Enhancement of Textural and Sensory Characteristics of Wheat Bread Using a Chickpea Sourdough Fermented with a Selected Autochthonous Microorganism

 ,
, 
 , and
, and
Abstract
:
1. Introduction
2. Materials and Methods
2.1. Raw Materials
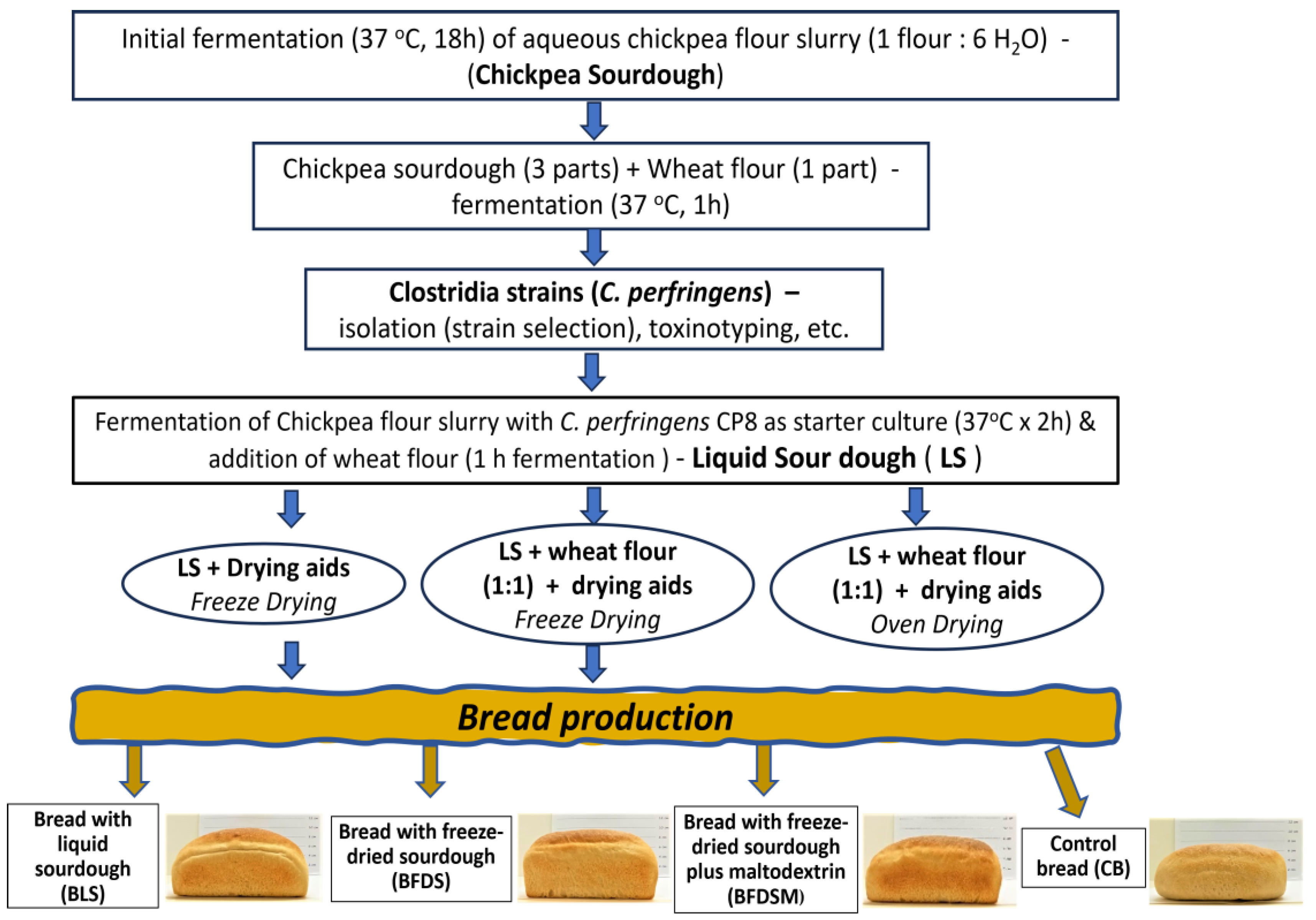
2.2. Preparation of Traditional Chickpea Sourdough
2.3. Isolation and Identification of Indigenous Clostridial Strains
2.3.1. Identification of Clostridia Isolates by SDS-PAGE of Cellular Proteins and 16S rDNA PCR
2.3.2. Toxinotyping Test by Multiplex PCR
2.4. Fermentation of Chickpea Flour with Clostridium Isolates as Starter Cultures
2.5. Dried Sourdough Making
2.6. Total Titratable Acidity and pH Determination
2.7. Breadmaking Procedure
2.8. Physical Properties of Breads
2.9. Organic Acids Analysis by HPLC
2.9.1. Sample Preparation
2.9.2. Equipment and Analytical Procedure
2.10. Differential Scanning Calorimetry (DSC)
2.11. Shelf Life Estimated by Mold Growth
2.12. FTIR Spectroscopy Analysis
2.13. Sensory Analysis of Breads
2.14. Statistical Analysis
3. Results and Discussions
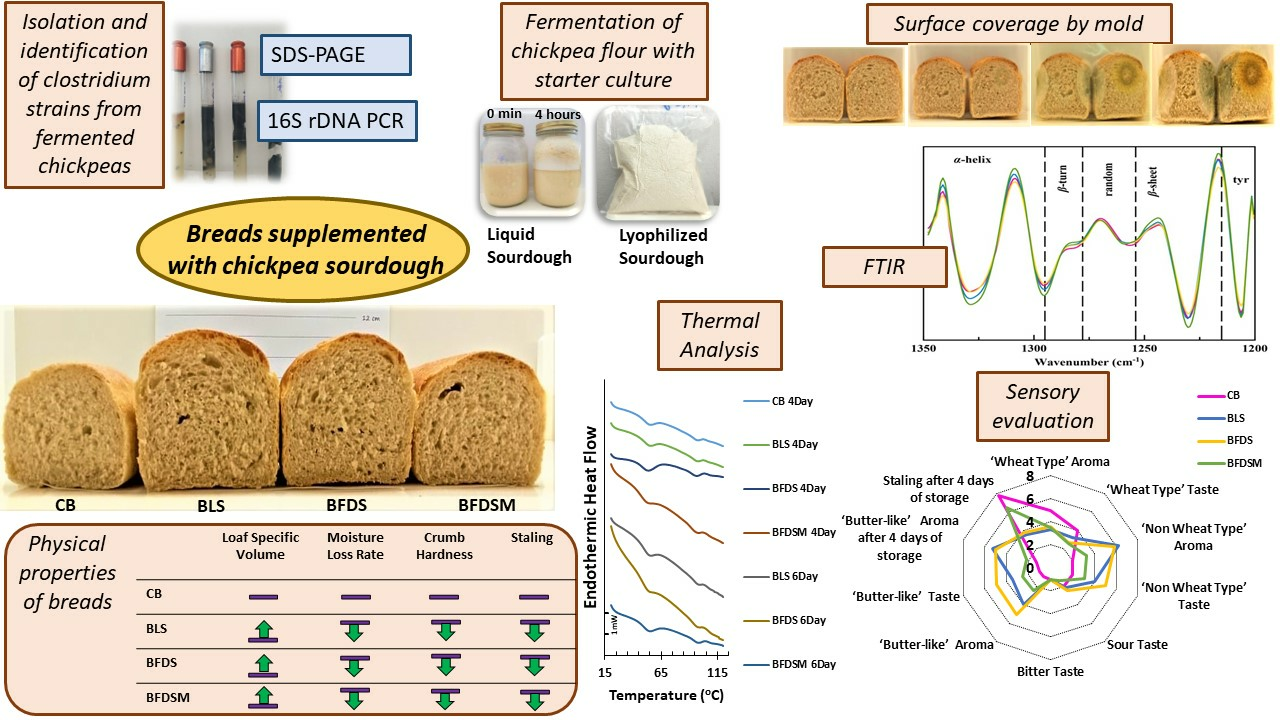
3.1. Identification of Clostridia from Chickpea Sourdough
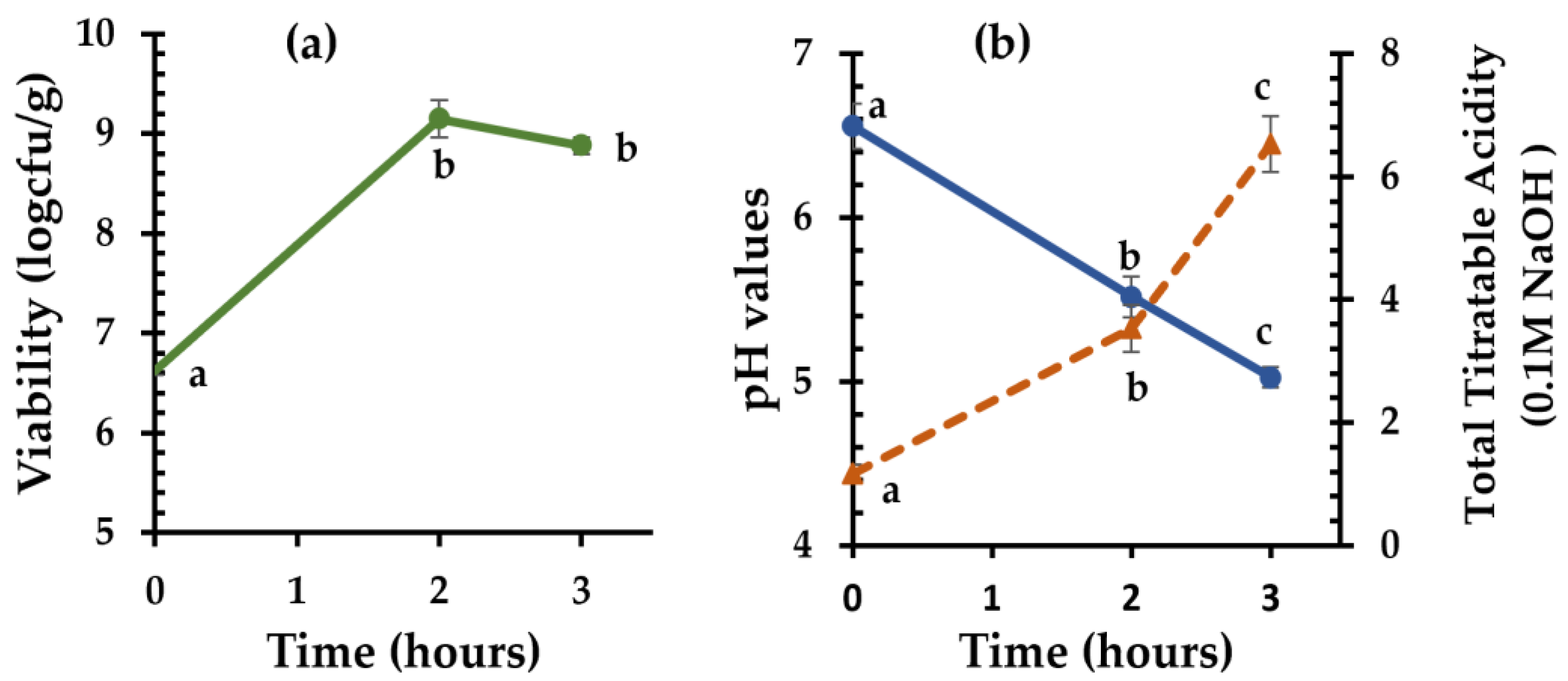
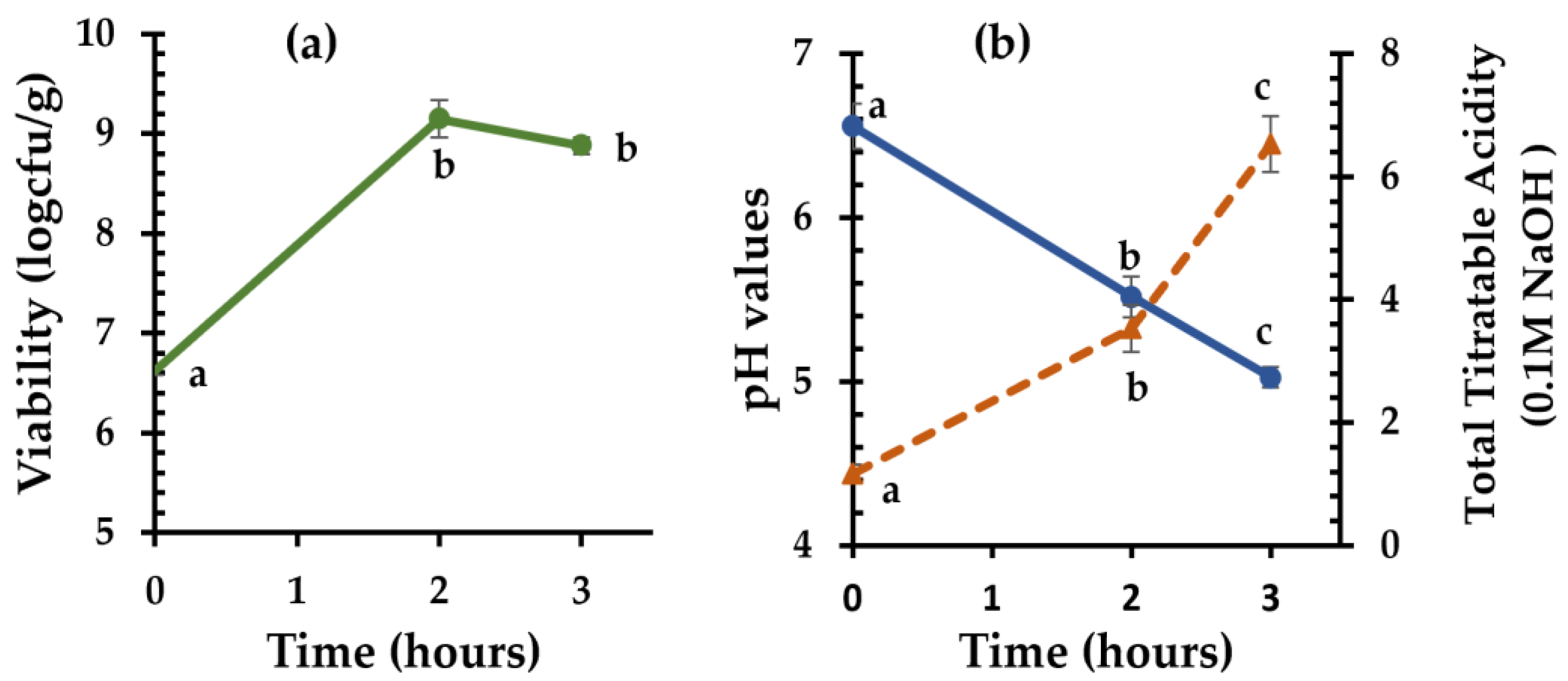
3.2. Sourdough with Clostridia as a Starter Culture
3.3. Drying of Liquid Sourdough (LS)
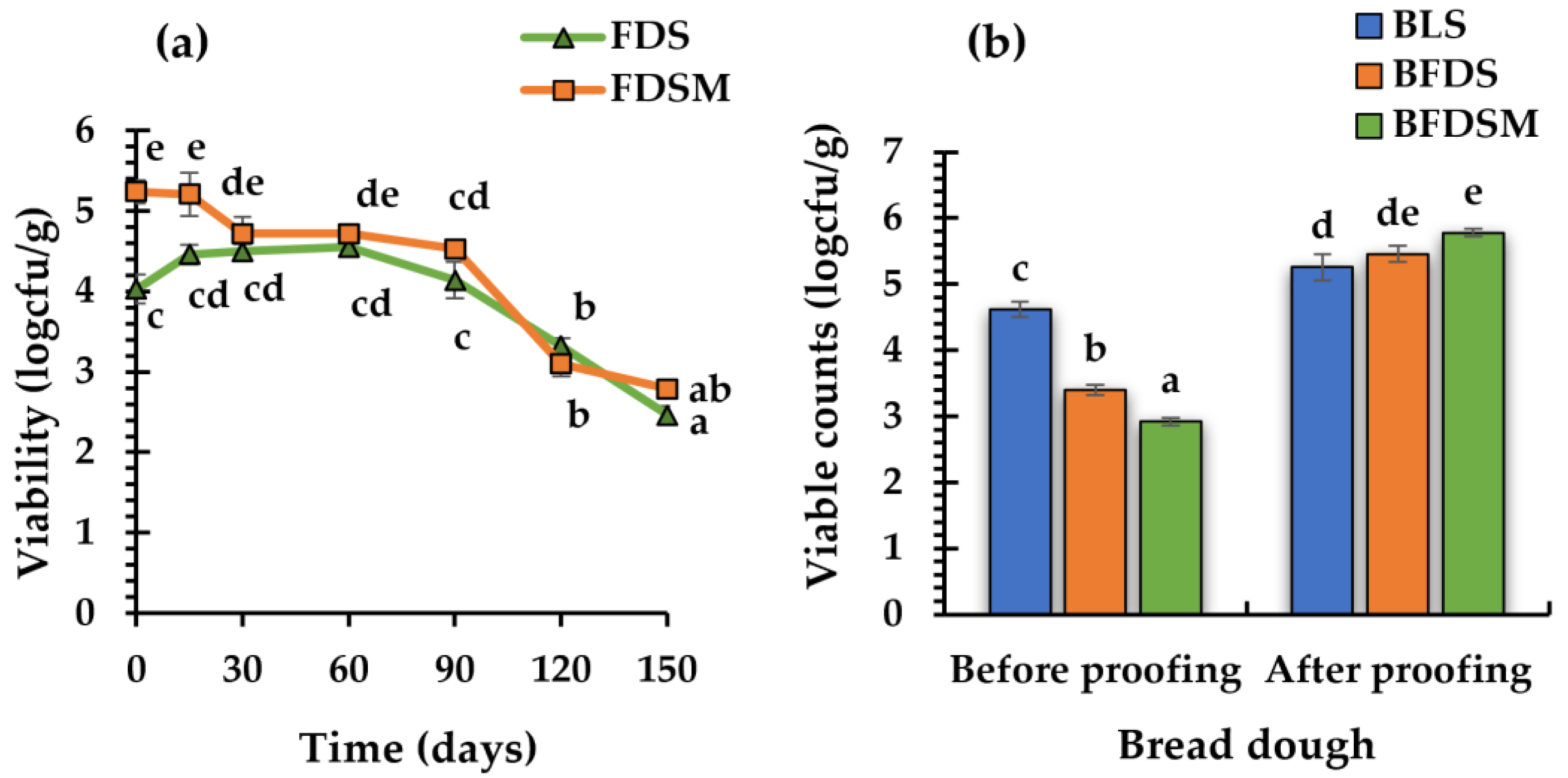
3.4. Storage Stability of Freeze-Dried Sourdoughs
3.5. C. Perfringens CP8 Growth during Breadmaking
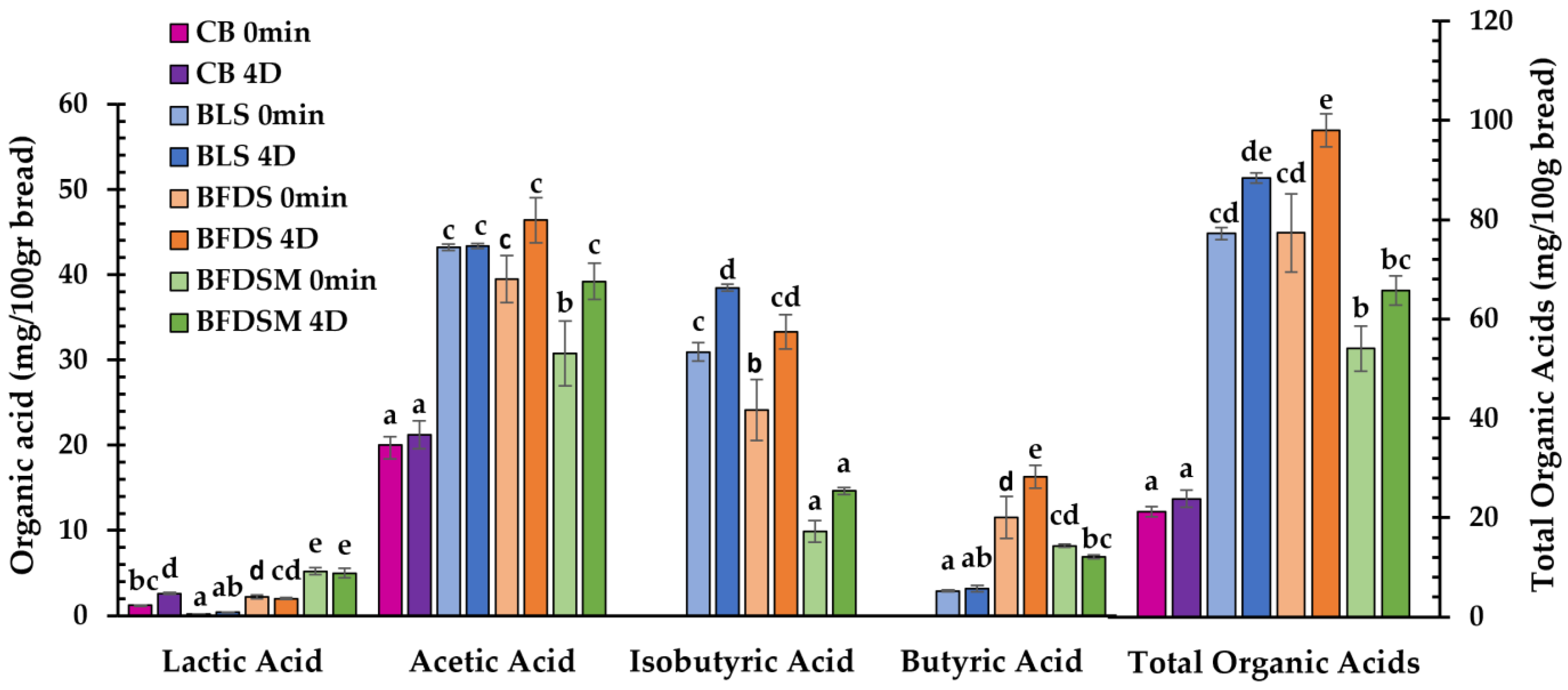
3.6. Organic Acids in Sourdough and Bread
3.7. Loaf-Specific Volume
3.8. Color Parameters
3.9. Moisture Content of Breads
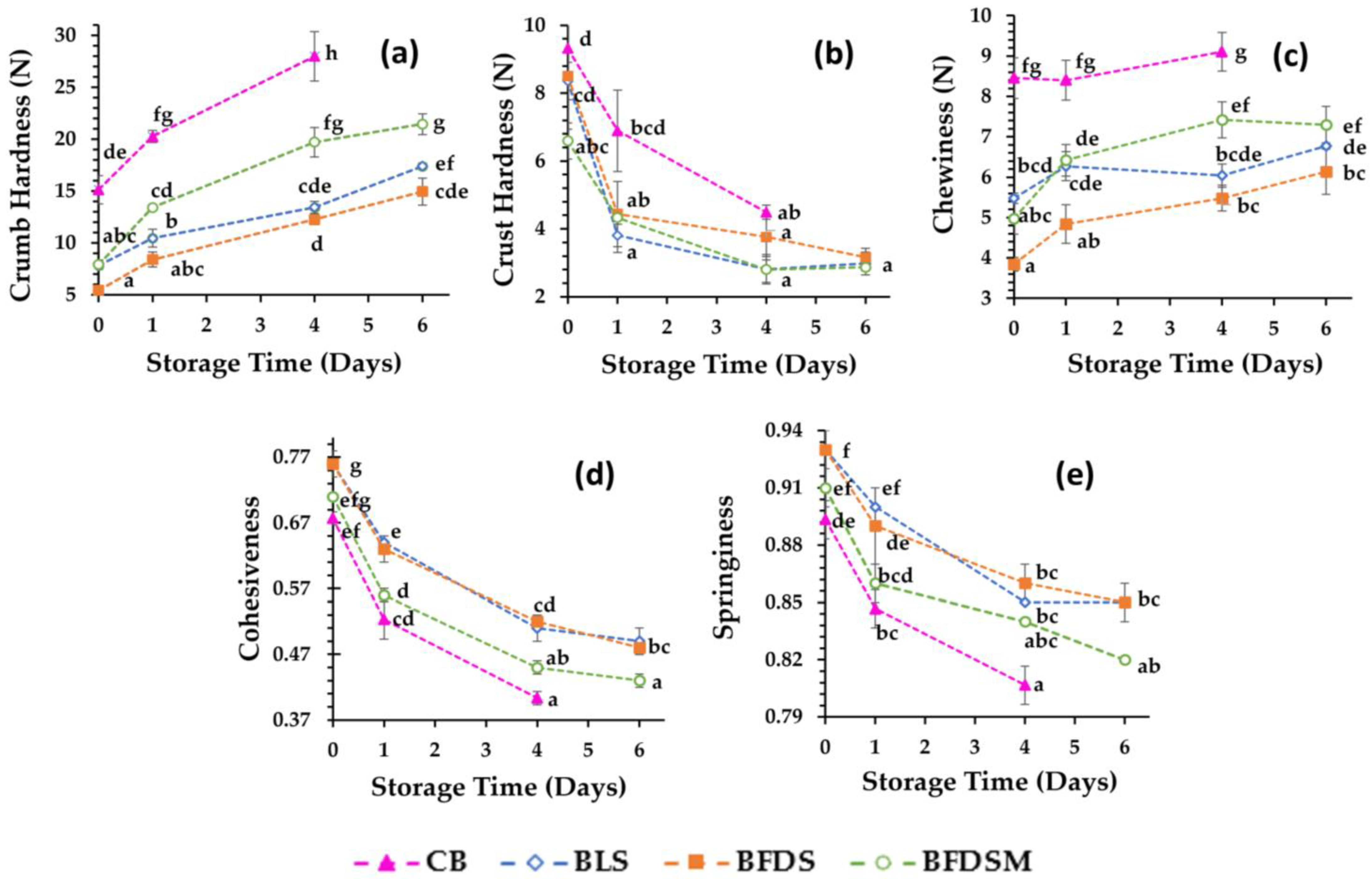
3.10. Texture Analysis of Breads
3.11. Starch Retrogradation in Bread Crumb
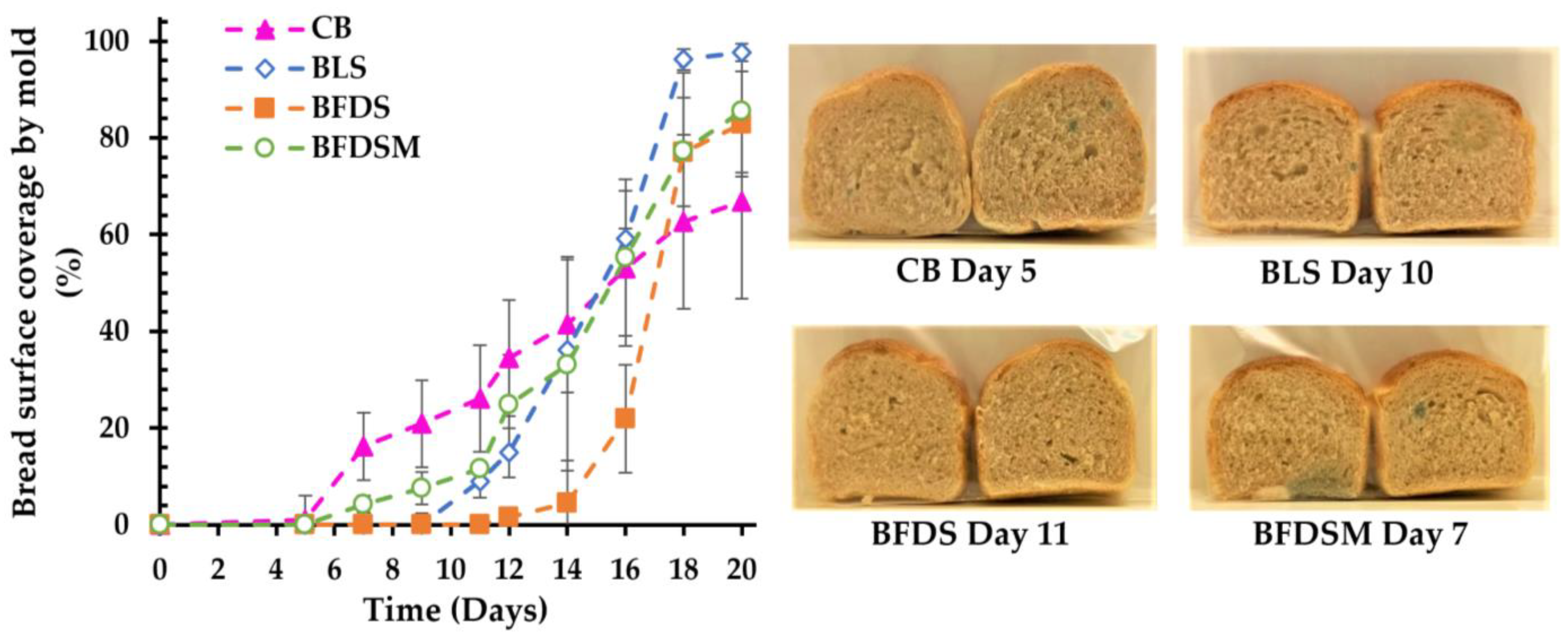
3.12. Mold Growth in Breads
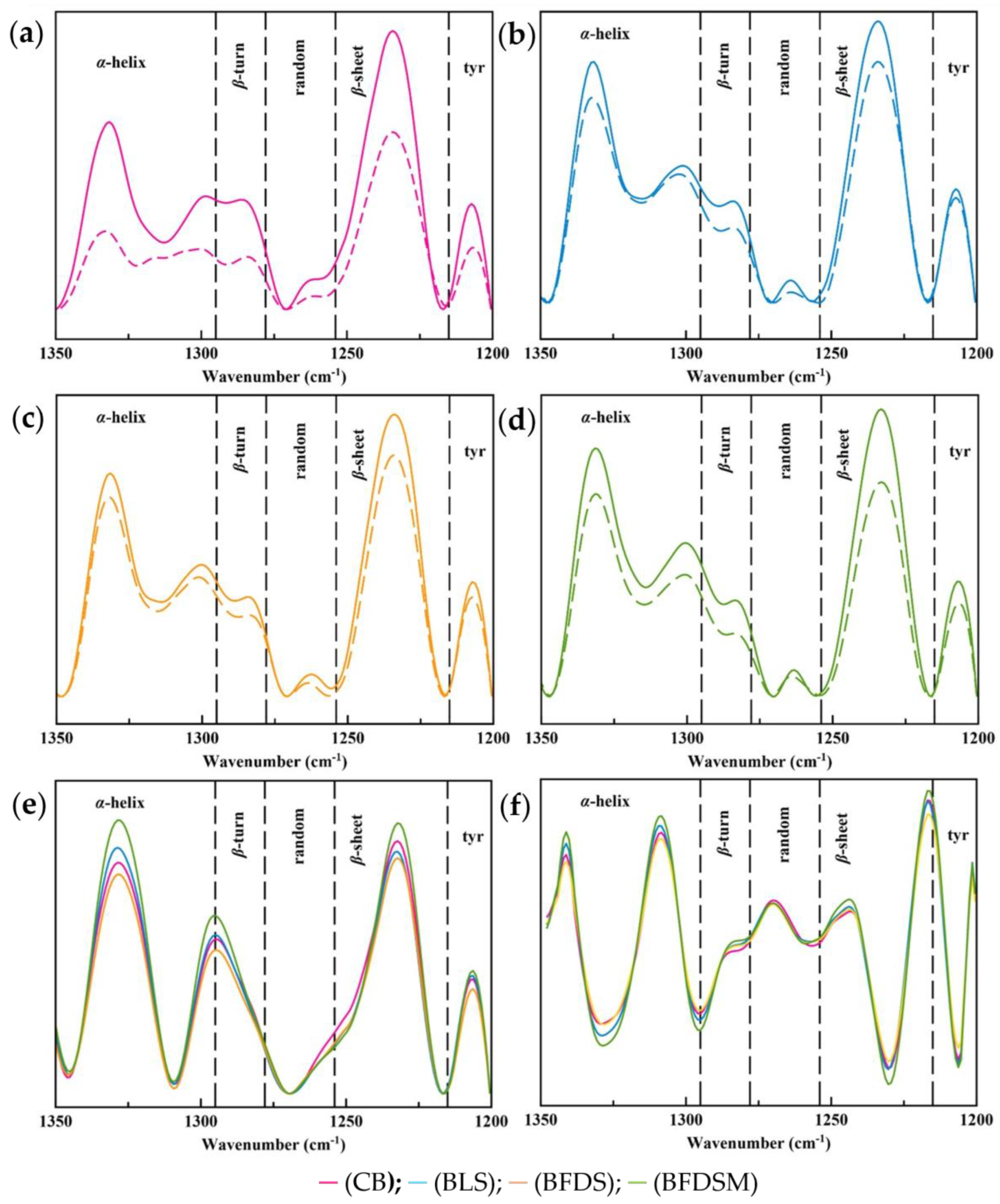
3.13. FTIR Spectroscopy Analysis in Sourdough and Breads
3.13.1. Protein Secondary Structure of Doughs
3.13.2. Protein Secondary Structure of Breads
3.13.3. Starch Chain Conformation of Breads
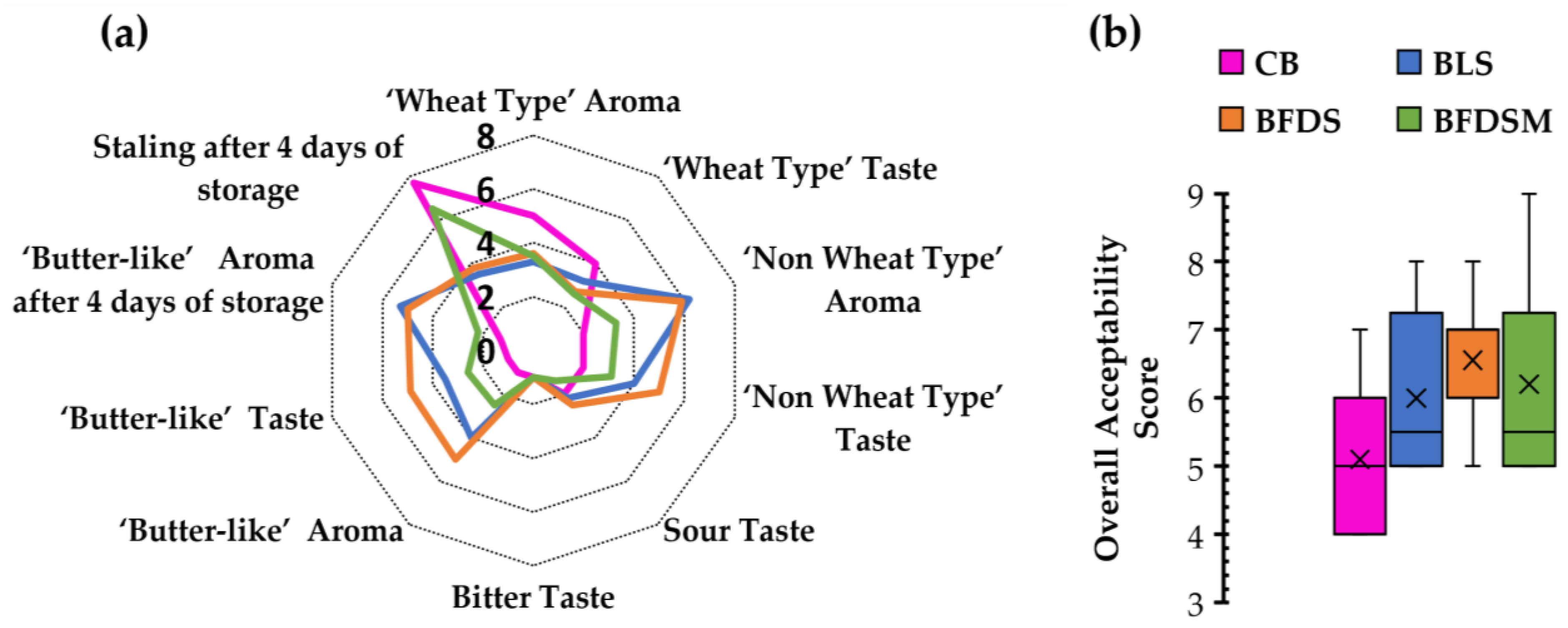
3.14. Sensory Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- De Vuyst, L.; Neysens, P. The Sourdough Microflora: Biodiversity and Metabolic Interactions. Trends Food Sci. Technol. 2005, 16, 43–56. [Google Scholar] [CrossRef]
- Corsetti, A.; Settanni, L. Lactobacilli in Sourdough Fermentation. Food Res. Int. 2007, 40, 539–558. [Google Scholar] [CrossRef]
- De Vuyst, L.; Vrancken, G.; Ravyts, F.; Rimaux, T.; Weckx, S. Biodiversity, Ecological Determinants, and Metabolic Exploitation of Sourdough Microbiota. Food Microbiol. 2009, 26, 666–675. [Google Scholar] [CrossRef] [PubMed]
- Tsafrakidou, P.; Michaelidou, A.-M.G.; Biliaderis, C. Fermented Cereal-Based Products: Nutritional Aspects, Possible Impact on Gut Microbiota and Health Implications. Foods 2020, 9, 734. [Google Scholar] [CrossRef]
- De Vos, M. Frontiers in Food Biotechnology–Fermentations and Functionality. Curr. Opin. Biotechnol. 2005, 2, 187–189. [Google Scholar] [CrossRef]
- Gobbetti, M.; Rizzello, C.G.; Di Cagno, R.; De Angelis, M. How the Sourdough May Affect the Functional Features of Leavened Baked Goods. Food Microbiol. 2014, 37, 30–40. [Google Scholar] [CrossRef]
- Ludewig, H.G. Methods of Prolonging Storage Life of Yeast-Raised Bakery Products. Brot Backwaren 1988, 36, 256–259. [Google Scholar]
- Ortola, C.; Barber, B.; Pelaez, M.T.; Benedito de Barber, C. Storage of packaged white bread, 1: Effects of water activity reduction and microbial contamination control on bread characteristics. Rev. Agroquim. Tecnol. Aliment. Spain 1989, 29, 384–398. [Google Scholar]
- Corsetti, A.; Gobbetti, M.; Balestrieri, F.; Paoletti, F.; Russi, L.; Rossi, J. Sourdough Lactic Acid Bacteria Effects on Bread Firmness and Stalin. J. Food Sci. 1998, 63, 347–351. [Google Scholar] [CrossRef]
- Collins, N.E.; Kirschner, L.A.M.; von Holy, A. Characterization of Bacillus Isolates from Ropey Bread, Bakery Equipment and Raw Materials. S. Afr. J. Sci. 1991, 87, 62–66. [Google Scholar]
- Manini, F.; Casiraghi, M.C.; Poutanen, K.; Brasca, M.; Erba, D.; Plumed-Ferrer, C. Characterization of Lactic Acid Bacteria Isolated from Wheat Bran Sourdough. LWT Food Sci. Technol. 2016, 66, 275–283. [Google Scholar] [CrossRef]
- Demirbaş, F.; İspirli, H.; Kurnaz, A.A.; Yilmaz, M.T.; Dertli, E. Antimicrobial and Functional Properties of Lactic Acid Bacteria Isolated from Sourdoughs. LWT Food Sci. Technol. 2017, 79, 361–366. [Google Scholar] [CrossRef]
- Liu, X.; Zhou, M.; Jiaxin, C.; Luo, Y.; Ye, F.; Jiao, S.; Hu, X.; Zhang, J.; Lü, X. Bacterial Diversity in Traditional Sourdough from Different Regions in China. LWT 2018, 96, 251–259. [Google Scholar] [CrossRef]
- Liu, A.; Jia, Y.; Zhao, L.; Gao, Y.; Liu, G.; Chen, Y.; Zhao, G.; Xu, L.; Shen, L.; Liu, Y.; et al. Diversity of Isolated Lactic Acid Bacteria in Ya’an Sourdoughs and Evaluation of Their Exopolysaccharide Production Characteristics. LWT 2018, 95, 17–22. [Google Scholar] [CrossRef]
- Boye, J.; Zare, F.; Pletch, A. Pulse Proteins: Processing, Characterization, Functional Properties and Applications in Food and Feed. Food Res. Int. 2010, 43, 414–431. [Google Scholar] [CrossRef]
- Moroni, A.V.; Dal Bello, F.; Arendt, E.K. Sourdough in Gluten-Free Bread-Making: An Ancient Technology to Solve a Novel Issue? Food Microbiol. 2009, 26, 676–684. [Google Scholar] [CrossRef]
- Hatzikamari, M.; Kyriakidis, D.A.; Tzanetakis, N.; Biliaderis, C.G.; Litopoulou-Tzanetaki, E. Biochemical Changes during a Submerged Chickpea Fermentation Used as a Leavening Agent for Bread Production. Eur. Food Res. Technol. 2006, 224, 715. [Google Scholar] [CrossRef]
- Boyaci Gunduz, C.P.; Gaglio, R.; Franciosi, E.; Settanni, L.; Erten, H. Molecular Analysis of the Dominant Lactic Acid Bacteria of Chickpea Liquid Starters and Doughs and Propagation of Chickpea Sourdoughs with Selected Weissella Confusa. Food Microbiol. 2020, 91, 103490. [Google Scholar] [CrossRef]
- Mygdalia, A.S.; Nouska, C.; Hatzikamari, M.; Biliaderis, C.G.; Lazaridou, A. A Sourdough Process Based on Fermented Chickpea Extract as Leavening and Anti-Staling Agent for Improving the Quality of Gluten-Free Breads. Food Res. Int. 2022, 159, 111593. [Google Scholar] [CrossRef]
- Gidari-Gounaridou, C.; Nouska, C.; Hatzikamari, M.; Kotsiou, K.; Biliaderis, C.G.; Lazaridou, A. Impact of Dry Sourdough Based on a Fermented Chickpea Starter on Quality Characteristics and Shelf Life of Gluten-Free Bread. Food Biosci. 2023, 53, 102780. [Google Scholar] [CrossRef]
- Hatzikamari, M.; Yiangou, M.; Tzanetakis, N.; Litopoulou-Tzanetaki, E. Changes in Numbers and Kinds of Bacteria during a Chickpea Submerged Fermentation Used as a Leavening Agent for Bread Production. Int. J. Food Microbiol. 2007, 116, 37–43. [Google Scholar] [CrossRef] [PubMed]
- Papatsaroucha, E.; Pavlidou, S.; Hatzikamari, M.; Lazaridou, A.; Torriani, S.; Gerasopoulos, D.; Tzanetaki, E.L. Preservation of Pears in Water in the Presence of Sinapis Arvensis Seeds: A Greek Tradition. Int. J. Food Microbiol. 2012, 159, 254–262. [Google Scholar] [CrossRef]
- Erol, I.; Goncuoglu, M.; Ayaz, N.D.; Ormanci, F.S.B.; Hildebrandt, G. Molecular Typing of Clostridium Perfringens Isolated from Turkey Meat by Multiplex PCR. Lett. Appl. Microbiol. 2008, 47, 31–34. [Google Scholar] [CrossRef]
- Ahsani, M.; Mohammadabadi, M.; Shamsaddini, M. Clostridium Perfringens Isolate Typing by Multiplex PCR. J. Venom. Anim. Toxins Trop. Dis. 2010, 16, 573–578. [Google Scholar] [CrossRef]
- Meer, R.R.; Songer, J.G. Multiplex Polymerase Chain Reaction Assay for Genotyping Clostridium Perfringens. Am. J. Vet. Res. 1997, 58, 702–705. [Google Scholar] [PubMed]
- Baums, C.G.; Schotte, U.; Amtsberg, G.; Goethe, R. Diagnostic Multiplex PCR for Toxin Genotyping of Clostridium Perfringens Isolates. Vet. Microbiol. 2004, 100, 11–16. [Google Scholar] [CrossRef] [PubMed]
- Brandt, M.J. Sourdough Products for Convenient Use in Baking. Food Microbiol. 2007, 24, 161–164. [Google Scholar] [CrossRef]
- Armero, E.; Collar, C. Texture Properties of Formulated Wheat Doughs. Z. Für Leb. Forsch. A 1997, 204, 136–145. [Google Scholar] [CrossRef]
- AACC International 44-15.02 (Moisture Content) Methods. In Approved Methods of the American Association of Cereal Chemists; American Association of Cereal Chemists International: St. Paul, MN, USA, 2010.
- Kotsiou, K.; Sacharidis, D.-D.; Matsakidou, A.; Biliaderis, C.G.; Lazaridou, A. Impact of Roasted Yellow Split Pea Flour on Dough Rheology and Quality of Fortified Wheat Breads. Foods 2021, 10, 1832. [Google Scholar] [CrossRef]
- Cai, S.; Singh, B.R. Identification of β-Turn and Random Coil Amide III Infrared Bands for Secondary Structure Estimation of Proteins. Biophys. Chem. 1999, 80, 7–20. [Google Scholar] [CrossRef]
- Muadiad, K.; Sirivongpaisal, P. Effect of Frozen Storage and Hydrocolloids on Structural and Rheological Properties of Frozen Roti Dough Supplemented with Rice Bran. Food Sci. Technol. 2021, 42, e44620. [Google Scholar] [CrossRef]
- Jung, Y.M.; Czarnik-Matusewicz, B.; Ozaki, Y. Two-Dimensional Infrared, Two-Dimensional Raman, and Two-Dimensional Infrared and Raman Heterospectral Correlation Studies of Secondary Structure of β-Lactoglobulin in Buffer Solutions. J. Phys. Chem. B 2000, 104, 7812–7817. [Google Scholar] [CrossRef]
- Lawless, H.T.; Heymann, H. Acceptance Testing. In Sensory Evaluation of Food: Principles and Practices; Lawless, H.T., Heymann, H., Eds.; Food Science Text Series; Springer: New York, NY, USA, 2010; pp. 325–347. ISBN 978-1-4419-6488-5. [Google Scholar]
- Lawless, H.T.; Heymann, H. Preference Testing. In Sensory Evaluation of Food: Principles and Practices; Lawless, H.T., Heymann, H., Eds.; Food Science Text Series; Springer: New York, NY, USA, 2010; pp. 303–324. ISBN 978-1-4419-6488-5. [Google Scholar]
- Kyyaly, R.; Lawand, M.; Khatib, F. Isolation Bacteria from Soaked Chickpea Using for Bakery Fermentation. Inter. J. Acad. Sci. Res. 2017, 5, 77–89. [Google Scholar]
- Katsaboxakis, K.; Mallidis, K. The Microflora of Soak Water during Natural Fermentation of Coarsely Ground Chickpea (Cicer arietinum) Seeds. Lett. Appl. Microbiol. 1996, 23, 261–265. [Google Scholar] [CrossRef]
- Hristoskova, S.P.; Yocheva, L.D.; Yankov, D.S.; Danova, S.T. Newly Characterized Butyrate Producing Clostridium Sp. Strain 4a1, Isolated from Chickpea Beans (Cicer arietinum L.). Bulg. Chem. Commun. 2018, 50, 459–466. [Google Scholar]
- Skjelkvåle, R.; Stringer, M.F.; Smart, J.L. Enterotoxin Production by Lecithinase-positive and Lecithinase-negative Clostridium Perfringens Isolated from Food Poisoning Outbreaks and Other Sources. J. Appl. Bacteriol. 1979, 47, 329–339. [Google Scholar] [CrossRef]
- Andersson, A.; Ronner, U.; Granum, P.E. What Problems Does the Food Industry Have with the Spore-Forming Pathogens Bacillus Cereus and Clostridium Perfringens? Int. J. Food Microbiol. 1995, 28, 145–155. [Google Scholar] [CrossRef]
- McClane, B.A.; Robertson, S.L.; Li, J. Clostridium perfringens. In Food Microbiology; Doyle, M.P., Buchanan, R.L., Eds.; ASM Press: Washington, DC, USA, 2013; pp. 465–489. ISBN 978-1-68367-058-2. [Google Scholar]
- Lattanzi, A.; Minervini, F.; Gobbetti, M. Assessment of Comparative Methods for Storing Type-I Wheat Sourdough. LWT Food Sci. Technol. 2014, 59, 948–955. [Google Scholar] [CrossRef]
- Reale, A.; Di Renzo, T.; Preziuso, M.; Panfili, G.; Cipriano, L.; Messia, M.C. Stabilization of Sourdough Starter by Spray Drying Technique: New Breadmaking Perspective. LWT 2019, 99, 468–475. [Google Scholar] [CrossRef]
- Carpenter, J.F.; Crowe, L.M.; Crowe, J.H. Stabilization of Phosphofructokinase with Sugars during Freeze-Drying: Characterization of Enhanced Protection in the Presence of Divalent Cations. Biochim. Biophys. Acta BBA Gen. Subj. 1987, 923, 109–115. [Google Scholar] [CrossRef]
- Carpenter, J.F.; Crowe, J.H. Modes of Stabilization of a Protein by Organic Solutes during Desiccation. Cryobiology 1988, 25, 459–470. [Google Scholar] [CrossRef]
- Carpenter, J.F.; Crowe, J.H. An Infrared Spectroscopic Study of the Interactions of Carbohydrates with Dried Proteins. Biochemistry 1989, 28, 3916–3922. [Google Scholar] [CrossRef]
- Taylor, L.S.; Zografi, G. Sugar–Polymer Hydrogen Bond Interactions in Lyophilized Amorphous Mixtures. J. Pharm. Sci. 1998, 87, 1615–1621. [Google Scholar] [CrossRef] [PubMed]
- Oldenhof, H.; Wolkers, W.F.; Fonseca, F.; Passot, S.; Marin, M. Effect of Sucrose and Maltodextrin on the Physical Properties and Survival of Air-Dried Lactobacillus Bulgaricus: An In Situ Fourier Transform Infrared Spectroscopy Study. Biotechnol. Prog. 2008, 21, 885–892. [Google Scholar] [CrossRef] [PubMed]
- Crowe, J.H.; Crowe, L.M.; Carpenter, J.F. Preserving Dry Biomaterials: The Water Replacement Hypothesis, Part 1. Biopharm 1993, 4, 28–32. [Google Scholar]
- Crowe, J.H.; Crowe, L.M.; Carpenter, J.F. Preserving Dry Biomaterials: The Water Replacement Hypothesis, Part 2. Biopharm 1993, 4, 40–43. [Google Scholar]
- Sharma, N.K.; Keerqin, C.; Wu, S.-B.; Choct, M.; Swick, R.A. Emissions of Volatile Odorous Metabolites by Clostridium Perfringens—In Vitro Study Using Two Broth Cultures. Poult. Sci. 2017, 96, 3291–3297. [Google Scholar] [CrossRef]
- Petrognani, C.; Boon, N.; Ganigué, R. Production of Isobutyric Acid from Methanol by Clostridium Luticellarii. Green Chem. 2020, 22, 8389–8402. [Google Scholar] [CrossRef]
- Lacumin, L.; Comi, G. A Survey of a Blown Pack Spoilage Produced by Clostridium Perfringens in Vacuum–Packaged Wurstel. Food Microbiol. 2021, 94, 103654. [Google Scholar] [CrossRef]
- Macfarlane, S.; Macfarlane, G. Proteolysis and amino acidfermentation. In Human Colonic Bacteria: Role in Nutrition, Physiology and Pathology; Gibson, G., Macfarlane, G., Eds.; CRC Press: Boca Raton, FL, USA, 1995. [Google Scholar]
- Aguilar Uscanga, M.G.; Délia, M.-L.; Strehaiano, P. Brettanomyces Bruxellensis: Effect of Oxygen on Growth and Acetic Acid Production. Appl. Microbiol. Biotechnol. 2003, 61, 157–162. [Google Scholar] [CrossRef]
- Walker, G.M.; Stewart, G.G. Saccharomyces Cerevisiae in the Production of Fermented Beverages. Beverages 2016, 2, 30. [Google Scholar] [CrossRef]
- Gül, H.; Hayıt, F.; Acar, C.; Kurt, N.; Dizlek, H. Effects of Chickpea-Based Leavening Extract on Physical, Textural and Sensory Properties of White Wheat Bread. Int. J. Food Eng. 2018, 14, 20170348. [Google Scholar] [CrossRef]
- Sayaslan, A.; Şahin, N. Effects of Fermented-Chickpea Liquor (Chickpea Yeast) on Whole-Grain Wheat Flour Bread Properties. Qual. Assur. Saf. Crops Foods 2018, 10, 183–192. [Google Scholar] [CrossRef]
- Witczak, M.; Korus, J.; Ziobro, R.; Juszczak, L. The Effects of Maltodextrins on Gluten-Free Dough and Quality of Bread. J. Food Eng. 2010, 96, 258–265. [Google Scholar] [CrossRef]
- Haros, M.; Rosell, C.; Benedito, C. Effect of Different Carbohydrases on Fresh Bread Texture and Bread Staling. Eur. Food Res. Technol. 2002, 215, 425–430. [Google Scholar] [CrossRef]
- Giannou, V.; Tzia, C. Cryoprotective Role of Exogenous Trehalose in Frozen Dough Products. Food Bioprocess Technol. 2008, 1, 276–284. [Google Scholar] [CrossRef]
- Sahi, S.S.; Little, K. Quality control. In Bakery Products: Science and Technology; Hui, Y., Corke, H., De Leyn, I., Nip, W., Cross, N., Eds.; Blackwell Publishing: Oxford, UK, 2006; pp. 319–336. [Google Scholar]
- Sze-Yin, S.; Lai-Hoong, C. Effects of Maltodextrin and Trehalose on the Physical Properties of Chinese Steamed Bread Made from Frozen Doughs. Int. Food Res. J. 2013, 20, 1529–1535. [Google Scholar]
- Gobbetti, M.; Corsetti, A.; Rossi, J. Interaction between Lactic Acid Bacteria and Yeasts in Sour-Dough Using a Rheofermentometer. World J. Microbiol. Biotechnol. 1995, 11, 625–630. [Google Scholar] [CrossRef]
- Corsetti, A.; Gobbetti, M.; De Marco, B.; Balestrieri, F.; Paoletti, F.; Russi, L.; Rossi, J. Combined Effect of Sourdough Lactic Acid Bacteria and Additives on Bread Firmness and Staling. J. Agric. Food Chem. 2000, 48, 3044–3051. [Google Scholar] [CrossRef]
- Dal Bello, F.; Clarke, C.I.; Ryan, L.A.M.; Ulmer, H.; Schober, T.J.; Ström, K.; Sjögren, J.; van Sinderen, D.; Schnürer, J.; Arendt, E.K. Improvement of the Quality and Shelf Life of Wheat Bread by Fermentation with the Antifungal Strain Lactobacillus Plantarum FST 1.7. J. Cereal Sci. 2007, 45, 309–318. [Google Scholar] [CrossRef]
- Wu, C.; Liu, R.; Huang, W.; Rayas-Duarte, P.; Wang, F.; Yao, Y. Effect of Sourdough Fermentation on the Quality of Chinese Northern-Style Steamed Breads. J. Cereal Sci. 2012, 56, 127–133. [Google Scholar] [CrossRef]
- Broyart, B.; Trystram, G.; Duquenoy, A. Predicting Colour Kinetics during Cracker Baking. J. Food Eng. 1998, 35, 351–368. [Google Scholar] [CrossRef]
- Ahrne, L.; Andersson, C.-G.; Floberg, P.; Rose’n, J.; Lingnert, H. Effect of Crust Temperature and Water Content on Acrylamide Formation during Baking of White Bread: Steam and Falling Temperature Baking. LWT Food Sci. Technol. 2007, 40, 1708–1715. [Google Scholar] [CrossRef]
- Nashat, S.; Abdullah, M.Z. Quality evaluation of bakery products. In Computer Vision Technology for Food Quality Evaluation; Academic Press: Amsterdam, The Netherlands, 2008; pp. 481–522. ISBN 978-0-12-373642-0. [Google Scholar]
- Rizzello, C.G.; Nionelli, L.; Coda, R.; Cagno, R.D.; Gobbetti, M. Use of Sourdough Fermented Wheat Germ for Enhancing the Nutritional, Texture and Sensory Characteristics of the White Bread. Eur. Food Res. Technol. 2010, 10, 645–654. [Google Scholar] [CrossRef]
- Purlis, E.; Salvadori, V.O. Bread Browning Kinetics during Baking. J. Food Eng. 2007, 80, 1107–1115. [Google Scholar] [CrossRef]
- Lara, E.; Cortés, P.; Briones, V.; Perez, M. Structural and Physical Modifications of Corn Biscuits during Baking Process. LWT Food Sci. Technol. 2011, 44, 622–630. [Google Scholar] [CrossRef]
- Ozulku, G.; Arıcı, M. Characterization of the Rheological and Technological Properties of the Frozen Sourdough Bread with Chickpea Flour Addition. J. Food Meas. Charact. 2017, 11, 1493–1500. [Google Scholar] [CrossRef]
- Ertop, M.H.; İbrahim Tuğkan, Ş. Optimization of The Amount of Chickpea Sourdough and Dry Yeast in Wheat Bread Formulation: Evaluation of Physicochemical, Sensory and Antioxidant Properties. Food Sci. Technol. Res. 2018, 24, 45–53. [Google Scholar] [CrossRef]
- Ronda, F.; Roos, Y.H. Staling of Fresh and Frozen Gluten-Free Bread. J. Cereal Sci. 2011, 53, 340–346. [Google Scholar] [CrossRef]
- Biliaderis, C.; Izydorczyk, M.; Rattan, O. Effect of Arabinoxylans on Bread-Making Quality of Wheat Flours. Food Chem. 1995, 53, 165–171. [Google Scholar] [CrossRef]
- Ziobro, R.; Korus, J.; Witczak, M.; Juszczak, L. Influence of Modified Starches on Properties of Gluten-Free Dough and Bread. Part II: Quality and Staling of Gluten-Free Bread. Food Hydrocoll. 2012, 29, 68–74. [Google Scholar] [CrossRef]
- Roman, L.; Reguilon, M.P.; Martinez, M.M.; Gomez, M. The Effects of Starch Cross-Linking, Stabilization and Pre-Gelatinization at Reducing Gluten-Free Bread Staling. LWT 2020, 132, 109908. [Google Scholar] [CrossRef]
- Schiraldi, A.; Fessas, D. Mechanism of staling: An overview. In Bread Staling; Chinachoti, P., Vodovotz, Y., Eds.; CRC Press: Boca Raton, FL, USA, 2001; pp. 1–17. ISBN 978-1-351-08724-7. [Google Scholar]
- Bosmans, G.M.; Lagrain, B.; Ooms, N.; Fierens, E.; Delcour, J.A. Biopolymer Interactions, Water Dynamics, and Bread Crumb Firming. J. Agric. Food Chem. 2013, 61, 4646–4654. [Google Scholar] [CrossRef]
- Vouris, D.G.; Lazaridou, A.; Mandala, I.G.; Biliaderis, C.G. Wheat Bread Quality Attributes Using Jet Milling Flour Fractions. LWT 2018, 92, 540–547. [Google Scholar] [CrossRef]
- Gómez, M.; Ronda, F.; Caballero, P.A.; Blanco, C.A.; Rosell, C.M. Functionality of Different Hydrocolloids on the Quality and Shelf-Life of Yellow Layer Cakes. Food Hydrocoll. 2007, 21, 167–173. [Google Scholar] [CrossRef]
- Rizzello, C.G.; Lorusso, A.; Montemurro, M.; Gobbetti, M. Use of Sourdough Made with Quinoa (Chenopodium Quinoa) Flour and Autochthonous Selected Lactic Acid Bacteria for Enhancing the Nutritional, Textural and Sensory Features of White Bread. Food Microbiol. 2016, 56, 1–13. [Google Scholar] [CrossRef]
- Ribotta, P.D.; Le Bail, A. Thermo-Physical Assessment of Bread during Staling. LWT Food Sci. Technol. 2007, 40, 879–884. [Google Scholar] [CrossRef]
- Biliaderis, C.G. Structures and phase transitions of starch polymers. In Polysaccharide Association Structures; Walter, R.H., Ed.; Marcel Dekker, Inc.: New York, NY, USA, 1998; pp. 57–168. [Google Scholar]
- Ribotta, P.D.; Cuffini, S.; Leon, A.E.; Anon, M.C. The Staling of Bread: An X-Ray Diffraction Study. Eur. Food Res. Technol. 2004, 218, 219–223. [Google Scholar] [CrossRef]
- Katina, K.; Heiniö, R.-L.; Autio, K.; Poutanen, K. Optimization of Sourdough Process for Improved Sensory Profile and Texture of Wheat Bread. LWT Food Sci. Technol. 2006, 39, 1189–1202. [Google Scholar] [CrossRef]
- Torrieri, E.; Pepe, O.; Ventorino, V.; Masi, P.; Cavella, S. Effect of Sourdough at Different Concentrations on Quality and Shelf Life of Bread. LWT Food Sci. Technol. 2014, 56, 508–516. [Google Scholar] [CrossRef]
- Zeleznak, K.J.; Hoseney, R.C. The Role of Water in the Retrogradation of Wheat Starch Gels and Bread Crumb. Cereal Chem. 1986, 63, 407–411. [Google Scholar]
- Biliaderis, C.G. Chapter 8—Structural Transitions and Related Physical Properties of Starch. In Starch, 3rd ed.; BeMiller, J., Whistler, R., Eds.; Food Science and Technology; Academic Press: San Diego, CA, USA, 2009; pp. 293–372. ISBN 978-0-12-746275-2. [Google Scholar]
- Pepe, O.; Blajotta, G.; Anastasio, M.; Moschetti, G.; Ercolini, D.; Villani, F. Technological and Molecular Diversity of Lactobacillus Plantarum Strains Isolated from Naturally Fermented Sourdoughs. Syst. Appl. Microbiol. 2004, 27, 443–453. [Google Scholar] [CrossRef]
- Kaditzky, S.; Seitter, M.; Hertel, C.; Vogel, R.F. Performance of Lactobacillus Sanfranciscensis TMW 1.392 and Its Levansucrase Deletion Mutant in Wheat Dough and Comparison of Their Impact on Bread Quality. Eur. Food Res. Technol. 2008, 227, 433–442. [Google Scholar] [CrossRef]
- Thiele, C.; Gänzle, M.G.; Vogel, R.F. Fluorescence Labeling of Wheat Proteins for Determination of Gluten Hydrolysis and Depolymerization during Dough Processing and Sourdough Fermentation. J. Agric. Food Chem. 2003, 51, 2745–2752. [Google Scholar] [CrossRef]
- Thiele, C.; Grassl, S.; Gänzle, M. Gluten Hydrolysis and Depolymerization during Sourdough Fermentation. J. Agric. Food Chem. 2004, 52, 1307–1314. [Google Scholar] [CrossRef] [PubMed]
- He, H.; Hoseney, R.C. Changes in Bread Firmness and Moisture during Long-Term Storage. Cereal Chem. 1990, 67, 603–605. [Google Scholar]
- Rinaldi, M. Sourdough Fermentation and Chestnut Flour in Gluten-Free Bread: A Shelf-Life Evaluation. Food Chem. 2017, 9, 144–152. [Google Scholar] [CrossRef]
- Hayta, M.; Ertop, M.H. Physicochemical, Textural and Microbiological Properties of Optimised Wheat Bread Formulations as Affected by Differently Fermented Sourdough. Qual. Assur. Saf. Crops Foods 2019, 11, 283–293. [Google Scholar] [CrossRef]
- Legan, J.D. Mould Spoilage of Bread: The Problem and Some Solutions. Int. Biodeterior. Biodegrad. 1993, 32, 33–53. [Google Scholar] [CrossRef]
- Vytřasová, J.; Přibáňová, P.; Marvanová, L. Occurrence of Xerophilic Fungi in Bakery Gingerbread Production. Int. J. Food Microbiol. 2002, 72, 91–96. [Google Scholar] [CrossRef]
- Debonne, E.; Van Schoors, F.; Maene, P.; Van Bockstaele, F.; Vermeir, P.; Verwaeren, J.; Eeckhout, M.; Devlieghere, F. Comparison of the Antifungal Effect of Undissociated Lactic and Acetic Acid in Sourdough Bread and in Chemically Acidified Wheat Bread. Int. J. Food Microbiol. 2020, 321, 108551. [Google Scholar] [CrossRef]
- Najafi, M.A.; Rezaei, K.; Safari, M.; Razavi, S.H. Effects of Several Starter Cultures on the Anti-Mold Activity and Sensory Attributes of a Traditional Flat Bread (Sangak) from Iran. Food Sci. Biotechnol. 2012, 21, 113–121. [Google Scholar] [CrossRef]
- Dalié, D.K.D.; Deschamps, A.M.; Richard-Forget, F. Lactic Acid Bacteria—Potential for Control of Mould Growth and Mycotoxins: A Review. Food Control 2010, 21, 370–380. [Google Scholar] [CrossRef]
- Le Lay, C.; Mounier, J.; Vasseur, V.; Weill, A.; Le Blay, G.; Barbier, G.; Coton, E. In Vitro and in Situ Screening of Lactic Acid Bacteria and Propionibacteria Antifungal Activities against Bakery Product Spoilage Molds. Food Control 2016, 60, 247–255. [Google Scholar] [CrossRef]
- Axel, C.; Zannini, E.; Arendt, E.K. Mold Spoilage of Bread and Its Biopreservation: A Review of Current Strategies for Bread Shelf Life Extension. Crit. Rev. Food Sci. Nutr. 2017, 57, 3528–3542. [Google Scholar] [CrossRef]
- Axel, C.; Brosnan, B.; Zannini, E.; Peyer, L.C.; Furey, A.; Coffey, A.; Arendt, E.K. Antifungal Activities of Three Different Lactobacillus Species and Their Production of Antifungal Carboxylic Acids in Wheat Sourdough. Appl. Microbiol. Biotechnol. 2016, 100, 1701–1711. [Google Scholar] [CrossRef]
- Kaddour, A.A.; Mondet, M.; Cuq, B. Description of Chemical Changes Implied During Bread Dough Mixing by FT-ATR Mid-Infrared Spectroscopy. Cereal Chem. 2008, 85, 673–678. [Google Scholar] [CrossRef]
- Kłosok, K.; Welc, R.; Fornal, E.; Nawrocka, A. Effects of Physical and Chemical Factors on the Structure of Gluten, Gliadins and Glutenins as Studied with Spectroscopic Methods. Molecules 2021, 26, 508. [Google Scholar] [CrossRef] [PubMed]
- Bock, J.E.; Connelly, R.K.; Damodaran, S. Impact of Bran Addition on Water Properties and Gluten Secondary Structure in Wheat Flour Doughs Studied by Attenuated Total Reflectance Fourier Transform Infrared Spectroscopy. Cereal Chem. 2013, 90, 377–386. [Google Scholar] [CrossRef]
- Bock, J.E. The Structural Evolution of Water and Gluten in Refined and Whole Grain Breads: A Study of Soft and Hard Wheat Breads from Postmixing to Final Product. Cereal Chem. 2019, 96, 520–531. [Google Scholar] [CrossRef]
- Zhang, Y.; Hong, T.; Yu, W.; Yang, N.; Jin, Z.; Xu, X. Structural, Thermal and Rheological Properties of Gluten Dough: Comparative Changes by Dextran, Weak Acidification and Their Combination. Food Chem. 2020, 330, 127154. [Google Scholar] [CrossRef] [PubMed]
- Nawrocka, A.; Miś, A.; Niewiadomski, Z. Dehydration of Gluten Matrix as a Result of Dietary Fibre Addition—A Study on Model Flour with Application of FT-IR Spectroscopy. J. Cereal Sci. 2017, 74, 86–94. [Google Scholar] [CrossRef]
- Nutter, J.; Saiz, A.I.; Iurlina, M.O. Microstructural and Conformational Changes of Gluten Proteins in Wheat-Rye Sourdough. J. Cereal Sci. 2019, 87, 91–97. [Google Scholar] [CrossRef]
- Feng, J.; Zhang, S.; Zhang, Y.; Wang, J. Correlation of Gluten Molecular Conformation with Dough Viscoelastic Properties during Storage. Grain Oil Sci. Technol. 2018, 1, 1–7. [Google Scholar] [CrossRef]
- Omedi, J.O.; Huang, J.; Huang, W.; Zheng, J.; Zeng, Y.; Zhang, B.; Zhou, L.; Zhao, F.; Li, N.; Gao, T. Suitability of Pitaya Fruit Fermented by Sourdough LAB Strains for Bread Making: Its Impact on Dough Physicochemical, Rheo-Fermentation Properties and Antioxidant, Antifungal and Quality Performance of Bread. Heliyon 2021, 7, e08290. [Google Scholar] [CrossRef] [PubMed]
- Kotsiou, K.; Sacharidis, D.-D.; Matsakidou, A.; Biliaderis, C.G.; Lazaridou, A. Physicochemical and Functional Aspects of Composite Wheat-Roasted Chickpea Flours in Relation to Dough Rheology, Bread Quality and Staling Phenomena. Food Hydrocoll. 2022, 124, 107322. [Google Scholar] [CrossRef]
- Li, W.; Zhang, S.; Feng, Y.; Wang, Z.; Han, H. Effects of Storage Time on Steamed Bread Quality and Gluten Protein Structure. IOP Conf. Ser. Mater. Sci. Eng. 2018, 392, 052006. [Google Scholar] [CrossRef]
- Sivam, A.S.; Sun-Waterhouse, D.; Perera, C.O.; Waterhouse, G.I.N. Application of FT-IR and Raman Spectroscopy for the Study of Biopolymers in Breads Fortified with Fibre and Polyphenols. Food Res. Int. 2013, 50, 574–585. [Google Scholar] [CrossRef]
- Nawrocka, A.; Krekora, M.; Niewiadomski, Z.; Miś, A. FTIR Studies of Gluten Matrix Dehydration after Fibre Polysaccharide Addition. Food Chem. 2018, 252, 198–206. [Google Scholar] [CrossRef]
- Mouzakitis, C.-K.; Sereti, V.; Matsakidou, A.; Kotsiou, K.; Biliaderis, C.G.; Lazaridou, A. Physicochemical Properties of Zein-Based Edible Films and Coatings for Extending Wheat Bread Shelf Life. Food Hydrocoll. 2022, 132, 107856. [Google Scholar] [CrossRef]
- Van Soest, J.J.G.; Benes, K.; De Wit, D. The Influence of Acid Hydrolysis of Potato Starch on the Stress-Strain Propoerties of Thermoplastic Starch. Starch Stärke 1995, 47, 429–434. [Google Scholar] [CrossRef]
- Smits, A.L.M.; Ruhnau, F.C.; Vliegenthart, J.F.G.; van Soest, J.J.G. Ageing of Starch Based Systems as Observed with FT-IR and Solid State NMR Spectroscopy. Starch Stärke 1998, 50, 478–483. [Google Scholar] [CrossRef]
- Warren, F.J.; Gidley, M.J.; Flanagan, B.M. Infrared Spectroscopy as a Tool to Characterise Starch Ordered Structure—A Joint FTIR–ATR, NMR, XRD and DSC Study. Carbohydr. Polym. 2016, 139, 35–42. [Google Scholar] [CrossRef] [PubMed]
- Nhouchi, Z.; Karoui, R. Application of Fourier-Transform Mid Infrared Spectroscopy for the Monitoring of Pound Cakes Quality during Storage. Food Chem. 2018, 252, 327–334. [Google Scholar] [CrossRef]
- Ringsted, T.; Siesler, H.W.; Engelsen, S.B. Monitoring the Staling of Wheat Bread Using 2D MIR-NIR Correlation Spectroscopy. J. Cereal Sci. 2017, 75, 92–99. [Google Scholar] [CrossRef]
- Xiao, Y.; Huang, L.; Chen, Y.; Zhang, S.; Rui, X.; Dong, M. Comparative Study of the Effects of Fermented and Non-Fermented Chickpea Flour Addition on Quality and Antioxidant Properties of Wheat Bread. CyTA J. Food 2016, 14, 621–631. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Toxin | Sequence 5′-3′ | Amplicon (bp) |
|---|---|---|
| α | GCTAATGTTACTGCCGTTGA | 324 |
| CCTCTGATACATCGTGTAAG | ||
| β | GCGAATATGCTGAATCATCTA | 196 |
| GCAGGAACATTAGTATATCTTC | ||
| ε | GCGGTGATATCCATCTATTC | 655 |
| CCACTTACTTGTCCTACTAAC | ||
| CPE | GGAGATGGTTGGATATTAGG | 233 |
| GGACCAGCAGTTGTAGATA | ||
| CPE | GGGGAACCCTCAGTAGTTTCA | 506 |
| ACCAGCTGGATTTGAGTTTAATG |
| Component (g) | CB | ΒLS | BFDS | BFDSM |
|---|---|---|---|---|
| Wheat flour | 92.50 | 75.00 | 75.00 | 75.00 |
| Chickpea flour | 7.50 | - | - | - |
| Water | 59.00 | 13.50 | 59.00 | 59.00 |
| Baker’s yeast | 1.00 | 0.20 | 0.20 | 0.20 |
| Liquid sourdough (g H2O + dry solids) | - | (45.50 + 25.00) | - | - |
| Dried FDS 1 | - | - | 25.00 | - |
| Dried FDSM 1 | - | - | - | 25.00 |
| Salt | 1.00 | 1.00 | 1.00 | 1.00 |
| Vinegar | 1.50 | 1.50 | 1.50 | 1.50 |
| Organic Acids (mg/g Sourdough) | ||||
|---|---|---|---|---|
| Lactic Acid | Acetic Acid | Isobutyric Acid | Butyric Acid | |
| LS 1 | 4.18 ± 0.27 a 2 | 39.05 ± 0.06 a | 6.54 ± 0.71 a | 4.69 ± 0.59 a |
| FDS | 4.24 ± 0.06 a | 32.83 ± 0.21 a | 6.90 ± 0.32 a | 13.58 ± 0.29 b |
| FDSM | 4.41 ± 0.22 a | 55.64 ± 6.07 b | 10.55 ± 1.27 b | 31.39 ± 3.41 c |
| CB 1 | BLS | BFDS | BFDSM | |
|---|---|---|---|---|
| Loaf-specific volume (mL/g) | 2.20 ± 0.04 a 2 | 2.64 ± 0.05 b | 3.19 ± 0.08 c | 2.69 ± 0.11 b |
| Crust color parameters | ||||
| L* | 61.07 ± 1.22 c | 50.82 ± 0.81 a | 56.35 ± 1.33 b | 50.62 ± 1.26 a |
| a* | 8.91 ± 0.28 a | 12.27 ± 0.05 b | 14.45 ± 0.01 d | 13.58 ± 0.44 c |
| b* | 33.30 ± 1.23 b | 28.22 ± 0.28 ab | 27.43 ± 0.96 a | 29.50 ± 0.35 ab |
| Bread Moisture (%) | |||||
|---|---|---|---|---|---|
| 0 Day | 1 Day | 4 Day | 6 Day | ||
| Crust | CB 1 | 17.84 ± 0.49 a 2 | 27.23 ± 0.11 b | 30.38 ± 0.69 c | |
| BLS | 20.60 ± 0.26 a | 27.34 ± 0.21 b | 31.25 ± 0.60 c | 32.69 ± 0.84 c | |
| BFDS | 16.19 ± 1.55 a | 22.31± 0.14 b | 27.85 ± 0.29 c | 26.64 ± 0.70 c | |
| BFDSM | 20.32 ± 0.45 a | 21.86 ± 0.38 a | 31.56 ± 0.60 b | 30.78 ± 1.23 b | |
| Crumb | CB | 45.02 ± 0.59 a | 44.04 ± 0.38 ab | 42.78 ± 0.64 b | |
| BLS | 46.08 ± 0.37 a | 45.96 ± 0.54 ab | 44.83 ± 0.62 b | 43.33 ± 1.81 b | |
| BFDS | 42.84 ± 0.18 a | 42.67 ± 0.57 b | 40.34 ± 0.11 b | 41.87 ± 0.06 b | |
| BFDSM | 42.84 ± 0.94 a | 42.68 ± 0.03 ab | 41.10 ± 1.02 b | 38.77 ± 0.98 c | |
| Melting Temperature of Retrogradation Endotherm | Retrogradation Melting Enthalpy | |||
|---|---|---|---|---|
| Samples 1 | Onset, To (°C) | Peak, Tp (°C) | Endset, Te (°C) | ΔHret, (mJ/mg Flour d.b.) |
| CB 2 h | 43.35 b 2 | 48.02 a | 60.18 b | 0.13 a |
| CB 96 h | 43.06 b | 52.44 de | 62.14 d | 2.43 de |
| BLS 2 h | 43.03 b | 47.92 a | 59.10 a | 0.13 a |
| BLS 96 h | 44.77 de | 53.15 e | 59.24 a | 1.09 b |
| BLS 144 h | 45.57 e | 52.95 de | 60.10 b | 1.47 bc |
| BFDS 2 h | 45.25 e | 50.42 bc | 60.55 b | 0.12 a |
| BFDS 96 h | 44.28 cd | 52.22 de | 60.35 b | 1.55 bc |
| BFDS 144 h | 43.81 bc | 52.64 de | 60.92 c | 1.88 cd |
| BFDSM 2 h | 43.07 b | 50.11 b | 60.35 b | 0.29 a |
| BFDSM 96 h | 41.70 a | 51.87 cd | 62.80 e | 1.89 cd |
| BFDSM 144 h | 43.25 b | 53.14 e | 63.415 f | 2.53 e |
| CB 1 (% Area) | BLS (% Area) | BFDS (% Area) | BFDSM (% Area) | |||||
|---|---|---|---|---|---|---|---|---|
| Secondary Structure Assignment | Dough | Leavened Dough | Dough | Leavened Dough | Dough | Leavened Dough | Dough | Leavened Dough |
| Tyr ring | 6.47 ± 0.06 a 2 | 7.13 ± 0.41 a | 6.36 ± 0.35 a | 6.98 ± 0.36 a | 6.72 ± 0.21 a | 6.89 ± 0.41 a | 6.57 ± 2.16 a | 6.89 ± 0.46 a |
| β-sheet | 39.85 ± 0.09 f | 36.96 ± 1.21 de | 39.50 ± 2.01 f | 35.79 ± 1.02 c | 37.72 ± 3.16 e | 36.22 ± 0.01 d | 34.54 ± 1.23 b | 32.79 ± 0.12 a |
| random | 3.24 ± 0.26 e | 0.25 ± 0.81 d | 1.74 ± 1.12 cd | 0.44 ± 0.00 a | 1.16 ± 0.01 c | 0.63 ± 0.35 ab | 1.02 ± 0.18 c | 1.21 ± 0.01 c |
| β-turn | 9.11 ± 0.56 d | 6.71 ± 0.24 a | 7.68 ± 1.51 ab | 8.54 ± 0.97 c | 8.46 ± 0.11 c | 8.66 ± 1.80 c | 8.43 ± 0.97 c | 7.23 ± 1.10 ab |
| β-turn/α-helix intermediate protein conformation | 14.95 ± 1.00 b | 21.28 ± 3.83 d | 16.22 ± 0.46 c | 15.66 ± 0.29 bc | 14.76 ± 3.72 b | 15.14 ± 0.56 bc | 13.57 ± 4.09 a | 13.52 ± 3.72 a |
| α-helix | 26.36 ± 0.67 a | 27.66 ± 3.94 b | 28.50 ± 5.14 b | 32.61 ± 1.32 c | 31.19 ± 2.16 c | 32.47 ± 2.60 c | 35.86 ± 3.71 d | 38.36 ± 2.19 e |
| Secondary Structure Assignment | Storage Time | CB 1 | BLS | BFDS | BFDSM |
|---|---|---|---|---|---|
| Days | %Area | %Area | %Area | %Area | |
| Tyr ring | 0 | 6.92 ± 0.12 aB 2 | 7.29 ± 0.08 bC | 6.64 ± 0.09 aA | 6.91 ± 0.03 bB |
| 4 | 6.92 ± 0.16 aA | 6.90 ± 0.08 aA | 6.87 ± 0.09 bA | 6.79 ± 0.07 bA | |
| 6 | - | 7.03 ± 0.09 aC | 6.79 ± 0.06 abB | 6.16 ± 0.03 aA | |
| β-sheet | 0 | 33.97 ± 0.09 aB | 33.54 ± 0.14 aB | 36.35 ± 0.34 aC | 32.56 ± 0.40 aA |
| 4 | 34.88 ± 0.54 aB | 42.10 ± 0.16 cD | 36.88 ± 0.70 aC | 33.14 ± 0.22 aA | |
| 6 | - | 36.65 ± 0.05 bB | 36.16 ± 0.15 aA | 45.50 ± 0.2 0 bC | |
| random | 0 | 4.15 ± 0.09 aC | 1.86 ± 0.08 bA | 1.51 ± 0.12 aA | 2.22 ± 0.11 bB |
| 4 | 4.19 ± 0.41 aC | 0.63 ± 0.02 aA | 2.03 ± 0.03 aB | 1.45 ± 0.10 aB | |
| 6 | - | 0.46 ± 0.01 aA | 1.71 ± 0.59 aB | 1.35 ± 0.22 aAB | |
| β-turn | 0 | 5.15 ± 0.03 aA | 5.13 ± 0.04 aA | 5.68 ± 0.08 aB | 5.13 ± 0.0 4 bA |
| 4 | 4.26 ± 0.40 aB | 5.69 ± 0.14 bC | 5.20 ± 0.05 aC | 3.22 ± 0.15 aA | |
| 6 | - | 7.35 ± 0.03 cB | 4.96 ± 0.66 aA | 5.59 ± 0.03 cA | |
| β-turn/α-helix intermediate protein conformation | 0 | 16.68 ± 0.13 aAB | 17.08 ± 0.10 cBC | 16.27 ± 0.27 aA | 17.43 ± 0.15 bC |
| 4 | 18.53 ± 0.25 bC | 14.09 ± 0.18 aA | 16.99 ± 0.13 aB | 21.03 ± 0.18 cD | |
| 6 | - | 15.36 ± 0.13 bB | 17.77 ± 1.36 aC | 12.87 ± 0.10 aA | |
| α-helix | 0 | 33.14 ± 0.18 bA | 35.10 ± 0.20 cB | 33.53 ± 0.35 aA | 35.75 ± 0.28 cB |
| 4 | 31.21 ± 0.6 4aAB | 30.59 ± 0.10 aA | 32.02 ± 0.46 aB | 34.36 ± 0.29 bC | |
| 6 | - | 33.15 ± 0.04 bB | 32.58 ± 1.21 aB | 28.52 ± 0.46 aA | |
| Ratio of Ordered to Amorphous Starch, R1047/1022 | |||||
| CB | BLS | BFDS | BFDSM | ||
| R1047/1022 | 0 | 0.694 ± 0.001 aAB | 0.682 ± 0.001 aA | 0.689 ± 0.002 aA | 0.695 ± 0.000 aB |
| 4 or 6 3 | 0.726 ± 0.002 bB | 0.718 ± 0.003 bA | 0.717 ± 0.004 bA | 0.747 ± 0.001 bC | |
| Rate of R1047/1022 increase (day−1) | +0.008 C | +0.006 B | +0.004 A | +0.009 D | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nouska, C.; Hatzikamari, M.; Matsakidou, A.; Biliaderis, C.G.; Lazaridou, A. Enhancement of Textural and Sensory Characteristics of Wheat Bread Using a Chickpea Sourdough Fermented with a Selected Autochthonous Microorganism. Foods 2023, 12, 3112. https://doi.org/10.3390/foods12163112
Nouska C, Hatzikamari M, Matsakidou A, Biliaderis CG, Lazaridou A. Enhancement of Textural and Sensory Characteristics of Wheat Bread Using a Chickpea Sourdough Fermented with a Selected Autochthonous Microorganism. Foods. 2023; 12(16):3112. https://doi.org/10.3390/foods12163112
Chicago/Turabian StyleNouska, Chrysanthi, Magdalini Hatzikamari, Anthia Matsakidou, Costas G. Biliaderis, and Athina Lazaridou. 2023. "Enhancement of Textural and Sensory Characteristics of Wheat Bread Using a Chickpea Sourdough Fermented with a Selected Autochthonous Microorganism" Foods 12, no. 16: 3112. https://doi.org/10.3390/foods12163112
APA StyleNouska, C., Hatzikamari, M., Matsakidou, A., Biliaderis, C. G., & Lazaridou, A. (2023). Enhancement of Textural and Sensory Characteristics of Wheat Bread Using a Chickpea Sourdough Fermented with a Selected Autochthonous Microorganism. Foods, 12(16), 3112. https://doi.org/10.3390/foods12163112