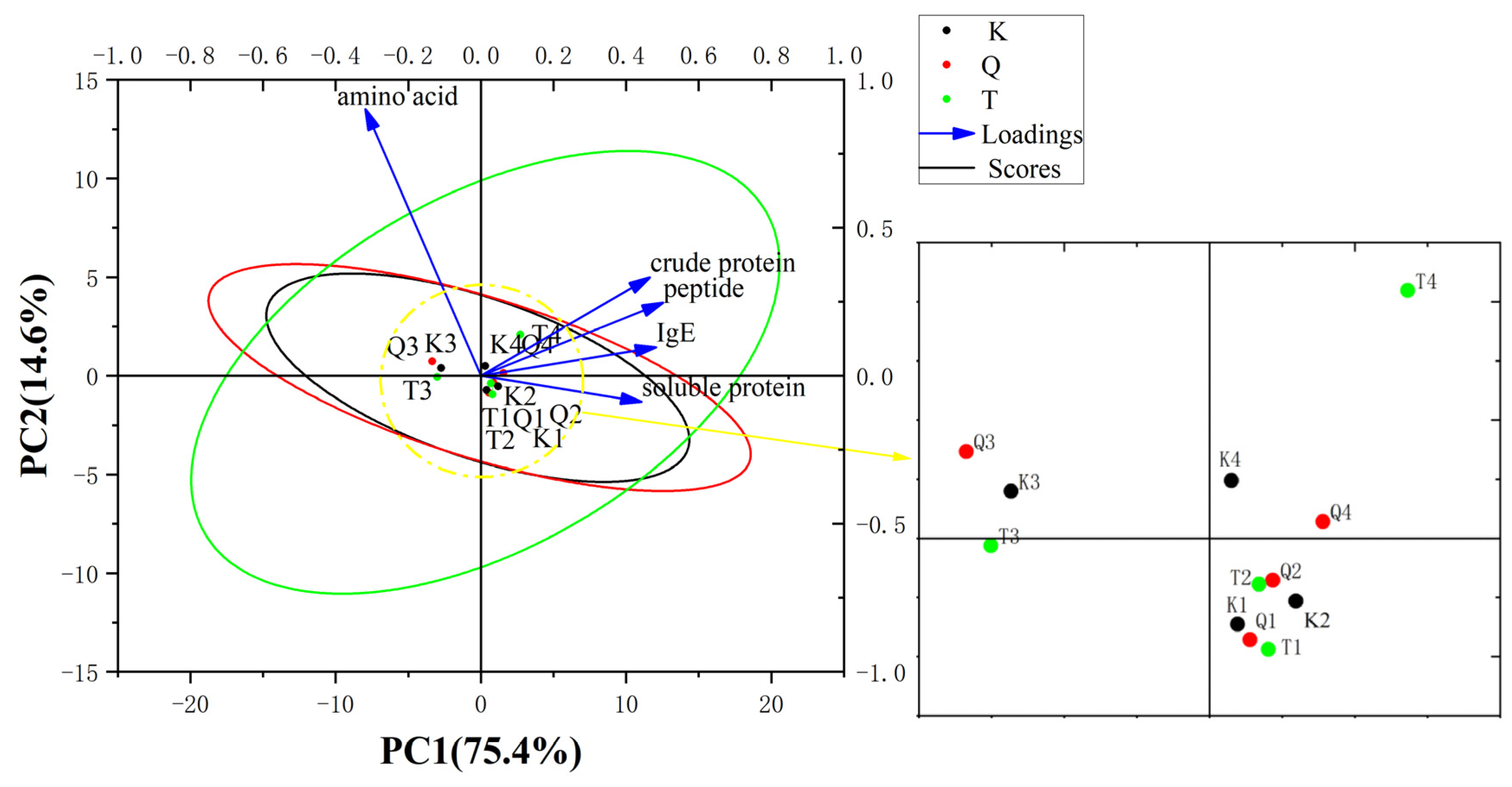
3. Results and Discussion
4. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Pawase, P. Standardization and formulation of fig mango mix fruit bar. Int. J. Chem. Res. 2018, 6, 394–398. Available online: https://www.chemijournal.com/archives/2018/vol6issue6/PartG/6-6-1-663.pdf (accessed on 28 September 2018).
- Evans, E.A.; Ballen, F.H.; Siddiq, M. Mango Production, Global Trade, Consumption Trends, and Postharvest Processing and Nutrition. In Handbook of Mango Fruit; Wiley Online Library: Hoboken, NJ, USA, 2017; pp. 1–16. [Google Scholar] [CrossRef]
- Food and Agriculture Organization (FAO). Statisitics. Available online: https://www.statista.com/statistics/577951/world-mango-production/ (accessed on 19 January 2023).
- Ntsoane, M.L.; Zude-Sasse, M.; Mahajan, P.; Sivakumar, D. Quality assesment and postharvest technology of mango: A review of its current status and future perspectives. Sci. Hortic. 2019, 249, 77–85. [Google Scholar] [CrossRef]
- Papanikolaou, Y.; Fulgoni, V.L. Mango consumption is associated with improved nutrient intakes, diet quality, and weight-related health outcomes. Nutrients 2022, 14, 59. [Google Scholar] [CrossRef]
- Rosas, M.; Pinneo, S.; O’Mealy, C.; Tsang, M.; Liu, C.; Kern, M.; Hooshmand, S.; Hong, M.Y. Effects of fresh mango consumption on cardiometabolic risk factors in overweight and obese adults. Nutr. Metab. Cardiovasc. Dis. 2022, 32, 494–503. [Google Scholar] [CrossRef]
- Asuncion, P.; Liu, C.; Castro, R.; Yon, V.; Rosas Jr, M.; Hooshmand, S.; Kern, M.; Hong, M.Y. The effects of fresh mango consumption on gut health and microbiome—Randomized controlled trial. Food Sci. Nutr. 2023, 11, 2069–2078. [Google Scholar] [CrossRef] [PubMed]
- Berghea, E.C.; Craiu, M.; Ali, S.; Corcea, S.L.; Bumbacea, R.S. Contact allergy induced by mango (Mangifera indica): A relevant topic? Medicina 2021, 57, 1240. [Google Scholar] [CrossRef] [PubMed]
- Paschke, A.; Kinder, H.; Zunker, K.; Wigotzki, M.; Steinhart, H.; Weßbecher, R.; Vieluf, I. Characterization of cross-reacting allergens in mango fruit. Allergy 2001, 56, 237–242. [Google Scholar] [CrossRef]
- Cardona, E.E.G.; Heathcote, K.; Teran, L.M.; Righetti, P.G.; Boschetti, E.; D’Amato, A. Novel low-abundance allergens from mango via combinatorial peptide libraries treatment: A proteomics study. Food Chem. 2018, 269, 652–660. [Google Scholar] [CrossRef]
- Tsai, W.-C.; Wu, T.-C.; Chiang, B.-L.; Wen, H.-W. Cloning, expression, and purification of recombinant major mango allergen Man i 1 in Escherichia coli. Protein Expr. Purif. 2017, 130, 35–43. [Google Scholar] [CrossRef]
- Paschke, A.; Kinder, H.; Zunker, K.; Wigotzki, M.; Wessbecher, R.; Vieluf, D.; Steinhart, H. Characterization of allergens in mango fruit and ripening dependence of the allergenic potency. Food Agric. Immunol. 2001, 13, 51–61. [Google Scholar] [CrossRef]
- Honglei, G.; Yanjun, C. Recent advances in the study of epitopes, allergens and immunologic cross-reactivity of edible mango. Food Sci. Hum. Wellness 2023, 13. [Google Scholar] [CrossRef]
- Trogen, B.; Jacobs, S.; Nowak-Wegrzyn, A. Early Introduction of Allergenic Foods and the Prevention of Food Allergy. Nutrients 2022, 14, 2565. [Google Scholar] [CrossRef] [PubMed]
- Sampath, V.; Abrams, E.M.; Adlou, B.; Akdis, C.; Akdis, M.; Brough, H.A.; Chan, S.; Chatchatee, P.; Chinthrajah, R.S.; Cocco, R.R.; et al. Food allergy across the globe. J. Allergy Clin. Immunol. 2021, 148, 1347–1364. [Google Scholar] [CrossRef] [PubMed]
- Pi, X.; Wan, Y.; Yang, Y.; Li, R.; Wu, X.; Xie, M.; Li, X.; Fu, G. Research progress in peanut allergens and their allergenicity reduction. Trends Food Sci. Technol. 2019, 93, 212–220. [Google Scholar] [CrossRef]
- Rahaman, T.; Vasiljevic, T.; Ramchandran, L. Effect of processing on conformational changes of food proteins related to allergenicity. Trends Food Sci. Technol. 2016, 49, 24–34. [Google Scholar] [CrossRef]
- Pi, X.; Sun, Y.; Cheng, J.; Fu, G.; Guo, M. A review on polyphenols and their potential application to reduce food allergenicity. Crit. Rev. Food Sci. Nutr. 2022, 1–18. [Google Scholar] [CrossRef]
- Ahmed, I.; Chen, H.; Li, J.; Wang, B.; Li, Z.; Huang, G. Enzymatic crosslinking and food allergenicity: A comprehensive review. Compr. Rev. Food Sci. Food Saf. 2021, 20, 5856–5879. [Google Scholar] [CrossRef]
- Pi, X.; Sun, Y.; Fu, G.; Wu, Z.; Cheng, J. Effect of processing on soybean allergens and their allergenicity. Trends Food Sci. Technol. 2021, 118, 316–327. [Google Scholar] [CrossRef]
- Pi, X.; Yang, Y.; Sun, Y.; Cui, Q.; Wan, Y.; Fu, G.; Chen, H.; Cheng, J. Recent advances in alleviating food allergenicity through fermentation. Crit. Rev. Food Sci. Nutr. 2022, 62, 7255–7268. [Google Scholar] [CrossRef]
- Yildiz, G.; Aadil, R.M. Comparative analysis of antibrowning agents, hot water and high-intensity ultrasound treatments to maintain the quality of fresh-cut mangoes. J. Food Sci. Technol. 2022, 59, 202–211. [Google Scholar] [CrossRef]
- Dube, M.; Zunker, K.; Neidhart, S.; Carle, R.; Steinhart, H.; Paschke, A. Effect of technological processing on the allergenicity of mangoes (Mangifera indica L.). J. Agric. Food Chem. 2004, 52, 3938–3945. [Google Scholar] [CrossRef] [PubMed]
- El Mecherfi, K.-E.; Todorov, S.D.; Cavalcanti de Albuquerque, M.A.; Denery-Papini, S.; Lupi, R.; Haertlé, T.; Dora Gombossy de Melo Franco, B.; Larré, C. Allergenicity of fermented foods: Emphasis on seeds protein-based products. Foods 2020, 9, 792. [Google Scholar] [CrossRef] [PubMed]
- El-Ghaish, S.; Ahmadova, A.; Hadji-Sfaxi, I.; El Mecherfi, K.E.; Bazukyan, I.; Choiset, Y.; Rabesona, H.; Sitohy, M.; Popov, Y.G.; Kuliev, A.A.; et al. Potential use of lactic acid bacteria for reduction of allergenicity and for longer conservation of fermented foods. Trends Food Sci. Technol. 2011, 22, 509–516. [Google Scholar] [CrossRef]
- Emkani, M.; Oliete, B.; Saurel, R. Effect of lactic acid fermentation on legume protein properties, a review. Fermentation 2022, 8, 244. [Google Scholar] [CrossRef]
- Fu, W.; Xue, W.; Liu, C.; Tian, Y.; Zhang, K.; Zhu, Z. Screening of lactic acid bacteria and yeasts from sourdough as starter cultures for reduced allergenicity wheat products. Foods 2020, 9, 751. [Google Scholar] [CrossRef] [PubMed]
- Anggraini, H.; Tongkhao, K.; Chanput, W. Reducing milk allergenicity of cow, buffalo, and goat milk using lactic acid bacteria fermentation. AIP Conf. Proc. 2018, 2021, 070010-1–070010-8. [Google Scholar] [CrossRef]
- El Mecherfi, K.-E.; Lupi, R.; Cherkaoui, M.; Albuquerque, M.A.C.; Todorov, S.D.; Tranquet, O.; Klingebiel, C.; Rogniaux, H.; Denery-Papini, S.; Onno, B.; et al. Fermentation of gluten by Lactococcus lactis LLGKC18 reduces its antigenicity and allergenicity. Probiotics Antimicrob. Proteins 2022, 14, 779–791. [Google Scholar] [CrossRef] [PubMed]
- Licandro, H.; Ho, P.H.; Nguyen, T.K.C.; Petchkongkaew, A.; Nguyen, H.V.; Chu-Ky, S.; Nguyen, T.V.A.; Lorn, D.; Waché, Y. How fermentation by lactic acid bacteria can address safety issues in legumes food products? Food Control 2020, 110, 106957. [Google Scholar] [CrossRef]
- Mandha, J.; Shumoy, H.; Devaere, J.; Schouteten, J.J.; Gellynck, X.; De Winne, A.; Matemu, A.O.; Raes, K. Effect of lactic acid fermentation on volatile compounds and sensory characteristics of mango (Mangifera indica) juices. Foods 2022, 11, 383. [Google Scholar] [CrossRef]
- Sadineni, V.; Kondapalli, N.; Obulam, V.S.R. Effect of co-fermentation with Saccharomyces cerevisiae and Torulaspora delbrueckii or Metschnikowia pulcherrima on the aroma and sensory properties of mango wine. Ann. Microbiol. 2012, 62, 1353–1360. [Google Scholar] [CrossRef]
- Akbarirad, H.; Mazaheri Assadi, M.; Pourahmad, R.; Mousavi Khaneghah, A. Employing of the different fruit juices substrates in vinegar kombucha preparation. Curr. Nutr. Food Sci. 2017, 13, 303–308. [Google Scholar] [CrossRef]
- Jung, S.; Rickert, D.A.; Deak, N.A.; Aldin, E.D.; Recknor, J.; Johnson, L.A.; Murphy, P.A. Comparison of kjeldahl and dumas methods for determining protein contents of soybean products. J. Am. Oil Chem. Soc. 2003, 80, 1169. [Google Scholar] [CrossRef]
- Smith, P.K.; Krohn, R.I.; Hermanson, G.T.; Mallia, A.K.; Gartner, F.H.; Provenzano, M.D.; Fujimoto, E.K.; Goeke, N.M.; Olson, B.J.; Klenk, D.C. Measurement of protein using bicinchoninic acid. Anal. Biochem. 1985, 150, 76–85. [Google Scholar] [CrossRef] [PubMed]
- Rui, X.; Huang, J.; Xing, G.; Zhang, Q.; Li, W.; Dong, M. Changes in soy protein immunoglobulin E reactivity, protein degradation, and conformation through fermentation with Lactobacillus plantarum strains. Lwt 2019, 99, 156–165. [Google Scholar] [CrossRef]
- Su, X.; Zhang, Z.; Zhang, Q.; Yang, J.; Gao, M.; Hu, H.; Liu, D. Optimization of yeast autolysis under solid-state fermentation conditions. Sheng Wu Gong Cheng Xue Bao = Chin. J. Biotechnol. 2019, 35, 726–736. [Google Scholar] [CrossRef]
- Huang, J.; Liu, Z.; Rui, X.; L’Hocine, L.; Zhang, Q.Q.; Li, W.; Dong, M.S. Assessment of the effect of lactic acid fermentation on the gastroduodenal digestibility and immunoglobulin E binding capacity of soy proteins via an in vitro dynamic gastrointestinal digestion model. Food Funct. 2020, 11, 10467–10479. [Google Scholar] [CrossRef]
- Peñas, E.; Gomez, R.; Frias, J.; Baeza, M.L.; Vidal-Valverde, C. High hydrostatic pressure effects on immunoreactivity and nutritional quality of soybean products. Food Chem. 2011, 125, 423–429. [Google Scholar] [CrossRef]
- Kaashyap, M.; Cohen, M.; Mantri, N. Microbial diversity and characteristics of kombucha as revealed by metagenomic and physicochemical analysis. Nutrients 2021, 13, 4446. [Google Scholar] [CrossRef]
- Reva, O.N.; Zaets, I.E.; Ovcharenko, L.P.; Kukharenko, O.E.; Shpylova, S.P.; Podolich, O.V.; de Vera, J.P.; Kozyrovska, N.O. Metabarcoding of the kombucha microbial community grown in different microenvironments. AMB Express 2015, 5, 35. [Google Scholar] [CrossRef]
- Landis Elizabeth, A.; Fogarty, E.; Edwards John, C.; Popa, O.; Eren, A.M.; Wolfe Benjamin, E. Microbial diversity and interaction specificity in kombucha tea fermentations. mSystems 2022, 7, e00157-22. [Google Scholar] [CrossRef]
- Wang, B.; Rutherfurd-Markwick, K.; Zhang, X.-X.; Mutukumira, A.N. Kombucha: Production and microbiological research. Foods 2022, 11, 3456. [Google Scholar] [CrossRef] [PubMed]
- Tran, T.; Grandvalet, C.; Verdier, F.; Martin, A.; Alexandre, H.; Tourdot-Maréchal, R. Microbial dynamics between yeasts and acetic acid bacteria in kombucha: Impacts on the chemical composition of the beverage. Foods 2020, 9, 963. [Google Scholar] [CrossRef] [PubMed]
- Villarreal-Soto, S.A.; Beaufort, S.; Bouajila, J.; Souchard, J.-P.; Renard, T.; Rollan, S.; Taillandier, P. Impact of fermentation conditions on the production of bioactive compounds with anticancer, anti-inflammatory and antioxidant properties in kombucha tea extracts. Process Biochem. 2019, 83, 44–54. [Google Scholar] [CrossRef]
- Villarreal-Soto, S.A.; Beaufort, S.; Bouajila, J.; Souchard, J.-P.; Taillandier, P. Understanding kombucha tea fermentation: A review. J. Food Sci. 2018, 83, 580–588. [Google Scholar] [CrossRef]
- Bourdichon, F.; Casaregola, S.; Farrokh, C.; Frisvad, J.C.; Gerds, M.L.; Hammes, W.P.; Harnett, J.; Huys, G.; Laulund, S.; Ouwehand, A.; et al. Food fermentations: Microorganisms with technological beneficial use. Int. J. Food Microbiol. 2012, 154, 87–97. [Google Scholar] [CrossRef]
- Christensen, L.F.; García-Béjar, B.; Bang-Berthelsen, C.H.; Hansen, E.B. Extracellular microbial proteases with specificity for plant proteins in food fermentation. Int. J. Food Microbiol. 2022, 381, 109889. [Google Scholar] [CrossRef]
- Gänzle, M.G. Food fermentations for improved digestibility of plant foods—An essential ex situ digestion step in agricultural societies? Curr. Opin. Food Sci. 2020, 32, 124–132. [Google Scholar] [CrossRef]
- Hrnjez, D.; Vaštag, Ž.; Milanović, S.; Vukić, V.; Iličić, M.; Popović, L.; Kanurić, K. The biological activity of fermented dairy products obtained by kombucha and conventional starter cultures during storage. J. Funct. Foods 2014, 10, 336–345. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| IgE (OD450) | Mango * | Juices Fermentation | Pieces Fermentation | ||||||
|---|---|---|---|---|---|---|---|---|---|
| Supernatant | Sediment | Supernatant | Sediment | ||||||
| Before | After | Before | After | Before | After | Before | After | ||
| IgE1 | Q | 0.68 ± 0.02 Aa | 0.64 ± 0.03 Aa | 0.85 ± 0.02 Aa | 0.73 ± 0.02 Ab | 0.43 ± 0.02 Bb | 0.54 ± 0.04 Aa | 0.86 ± 0.02 Aa | 0.75 ± 0.01 Ab |
| K | 0.66 ± 0.03 Aa | 0.61 ± 0.01 Ab | 0.83 ± 0.02 Ba | 0.79 ± 0.01 Aa | 0.50 ± 0.01 Bb | 0.56 ± 0.01 Aa | 0.95 ± 0.02 Aa | 0.83 ± 0.01 Ab | |
| T | 0.43 ± 0.03 Aa | 0.40 ± 0.01 Ba | 0.57 ± 0.03 Ba | 0.42 ± 0.04 Bb | 0.50 ± 0.01 Ab | 0.66 ± 0.01 Aa | 0.78 ± 0.02 Aa | 0.59 ± 0.06 Ab | |
| IgE2 | Q | 0.22 ± 0.02 Aa | 0.16 ± 0.02 Bb | 0.46 ± 0.02 Aa | 0.36 ± 0.01 Ab | 0.30 ± 0.01 Ab | 0.56 ± 0.02 Aa | 0.42 ± 0.04 Aa | 0.31 ± 0.01 Ab |
| K | 0.26 ± 0.02 Aa | 0.23 ± 0.05 Aa | 0.46 ± 0.01 Aa | 0.28 ± 0.02 Ab | 0.27 ± 0.04 Aa | 0.31 ± 0.03 Aa | 0.38 ± 0.01 Aa | 0.35 ± 0.05 Aa | |
| T | 0.43 ± 0.01 Aa | 0.36 ± 0.01 Ab | 0.51 ± 0.01 Ba | 0.39 ± 0.01 Bb | 0.23 ± 0.06 Bb | 0.40 ± 0.02 Aa | 0.65 ± 0.01 Aa | 0.58 ± 0.03 Ab | |
| IgE3 | Q | 0.19 ± 0.02 Aa | 0.16 ± 0.01 Aa | 0.30 ± 0.01 Aa | 0.20 ± 0.01 Ab | 0.21 ± 0.06 Aa | 0.23 ± 0.09 Aa | 0.26 ± 0.07 Aa | 0.22 ± 0.04 Aa |
| K | 0.39 ± 0.05 Aa | 0.30 ± 0.06 Ab | 0.41 ± 0.1 Aa | 0.38 ± 0.1 Aa | 0.28 ± 0.06 Ab | 0.39 ± 0.1 Aa | 0.44 ± 0.2 Aa | 0.29 ± 0.08 Ab | |
| T | 0.26 ± 0.03 Aa | 0.19 ± 0.02 Ab | 0.24 ± 0.02 Aa | 0.17 ± 0.01 Ab | 0.18 ± 0.01 Aa | 0.20 ± 0.04 Aa | 0.30 ± 0.1 Aa | 0.22 ± 0.04 Ab | |
| IgE4 | Q | 0.32 ± 0.01 Aa | 0.29 ± 0.01 Aa | 0.21 ± 0.01 Aa | 0.17 ± 0.01 Aa | 0.18 ± 0.01 Bb | 0.23 ± 0.01 Aa | 0.21 ± 0.01 Aa | 0.17 ± 0.01 Aa |
| K | 0.30 ± 0.01 Aa | 0.27 ± 0.01 Aa | 0.34 ± 0.8 Aa | 0.18 ± 0.1 Ab | 0.12 ± 0.01 Ba | 0.15 ± 0.01 Ba | 0.19 ± 0.01 Ba | 0.18 ± 0.01 Aa | |
| T | 0.41 ± 0.01 Aa | 0.40 ± 0.1 Aa | 0.20 ± 0.01 Aa | 0.18 ± 0.01 Aa | 0.15 ± 0.01 Bb | 0.22 ± 0.01 Ba | 0.23 ± 0.01 Aa | 0.16 ± 0.01 Ab | |
| IgE5 | Q | 0.26 ± 0.01 Aa | 0.23 ± 0.01 Aa | 0.27 ± 0.01 Aa | 0.26 ± 0.03 Aa | 0.24 ± 0.02 Aa | 0.26 ± 0.01 Aa | 0.34 ± 0.01 Aa | 0.29 ± 0.01 Ab |
| K | 0.24 ± 0.02 Aa | 0.21 ± 0.02 Aa | 0.28 ± 0.9 Aa | 0.24 ± 0.2 Aa | 0.24 ± 0.02 Aa | 0.24 ± 0.01 Aa | 0.27 ± 0.01 Aa | 0.25 ± 0.02 Aa | |
| T | 0.26 ± 0.06 Aa | 0.20 ± 0.3 Ab | 0.23 ± 0.02 Aa | 0.20 ± 0.07 Aa | 0.25 ± 0.01 Aa | 0.26 ± 0.04 Aa | 0.3 ± 0.01 Aa | 0.29 ± 0.01 Ab | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tian, M.; Zhang, Q.; Zeng, X.; Rui, X.; Jiang, M.; Chen, X. The Differences in Protein Degradation and Sensitization Reduction of Mangoes between Juices and Pieces Fermentation. Foods 2023, 12, 3465. https://doi.org/10.3390/foods12183465
Tian M, Zhang Q, Zeng X, Rui X, Jiang M, Chen X. The Differences in Protein Degradation and Sensitization Reduction of Mangoes between Juices and Pieces Fermentation. Foods. 2023; 12(18):3465. https://doi.org/10.3390/foods12183465
Chicago/Turabian StyleTian, Mengtian, Qiuqin Zhang, Xianming Zeng, Xin Rui, Mei Jiang, and Xiaohong Chen. 2023. "The Differences in Protein Degradation and Sensitization Reduction of Mangoes between Juices and Pieces Fermentation" Foods 12, no. 18: 3465. https://doi.org/10.3390/foods12183465
APA StyleTian, M., Zhang, Q., Zeng, X., Rui, X., Jiang, M., & Chen, X. (2023). The Differences in Protein Degradation and Sensitization Reduction of Mangoes between Juices and Pieces Fermentation. Foods, 12(18), 3465. https://doi.org/10.3390/foods12183465