
The Impacts of Acidophilic Lactic Acid Bacteria on Food and Human Health: A Review of the Current Knowledge
 ,
,  ,
,  ,
,  and
and 
Abstract
1. Introduction
2. Overview of Acidophilic Microorganisms
3. Acidophilic Lactic Acid Bacteria in Foods
3.1. Their Utilization in Fermentation Technology as Starter Cultures
3.2. Food Safety and Stability Issues
3.3. Effects of Acidophilic Lactic Acid Bacteria on the Nutritional Value of Foods
4. Acidophilic Lactic Acid Bacteria’s Role in the Human Diet and Health
4.1. Their Probiotic Potentials
4.2. Beneficial and Toxic Compounds Released by Acidophilic Lactic Acid Bacteria
4.2.1. Organic Acids
4.2.2. B Group Vitamins
4.2.3. Gamma-Aminobutyric Acid (GABA)
4.2.4. Enzymes
4.2.5. Bacteriocins
4.2.6. Biogenic Amines
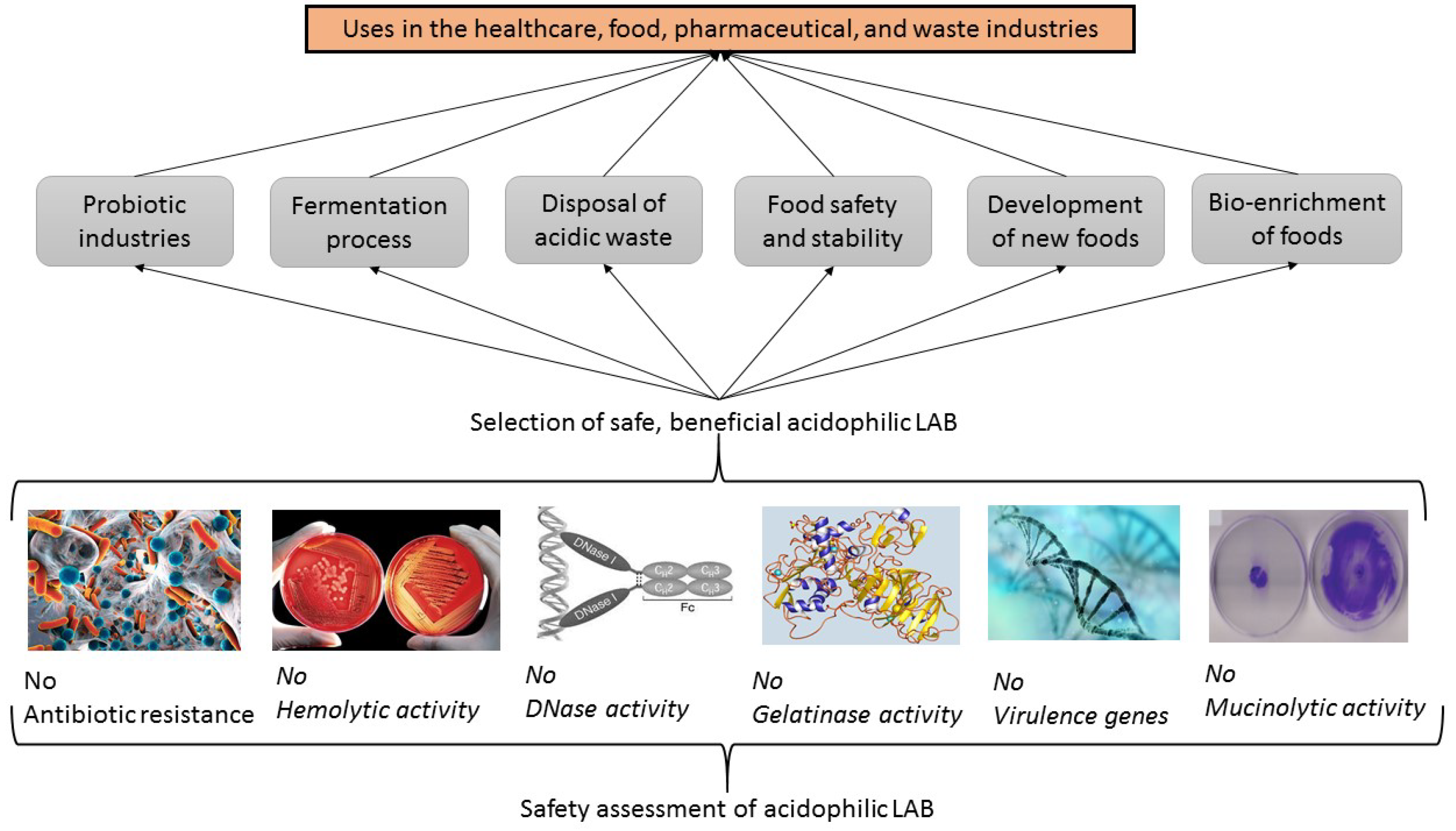
4.3. Safety Assessments of Acidophilic LAB with Probiotic Potentials
4.3.1. Antibiotic Resistance
4.3.2. Hemolytic Activity
4.3.3. DNase Activity
4.3.4. Gelatinase Activity
4.3.5. Presence of Virulence Genes
4.3.6. Mucinolytic Activity
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ayivi, R.D.; Gyawali, R.; Krastanov, A.; Aljaloud, S.O.; Worku, M.; Tahergorabi, R.; Silva, R.C.D.; Ibrahim, S.A. Lactic acid bacteria: Food safety and human health applications. Dairy 2020, 1, 202–232. [Google Scholar] [CrossRef]
- Johnson, D.B.; Quatrini, R. Acidophile microbiology in space and time. Curr. Issues Mol. Biol. 2020, 39, 63–76. [Google Scholar] [CrossRef]
- Bintsis, T. Lactic acid bacteria as starter cultures: An update in their metabolism and genetics. AIMS Microbiol. 2018, 4, 665. [Google Scholar] [CrossRef] [PubMed]
- Bajaj, B.K.; Claes, I.J.; Lebeer, S. Functional mechanisms of probiotics. J. Microbiol. Biotechnol. Food Sci. 2021, 2021, 321–327. [Google Scholar]
- Plessas, S.; Nouska, C.; Karapetsas, A.; Kazakos, S.; Alexopoulos, A.; Mantzourani, I.; Chondrou, P.; Fournomiti, M.; Galanis, A.; Bezirtzoglou, E. Isolation, characterization and evaluation of the probiotic potential of a novel Lactobacillus strain isolated from Feta-type cheese. Food Chem. 2017, 226, 102–108. [Google Scholar] [CrossRef] [PubMed]
- Bin Masalam, M.S.; Bahieldin, A.; Alharbi, M.G.; Al-Masaudi, S.; Al-Jaouni, S.K.; Harakeh, S.M.; Al-Hindi, R.R. Isolation, molecular characterization and probiotic potential of lactic acid bacteria in Saudi raw and fermented milk. Evid.-Based Complement. Altern. Med. 2018, 2018, 7970463. [Google Scholar] [CrossRef] [PubMed]
- Obafemi, Y.D.; Oranusi, S.U.; Ajanaku, K.O.; Akinduti, P.A.; Leech, J.; Cotter, P.D. African fermented foods: Overview, emerging benefits, and novel approaches to microbiome profiling. NPJ Sci. Food 2022, 6, 15. [Google Scholar] [CrossRef]
- Pérez-Armendáriz, B.; Cardoso-Ugarte, G.A. Traditional fermented beverages in Mexico: Biotechnological, nutritional, and functional approaches. Food Res. Int. 2020, 136, 109307. [Google Scholar] [CrossRef]
- Lorenzo, J.M.; Munekata, P.E.; Dominguez, R.; Pateiro, M.; Saraiva, J.A.; Franco, D. Main groups of microorganisms of relevance for food safety and stability: General aspects and overall description. In Innovative Technologies for Food Preservation; Academic Press: Cambridge, MA, USA, 2018; pp. 53–107. [Google Scholar]
- Gunyakti, A.; Asan-Ozusaglam, M. Lactobacillus gasseri from human milk with probiotic potential and some technological properties. LWT 2019, 109, 261–269. [Google Scholar] [CrossRef]
- Margalho, L.P.; Jorge, G.P.; Noleto, D.A.; Silva, C.E.; Abreu, J.S.; Piran, M.V.; Brocchi, M.; Sant’Ana, A.S. Biopreservation and probiotic potential of a large set of lactic acid bacteria isolated from Brazilian artisanal cheeses: From screening to in product approach. Microbiol. Res. 2021, 242, 126622. [Google Scholar] [CrossRef]
- Padmavathi, T.P.; Bhargavi, R.; Priyanka, P.R.; Niranjan, N.R.; Pavitra, P.V. Screening of potential probiotic lactic acid bacteria and production of amylase and its partial purification. J. Genet. Eng. Biotechnol. 2018, 16, 357–362. [Google Scholar] [CrossRef]
- Barbieri, F.; Montanari, C.; Gardini, F.; Tabanelli, G. Biogenic amine production by lactic acid bacteria: A review. Foods 2019, 8, 17. [Google Scholar] [CrossRef] [PubMed]
- Alvarez, M.A.; Moreno-Arribas, M.V. The problem of biogenic amines in fermented foods and the use of potential biogenic amine-degrading microorganisms as a solution. Trends Food Sci. Technol. 2014, 39, 146–155. [Google Scholar] [CrossRef]
- Razia, S.; Hadibarata, T.; Lau, S.Y. Acidophilic microorganisms in remediation of contaminants present in extremely acidic conditions. Bioprocess Biosyst. Eng. 2023, 46, 341–358. [Google Scholar] [CrossRef]
- Montaño López, J.; Duran, L.; Avalos, J.L. Physiological limitations and opportunities in microbial metabolic engineering. Nat. Rev. Microbiol. 2022, 20, 35–48. [Google Scholar] [CrossRef] [PubMed]
- Mathur, S.; Singh, R. Antibiotic resistance in food lactic acid bacteria—A review. Int. J. Food Microbiol. 2005, 105, 281–295. [Google Scholar] [CrossRef]
- Hawaz, E. Isolation and identification of probiotic lactic acid bacteria from curd and in vitro evaluation of its growth inhibition activities against pathogenic bacteria. Afr. J. Microbiol. Res. 2014, 8, 1419–1425. [Google Scholar]
- Cervantes-Elizarrarás, A.; Cruz-Cansino, N.d.S.; Ramírez-Moreno, E.; Vega-Sánchez, V.; Velázquez-Guadarrama, N.; Zafra-Rojas, Q.Y.; Piloni-Martini, J. In vitro probiotic potential of lactic acid bacteria isolated from aguamiel and pulque and antibacterial activity against pathogens. Appl. Sci. 2019, 9, 601. [Google Scholar] [CrossRef]
- Rodrigues, N.P.A.; Garcia, E.F.; de Souza, E.L. Selection of lactic acid bacteria with promising probiotic aptitudes from fruit and ability to survive in different food matrices. Braz. J. Microbiol. 2021, 52, 2257–2269. [Google Scholar] [CrossRef]
- Sakoui, S.; Derdak, R.; Addoum, B.; Pop, O.L.; Vodnar, D.C.; Suharoschi, R.; Soukri, A.; El Khalfi, B. The first study of probiotic properties and biological activities of lactic acid bacteria isolated from Bat guano from Er-rachidia, Morocco. LWT 2022, 159, 113224. [Google Scholar] [CrossRef]
- Pinto, A.; Barbosa, J.; Albano, H.; Isidro, J.; Teixeira, P. Screening of bacteriocinogenic lactic acid bacteria and their characterization as potential probiotics. Microorganisms 2020, 8, 393. [Google Scholar] [CrossRef]
- Irwin, J.A. Overview of extremophiles and their food and medical applications. In Physiological and Biotechnological Aspects of Extremophiles; Academic Press: Cambridge, MA, USA, 2020; pp. 65–87. [Google Scholar]
- Souza, T.S.P.d.; de Andrade, C.J.; Koblitz, M.G.B.; Fai, A.E.C. Microbial Peptidase in Food Processing: Current State of the Art and Future Trends. Catal. Lett. 2022, 153, 114–137. [Google Scholar] [CrossRef]
- Sharma, A.; Kawarabayasi, Y.; Satyanarayana, T. Acidophilic bacteria and archaea: Acid stable biocatalysts and their potential applications. Extremophiles 2012, 16, 1–19. [Google Scholar] [CrossRef] [PubMed]
- Moat, A.G.; Foster, J.W.; Spector, M.P. Microbial physiology; John Wiley & Sons: New York, NY, USA, 2002. [Google Scholar]
- Chen, X. Thriving at Low pH: Adaptation Mechanisms of Acidophiles. In Acidophiles—Fundamentals and Applications; IntechOpen: London, UK, 2021. [Google Scholar]
- Xu, J.; Guo, L.; Zhao, N.; Meng, X.; Zhang, J.; Wang, T.; Wei, X.; Fan, M. Response mechanisms to acid stress of acid-resistant bacteria and biotechnological applications in the food industry. Crit. Rev. Biotechnol. 2022, 43, 258–274. [Google Scholar] [CrossRef] [PubMed]
- YÖRÜK, G.; GÜNER, A. Laktik asit bakterilerinin sınıflandırılması ve Weissella türlerinin gıda mikrobiyolojisinde önemi. Atatürk Üniversitesi Vet. Bilim. Derg. 2011, 6, 163–176. [Google Scholar]
- Smit, G.; Smit, B.A.; Engels, W.J. Flavour formation by lactic acid bacteria and biochemical flavour profiling of cheese products. FEMS Microbiol. Rev. 2005, 29, 591–610. [Google Scholar] [CrossRef]
- Johansen, E. Use of natural selection and evolution to develop new starter cultures for fermented foods. Annu. Rev. Food Sci. Technol. 2018, 9, 411–428. [Google Scholar] [CrossRef]
- Garvie, E.I. Bacterial lactate dehydrogenases. Microbiol. Rev. 1980, 44, 106–139. [Google Scholar] [CrossRef] [PubMed]
- Mannaa, M.; Han, G.; Seo, Y.S.; Park, I. Evolution of Food Fermentation Processes and the Use of Multi-Omics in Deciphering the Roles of the Microbiota. Foods 2021, 10, 2861. [Google Scholar] [CrossRef]
- du Toit, M.; Engelbrecht, L.; Lerm, E.; Krieger-Weber, S. Lactobacillus: The next generation of malolactic fermentation starter cultures—An overview. Food Bioprocess Technol. 2011, 4, 876–906. [Google Scholar] [CrossRef]
- Gürsoy, O.; kınık, Ö. Lactobacıllı and theır usage potentıal ın probıotıc cheese productıon. J. Eng. Sci. 2005, 11, 361–371. [Google Scholar]
- Courtin, P.; Rul, F. Interactions between microorganisms in a simple ecosystem: Yogurt bacteria as a study model. Le Lait 2004, 84, 125–134. [Google Scholar] [CrossRef]
- Aslam, S.; Qazi, J.I. Isolation of acidophilic lactic acid bacteria antagonistic to microbial contaminants. Pak. J. Zool. 2010, 42, 567–573. [Google Scholar]
- Laranjo, M.; Potes, M.E.; Elias, M. Role of Starter Cultures on the Safety of Fermented Meat Products. Front. Microbiol. 2019, 10, 853. [Google Scholar] [CrossRef] [PubMed]
- Kleerebezem, M.; Bachmann, H.; van Pelt-KleinJan, E.; Douwenga, S.; Smid, E.J.; Teusink, B.; van Mastrigt, O. Lifestyle, metabolism and environmental adaptation in Lactococcus lactis. FEMS Microbiol. Rev. 2020, 44, 804–820. [Google Scholar] [CrossRef] [PubMed]
- Bourdichon, F.; Boyaval, P.; Casaregola, S.; Dupont, J.; Farrokh, C.; Frisvad, J.; Hammes, W.; Huys, G.; Jany, J.; Laulund, S. 3 The 2012 Inventory of Microbial Species with technological beneficial role in fermented food products. Bull. Int. Dairy Fed. 2012, 455, 22–61. [Google Scholar]
- Tzora, A.; Nelli, A.; Voidarou, C.; Fthenakis, G.; Rozos, G.; Theodorides, G.; Bonos, E.; Skoufos, I. Microbiota “Fingerprint” of Greek Feta Cheese through Ripening. Appl. Sci. 2021, 11, 5631. [Google Scholar] [CrossRef]
- Russell, J.B.; Diez-Gonzalez, F. The effects of fermentation acids on bacterial growth. Adv. Microb. Physiol. 1997, 39, 205–234. [Google Scholar]
- De Clerck, E.; Rodriguez-Diaz, M.; Forsyth, G.; Lebbe, L.; Logan, N.A.; DeVos, P. Polyphasic characterization of Bacillus coagulans strains, illustrating heterogeneity within this species, and emended description of the species. Syst. Appl. Microbiol. 2004, 27, 50–60. [Google Scholar] [CrossRef]
- Haberbeck, L.U.; da Silva Riehl, C.A.; Salomão, B.d.C.M.; De Aragao, G.M.F. Bacillus coagulans spore inactivation through the application of oregano essential oil and heat. LWT-Food Sci. Technol. 2012, 46, 267–273. [Google Scholar] [CrossRef][Green Version]
- Lucas, R.; Grande, M.J.; Abriouel, H.; Maqueda, M.; Omar, N.B.; Valdivia, E.; Martínez-Cañamero, M.; Gálvez, A. Application of the broad-spectrum bacteriocin enterocin AS-48 to inhibit Bacillus coagulans in canned fruit and vegetable foods. Food Chem. Toxicol. 2006, 44, 1774–1781. [Google Scholar] [CrossRef] [PubMed]
- Silva, F.V.; Gibbs, P. Target selection in designing pasteurization processes for shelf-stable high-acid fruit products. Crit. Rev. Food Sci. Nutr. 2004, 44, 353–360. [Google Scholar] [CrossRef] [PubMed]
- Danyluk, M.D.; Friedrich, L.M.; Jouquand, C.; Goodrich-Schneider, R.; Parish, M.E.; Rouseff, R. Prevalence, concentration, spoilage, and mitigation of Alicyclobacillus spp. in tropical and subtropical fruit juice concentrates. Food Microbiol. 2011, 28, 472–477. [Google Scholar] [CrossRef] [PubMed]
- Chizhayeva, A.; Amangeldi, A.; Oleinikova, Y.; Alybaeva, A.; Sadanov, A. Lactic acid bacteria as probiotics in sustainable development of aquaculture. Aquat. Living Resour. 2022, 35, 10. [Google Scholar] [CrossRef]
- Sharma, H.; Ozogul, F.; Bartkiene, E.; Rocha, J.M. Impact of lactic acid bacteria and their metabolites on the techno-functional properties and health benefits of fermented dairy products. Crit. Rev. Food Sci. Nutr. 2023, 63, 4819–4841. [Google Scholar] [CrossRef] [PubMed]
- Grattepanche, F.; Miescher-Schwenninger, S.; Meile, L.; Lacroix, C. Recent developments in cheese cultures with protective and probiotic functionalities. Dairy Sci. Technol. 2008, 88, 421–444. [Google Scholar] [CrossRef]
- Hammes, W.P.; Tichaczek, P.S. The potential of lactic acid bacteria for the production of safe and wholesome food. Z. Lebensm.-Unters. Und-Forsch. 1994, 198, 193–201. [Google Scholar] [CrossRef]
- Wafula, E.N.; Muhonja, C.N.; Kuja, J.O.; Owaga, E.E.; Makonde, H.M.; Mathara, J.M.; Kimani, V.W. Lactic Acid Bacteria from African Fermented Cereal-Based Products: Potential Biological Control Agents for Mycotoxins in Kenya. J. Toxicol. 2022, 2022, 2397767. [Google Scholar] [CrossRef]
- Emkani, M.; Oliete, B.; Saurel, R. Effect of Lactic Acid Fermentation on Legume Protein Properties, a Review. Fermentation 2022, 8, 244. [Google Scholar] [CrossRef]
- Kumari, M.; Kokkiligadda, A.; Dasriya, V.; Naithani, H. Functional relevance and health benefits of soymilk fermented by lactic acid bacteria. J. Appl. Microbiol. 2022, 133, 104–119. [Google Scholar] [CrossRef]
- Zhu, L.; Mu, T.; Ma, M.; Sun, H.; Zhao, G. Nutritional composition, antioxidant activity, volatile compounds, and stability properties of sweet potato residues fermented with selected lactic acid bacteria and bifidobacteria. Food Chem. 2022, 374, 131500. [Google Scholar] [CrossRef]
- Burakova, I.; Smirnova, Y.; Gryaznova, M.; Syromyatnikov, M.; Chizhkov, P.; Popov, E.; Popov, V. The Effect of Short-Term Consumption of Lactic Acid Bacteria on the Gut Microbiota in Obese People. Nutrients 2022, 14, 3384. [Google Scholar] [CrossRef] [PubMed]
- FAO; WHO. Guidelines for the Evaluation of Probiotics in Food, Report of a Joint FAO/WHO Working Group on Drafting Guideline for the Evaluation of Probiotic in Food; World Health Organization: Geneva, Switzerland, 2002. [Google Scholar]
- Prabhurajeshwar, C.; Chandrakanth, R.K. Probiotic potential of Lactobacilli with antagonistic activity against pathogenic strains: An in vitro validation for the production of inhibitory substances. Biomed. J. 2017, 40, 270–283. [Google Scholar] [CrossRef] [PubMed]
- Patel, A.K.; Ahire, J.J.; Pawar, S.P.; Chaudhari, B.L.; Chincholkar, S.B. Comparative accounts of probiotic characteristics of Bacillus spp. isolated from food wastes. Food Res. Int. 2009, 42, 505–510. [Google Scholar] [CrossRef]
- Lee, H.-W.; Park, Y.-S.; Jung, J.-S.; Shin, W.-S.J.A. Chitosan oligosaccharides, dp 2–8, have prebiotic effect on the Bifidobacterium bifidium and Lactobacillus sp. Anaerobe 2002, 8, 319–324. [Google Scholar] [CrossRef]
- Kaur, M.; Singh, H.; Jangra, M.; Kaur, L.; Jaswal, P.; Dureja, C.; Nandanwar, H.; Chaudhuri, S.R.; Raje, M.; Mishra, S. Lactic acid bacteria isolated from yak milk show probiotic potential. Appl. Microbiol. Biotechnol. 2017, 101, 7635–7652. [Google Scholar] [CrossRef]
- Saxami, G.; Ypsilantis, P.; Sidira, M.; Simopoulos, C.; Kourkoutas, Y.; Galanis, A. Distinct adhesion of probiotic strain Lactobacillus casei ATCC 393 to rat intestinal mucosa. Anaerobe 2012, 18, 417–420. [Google Scholar] [CrossRef] [PubMed]
- El-Sayed, A.I.; El-Borai, A.M.; Akl, S.H.; El-Aassar, S.A.; Abdel-Latif, M.S. Identification of Lactobacillus strains from human mother milk and cottage cheese revealed potential probiotic properties with enzymatic activity. Sci. Rep. 2022, 12, 22522. [Google Scholar] [CrossRef]
- Vemuri, R.; Shinde, T.; Shastri, M.D.; Perera, A.P.; Tristram, S.; Martoni, C.J.; Gundamaraju, R.; Ahuja, K.D.; Ball, M.; Eri, R. A human origin strain Lactobacillus acidophilus DDS-1 exhibits superior in vitro probiotic efficacy in comparison to plant or dairy origin probiotics. Int. J. Med. Sci. 2018, 15, 840. [Google Scholar] [CrossRef]
- Byakika, S.; Mukisa, I.M.; Byaruhanga, Y.B.; Muyanja, C. A review of criteria and methods for evaluating the probiotic potential of microorganisms. Food Rev. Int. 2019, 35, 427–466. [Google Scholar] [CrossRef]
- Lee, K.W.; Shim, J.M.; Park, S.-K.; Heo, H.-J.; Kim, H.-J.; Ham, K.-S.; Kim, J.H. Isolation of lactic acid bacteria with probiotic potentials from kimchi, traditional Korean fermented vegetable. LWT-Food Sci. Technol. 2016, 71, 130–137. [Google Scholar] [CrossRef]
- Bao, Y.; Zhang, Y.; Zhang, Y.; Liu, Y.; Wang, S.; Dong, X.; Wang, Y.; Zhang, H. Screening of potential probiotic properties of Lactobacillus fermentum isolated from traditional dairy products. Food Control 2010, 21, 695–701. [Google Scholar] [CrossRef]
- Osmanagaoglu, O.; Kiran, F.; Ataoglu, H. Evaluation of in vitro probiotic potential of Pediococcus pentosaceus OZF isolated from human breast milk. Probiotics Antimicrob. Proteins 2010, 2, 162–174. [Google Scholar] [CrossRef] [PubMed]
- Bujnakova, D.; Strakova, E.; Kmet, V. In vitro evaluation of the safety and probiotic properties of Lactobacilli isolated from chicken and calves. Anaerobe 2014, 29, 118–127. [Google Scholar] [CrossRef] [PubMed]
- Comerlato, C.B.; Prichula, J.; Siqueira, F.M.; Ritter, A.C.; Varela, A.P.M.; Mayer, F.Q.; Brandelli, A. Genomic analysis of Enterococcus durans LAB18S, a potential probiotic strain isolated from cheese. Genet. Mol. Biol. 2022, 45, e20210201. [Google Scholar] [CrossRef]
- Srinivash, M.; Krishnamoorthi, R.; Mahalingam, P.U.; Malaikozhundan, B.; Keerthivasan, M. Probiotic potential of exopolysaccharide producing lactic acid bacteria isolated from homemade fermented food products. J. Agric. Food Res. 2023, 11, 100517. [Google Scholar] [CrossRef]
- Falfán-Cortés, R.N.; Mora-Peñaflor, N.; Gómez-Aldapa, C.A.; Rangel-Vargas, E.; Acevedo-Sandoval, O.A.; Franco-Fernández, M.J.; Castro-Rosas, J. Characterization and evaluation of the probiotic potential in vitro and in situ of Lacticaseibacillus paracasei isolated from tenate cheese. J. Food Prot. 2022, 85, 112–121. [Google Scholar] [CrossRef]
- Guo, X.; Long, R.; Kreuzer, M.; Ding, L.; Shang, Z.; Zhang, Y.; Yang, Y.; Cui, G. Importance of functional ingredients in yak milk-derived food on health of Tibetan nomads living under high-altitude stress: A review. Crit. Rev. Food Sci. Nutr. 2014, 54, 292–302. [Google Scholar] [CrossRef]
- Chen, X.; Du, X.; Wang, W.; Zhang, J.; Sun, Z.; Lıu, W.; Li, L.; Sun, T.; Zhang, H. Isolation and identification of cultivable lactic acid bacteria in traditional fermented milk of Tibet in China. Int. J. Dairy Technol. 2010, 63, 437–444. [Google Scholar]
- Stoeker, L.; Nordone, S.; Gunderson, S.; Zhang, L.; Kajikawa, A.; LaVoy, A.; Miller, M.; Klaenhammer, T.R.; Dean, G.A. Assessment of Lactobacillus gasseri as a candidate oral vaccine vector. Clin. Vaccine Immunol. 2011, 18, 1834–1844. [Google Scholar] [CrossRef]
- Sharma, A.; Lavania, M.; Singh, R.; Lal, B. Identification and probiotic potential of lactic acid bacteria from camel milk. Saudi J. Biol. Sci. 2021, 28, 1622–1632. [Google Scholar] [CrossRef]
- Yasmin, I.; Saeed, M.; Khan, W.A.; Khaliq, A.; Chughtai, M.F.J.; Iqbal, R.; Tehseen, S.; Naz, S.; Liaqat, A.; Mehmood, T. In vitro probiotic potential and safety evaluation (hemolytic, cytotoxic activity) of Bifidobacterium strains isolated from raw camel milk. Microorganisms 2020, 8, 354. [Google Scholar] [CrossRef] [PubMed]
- Akinyemi, M.O.; Ogunremi, O.R.; Adeleke, R.A.; Ezekiel, C.N. Probiotic Potentials of Lactic Acid Bacteria and Yeasts from Raw Goat Milk in Nigeria. Probiotics Antimicrob. Proteins 2022, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Islam, M.Z.; Uddin, M.E.; Rahman, M.T.; Islam, M.A.; Harun-ur-Rashid, M. Isolation and characterization of dominant lactic acid bacteria from raw goat milk: Assessment of probiotic potential and technological properties. Small Rumin. Res. 2021, 205, 106532. [Google Scholar] [CrossRef]
- Grujović, M.Ž.; Mladenović, K.G.; Semedo-Lemsaddek, T.; Laranjo, M.; Stefanović, O.D.; Kocić-Tanackov, S.D. Advantages and disadvantages of non-starter lactic acid bacteria from traditional fermented foods: Potential use as starters or probiotics. Compr. Rev. Food Sci. Food Saf. 2022, 21, 1537–1567. [Google Scholar] [CrossRef]
- Sadeghi, M.; Panahi, B.; Mazlumi, A.; Hejazi, M.A.; Komi, D.E.A.; Nami, Y. Screening of potential probiotic lactic acid bacteria with antimicrobial properties and selection of superior bacteria for application as biocontrol using machine learning models. LWT 2022, 162, 113471. [Google Scholar] [CrossRef]
- Khan, I.; Kang, S.C. Probiotic potential of nutritionally improved Lactobacillus plantarum DGK-17 isolated from Kimchi–A traditional Korean fermented food. Food Control 2016, 60, 88–94. [Google Scholar] [CrossRef]
- Sagdic, O.; Ozturk, I.; Yapar, N.; Yetim, H. Diversity and probiotic potentials of lactic acid bacteria isolated from gilaburu, a traditional Turkish fermented European cranberrybush (Viburnum opulus L.) fruit drink. Food Res. Int. 2014, 64, 537–545. [Google Scholar] [CrossRef] [PubMed]
- Metrouh, R.; Fares, R.; Mechai, A.; Debabza, M.; Menasria, T. Technological properties and probiotic potential of Lactiplantibacillus plantarum SJ14 isolated from Algerian traditional cheese “Jben”. J. Food Process. Preserv. 2022, 46, e16482. [Google Scholar] [CrossRef]
- Coimbra-Gomes, J.; Reis, P.J.; Tavares, T.G.; Malcata, F.X.; Macedo, A.C. Study of lactic acid bacteria biodiversity in fermented Cobrançosa table olives to determine their probiotic potential. Foods 2022, 11, 3050. [Google Scholar] [CrossRef]
- Liu, C.; Xue, W.-j.; Ding, H.; An, C.; Ma, S.-j.; Liu, Y. Probiotic Potential of Lactobacillus Strains Isolated From Fermented Vegetables in Shaanxi, China. Front. Microbiol. 2022, 12, 4168. [Google Scholar] [CrossRef]
- Akmal, U.; Ghori, I.; Elasbali, A.M.; Alharbi, B.; Farid, A.; Alamri, A.S.; Muzammal, M.; Asdaq, S.M.B.; Naiel, M.A.; Ghazanfar, S. Probiotic and antioxidant potential of the Lactobacillus spp. isolated from artisanal fermented pickles. Fermentation 2022, 8, 328. [Google Scholar] [CrossRef]
- Leite, A.M.; Miguel, M.; Peixoto, R.; Ruas-Madiedo, P.; Paschoalin, V.; Mayo, B.; Delgado, S. Probiotic potential of selected lactic acid bacteria strains isolated from Brazilian kefir grains. J. Dairy Sci. 2015, 98, 3622–3632. [Google Scholar] [CrossRef]
- Olajugbagbe, T.E.; Elugbadebo, O.E.; Omafuvbe, B.O. Probiotic potentials of Pediococuss acidilactici isolated from wara; A Nigerian unripened soft cheese. Heliyon 2020, 6, e04889. [Google Scholar] [CrossRef] [PubMed]
- Salomskiene, J.; Jonkuviene, D.; Macioniene, I.; Abraitiene, A.; Zeime, J.; Repeckiene, J.; Vaiciulyte-Funk, L. Differences in the occurence and efficiency of antimicrobial compounds produced by lactic acid bacteria. Eur. Food Res. Technol. 2019, 245, 569–579. [Google Scholar] [CrossRef]
- Bangar, S.P.; Suri, S.; Trif, M.; Ozogul, F. Organic acids production from lactic acid bacteria: A preservation approach. Food Biosci. 2022, 46, 101615. [Google Scholar] [CrossRef]
- Wang, C.; Chang, T.; Yang, H.; Cui, M. Antibacterial mechanism of lactic acid on physiological and morphological properties of Salmonella Enteritidis, Escherichia coli and Listeria monocytogenes. Food Control 2015, 47, 231–236. [Google Scholar] [CrossRef]
- Hati, S.; Patel, M.; Mishra, B.K.; Das, S. Short-chain fatty acid and vitamin production potentials of Lactobacillus isolated from fermented foods of Khasi Tribes, Meghalaya, India. Ann. Microbiol. 2019, 69, 1191–1199. [Google Scholar] [CrossRef]
- Gonmei, Z.; Toteja, G. Micronutrient status of Indian population. Indian J. Med. Res. 2018, 148, 511. [Google Scholar]
- LeBlanc, J.; Laiño, J.E.; Del Valle, M.J.; Vannini, V.v.; van Sinderen, D.; Taranto, M.P.; de Valdez, G.F.; de Giori, G.S.; Sesma, F. B-Group vitamin production by lactic acid bacteria–current knowledge and potential applications. J. Appl. Microbiol. 2011, 111, 1297–1309. [Google Scholar] [CrossRef]
- Goswami, G.; Bora, S.S.; Parveen, A.; Boro, R.C.; Barooah, M. Identification and functional properties of dominant lactic acid bacteria isolated from Kahudi, a traditional rapeseed fermented food product of Assam, India. J. Ethn. Foods 2017, 4, 187–197. [Google Scholar] [CrossRef]
- LeBlanc, J.G.; Levit, R.; Savoy de Giori, G.; de Moreno de LeBlanc, A. Application of vitamin-producing lactic acid bacteria to treat intestinal inflammatory diseases. Appl. Microbiol. Biotechnol. 2020, 104, 3331–3337. [Google Scholar] [CrossRef] [PubMed]
- Santos, F.; Wegkamp, A.; de Vos, W.M.; Smid, E.J.; Hugenholtz, J. High-level folate production in fermented foods by the B12 producer Lactobacillus reuteri JCM1112. Appl. Environ. Microbiol. 2008, 74, 3291–3294. [Google Scholar] [CrossRef]
- Masuda, M.; Ide, M.; Utsumi, H.; Niiro, T.; Shimamura, Y.; Murata, M. Production potency of folate, vitamin B12, and thiamine by lactic acid bacteria isolated from Japanese pickles. Biosci. Biotechnol. Biochem. 2012, 76, 2061–2067. [Google Scholar] [CrossRef]
- Carrizo, S.L.; de LeBlanc, A.d.M.; LeBlanc, J.G.; Rollán, G.C. Quinoa pasta fermented with lactic acid bacteria prevents nutritional deficiencies in mice. Food Res. Int. 2020, 127, 108735. [Google Scholar] [CrossRef] [PubMed]
- Laiño, J.E.; del Valle, M.J.; de Giori, G.S.; LeBlanc, J.G.J. Development of a high folate concentration yogurt naturally bio-enriched using selected lactic acid bacteria. LWT-Food Sci. Technol. 2013, 54, 1–5. [Google Scholar] [CrossRef]
- Laiño, J.E.; LeBlanc, J.G.; Savoy de Giori, G. Production of natural folates by lactic acid bacteria starter cultures isolated from artisanal Argentinean yogurts. Can. J. Microbiol. 2012, 58, 581–588. [Google Scholar] [CrossRef]
- Molina, V.; Médici, M.; de Valdez, G.F.; Taranto, M.P. Soybean-based functional food with vitamin B12-producing lactic acid bacteria. J. Funct. Foods 2012, 4, 831–836. [Google Scholar] [CrossRef]
- Thakur, K.; Tomar, S.K.; De, S. Lactic acid bacteria as a cell factory for riboflavin production. Microb. Biotechnol. 2016, 9, 441–451. [Google Scholar] [CrossRef]
- Jitpakdee, J.; Kantachote, D.; Kanzaki, H.; Nitoda, T. Selected probiotic lactic acid bacteria isolated from fermented foods for functional milk production: Lower cholesterol with more beneficial compounds. LWT 2021, 135, 110061. [Google Scholar] [CrossRef]
- Ratanaburee, A.; Kantachote, D.; Charernjiratrakul, W.; Sukhoom, A. Selection of γ-aminobutyric acid-producing lactic acid bacteria and their potential as probiotics for use as starter cultures in T hai fermented sausages (N ham). Int. J. Food Sci. Technol. 2013, 48, 1371–1382. [Google Scholar] [CrossRef]
- Ratanaburee, A.; Kantachote, D.; Charernjiratrakul, W.; Penjamras, P.; Chaiyasut, C. Enhancement of γ-aminobutyric acid in a fermented red seaweed beverage by starter culture Lactobacillus plantarum DW12. Electron. J. Biotechnol. 2011, 14, 1. [Google Scholar]
- Park, K.-B.; Oh, S.-H. Production of yogurt with enhanced levels of gamma-aminobutyric acid and valuable nutrients using lactic acid bacteria and germinated soybean extract. Bioresour. Technol. 2007, 98, 1675–1679. [Google Scholar] [CrossRef] [PubMed]
- Ratanaburee, A.; Kantachote, D.; Charernjiratrakul, W.; Sukhoom, A. Enhancement of γ-aminobutyric acid (GABA) in Nham (Thai fermented pork sausage) using starter cultures of Lactobacillus namurensis NH2 and Pediococcus pentosaceus HN8. Int. J. Food Microbiol. 2013, 167, 170–176. [Google Scholar] [CrossRef] [PubMed]
- Lozano, J.; Fernández-Ciganda, S.; González Revello, Á.; Hirigoyen, D.; Martínez, M.; Scorza, C.; Zunino, P. Probiotic potential of GABA-producing lactobacilli isolated from Uruguayan artisanal cheese starter cultures. J. Appl. Microbiol. 2022, 133, 1610–1619. [Google Scholar] [CrossRef]
- Kim, J.Y.; Lee, M.Y.; Ji, G.E.; Lee, Y.S.; Hwang, K.T. Production of γ-aminobutyric acid in black raspberry juice during fermentation by Lactobacillus brevis GABA100. Int. J. Food Microbiol. 2009, 130, 12–16. [Google Scholar] [CrossRef]
- Kolev, P.; Rocha-Mendoza, D.; Ruiz-Ramírez, S.; Ortega-Anaya, J.; Jiménez-Flores, R.; García-Cano, I. Screening and characterization of β-galactosidase activity in lactic acid bacteria for the valorization of acid whey. JDS Commun. 2022, 3, 1–6. [Google Scholar] [CrossRef]
- Panda, S.H.; Ray, R.C. Amylolytic Lactic Acid Bacteria. İn Fermented Foods, Part I: Biochemistry and Biotechnology; CRC Press: Boca Raton, FL, USA, 2016; p. 133. [Google Scholar]
- Xu, Y.; Zhou, T.; Tang, H.; Li, X.; Chen, Y.; Zhang, L.; Zhang, J. Probiotic potential and amylolytic properties of lactic acid bacteria isolated from Chinese fermented cereal foods. Food Control 2020, 111, 107057. [Google Scholar] [CrossRef]
- Paul, J.S.; Gupta, N.; Beliya, E.; Tiwari, S.; Jadhav, S.K. Aspects and recent trends in microbial α-amylase: A review. Appl. Biochem. Biotechnol. 2021, 193, 2649–2698. [Google Scholar] [CrossRef]
- Pasin, T.M.; dos Anjos Moreira, E.; de Lucas, R.C.; Benassi, V.M.; Ziotti, L.S.; Cereia, M.; Polizeli, M.d.L.T.d.M. Novel amylase-producing fungus hydrolyzing wheat and brewing residues, Aspergillus carbonarius, discovered in tropical forest remnant. Folia Microbiol. 2020, 65, 173–184. [Google Scholar] [CrossRef]
- Niyonzima, F.N.; Veena, S.; More, S.S. Industrial production and optimization of microbial enzymes. In Microbial Enzymes: Roles and Applications in İndustries; Springer: Singapore, 2020; pp. 115–135. [Google Scholar]
- Mokoena, M.P. Lactic acid bacteria and their bacteriocins: Classification, biosynthesis and applications against uropathogens: A mini-review. Molecules 2017, 22, 1255. [Google Scholar] [CrossRef]
- Parada, J.L.; Caron, C.R.; Medeiros, A.B.P.; Soccol, C.R. Bacteriocins from lactic acid bacteria: Purification, properties and use as biopreservatives. Braz. Arch. Biol. Technol. 2007, 50, 512–542. [Google Scholar] [CrossRef]
- da Costa, R.J.; Voloski, F.L.; Mondadori, R.G.; Duval, E.H.; Fiorentini, Â.M. Preservation of meat products with bacteriocins produced by lactic acid bacteria isolated from meat. J. Food Qual. 2019, 2019, 4726510. [Google Scholar] [CrossRef]
- Vieco-Saiz, N.; Belguesmia, Y.; Raspoet, R.; Auclair, E.; Gancel, F.; Kempf, I.; Drider, D. Benefits and inputs from lactic acid bacteria and their bacteriocins as alternatives to antibiotic growth promoters during food-animal production. Front. Microbiol. 2019, 10, 57. [Google Scholar] [CrossRef] [PubMed]
- Brown, K.; Uwiera, R.R.; Kalmokoff, M.L.; Brooks, S.P.; Inglis, G.D. Antimicrobial growth promoter use in livestock: A requirement to understand their modes of action to develop effective alternatives. Int. J. Antimicrob. Agents 2017, 49, 12–24. [Google Scholar] [CrossRef] [PubMed]
- Laxminarayan, R.; Van Boeckel, T.; Teillant, A. The Economic Costs of Withdrawing Antimicrobial Growth Promoters from the Livestock Sector; OECD Food, Agriculture and Fisheries Papers, No. 78; OECD Publishing: Paris, France, 2015. [Google Scholar] [CrossRef]
- Seal, B.S.; Drider, D.; Oakley, B.B.; Brüssow, H.; Bikard, D.; Rich, J.O.; Miller, S.; Devillard, E.; Kwan, J.; Bertin, G. Microbial-derived products as potential new antimicrobials. Vet. Res. 2018, 49, 66. [Google Scholar] [CrossRef] [PubMed]
- Egan, K.; Field, D.; Rea, M.C.; Ross, R.P.; Hill, C.; Cotter, P.D. Bacteriocins: Novel solutions to age old spore-related problems? Front. Microbiol. 2016, 7, 461. [Google Scholar] [CrossRef]
- Liu, S.; Wilkinson, B.J.; Bischoff, K.M.; Hughes, S.R.; Rich, J.O.; Cotta, M.A. Novel antibacterial polypeptide laparaxin produced by Lactobacillus paracasei strain NRRL B-50314 via fermentation. J. Pet. Environ. Biotechnol. 2012, 3. [Google Scholar] [CrossRef]
- Todorov, S.D.; Dicks, L.M. Pediocin ST18, an anti-listerial bacteriocin produced by Pediococcus pentosaceus ST18 isolated from boza, a traditional cereal beverage from Bulgaria. Process Biochem. 2005, 40, 365–370. [Google Scholar] [CrossRef]
- Jiao, D.; Liu, Y.; Zeng, R.; Hou, X.; Nie, G.; Sun, L.; Fang, Z. Preparation of phosphatidylcholine nanovesicles containing bacteriocin CAMT2 and their anti-listerial activity. Food Chem. 2020, 314, 126244. [Google Scholar] [CrossRef]
- Tiwari, S.K.; Dicks, L.M.; Popov, I.V.; Karaseva, A.; Ermakov, A.M.; Suvorov, A.; Tagg, J.R.; Weeks, R.; Chikindas, M.L. Probiotics at war against viruses: What is missing from the picture? Front. Microbiol. 2020, 11, 1877. [Google Scholar] [CrossRef]
- Le Lay, C.; Fernandez, B.; Hammami, R.; Ouellette, M.; Fliss, I. On Lactococcus lactis UL719 competitivity and nisin (Nisaplin®) capacity to inhibit Clostridium difficile in a model of human colon. Front. Microbiol. 2015, 6, 1020. [Google Scholar] [CrossRef] [PubMed]
- Osmanağaoğlu, Ö. Detection and characterization of Leucocin OZ, a new anti-listerial bacteriocin produced by Leuconostoc carnosum with a broad spectrum of activity. Food Control 2007, 18, 118–123. [Google Scholar] [CrossRef]
- Vardakas, K.Z.; Polyzos, K.A.; Patouni, K.; Rafailidis, P.I.; Samonis, G.; Falagas, M.E. Treatment failure and recurrence of Clostridium difficile infection following treatment with vancomycin or metronidazole: A systematic review of the evidence. Int. J. Antimicrob. Agents 2012, 40, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Blay, G.L.; Lacroix, C.; Zihler, A.; Fliss, I. In vitro inhibition activity of nisin A, nisin Z, pediocin PA-1 and antibiotics against common intestinal bacteria. Lett. Appl. Microbiol. 2007, 45, 252–257. [Google Scholar] [CrossRef] [PubMed]
- Lay, C.L.; Dridi, L.; Bergeron, M.G.; Ouellette, M.; Fliss, I.l. Nisin is an effective inhibitor of Clostridium difficile vegetative cells and spore germination. J. Med. Microbiol. 2016, 65, 169–175. [Google Scholar] [CrossRef]
- Al Kassaa, I.; Hober, D.; Hamze, M.; Chihib, N.E.; Drider, D. Antiviral potential of lactic acid bacteria and their bacteriocins. Probiotics Antimicrob. Proteins 2014, 6, 177–185. [Google Scholar] [CrossRef]
- Maeda, N.; Nakamura, R.; Hirose, Y.; Murosaki, S.; Yamamoto, Y.; Kase, T.; Yoshikai, Y. Oral administration of heat-killed Lactobacillus plantarum L-137 enhances protection against influenza virus infection by stimulation of type I interferon production in mice. Int. Immunopharmacol. 2009, 9, 1122–1125. [Google Scholar] [CrossRef]
- Fernández, M.; Hudson, J.A.; Korpela, R.; de los Reyes-Gavilán, C.G. Impact on human health of microorganisms present in fermented dairy products: An overview. BioMed Res. Int. 2015, 2015, 412714. [Google Scholar] [CrossRef]
- Deepika Priyadarshani, W.M.; Rakshit, S.K. Screening selected strains of probiotic lactic acid bacteria for their ability to produce biogenic amines (histamine and tyramine). Int. J. Food Sci. Technol. 2011, 46, 2062–2069. [Google Scholar] [CrossRef]
- Pegg, A.E. Toxicity of polyamines and their metabolic products. Chem. Res. Toxicol. 2013, 26, 1782–1800. [Google Scholar] [CrossRef] [PubMed]
- Marcobal, A.; De Las Rivas, B.; Landete, J.M.; Tabera, L.; Muñoz, R. Tyramine and phenylethylamine biosynthesis by food bacteria. Crit. Rev. Food Sci. Nutr. 2012, 52, 448–467. [Google Scholar] [CrossRef] [PubMed]
- Hazards, E.P.o.B. Scientific opinion on risk based control of biogenic amine formation in fermented foods. Efsa J. 2011, 9, 2393. [Google Scholar]
- Álvarez-Cisneros, Y.M.; Ponce-Alquicira, E. Antibiotic resistance in lactic acid bacteria. In Antimicrobial Resistance—A Global Threat; IntechOpen: London, UK, 2018. [Google Scholar]
- EFSA Panel on Additives and Products or Substances used in Animal Feed (FEEDAP). Guidance on the assessment of bacterial susceptibility to antimicrobials of human and veterinary importance. EFSA J. 2012, 10, 2740. [Google Scholar]
- Wang, X.; Wang, W.; Lv, H.; Zhang, H.; Liu, Y.; Zhang, M.; Wang, Y.; Tan, Z. Probiotic potential and wide-spectrum antimicrobial activity of lactic acid bacteria isolated from infant feces. Probiotics Antimicrob. Proteins 2021, 13, 90–101. [Google Scholar] [CrossRef]
- Mangia, N.P.; Saliba, L.; Deiana, P. Functional and safety characterization of autochthonous Lactobacillus paracasei FS103 isolated from sheep cheese and its survival in sheep and cow fermented milks during cold storage. Ann. Microbiol. 2019, 69, 161–170. [Google Scholar] [CrossRef]
- Abushelaibi, A.; Al-Mahadin, S.; El-Tarabily, K.; Shah, N.P.; Ayyash, M. Characterization of potential probiotic lactic acid bacteria isolated from camel milk. LWT-Food Sci. Technol. 2017, 79, 316–325. [Google Scholar] [CrossRef]
- Oh, Y.J.; Jung, D.S. Evaluation of probiotic properties of Lactobacillus and Pediococcus strains isolated from Omegisool, a traditionally fermented millet alcoholic beverage in Korea. LWT-Food Sci. Technol. 2015, 63, 437–444. [Google Scholar] [CrossRef]
- Argyri, A.A.; Zoumpopoulou, G.; Karatzas, K.-A.G.; Tsakalidou, E.; Nychas, G.-J.E.; Panagou, E.Z.; Tassou, C.C. Selection of potential probiotic lactic acid bacteria from fermented olives by in vitro tests. Food Microbiol. 2013, 33, 282–291. [Google Scholar] [CrossRef]
- Ismael, M.; Gu, Y.; Cui, Y.; Wang, T.; Yue, F.; Yantin, Q.; Lü, X. Lactic acid bacteria isolated from Chinese traditional fermented milk as novel probiotic strains and their potential therapeutic applications. 3 Biotech 2022, 12, 337. [Google Scholar] [CrossRef]
- Singhal, N.; Singh, N.S.; Mohanty, S.; Singh, P.; Virdi, J.S. Evaluation of probiotic characteristics of lactic acid bacteria isolated from two commercial preparations available in Indian market. Indian J. Microbiol. 2019, 59, 112–115. [Google Scholar] [CrossRef] [PubMed]
- Motey, G.A.; Owusu-Kwarteng, J.; Obiri-Danso, K.; Ofori, L.A.; Ellis, W.O.; Jespersen, L. In vitro properties of potential probiotic lactic acid bacteria originating from Ghanaian indigenous fermented milk products. World J. Microbiol. Biotechnol. 2021, 37, 52. [Google Scholar] [CrossRef]
- dos Santos Leandro, E.; Ginani, V.C.; de Alencar, E.R.; Pereira, O.G.; Rose, E.C.P.; do Vale, H.M.M.; Pratesi, R.; Hecht, M.M.; Cavalcanti, M.H.; Tavares, C.S.O. Isolation, identification, and screening of lactic acid bacteria with probiotic potential in silage of different species of forage plants, cocoa beans, and artisanal salami. Probiotics Antimicrob. Proteins 2021, 13, 173–186. [Google Scholar] [CrossRef]
- de Amorim Trindade, D.P.; Barbosa, J.P.; Martins, E.M.F.; Tette, P.A.S. Isolation and identification of lactic acid bacteria in fruit processing residues from the Brazilian Cerrado and its probiotic potential. Food Biosci. 2022, 48, 101739. [Google Scholar] [CrossRef]
- Somashekaraiah, R.; Shruthi, B.; Deepthi, B.; Sreenivasa, M. Probiotic properties of lactic acid bacteria isolated from neera: A naturally fermenting coconut palm nectar. Front. Microbiol. 2019, 10, 1382. [Google Scholar] [CrossRef] [PubMed]
- Colombo, M.; Nero, L.A.; Todorov, S.D. Safety profiles of beneficial lactic acid bacteria isolated from dairy systems. Braz. J. Microbiol. 2020, 51, 787–795. [Google Scholar] [CrossRef] [PubMed]
- Meena, K.K.; Taneja, N.K.; Jain, D.; Ojha, A.; Kumawat, D.; Mishra, V. In Vitro Assessment of Probiotic and Technological Properties of Lactic Acid Bacteria Isolated from Indigenously Fermented Cereal-Based Food Products. Fermentation 2022, 8, 529. [Google Scholar] [CrossRef]
- da Silva Ferrari, I.; de Souza, J.V.; Ramos, C.L.; da Costa, M.M.; Schwan, R.F.; Dias, F.S. Selection of autochthonous lactic acid bacteria from goat dairies and their addition to evaluate the inhibition of Salmonella typhi in artisanal cheese. Food Microbiol. 2016, 60, 29–38. [Google Scholar] [CrossRef]
- Missaoui, J.; Saidane, D.; Mzoughi, R.; Minervini, F. Fermented seeds (“Zgougou”) from aleppo pine as a novel source of potentially probiotic lactic acid bacteria. Microorganisms 2019, 7, 709. [Google Scholar] [CrossRef]
- Javed, G.; Arshad, N.; Munir, A.; Khan, S.; Rasheed, S.; Hussain, I. Signature probiotic and pharmacological attributes of lactic acid bacteria isolated from human breast milk. Int. Dairy J. 2022, 127, 105297. [Google Scholar] [CrossRef]
- Bindu, A.; Lakshmidevi, N. Identification and in vitro evaluation of probiotic attributes of lactic acid bacteria isolated from fermented food sources. Arch. Microbiol. 2021, 203, 579–595. [Google Scholar] [CrossRef] [PubMed]
- Muñoz-Atienza, E.; Gómez-Sala, B.; Araújo, C.; Campanero, C.; Del Campo, R.; Hernández, P.E.; Herranz, C.; Cintas, L.M. Antimicrobial activity, antibiotic susceptibility and virulence factors of lactic acid bacteria of aquatic origin intended for use as probiotics in aquaculture. BMC Microbiol. 2013, 13, 15. [Google Scholar] [CrossRef] [PubMed]
- Ben Farhat, L.; Romeo, F.V.; Foti, P.; Russo, N.; Randazzo, C.L.; Caggia, C.; Abidi, F. Multi-Functional Potential of Lactic Acid Bacteria Strains and Antimicrobial Effects in Minimally Processed Pomegranate (Punica granatum L. cv Jolly Red) Arils. Microorganisms 2022, 10, 1876. [Google Scholar] [CrossRef] [PubMed]
- Banwo, K.; Asogwa, F.C.; Ogunremi, O.R.; Adesulu-Dahunsi, A.; Sanni, A. Nutritional profile and antioxidant capacities of fermented millet and sorghum gruels using lactic acid bacteria and yeasts. Food Biotechnol. 2021, 35, 199–220. [Google Scholar] [CrossRef]
- Kaktcham, P.M.; Temgoua, J.-B.; Zambou, F.N.; Diaz-Ruiz, G.; Wacher, C.; Pérez-Chabela, M.d.L. In vitro evaluation of the probiotic and safety properties of bacteriocinogenic and non-bacteriocinogenic lactic acid bacteria from the intestines of Nile tilapia and common carp for their use as probiotics in aquaculture. Probiotics Antimicrob. Proteins 2018, 10, 98–109. [Google Scholar] [CrossRef]
- Domingos-Lopes, M.; Stanton, C.; Ross, P.; Dapkevicius, M.; Silva, C. Genetic diversity, safety and technological characterization of lactic acid bacteria isolated from artisanal Pico cheese. Food Microbiol. 2017, 63, 178–190. [Google Scholar] [CrossRef]
- Ribeiro, S.; Coelho, M.; Todorov, S.D.; Franco, B.D.G.d.M.; Dapkevicius, M.; Silva, C. Technological properties of bacteriocin-producing lactic acid bacteria isolated from Pico cheese an artisanal cow’s milk cheese. J. Appl. Microbiol. 2014, 116, 573–585. [Google Scholar] [CrossRef]
- Rabaoui, G.; Sánchez-Juanes, F.; Tebini, M.; Naghmouchi, K.; Bellido, J.L.M.; Ben-Mahrez, K.; Réjiba, S. Potential Probiotic Lactic Acid Bacteria with Anti-Penicillium expansum Activity from Different Species of Tunisian Edible Snails. Probiotics Antimicrob. Proteins 2022, 15, 82–106. [Google Scholar] [CrossRef]
- Rastogi, S.; Mittal, V.; Singh, A. Selection of potential probiotic bacteria from exclusively breastfed infant faeces with antagonistic activity against multidrug-resistant ESKAPE pathogens. Probiotics Antimicrob. Proteins 2021, 13, 739–750. [Google Scholar] [CrossRef]
- Monteagudo-Mera, A.; Rodríguez-Aparicio, L.; Rúa, J.; Martínez-Blanco, H.; Navasa, N.; García-Armesto, M.R.; Ferrero, M.Á. In vitro evaluation of physiological probiotic properties of different lactic acid bacteria strains of dairy and human origin. J. Funct. Foods 2012, 4, 531–541. [Google Scholar] [CrossRef]
- Le, B.; Anh, P.T.N.; Kim, J.-E.; Cheng, J.; Yang, S.H. Rice bran fermentation by lactic acid bacteria to enhance antioxidant activities and increase the ferulic acid, ρ-coumaric acid, and γ-oryzanol content. J. Appl. Biol. Chem. 2019, 62, 257–264. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Microorganism | Food in Which İt İs İsolated/Used |
|---|---|
| Lactobacillus delbrueckii subsp. bulgaricus | Dairy products |
| Ligilactobacillus acidipiscis | |
| Lactobacillus acidophilus | |
| Lactobacillus delbrueckii subsp. lactis | |
| Levilactobacillus brevis | |
| Lacticaseibacillus casei | |
| Lactiplantibacillus plantarum | |
| Loigolactobacillus coryniformis subsp. coryniformis | |
| Lactobacillus helveticus | |
| Levilactobacillus parabrevis | |
| Lacticaseibacillus paracasei subsp. paracasei | |
| Lactiplantibacillus pentosus | |
| Lactiplantibacillus plantarum subsp. plantarum | |
| Weissella hellenica | |
| Weisella paramesenteroides | |
| Weisella confusa | |
| Lactobacillus helveticus | Probiotic Dairy Products |
| Lacticaseibacillus casei | |
| Lacticaseibacillus rhamnosus | |
| Limosilactobacillus reuteri | |
| Lactiplantibacillus plantarum | Sauerkraut |
| Lactiplantibacillus plantarum | Meat, Meat Products |
| Companilactobacillus alimentarius | |
| Latilactobacillus curvatus subps. curvatus | |
| Lapidilactobacillus dextrinicus | |
| Lacticaseibacillus paracasei subsp. paracasei | |
| Lactiplantibacillus plantarum subsp. plantarum | |
| Latilactobacillus sakei subsp. sakei | |
| Weisella hellenica | |
| Weisella viridescens | |
| Weisella paramesenteroides | |
| Weisella confusa | |
| Lactobacillus acetotolerans | Vegetables |
| Levilactobacillus brevis | |
| Lactobacillus acidophilus | |
| Schleiferilactobacillus harbinensis | |
| Lactobacillus helveticus | |
| Companilactobacillus kimchii | |
| Lentilactobacillus kisonensis | |
| Levilactobacillus parabrevis | |
| Lactiplantibacillus plantarum subsp. plantarum | |
| Weisella paramesenteroides | |
| Weisella confusa | |
| Levilactobacillus acidifarinae | Sourdough |
| Lactobacillus amylolyticus | |
| Lactobacillus amylovorus | |
| Lentilactobacillus parabuchneri | |
| Lactobacillus crispatus | |
| Limosilactobacillus fermentum | |
| Companilactobacillus crustorum | |
| Lactobacillus gasseri | |
| Levilactobacillus hammesii | |
| Fructilactobacillus fructivorans | |
| Lactobacillus jensenii | |
| Lactobacillus johnsonii | |
| Lacticaseibacillus manihotivorans | |
| Companilactobacillus mindensis | |
| Limosilactobacillus mucosae | |
| Weisella cibaria | |
| Weisella confusa | |
| Ligilactobacillus acidipiscis | Fish |
| Companilactobacillus alimentarius | |
| Companilactobacillus farciminis | |
| Weisella thailandensis | |
| Lentilactobacillus parabuchneri | Wine |
| Lentilactobacillus hilgardii | |
| Liquorilactobacillus oeni | |
| Lactiplantibacillus pentosus | |
| Liquorilactobacillus cacaonum | Cocoa |
| Lactiplantibacillus fabifermentans | |
| Liquorilactobacillus ghanensis | |
| Liquorilactobacillus nagelii | |
| Secundilactobacillus collinoides | Fruits |
| Liquorilactobacillus mali | |
| Lactiplantibacillus pentosus | |
| Paucilactobacillus suebicus | |
| Agrilactobacillus composti | Beverages |
| Fructilactobacillus fructivorans | |
| Liquorilactobacillus hordei | |
| Latilactobacillus sakei subsp. sakeii | |
| Liquorilactobacillus mali | |
| Lentilactobacillus diolivorans | Cereals |
| Limosilactobacillus frumenti | |
| Companilactobacillus farciminis | Soy |
| Author and Reference | Country | Traditional Fermented Foods | Study Design | Microorganisms | Conclusion |
|---|---|---|---|---|---|
| Sadeghi et al. [81] (2022) | Iranian | Dairy products | The probiotic qualities of 144 different strains of LAB were investigated. | -Lacticaseibacillus paracasei S23 -Lactiplantibacillus plantarum S57 -Lactiplantibacillus plantarum S70 -Lacticaseibacillus casei S81 | These four acidophilic–aciduric LAB show great probiotic potential and antimicrobial activity with their high autoaggregation, coaggregation and hydrophobicity, and high biofilm formation capacity. |
| Khan et al. [82] (2016) | Korea | Kimchi | The functional properties of Lactiplantibacillus plantarum DGK-17 isolated from Kimchi were investigated. | -Lactiplantibacillus plantarum DGK-17 | This acidophilic microorganism has a strong probiotic potential and antimicrobial activity. |
| Bin Masalam et al. [6] (2018) | Saudi Arabia | Milk | Thirteen distinct types of raw and fermented milk were used to isolate 93 possible LAB. | -Lacticaseibacillus casei MSJ1 -Lacticaseibacillus casei Dwan5 -Lactobacillus plantarum EyLan2 -Enterococcus faecium Gail-BawZir8 | In terms of tolerance to acidic pH, bile resistance, and antibacterial activity, it was determined that these four LAB exhibited the best probiotic properties. |
| Sagdic et al. [83] (2014) | Turkey | Gilaburu | The probiotic potentials of LAB isolated from traditional Turkish fermented European cranberrybush (Viburnum opulus L.; Turkish name is gilaburu) fruit juice were determined. | -Lacticaseibacillus casei G20a -Lactiplantibacillus plantarum G19e | Lacticaseibacillus casei (G20a) and Lactiplantibacillus plantarum (G19e) were identified as the strains with the highest cell hydrophobicity degrees. Lactiplantibacillus plantarum strains were found to be more tolerant to acidic pH than other strains and could grow at pH 2.5. It was concluded that, in addition to the Lactobacillusplantarum strain, which is dominant in fermented gilaburu juice, 11 different identified LAB strains can be used as probiotic bacteria. |
| Margalho et al. [11] (2021) | Brazil | Artisanal cheeses | The probiotic potential of LAB strains (n = 220) isolated from Brazilian artisanal cheeses was investigated. | -Lactiplantibacillus plantarum -19 more acidophilic LAB | Twenty acidophilic isolates were evaluated as probiotics because they met these criteria according to their low pH resistance, bile salts, GI tolerance and adhesion properties. Among them, Lactiplantibacillus plantarum significantly reduced the number of Staphylococcus aureus and Listeria monocytogenes pathogens. |
| Metrouh et al. [84] (2022) | Algeria | Traditional cheeses “Jben” | The probiotic potential of Lactiplantibacillus plantarum SJ14 isolated from Algerian traditional cheeses (Jben) was evaluated. | -Lactiplantibacillus plantarum SJ14 | This low-pH microorganism showed desirable probiotic characteristics. |
| Coimbra-Gomes et al. [85] (2022) | Portugal | Cobrançosa Table Olives | Nineteen native LAB strains isolated from Cobrançosa table olives were studied. | -Lactiplantibacillus paraplantarum i101 -Lactiplantibacillus pentosus i53 -Lactiplantibacillus pentosus i106 | This acidophilic strain showed the highest survival rate. |
| Liu et al. [86] (2022) | China | Fermented Vegetables | The Lactobacillus strains that were obtained from traditionally fermented vegetables were put through an in vitro probiotic test. | -Lactiplantibacillus plantarum -Levilactobacillus brevis -Weissella viridescens | Out of the 74 strains that were isolated from fermented vegetables, 26 demonstrated high survival rates in the presence of bile salts and a low pH. A total of 15 of these strains are Lactiplantibacillus plantarum strains, 9 are Levilactobacillus brevis strains, and the final two are Weissella viridescens strains. |
| Akmal et al. [87] (2022) | Pakistan | Artisanal Fermented Pickles | Nine Lactobacillus strains isolates with promising probiotic potential from fifty different traditional fermented pickle samples were investigated. | -Lactiplantibacillus plantarum -Lacticaseibacillus paracasei -Levilactobacillus brevis | It was determined that three of the strains with probiotic potential belonged to Lactiplantibacillus plantarum, five to Lacticaseibacillus paracasei, and one to Levilactobacillus brevis. |
| Leite et al. [88] (2015) | Brazil | Kefir grains | In this study, the potential probiotic qualities of 34 LAB were investigated after being isolated from a variety of Brazilian kefir grains. | -Lacticaseibacillus paracasei MRS59 | This acidophilic strain has been recognized as a promising probiotic candidate. |
| Lee et al. [66] (2016) | Korea | Kimchi | The probiotic potential of four LAB strains isolated from kimchi was investigated. | -Lactiplantibacillus plantarum C182 -Three Leuconostoc strains | Leuconostoc mesenteroides F27 and Lactiplantibacillus plantarum C182 strains can be used as potential probiotics. |
| Olajugbagbe et al. [89] (2020) | Nigeria | Wara | The probiotic potential of Pediococcus acidilactici isolated from Wara, a Nigerian unripened soft cheese from cow milk, was studied. | -Pediococcus acidilactici | It was concluded that this strain, which was demonstrated to survive at pH 2 and 1.5% bile salt concentration, and to have a high auto-aggregation ability and hydrophobicity, might be a useful probiotic for the development of functional food products. |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Icer, M.A.; Özbay, S.; Ağagündüz, D.; Kelle, B.; Bartkiene, E.; Rocha, J.M.F.; Ozogul, F. The Impacts of Acidophilic Lactic Acid Bacteria on Food and Human Health: A Review of the Current Knowledge. Foods 2023, 12, 2965. https://doi.org/10.3390/foods12152965
Icer MA, Özbay S, Ağagündüz D, Kelle B, Bartkiene E, Rocha JMF, Ozogul F. The Impacts of Acidophilic Lactic Acid Bacteria on Food and Human Health: A Review of the Current Knowledge. Foods. 2023; 12(15):2965. https://doi.org/10.3390/foods12152965
Chicago/Turabian StyleIcer, Mehmet Arif, Sena Özbay, Duygu Ağagündüz, Bayram Kelle, Elena Bartkiene, João Miguel F. Rocha, and Fatih Ozogul. 2023. "The Impacts of Acidophilic Lactic Acid Bacteria on Food and Human Health: A Review of the Current Knowledge" Foods 12, no. 15: 2965. https://doi.org/10.3390/foods12152965
APA StyleIcer, M. A., Özbay, S., Ağagündüz, D., Kelle, B., Bartkiene, E., Rocha, J. M. F., & Ozogul, F. (2023). The Impacts of Acidophilic Lactic Acid Bacteria on Food and Human Health: A Review of the Current Knowledge. Foods, 12(15), 2965. https://doi.org/10.3390/foods12152965