

Functional and Nutritional Characteristics of Natural or Modified Wheat Bran Non-Starch Polysaccharides: A Literature Review



Abstract
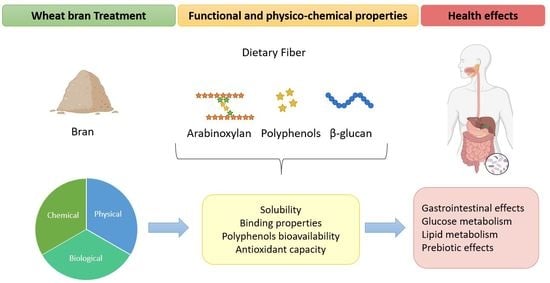
1. Introduction

2. Wheat Bran Characteristics and Properties
2.1. Chemical Composition of Wheat Bran
2.1.1. Wheat Bran Fibre Carbohydrates

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Molecular Group | Most Used Method in Literature | Values in Total WB | Values in WB SDF | Values in WB IDF | References |
|---|---|---|---|---|---|
| DF (g/100 g WB DW) | Gravimetric-enzymatic (AOAC methods (991.43; 985.29), AACC methods (32-07.01; 32-05.01; 32-21.01; 32-06.01)) | 33.4–62.4 | 2.3–9.8 | 38.5–60.1 | [28,37,56,57,58,59,60] |
| Total starch (g/100 g DW) | Colourimetric methods (AOAC 996.11, AACC method 76–12) | 11.3–23.5 | X | 7.6 | [37,57,58,59,60,61,62,63] |
| Nitrogen (g proteins/100 g DW) | Kjeldahl method (AACC 46-10) combustion methods (Dumas ICC standard 167) | 14.5–20.9 | X | 9.9 | [59,60,63,64,65,66] |
| Simple sugars (g/100 g WB DW) | Chromatography (HPLC-ELSD, HPAEC) | 0.14–0.63 | X | X | [67,68] |
| Ash (g ash/100 g DW) | Gravimetric (AACC-08-01) | 4.4–6.4 | X | 3.9 | [58,62,63] |
| Cellulose (% DW) | 5.5–31.1 | X | X | [41,42,69] | |
| Lignin content | Gravimetric (Klason) | 8–15 | X | X | [41,42,54] |
| Total β-glucans | AACC 32-23.01 AOAC 995.16 | 2.1–2.3 | 1.9 | 2.3 | [62,70] |
| Total arabinoxylans (% of DW) | Colourimetry (phloroglucinol), Chromatography (GC-MS) | 5.0–26.9 | X | X | [36,37,38] |
| Physico-Chemical Property | Methods | Value in Wheat Bran | References |
|---|---|---|---|
| WEAX (g/g DW) | Chromatography | 0.65 | [70] |
| Molecular Bounds | Spectroscopy FTIR | 3300 cm−1: O-H (cellulose, hemicellulose) 2930 cm−1 C-H CH2 (polysaccharides) 1660 cm−1 (lignin) | [63,71,72,73] |
| Ratio Arabinose/Xylose | Chromatography (HPLC-ELSD, HPAEC-PAD) | 0.2–1.1 | [74,75,76,77] |
| AX Solubility (%) | Gravimetric (extractions) | 15 | |
| AX Molecular weight (kDa) | Asymmetrical Flow Field-Flow Fractionation (AFFFF) | 20–600 | [54,75,78] |
| β-glucans Mw (kDa) | AFFFF | 258–635 | [49,51,79] |
| β-glucans Polymerisation degree (DP) | Enzymatic and Chromatographic (HPLC) | 5–28 |
2.1.2. Wheat Bran Fibre-Associated Phenolic Compounds
| Polyphenol | Quantity µg/g of Wheat Bran DW | References |
|---|---|---|
| Polyphenol | ||
| Free Phenolic Compounds | ||
| Phenolic acid—Hydroxybenzoic acids | ||
| Gallic acid | 0.3–1.0 | [44,58] |
| Vanillic | 2.2–28.5 | [58,95,96] |
| Vanillic acid isomer | 8.1–15.5 | [97] |
| 4-hydroxybenzoic = p-hydroxybenzoic acid | 2.5–5.8 | [58,96] |
| Protocathechuic | Nd–8.9 | [44,96] |
| Syringic | 1–8.0 | [44,95,96] |
| Salicylic | 6.4 | [96] |
| Ellagic acid | [98] | |
| Phenolic acid—Hydroxycinnamic acids | ||
| 3.4- Dimethoxycinnamic | 1.6 | [96] |
| Caffeic acid | 0.1–1.6 | [44,58,96] |
| Trans-caffeic | 2.5–8.2 | [97] |
| Chlorogenic acid | 1.9–3.1 | [44,58] |
| p-coumaric | 10.6–50.2 | [58,96] |
| Trans-p-coumaric | 1.1–2.3 | [97] |
| Ferulic | Free: 0.2–19Total: 1375–5670 | [44,58,95,99] |
| Trans-ferulic | 8.4–20.2 | [96] |
| Cis-ferulic | Nd–0.7 | [96] |
| Sinapic | 1.6–5.8 | [44,58,96] |
| Phenolic acid—Phenylethanoid | ||
| Hydroxytyrosol | 5.6–12.4 | [97] |
| Flavonoids | ||
| Catechin | 3.5–50.1 | [44] |
| Epicatechin | 1.1–3.3 | [44] |
| Apigenin-6-C-arabinose-8-C-hexoside | 101–149 | [97] |
| Apigenin-6-C-β-galactosyl-8-C-β-glucosyl-O-glucopyranoside | 37.9–49.3 | [97] |
| Apigenin-6,8-di-C-glucoside | 3.4–6.5 | [97] |
| Anthocyanins | ||
| Malvidin | [98] | |
| Total contents | ||
| Total phenolics (FC) GAE | 1467–11,100 | [30,60,69] |
| Bound polyphenols GAE | 2451–8500 | [60] |
| Free polyphenols GAE | 1175–2600 | [60] |
2.2. WB Physico-Chemical Properties
2.2.1. Fibre Functional Properties
Wheat Bran Fibre Solubility
Wheat Bran Viscosity
Binding Capacities
Water Holding Capacity and Water Swelling Capacity
Oil-Binding Capacity
Glucose Adsorption Capacity
Cholesterol Adsorption Capacity
Sodium Cholate Adsorption Capacity
Cation Exchange Capacity
Nitrite Ion Adsorption Capacity
| Functional Property | Value in WB DW | Value in WB SDF | Value in WB IDF | References |
|---|---|---|---|---|
| Water holding capacity (g water/g DW) | 2.2–7.3 | 2.2 | 3.0–4.3 | [37,56,59,60,62,63,70,71,73,108,130,142,151,152,153,154] |
| WSC (mL/g) | 1.7–2.7 | [59,142] | ||
| Oil holding/retention capacity (g oil/g DW) | 2.3–2.5 | 0.3–1.5 | 1.5–3.5 | [59,63,73,86,154] |
| Oil binding capacity (mL oil/g DW) | 1.2–5 | [130,155] | ||
| Cholesterol adsorption capacity (mg/g) | 2.2 (pH 2) | 20.5–32.9 (pH 2) 3.5–5.3 (pH 7) | 14.5–18.5 (pH 2) 18.1 (pH 7) | [56,63,71,73,130,151,156] |
| Glucose adsorption capacity (mmol/g) | 2.3 (50 MM) | 2.0 (50 MM) | [63,71,154] | |
| 2.1–7.3 (100 MM) | 4–5 (100 MM) | |||
| 15.7 (200 MM) | 8.0 (200 MM) | |||
| Sodium cholate adsorption capacity (mg/g) | 60.6–67.5 | 3.2 | 10–37 | [63,73,130,142,151] |
| Cation exchange capacity (mmol/g) | 0.51 | 0.122–0.132 | [63,73,154] | |
| Nitrite ion adsorption capacity (µmol/g) | 37 (pH 2) 15 (pH 7) | [73] | ||
| Viscosity (mPa·s) | 1.27–1.33 | [62,112] | ||
| Viscosity (CentiPoise) | 570 | [157] |
2.2.2. Physico-Chemical Properties of Fibre-Associated Phenolic Compounds
Antioxidant Activities
| Antioxidant Capacity Value | Wheat Bran Sample | References |
|---|---|---|
| DPPH | ||
| 59.42% | SDF | [71] |
| 2.91% | IDF residue | |
| 84.11% | IDF bound polyphenols | |
| 64.7% | Wheat IDF | [73] |
| 3.6 mm TEAC/g | Wheat bran (free) | [179] |
| 17.7 mm TEAC/g | Wheat bran (bound) | |
| 20.0 mm TEAC/g | Wheat bran (total) | |
| 6.42 EC50 mg/mL | Wheat bran | [167] |
| 7.02 IC50 mg/mL | Wheat bran | |
| 5.2% (discolouration) | Wheat bran (free) | [96] |
| 9.7% | Wheat bran (acid hydrolysis) | |
| 15.0% | Wheat bran (alkaline hydrolysis) | |
| 4.2–4.7 µmol TE/g DW | Wheat bran (free) | [44] |
| 7.5–8.3 µmol TE/g DW | Wheat bran (bound) | |
| 11.7–13.0 µmol TE/g DW | Wheat bran (total) | |
| 14.5% (inhibition) | Wheat bran coarse (soluble) | [175] |
| 15.9% | Wheat bran medium (soluble) | |
| 13.7% | Wheat bran fine (soluble) | |
| 41.3% | Wheat bran coarse (bound) | |
| 43.0% | Wheat bran medium (bound) | |
| 32.6% | Wheat bran fine (bound) | |
| ABTS | ||
| 88.42% | Wheat IDF | [73] |
| 10.2 mm TEAC/g | Wheat bran (free) | [179] |
| 40.5 mm TEAC/g | Wheat bran (bound) | |
| 50.7 mm TEAC/g | Wheat bran (total) | |
| 6.6 IC50 mg/mL | Wheat bran | [167] |
| 2.9 µM trolox equivalent | Wheat bran (free) | [96] |
| 6.0 µM trolox equivalent | Wheat bran (acid hydrolysis) | |
| 9.1 µM trolox equivalent | Wheat bran alkaline hydrolysis | |
| FRAP | ||
| 53.04 µmol FeSO4/g | Wheat bran (coarse) | [175] |
| 40.84 µmol FeSO4/g | Wheat bran (fine) | |
| 23.8 µmol FeSO4/g of defatted bran | Wheat bran coarse (soluble) | |
| 8.9 µmol FeSO4/g of defatted bran | Wheat bran medium (soluble) | |
| 12.3 µmol FeSO4/g of defatted bran | Wheat bran fine (soluble) | |
| 229.2 µmol FeSO4/g of defatted bran | Wheat bran coarse (bound) | |
| 28.3 µmol FeSO4/g of defatted bran | Wheat bran medium (bound) | |
| 28.6 µmol FeSO4/g of defatted bran | Wheat bran fine (bound) | |
| 53 nmol TE/g grain | Wheat bran | [167] |
| 58.4 µmol/g | Soluble AX from wheat bran | [180] |
| 21.42 µmol TE/g | SDF | [139] |
| 0.36 mmol/L | SDF | [71] |
| 0.09 mmol/L | IDF residue | |
| 1.59 mmol/L | IDF bound polyphenols | |
| 11.0 mm TEAC/g | Wheat bran (free) | [179] |
| 34.5 mm TEAC/g | Wheat bran (bound) | |
| 48.9 mm TEAC/g | Wheat bran (total) | |
3. Heath Benefits of Wheat Bran
3.1. General Effects Linked with WB Functional Properties
3.2. Effects on Glucose Metabolism
3.3. Effects on Lipid Metabolism
3.4. Effects on Microbiota
3.5. Immunomodulatory Activity
3.6. Antiproliferative Activity on Cancer
3.7. Phenolic Compounds Effects on Health
| Type of Fibre, Quantity | Clinical Study Duration | Volunteers (Disease, Number, Sex) | Results Linked with Fibre Consumption | References |
|---|---|---|---|---|
| High and low Mw concentrated barley β-glucan extract 3 g and 5 g doses | 10 weeks | Hypercholesterolaemia 155 M/F | ↘ LDL-Cholesterol ↘ Total cholesterol = HDL Cholesterol levels | [201] |
| High and low Mw concentrated barley β-glucan 6 g/day | 6 weeks | Hypercholesterolemia 90 M/F | Low Mw group ↘ Ratio TC/HDL ↗ Body weight ↘ Hunger High Mw group ↗ Ratio TC/HDL ↘ Body weight ↘ Hunger = Blood pressure, glucose, insulin, gastrointestinal symptoms | [252] |
| 5 groups: A: 3 g/day wheat fibre B: 3 g/day oat high Mw β-glucan C: 4 g/day oat medium Mw β-glucan D: 3 g/day oat medium Mw β-glucan E: 4 g/day oat low Mw β-glucan | 4 weeks | 5 mmol/L > LDL-C > 3 mmol/L 345 M/F | LDL-C in groups B, C and D < group A Non-significant for sex, age, baseline LDL-C | [202] |
| 3 g barley high Mw β-glucan 5 g barley low Mw β-glucan 3 g barley Low Mw β-glucan | 5 weeks | Mildly hypercholesterolemic subjects 30 | ↗ Lithocholic acid excretion ↗ Fermentability of fibre ↗ Total faecal SCFA in group 3 g/d high Mw | [203] |
| Control 3 g/d reduced viscosity barley β-glucan 6 g/d reduced viscosity barley β-glucan | 12 weeks | Healthy subjects 50 | ↗ Insulin sensitivity for 6 g/d = Body weight | [191] |
| Control: microcrystalline cellulose Oat β-glucan (5 g/day) | 12 weeks | Type-2 diabetes patients 37 | ↘ HbA1c ↘ Insulin and GLP-1 ↘ C-peptide ↘ HOMA ↘ Lactobacillus spp. and Butyrate-producing bacteria | [187] |
| Control group: Placebo 15 g AX | 6 weeks | Impaired glucose tolerance, insulin resistant, slightly elevated LDL and low HDL cholesterol concentration. Body mass index (BMI): 30.1 kg/m2 7 women 4 males | ↘ Postprandial response in serum glucose, insulin, triglycerides, plasma total ghrelin. = Plasma acylated ghrelin | [193] |
| 7.5 g/day AX (n = 16) 15 g/d AX (n = 17) 15 g/d placebo (control n = 14) | 6 weeks | Overweight 47 | = Gastrointestinal permeability and tight junction ↘ Faecal microbiota diversity ↗ Faecal pH ↗ Faecal concentration total SCFA ↘ TNFα by simulated PBMCs | [205] |
| 10 g placebo (control) 10 g wheat AXOS | 3 weeks | Healthy subjects 20 | = Blood lipid = Gastrointestinal symptoms ↗ Flatulence ↗ Bifidobacteria ↘ Urinary p-cresol (after 2 weeks) | [206] |
| 0, 3 and 10 g/day wheat bran extract enriched in AXOS | 3 weeks | Healthy volunteers 63 | ↘ Urinary p-cresol (10 g/d) ↗ Faecal bifidobacteria (10 g/d) ↗ Faecal SCFA ↗ Flatulence frequency distress severity (10 g/d) ↘ Faecal pH ↘ Constipation occurrence frequency | [207] |
| 10.4 g/d AXOS from wheat bran extracts and 3.6 g/d polyunsaturated fatty acids | 12 weeks | Overweight individuals with indices of metabolic syndrome 55 | ↗ Bifidobacteria ↗ Butyrate producing bacteria | [208] |
| Trial 1: Ready to eat cereal incorporated with fibres Low-fibre (4 g) AXOS or AX from flax (FLAX) High fibre (15 g) AXOS or AX from flax (FLAX) Trial 2: Low fibre High fibre AXOS High fibre FLAX Low fibre isocaloric | Overweight Women BMI 25.0–29.9 kg/m2 | = postprandial appetite ↗ GLP-1 ↗ PYY (High fibre AXOS and High fibre FLAX vs. Low fibre isocaloric) | [196] |
3.8. Nutritional and Health Claims of Wheat Bran
4. Improving Wheat Bran Characteristics for Increasing Health Benefits
4.1. Biological Treatments
4.2. Physical Treatments
| Treatment Type | Objective | Effects of Particle Size Reduction | References |
|---|---|---|---|
| Milling | To study if particle size distribution of whole wheat bran affects the phytochemical extractability and antioxidant properties. | ↗ Phenolic acids, flavonoid, carotenoids, anthocyanins ↗ ORAC (+80%) ↘ DPPH, FRAP, TAOC | [175] |
| Milling | To understand wheat bran’s hydration properties. | No chemical modification ↗ WEAX ↗ Damaged starch ↘ WHC | [152] |
| Milling | To study the effect of wheat bran particle size on in vitro gastrointestinal digestion. | ↗ Release of Phenolics compounds ↗ Antioxidant capacities ↗ Carbohydrate-digestive enzymes inhibitory activities ↘ Starch digestibility (retard starch digestion) | [58] |
| Air-flow impact mill | To study the link between structure and hydration properties of milled wheat bran. | ↗ SDF ↘ WRC, WSC, ORC | [86] |
| Micronisation = superfine grinding | To investigate the effect of wheat bran micronisation on its functionality and dough properties. | ↗ SDF ↗ WEAX ↗ Damage starch ↗ Phenolic acids liberation ↗ Free ferulic acid ↗ TPC, ABTS, DPPH ↗ Aleurone cells disruption ↘ Dough extensibility ↘ IDF ↘ WRC | [271] |
| Microfluidisation | ↗ WEAX ↗ Extractible B-glucans ↗ Viscosity ↗ Water extractability ↗ WRC ↗ Phytate (aleurone breaking) ↗ Free glucose content ↗ Maltose (starch degradation) ↗ Water extractible proteins | [112] | |
| Microfluidisation | Improve bran’s physiological properties | ↗ WHC, WSC ↗ OHC ↗ CEC ↘ Bulk density | [277] |
| Microfluidisation | Improving antioxidant properties of bound phenolic compounds. | ↗ Surface reactive (×3.8) and bound phenolics ↗ Antioxidant capacities | [278] |
| Extrusion | ↗ Fibre solubility ↘ Total dietary fibre ↘ WBC ↘ Phytic acid content | [56] | |
| Extrusion | ↘ Phytic acid (64.4%) Ture 115 °C 20% moisture ↘ Polyphenols ↘ Oxalates (36.8%, 140 °C, 20% moisture) ↘ Trypsin inhibitors (71.2%, 140 °C, 20% moisture) ↗ Bulk density ↗ Redness ↘ Yellowness | [279] | |
| Extrusion | ↗ Soluble fibre (70–100% RPM max) | [280] | |
| Extrusion | ↗ SDF ↗ WEAX (+33.6%) ↗ WHC, WSC ↗ Total phenolic ↗ FA ↗ Folic acid ↗ ORAC ↘ OHC ↘ Vitamin B1/B2 ↘ Phytic acid ↘ Fat, starch, protein | [25] | |
| Blasting-extrusion | ↗ SDF (+70%) ↗ WRC, WSC ↗ OBC ↗ Water soluble polysaccharides ↗ Antioxidant | [28] | |
| CO2 Blasting-extrusion | ↗ SDF ↗ WRC, WSC ↗ ORC | [59] | |
| Steam explosion (215 °C 120 s) | ↗ Free soluble phenolic acids | [281] | |
| Steam explosion | ↗ WEAX ↗ Reducing sugar ↘ Baking quality ↘ Phytic acid | [276] | |
| Steam explosion (high-pressure steam 0.8 MPa, 5 min) | Improve the value of wheat bran to provide a reference for the development of WB treatment | ↘ Particle size ↗ WHC, WSC (↘ WHC with particle size reduction) ↘ SCAC (and with particle size reduction) ↘ CEC ↘ Phytate content | [142] |
4.3. Chemical Treatments
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- FAO. World Food Situation. Available online: https://www.fao.org/worldfoodsituation/csdb/en/ (accessed on 29 January 2023).
- US Wheat Associates. Human Wheat Consumption Sets New Record. Available online: https://www.uswheat.org/wheatletter/human-wheat-consumption-sets-new-record/ (accessed on 29 January 2023).
- Hipsley, E.H. Dietary “Fibre” and Pregnancy Toxaemia. Br. Med. J. 1953, 2, 420–422. [Google Scholar] [CrossRef]
- Smith, C.K. Guidelines on Nutrition Labelling; Food and Agriculture Organization: Rome, Italy, 2013; Available online: https://www.fao.org/home/en (accessed on 14 June 2023).
- EFSA Panel on Dietetic Products. Scientific Opinion on the Substantiation of Health Claims Related to Resistant Maltodextrin and Reduction of Post-Prandial Glycaemic Responses (ID 796), Maintenance of Normal Blood LDL-Cholesterol Concentrations (ID 2927), Maintenance of Normal (Fasting) Blood Concentrations of Triglycerides (ID 2927) and Changes in Bowel Function (ID 797) Pursuant to Article 13(1) of Regulation (EC) No 1924/2006. EFSA J. 2011, 9, 2070. [Google Scholar] [CrossRef]
- Food and Drug Administration. Food labeling: Revision of the nutrition and supplement facts labels. Final rule. US Fed. Regist. 2016, 81, 33741–33999. [Google Scholar]
- Bureau of Nutritional Sciences; Health Canada. Policy for Labelling and Advertising of Dietary Fibre-Containing Food Products. 2012. Available online: https://www.canada.ca/content/dam/hc-sc/migration/hc-sc/fn-an/alt_formats/pdf/legislation/pol/fibre-label-etiquetage-eng.pdf (accessed on 27 June 2023).
- Australian Government. Australia New Zealand Food Standards Code—Standard 1.2.7—Nutrition, Health and Related Claims; Attorney-General’s Department: Australian Capital Territory, Australia, 2016. [Google Scholar]
- Health Canada; Bureau of Nutritional Sciences; Food Directorate; Health Products and Food Branch. Policy for Labelling and Advertising of Dietary Fibre-Containing Food Products; Policy for Labelling and Advertising of Dietary Fibre-Containing Food Products: Ottawa, Canada, 2017. [Google Scholar]
- Stephen, A.M.; Champ, M.M.-J.; Cloran, S.J.; Fleith, M.; van Lieshout, L.; Mejborn, H.; Burley, V.J. Dietary Fibre in Europe: Current State of Knowledge on Definitions, Sources, Recommendations, Intakes and Relationships to Health. Nutr. Res. Rev. 2017, 30, 149–190. [Google Scholar] [CrossRef] [PubMed]
- Marcotuli, I.; Colasuonno, P.; Hsieh, Y.S.Y.; Fincher, G.B.; Gadaleta, A. Non-Starch Polysaccharides in Durum Wheat: A Review. Int. J. Mol. Sci. 2020, 21, 2933. [Google Scholar] [CrossRef] [PubMed]
- Kurek, M.; Wyrwisz, J. The Application of Dietary Fiber in Bread Products. J. Food Process. Technol. 2015, 6, 447–450. [Google Scholar] [CrossRef]
- Anderson, J.W.; Baird, P.; Davis, R.H., Jr.; Ferreri, S.; Knudtson, M.; Koraym, A.; Waters, V.; Williams, C.L. Health Benefits of Dietary Fiber. Nutr. Rev. 2009, 67, 188–205. [Google Scholar] [CrossRef]
- Onipe, O.O.; Jideani, A.I.O.; Beswa, D. Composition and Functionality of Wheat Bran and Its Application in Some Cereal Food Products. Int. J. Food Sci. Technol. 2015, 50, 2509–2518. [Google Scholar] [CrossRef]
- Benisi-Kohansal, S.; Saneei, P.; Salehi-Marzijarani, M.; Larijani, B.; Esmaillzadeh, A. Whole-Grain Intake and Mortality from All Causes, Cardiovascular Disease, and Cancer: A Systematic Review and Dose-Response Meta-Analysis of Prospective Cohort Studies. Adv. Nutr. 2016, 7, 1052–1065. [Google Scholar] [CrossRef]
- Della Pepa, G.; Vetrani, C.; Vitale, M.; Riccardi, G. Wholegrain Intake and Risk of Type 2 Diabetes: Evidence from Epidemiological and Intervention Studies. Nutrients 2018, 10, 1288. [Google Scholar] [CrossRef] [PubMed]
- Chung, S.; Hwang, J.-T.; Park, S.-H. Physiological Effects of Bioactive Compounds Derived from Whole Grains on Cardiovascular and Metabolic Diseases. Appl. Sci. 2022, 12, 658. [Google Scholar] [CrossRef]
- Borczak, B.; Sikora, M.; Sikora, E.; Dobosz, A.; Kapusta-Duch, J. Glycaemic Index of Wheat Bread. Starch–Stärke 2018, 70, 1700022. [Google Scholar] [CrossRef]
- Izydorczyk, M.S.; Dexter, J.E. Barley β-Glucans and Arabinoxylans: Molecular Structure, Physicochemical Properties, and Uses in Food Products–a Review. Food Res. Int. 2008, 41, 850–868. [Google Scholar] [CrossRef]
- Korczak, R.; Slavin, J.L. Definitions, Regulations, and New Frontiers for Dietary Fiber and Whole Grains. Nutr. Rev. 2020, 78, 6–12. [Google Scholar] [CrossRef]
- Cheng, W.; Sun, Y.; Fan, M.; Li, Y.; Wang, L.; Qian, H. Wheat Bran, as the Resource of Dietary Fiber: A Review. Crit. Rev. Food Sci. Nutr. 2022, 62, 7269–7281. [Google Scholar] [CrossRef]
- Mendis, M.; Simsek, S. Arabinoxylans and Human Health. Food Hydrocoll. 2014, 42, 239–243. [Google Scholar] [CrossRef]
- Izydorczyk, M.S. Functional Properties of Cereal Cell Wall Polysaccharides. In Carbohydrates in Food; CRC Press: Boca Raton, FL, USA, 2017; ISBN 978-1-315-37282-2. [Google Scholar]
- Gan, J.; Xie, L.; Peng, G.; Xie, J.; Chen, Y.; Yu, Q. Systematic Review on Modification Methods of Dietary Fiber. Food Hydrocoll. 2021, 119, 106872. [Google Scholar] [CrossRef]
- Ye, G.; Wu, Y.; Wang, L.; Tan, B.; Shen, W.; Li, X.; Liu, Y.; Tian, X.; Zhang, D. Comparison of Six Modification Methods on the Chemical Composition, Functional Properties and Antioxidant Capacity of Wheat Bran. LWT 2021, 149, 111996. [Google Scholar] [CrossRef]
- Finnie, S.; Atwell, W.A. Wheat Flour; Handbook series; Elsevier: Amsterdam, The Netherlands, 2016; pp. 1–16. ISBN 978-1-891127-90-8. [Google Scholar]
- Prosky, L.; Asp, N.G.; Schweizer, T.F.; DeVries, J.W.; Furda, I. Determination of Insoluble, Soluble, and Total Dietary Fiber in Foods and Food Products: Interlaboratory Study. J. Assoc. Off. Anal. Chem. 1988, 71, 1017–1023. [Google Scholar] [CrossRef]
- Yan, X.; Ye, R.; Chen, Y. Blasting Extrusion Processing: The Increase of Soluble Dietary Fiber Content and Extraction of Soluble-Fiber Polysaccharides from Wheat Bran. Food Chem. 2015, 180, 106–115. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.; Liu, R.; Wu, T.; Zhang, M. Aggregation and Rheological Behavior of Soluble Dietary Fibers from Wheat Bran. Food Res. Int. 2017, 102, 291–302. [Google Scholar] [CrossRef]
- Murlidhar Sonkar, R.; Savata Gade, P.; Bokade, V.; Mudliar, S.N.; Bhatt, P. Ozone Assisted Autohydrolysis of Wheat Bran Enhances Xylooligosaccharide Production with Low Generation of Inhibitor Compounds: A Comparative Study. Bioresour. Technol. 2021, 338, 125559. [Google Scholar] [CrossRef] [PubMed]
- Kulathunga, J.; Simsek, S. Dietary Fiber Variation in Ancient and Modern Wheat Species: Einkorn, Emmer, Spelt and Hard Red Spring Wheat. J. Cereal Sci. 2022, 104, 103420. [Google Scholar] [CrossRef]
- Merali, Z.; Collins, S.R.A.; Elliston, A.; Wilson, D.R.; Käsper, A.; Waldron, K.W. Characterization of Cell Wall Components of Wheat Bran Following Hydrothermal Pretreatment and Fractionation. Biotechnol. Biofuels 2015, 8, 23. [Google Scholar] [CrossRef] [PubMed]
- Kapreliants, L.; Zhurlova, O. Technology of Wheat and Rye Bran Biotransformation into Functional Ingredients. Int. Food Res. J. 2017, 24, 1975–1979. [Google Scholar]
- Izydorczyk, M.; Biliaderis, C.G.; Bushuk, W. Physical Properties of Water-Soluble Pentosans from Different Wheat Varieties. Cereal Chem. 1991, 68, 145–150. [Google Scholar]
- Weightman, R.M.; Davis-Knight, H.; Wang, R.H.; Misailidis, N.; Villanueva, A.L.; Campbell, G.M. Towards Defining Optimal Feedstocks for a Wheat Biorefinery: Co-Production of Arabinoxylans with Bioethanol. Asp. Appl. Biol. 2008, 90, 153–160. [Google Scholar]
- Douglas, S. A Rapid Method for the Determination of Pentosans in Wheat Flour. Food Chem. 1981, 7, 139–145. [Google Scholar] [CrossRef]
- Santala, O.; Kiran, A.; Sozer, N.; Poutanen, K.; Nordlund, E. Enzymatic Modification and Particle Size Reduction of Wheat Bran Improves the Mechanical Properties and Structure of Bran-Enriched Expanded Extrudates. J. Cereal Sci. 2014, 60, 448–456. [Google Scholar] [CrossRef]
- Barron, C.; Bar-L’Helgouac’h, C.; Champ, M.; Saulnier, L. Arabinoxylan Content and Grain Tissue Distribution Are Good Predictors of the Dietary Fibre Content and Their Nutritional Properties in Wheat Products. Food Chem. 2020, 328, 127111. [Google Scholar] [CrossRef]
- Gebruers, K.; Dornez, E.; Bedõ, Z.; Rakszegi, M.; Frás, A.; Boros, D.; Courtin, C.M.; Delcour, J.A. Environment and Genotype Effects on the Content of Dietary Fiber and Its Components in Wheat in the HEALTHGRAIN Diversity Screen. J. Agric. Food Chem. 2010, 58, 9353–9361. [Google Scholar] [CrossRef]
- Scheller, H.V.; Ulvskov, P. Hemicelluloses. Annu. Rev. Plant Biol. 2010, 61, 263–289. [Google Scholar] [CrossRef]
- Xiao, Y.; Liu, Y.; Wang, X.; Li, M.; Lei, H.; Xu, H. Cellulose Nanocrystals Prepared from Wheat Bran: Characterization and Cytotoxicity Assessment. Int. J. Biol. Macromol. 2019, 140, 225–233. [Google Scholar] [CrossRef]
- Arzami, A.N.; Ho, T.M.; Mikkonen, K.S. Valorization of Cereal By-Product Hemicelluloses: Fractionation and Purity Considerations. Food Res. Int. 2022, 151, 110818. [Google Scholar] [CrossRef]
- Si, D.; Shang, T.; Liu, X.; Zheng, Z.; Hu, Q.; Hu, C.; Zhang, R. Production and Characterization of Functional Wheat Bran Hydrolysate Rich in Reducing Sugars, Xylooligosaccharides and Phenolic Acids. Biotechnol. Rep. 2020, 27, e00511. [Google Scholar] [CrossRef] [PubMed]
- Yadav, M.P.; Kaur, A.; Singh, B.; Simon, S.; Kaur, N.; Powell, M.; Sarker, M. Extraction and Characterization of Lipids and Phenolic Compounds from the Brans of Different Wheat Varieties. Food Hydrocoll. 2021, 117, 106734. [Google Scholar] [CrossRef]
- Delcour, J.A.; Van Win, H.; Grobet, P.J. Distribution and Structural Variation of Arabinoxylans in Common Wheat Mill Streams. J. Agric. Food Chem. 1999, 47, 271–275. [Google Scholar] [CrossRef]
- de Oliveira, D.M.; Finger-Teixeira, A.; Rodrigues Mota, T.; Salvador, V.H.; Moreira-Vilar, F.C.; Correa Molinari, H.B.; Craig Mitchell, R.A.; Marchiosi, R.; Ferrarese-Filho, O.; Dantas dos Santos, W. Ferulic Acid: A Key Component in Grass Lignocellulose Recalcitrance to Hydrolysis. Plant Biotechnol. J. 2015, 13, 1224–1232. [Google Scholar] [CrossRef] [PubMed]
- Kamal-Eldin, A.; Lærke, H.N.; Knudsen, K.-E.B.; Lampi, A.-M.; Piironen, V.; Adlercreutz, H.; Katina, K.; Poutanen, K.; Man, P. Physical, Microscopic and Chemical Characterisation of Industrial Rye and Wheat Brans from the Nordic Countries. Food Nutr. Res. 2009, 53, 1912. [Google Scholar] [CrossRef] [PubMed]
- Van Craeyveld, V.; Holopainen, U.; Selinheimo, E.; Poutanen, K.; Delcour, J.A.; Courtin, C.M. Extensive Dry Ball Milling of Wheat and Rye Bran Leads to in Situ Production of Arabinoxylan Oligosaccharides through Nanoscale Fragmentation. J. Agric. Food Chem. 2009, 57, 8467–8473. [Google Scholar] [CrossRef]
- Zhao, Q.; Hu, X.; Guo, Q.; Cui, S.W.; Xian, Y.; You, S.; Chen, X.; Xu, C.; Gao, X. Physicochemical Properties and Regulatory Effects on Db/Db Diabetic Mice of β-Glucans Extracted from Oat, Wheat and Barley. Food Hydrocoll. 2014, 37, 60–68. [Google Scholar] [CrossRef]
- Lazaridou, A.; Biliaderis, C.G. Molecular Aspects of Cereal β-Glucan Functionality: Physical Properties, Technological Applications and Physiological Effects. J. Cereal Sci. 2007, 46, 101–118. [Google Scholar] [CrossRef]
- Cui, W.; Wood, P.J.; Blackwell, B.; Nikiforuk, J. Physicochemical Properties and Structural Characterization by Two-Dimensional NMR Spectroscopy of Wheat b-D-Glucan—Comparison with Other Cereal b-D-Glucans. Carbohydr. Polym. 2000, 41, 249–258. [Google Scholar] [CrossRef]
- Campbell, N.A.; Reece, J.B.; Lachaîne, R.; Faucher, J. Campbell Biologie, 9th ed.; Pearson: Montréal, QC, Canada; Toronto, ON, Canada; Paris, France, 2012; ISBN 978-2-7613-5065-5. [Google Scholar]
- Voragen, A.G.J.; Coenen, G.-J.; Verhoef, R.P.; Schols, H.A. Pectin, a Versatile Polysaccharide Present in Plant Cell Walls. Struct. Chem. 2009, 20, 263–275. [Google Scholar] [CrossRef]
- Feillet, P. Le Grain de Blé—Composition et Utilisation; INRA editions; Quae: Paris, France, 2000; ISBN 2-7380-0896-8. [Google Scholar]
- Lin, D.; Ma, Y.; Qin, W.; Loy, D.A.; Chen, H.; Zhang, Q. The Structure, Properties and Potential Probiotic Properties of Starch-Pectin Blend: A Review. Food Hydrocoll. 2022, 129, 107644. [Google Scholar] [CrossRef]
- Aktas-Akyildiz, E.; Masatcioglu, M.T.; Köksel, H. Effect of Extrusion Treatment on Enzymatic Hydrolysis of Wheat Bran. J. Cereal Sci. 2020, 93, 102941. [Google Scholar] [CrossRef]
- Dust, J.M.; Gajda, A.M.; Flickinger, E.A.; Burkhalter, T.M.; Merchen, N.R.; Fahey, G.C. Extrusion Conditions Affect Chemical Composition and in Vitro Digestion of Select Food Ingredients. J. Agric. Food Chem. 2004, 52, 2989–2996. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Li, M.; Wang, L.; Li, Z. Effect of Particle Size on the Release Behavior and Functional Properties of Wheat Bran Phenolic Compounds during in Vitro Gastrointestinal Digestion. Food Chem. 2022, 367, 130751. [Google Scholar] [CrossRef] [PubMed]
- Long, D.; Ye, F.; Zhao, G. Optimization and Characterization of Wheat Bran Modified by in Situ Enhanced CO2 Blasting Extrusion. LWT—Food Sci. Technol. 2014, 59, 605–611. [Google Scholar] [CrossRef]
- Lee, Y.Y.; Ma, F.; Byars, J.A.; Felker, F.C.; Liu, S.; Mosier, N.S.; Lee, J.H.; Kenar, J.A.; Baik, B. Influences of Hydrothermal and Pressure Treatments on Compositional and Hydration Properties of Wheat Bran and Dough Mixing Properties of Whole Wheat Meal. Cereal Chem. 2021, 98, 673–682. [Google Scholar] [CrossRef]
- Ralet, M.-C.; Thibault, J.-F.; Della Valle, G. Influence of Extrusion-Cooking on the Physico-Chemical Properties of Wheat Bran. J. Cereal Sci. 1990, 11, 249–259. [Google Scholar] [CrossRef]
- Roye, C.; Bulckaen, K.; De Bondt, Y.; Liberloo, I.; Van De Walle, D.; Dewettinck, K.; Courtin, C.M. Side-by-side Comparison of Composition and Structural Properties of Wheat, Rye, Oat, and Maize Bran and Their Impact on in Vitro Fermentability. Cereal Chem. 2020, 97, 20–33. [Google Scholar] [CrossRef]
- Wang, L.; Tian, Y.; Chen, Y.; Chen, J. Effects of Acid Treatment on the Physicochemical and Functional Properties of Wheat Bran Insoluble Dietary Fiber. Cereal Chem. 2021, 99, 343–354. [Google Scholar] [CrossRef]
- Noort, M.W.J.; van Haaster, D.; Hemery, Y.; Schols, H.A.; Hamer, R.J. The Effect of Particle Size of Wheat Bran Fractions on Bread Quality—Evidence for Fibre–Protein Interactions. J. Cereal Sci. 2010, 52, 59–64. [Google Scholar] [CrossRef]
- Müller, D.J. Dumas or Kjeldahl for Reference Analysis? FOSS: Hilleroed, Denmark, 2017. [Google Scholar]
- Call, L.; Kapeller, M.; Grausgruber, H.; Reiter, E.; Schoenlechner, R.; D’Amico, S. Effects of Species and Breeding on Wheat Protein Composition. J. Cereal Sci. 2020, 93, 102974. [Google Scholar] [CrossRef]
- van den Borne, J.J.G.C.; Kabel, M.A.; Briens, M.; van der Poel, A.F.B.; Hendriks, W.H. Effects of Pretreatment of Wheat Bran on the Quality of Protein-Rich Residue for Animal Feeding and on Monosaccharide Release for Ethanol Production. Bioresour. Technol. 2012, 124, 446–454. [Google Scholar] [CrossRef]
- Aguedo, M.; Fougnies, C.; Dermience, M.; Richel, A. Extraction by Three Processes of Arabinoxylans from Wheat Bran and Characterization of the Fractions Obtained. Carbohydr. Polym. 2014, 105, 317–324. [Google Scholar] [CrossRef]
- Žilić, S.; Serpen, A.; Akıllıoğlu, G.; Janković, M.; Gökmen, V. Distributions of Phenolic Compounds, Yellow Pigments and Oxidative Enzymes in Wheat Grains and Their Relation to Antioxidant Capacity of Bran and Debranned Flour. J. Cereal Sci. 2012, 56, 652–658. [Google Scholar] [CrossRef]
- Renzetti, S.; Theunissen, M.; Horrevorts, K. A Systematic Comparison of the Intrinsic Properties of Wheat and Oat Bran Fractions and Their Effects on Dough and Bread Properties: Elucidation of Chemical Mechanisms, Water Binding, and Steric Hindrance. Foods 2021, 10, 2311. [Google Scholar] [CrossRef]
- Chen, H.; Xiong, M.; Bai, T.; Chen, D.; Zhang, Q.; Lin, D.; Liu, Y.; Liu, A.; Huang, Z.; Qin, W. Comparative Study on the Structure, Physicochemical, and Functional Properties of Dietary Fiber Extracts from Quinoa and Wheat. LWT 2021, 149, 111816. [Google Scholar] [CrossRef]
- Chu, J.; Zhao, H.; Lu, Z.; Lu, F.; Bie, X.; Zhang, C. Improved Physicochemical and Functional Properties of Dietary Fiber from Millet Bran Fermented by Bacillus Natto. Food Chem. 2019, 294, 79–86. [Google Scholar] [CrossRef] [PubMed]
- Liao, A.-M.; Zhang, J.; Yang, Z.-L.; Huang, J.-H.; Pan, L.; Hou, Y.-C.; Li, X.-X.; Zhao, P.-H.; Dong, Y.-Q.; Hu, Z.-Y.; et al. Structural, Physicochemical, and Functional Properties of Wheat Bran Insoluble Dietary Fiber Modified With Probiotic Fermentation. Front. Nutr. 2022, 9, 803440. [Google Scholar] [CrossRef] [PubMed]
- Beaugrand, J.; Crônier, D.; Thiebeau, P.; Schreiber, L.; Debeire, P.; Chabbert, B. Structure, Chemical Composition, and Xylanase Degradation of External Layers Isolated from Developing Wheat Grain. J. Agric. Food Chem. 2004, 52, 7108–7117. [Google Scholar] [CrossRef]
- Wang, P.; Hou, C.; Zhao, X.; Tian, M.; Gu, Z.; Yang, R. Molecular Characterization of Water-Extractable Arabinoxylan from Wheat Bran and Its Effect on the Heat-Induced Polymerization of Gluten and Steamed Bread Quality. Food Hydrocoll. 2019, 87, 570–581. [Google Scholar] [CrossRef]
- Wang, J.; Bai, J.; Fan, M.; Li, T.; Li, Y.; Qian, H.; Wang, L.; Zhang, H.; Qi, X.; Rao, Z. Cereal-Derived Arabinoxylans: Structural Features and Structure–Activity Correlations. Trends Food Sci. Technol. 2020, 96, 157–165. [Google Scholar] [CrossRef]
- Kaur, A.; Yadav, M.P.; Singh, B.; Bhinder, S.; Simon, S.; Singh, N. Isolation and Characterization of Arabinoxylans from Wheat Bran and Study of Their Contribution to Wheat Flour Dough Rheology. Carbohydr. Polym. 2019, 221, 166–173. [Google Scholar] [CrossRef]
- Liu, Y.; Wang, S.; Kang, J.; Wang, N.; Xiao, M.; Li, Z.; Wang, C.; Guo, Q.; Hu, X. Arabinoxylan from Wheat Bran: Molecular Degradation and Functional Investigation. Food Hydrocoll. 2020, 107, 105914. [Google Scholar] [CrossRef]
- Gozé, P. Ozonation Des Grains de Blé Tendre: ÉTude de L’Oxydation Des Constituants Biochimiques Majeurs. Ph.D. Thesis, Sorbonne Universités—Université de Technologie de Compiègne, Compiègne, France, 2016. [Google Scholar]
- Adom, K.K.; Sorrells, M.E.; Liu, R.H. Phytochemicals and Antioxidant Activity of Milled Fractions of Different Wheat Varieties. J. Agric. Food Chem. 2005, 53, 2297–2306. [Google Scholar] [CrossRef]
- Beta, T.; Nam, S.; Dexter, J.E.; Sapirstein, H.D. Phenolic Content and Antioxidant Activity of Pearled Wheat and Roller-Milled Fractions. Cereal Chem. J. 2005, 82, 390–393. [Google Scholar] [CrossRef]
- Liyana-Pathirana, C.M.; Shahidi, F. Importance of Insoluble-Bound Phenolics to Antioxidant Properties of Wheat. J. Agric. Food Chem. 2006, 54, 1256–1264. [Google Scholar] [CrossRef] [PubMed]
- Fernandez-Orozco, R.; Li, L.; Harflett, C.; Shewry, P.R.; Ward, J.L. Effects of Environment and Genotype on Phenolic Acids in Wheat in the HEALTHGRAIN Diversity Screen. J. Agric. Food Chem. 2010, 58, 9341–9352. [Google Scholar] [CrossRef]
- Li, L.; Shewry, P.R.; Ward, J.L. Phenolic Acids in Wheat Varieties in the HEALTHGRAIN Diversity Screen. J. Agric. Food Chem. 2008, 56, 9732–9739. [Google Scholar] [CrossRef]
- Menga, V.; Fares, C.; Troccoli, A.; Cattivelli, L.; Baiano, A. Effects of Genotype, Location and Baking on the Phenolic Content and Some Antioxidant Properties of Cereal Species: Antioxidant Properties in Cereals. Int. J. Food Sci. Technol. 2009, 45, 7–16. [Google Scholar] [CrossRef]
- Li, Y.; Wang, L.; Wang, H.; Li, Z.; Qiu, J.; Wang, L. Correlation of Microstructure, Pore Characteristics and Hydration Properties of Wheat Bran Modified by Airflow Impact Mill. Innov. Food Sci. Emerg. Technol. 2022, 77, 102977. [Google Scholar] [CrossRef]
- Sosulski, F.; Krygier, K.; Hogge, L. Free, Esterified, and Insoluble-Bound Phenolic Acids. 3. Composition of Phenolic Acids in Cereal and Potato Flours. J. Agric. Food Chem. 1982, 30, 337–340. [Google Scholar] [CrossRef]
- Antoine, C.; Peyron, S.; Lullien-Pellerin, V.; Abecassis, J.; Rouau, X. Wheat Bran Tissue Fractionation Using Biochemical Markers. J. Cereal Sci. 2004, 39, 387–393. [Google Scholar] [CrossRef]
- Adom, K.K.; Liu, R.H. Antioxidant Activity of Grains. J. Agric. Food Chem. 2002, 50, 6182–6187. [Google Scholar] [CrossRef]
- Renger, A.; Steinhart, H. Ferulic Acid Dehydrodimers as Structural Elements in Cereal Dietary Fibre. Eur. Food Res. Technol. 2000, 211, 422–428. [Google Scholar] [CrossRef]
- Ross, A.B.; Chen, Y.; Frank, J.; Swanson, J.E.; Parker, R.S.; Kozubek, A.; Lundh, T.; Vessby, B.; AÅman, P.; Kamal-Eldin, A. Cereal Alkylresorcinols Elevate γ-Tocopherol Levels in Rats and Inhibit γ-Tocopherol Metabolism In Vitro. J. Nutr. 2004, 134, 506–510. [Google Scholar] [CrossRef]
- Shah, I.; Shah, M.A.; Nawaz, M.A.; Pervez, S.; Noreen, N.; Vargas-de la Cruz, C.; Khan, F.; Blundell, R.; Briffa, J.; Azzopardi, J.; et al. Analysis of Other Phenolics (Capsaicin, Gingerol, and Alkylresorcinols). In Recent Advances in Natural Products Analysis; Elsevier: Amsterdam, The Netherlands, 2020; pp. 255–271. ISBN 978-0-12-816455-6. [Google Scholar]
- Ross, A.B.; Shepherd, M.J.; Schüpphaus, M.; Sinclair, V.; Alfaro, B.; Kamal-Eldin, A.; Åman, P. Alkylresorcinols in Cereals and Cereal Products. J. Agric. Food Chem. 2003, 51, 4111–4118. [Google Scholar] [CrossRef] [PubMed]
- Andersson, A.A.M.; Kamal-Eldin, A.; Fraś, A.; Boros, D.; Åman, P. Alkylresorcinols in Wheat Varieties in the HEALTHGRAIN Diversity Screen. J. Agric. Food Chem. 2008, 56, 9722–9725. [Google Scholar] [CrossRef]
- Kim, K.-H.; Tsao, R.; Yang, R.; Cui, S.W. Phenolic Acid Profiles and Antioxidant Activities of Wheat Bran Extracts and the Effect of Hydrolysis Conditions. Food Chem. 2006, 95, 466–473. [Google Scholar] [CrossRef]
- Verma, B.; Hucl, P.; Chibbar, R.N. Phenolic Acid Composition and Antioxidant Capacity of Acid and Alkali Hydrolysed Wheat Bran Fractions. Food Chem. 2009, 116, 947–954. [Google Scholar] [CrossRef]
- Bueno-Herrera, M.; Pérez-Magariño, S. Validation of an Extraction Method for the Quantification of Soluble Free and Insoluble Bound Phenolic Compounds in Wheat by HPLC-DAD. J. Cereal Sci. 2020, 93, 102984. [Google Scholar] [CrossRef]
- Arranz, S.; Saura Calixto, F. Analysis of Polyphenols in Cereals May Be Improved Performing Acidic Hydrolysis: A Study in Wheat Flour and Wheat Bran and Cereals of the Diet. J. Cereal Sci. 2010, 51, 313–318. [Google Scholar] [CrossRef]
- Mateo Anson, N.; van den Berg, R.; Havenaar, R.; Bast, A.; Haenen, G.R.M.M. Bioavailability of Ferulic Acid Is Determined by Its Bioaccessibility. J. Cereal Sci. 2009, 49, 296–300. [Google Scholar] [CrossRef]
- Höije, A.; Sternemalm, E.; Heikkinen, S.; Tenkanen, M.; Gatenholm, P. Material Properties of Films from Enzymatically Tailored Arabinoxylans. Biomacromolecules 2008, 9, 2042–2047. [Google Scholar] [CrossRef] [PubMed]
- Köhnke, T.; Östlund, Å.; Brelid, H. Adsorption of Arabinoxylan on Cellulosic Surfaces: Influence of Degree of Substitution and Substitution Pattern on Adsorption Characteristics. Biomacromolecules 2011, 12, 2633–2641. [Google Scholar] [CrossRef]
- Gruppen, H.; Hamer, R.J.; Voragen, A.G.J. Water-Unextractable Cell Wall Material from Wheat Flour. 1. Extraction of Polymers with Alkali. J. Cereal Sci. 1992, 16, 41–51. [Google Scholar] [CrossRef]
- Doehlert, D.C.; Simsek, S. Variation in β-Glucan Fine Structure, Extractability, and Flour Slurry Viscosity in Oats Due to Genotype and Environment. Cereal Chem. 2012, 89, 242–246. [Google Scholar] [CrossRef]
- Herrera, M.P.; Gao, J.; Vasanthan, T.; Temelli, F.; Henderson, K. β-Glucan Content, Viscosity, and Solubility of Canadian Grown Oat as Influenced by Cultivar and Growing Location. Can. J. Plant Sci. 2016, 96, 183–196. [Google Scholar] [CrossRef]
- Southgate, D.A.T.; Hudson, G.J.; Englyst, H. The Analysis of Dietary Fibre—The Choices for the Analyst. J. Sci. Food Agric. 1978, 29, 979–988. [Google Scholar] [CrossRef] [PubMed]
- McCleary, B.V.; DeVries, J.W.; Rader, J.I.; Cohen, G.; Prosky, L.; Mugford, D.C.; Champ, M.; Okuma, K. Determination of Total Dietary Fiber (CODEX Definition) by Enzymatic-Gravimetric Method and Liquid Chromatography: Collaborative Study. J. AOAC Int. 2010, 93, 221–233. [Google Scholar] [CrossRef] [PubMed]
- Englyst, H.N.; Quigley, M.E.; Hudson, G.J. Determination of Dietary Fibre as Non-Starch Polysaccharides with Gas–Liquid Chromatographic, High-Performance Liquid Chromatographic or Spectrophotometric Measurement of Constituent Sugars. Analyst 1994, 119, 1497–1509. [Google Scholar] [CrossRef] [PubMed]
- Elleuch, M.; Bedigian, D.; Roiseux, O.; Besbes, S.; Blecker, C.; Attia, H. Dietary Fibre and Fibre-Rich by-Products of Food Processing: Characterisation, Technological Functionality and Commercial Applications: A Review. Food Chem. 2011, 124, 411–421. [Google Scholar] [CrossRef]
- Megazyme. Total Dietary Fiber Assay Procedure K-TDFR-200A; Megazyme: Bray, Ireland, 2017. [Google Scholar]
- McCleary, B.; Sloane, N.; Draga, A.; Lazewska, I. Measurement of Total Dietary Fiber Using AOAC Method 2009.01 (AACC International Approved Method 32-45.01): Evaluation and Updates. Cereal Chem. 2013, 90, 396–414. [Google Scholar] [CrossRef]
- McCleary, B.V. Modification to AOAC Official Methods 2009.01 and 2011.25 to Allow for Minor Overestimation of Low Molecular Weight Soluble Dietary Fiber in Samples Containing Starch. J. AOAC Int. 2014, 97, 896–901. [Google Scholar] [CrossRef]
- De Bondt, Y.; Rosa-Sibakov, N.; Liberloo, I.; Roye, C.; Van de Walle, D.; Dewettinck, K.; Goos, P.; Nordlund, E.; Courtin, C.M. Study into the Effect of Microfluidisation Processing Parameters on the Physicochemical Properties of Wheat (Triticum Aestivum L.) Bran. Food Chem. 2020, 305, 125436. [Google Scholar] [CrossRef]
- Rosa-Sibakov, N.; Sibakov, J.; Lahtinen, P.; Poutanen, K. Wet Grinding and Microfluidization of Wheat Bran Preparations: Improvement of Dispersion Stability by Structural Disintegration. J. Cereal Sci. 2015, 64, 1–10. [Google Scholar] [CrossRef]
- Åman, P.; Rimsten, L.; Andersson, R. Molecular Weight Distribution of β-Glucan in Oat-Based Foods. Cereal Chem. 2004, 81, 356–360. [Google Scholar] [CrossRef]
- Martinant, J.P.; Cadalen, T.; Billot, A.; Chartier, S.; Leroy, P.; Bernard, M.; Saulnier, L.; Branlard, G. Genetic Analysis of Water-Extractable Arabinoxylans in Bread Wheat Endosperm. Appl. Genet. 1998, 97, 1069–1075. [Google Scholar] [CrossRef]
- Maes, C.; Delcour, J.A. Alkaline Hydrogen Peroxide Extraction of Wheat Bran Non-Starch Polysaccharides. J. Cereal Sci. 2001, 34, 29–35. [Google Scholar] [CrossRef]
- Gómez, C.; Navarro, A.; Manzanares, P.; Horta, A.; Carbonell, J.V. Physical and Structural Properties of Barley (1 → 3),(1 → 4)-β-d-Glucan. Part II. Viscosity, Chain Stiffness and Macromolecular Dimensions. Carbohydr. Polym. 1997, 32, 17–22. [Google Scholar] [CrossRef]
- Yao, W.; Gong, Y.; Li, L.; Hu, X.; You, L. The Effects of Dietary Fibers from Rice Bran and Wheat Bran on Gut Microbiota: An Overview. Food Chem. X 2022, 13, 100252. [Google Scholar] [CrossRef]
- Chau, C.F.; Huang, Y.L. Characterization of Passion Fruit Seed Fibres—A Potential Fibre Source. Food Chem. 2004, 85, 189–194. [Google Scholar] [CrossRef]
- Kethireddipalli, P.; Hung, Y.-C.; Phillips, R.D.; Mcwatters, K.H. Evaluating the Role of Cell Wall Material and Soluble Protein in the Functionality of Cowpea (Vigna Unguiculata) Pastes. J. Food Sci. 2002, 67, 53–59. [Google Scholar] [CrossRef]
- Kweon, M.; Slade, L.; Levine, H. Solvent Retention Capacity (SRC) Testing of Wheat Flour: Principles and Value in Predicting Flour Functionality in Different Wheat-Based Food Processes and in Wheat Breeding—A Review. Cereal Chem. 2011, 88, 537–552. [Google Scholar] [CrossRef]
- Courtin, C.M.; Delcour, J.A. Arabinoxylans and Endoxylanases in Wheat Flour Bread-Making. J. Cereal Sci. 2002, 35, 225–243. [Google Scholar] [CrossRef]
- Lovegrove, A.; Wood, A.J.; Hassall, K.L.; Howes, L.; Poole, M.; Tosi, P.; Shewry, P. The Contribution of Fiber Components to Water Absorption of Wheat Grown in the UK. Cereal Chem. 2020, 97, 940–948. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.; Cui, S.W.; Gu, X.; Zhang, J. Isolation and Structural Characterization of Water Unextractable Arabinoxylans from Chinese Black-Grained Wheat Bran. Carbohydr. Polym. 2011, 85, 615–621. [Google Scholar] [CrossRef]
- Izydorczyk, M.S.; Biliaderis, C.G. Cereal Arabinoxylans: Advances in Structure and Physicochemical Properties. Carbohydr. Polym. 1995, 28, 33–48. [Google Scholar] [CrossRef]
- Ahmad, A.; Kaleem, M. Chapter 11—β-Glucan as a Food Ingredient. In Biopolymers for Food Design; Handbook of Food Bioengineering; Grumezescu, A.M., Holban, A.M., Eds.; Academic Press: Cambridge, MA, USA, 2018; pp. 351–381. ISBN 978-0-12-811449-0. [Google Scholar]
- Tejinder, S.; Bhupinder, K.; Harinder, K. Flow Behavior and Functional Properties of Barley and Oat Water-soluble Β-D-glucan Rich Extractions. Int. J. Food Prop. 2000, 3, 259–274. [Google Scholar] [CrossRef]
- Capuano, E. The Behavior of Dietary Fiber in the Gastrointestinal Tract Determines Its Physiological Effect. Crit. Rev. Food Sci. Nutr. 2017, 57, 3543–3564. [Google Scholar] [CrossRef] [PubMed]
- Lu, X.; Jing, Y.; Li, Y.; Zhang, N.; Cao, Y. Eurotium Cristatum Produced β-Hydroxy Acid Metabolite of Monacolin K and Improved Bioactive Compound Contents as Well as Functional Properties in Fermented Wheat Bran. LWT 2022, 158, 113088. [Google Scholar] [CrossRef]
- Lai, S.; Chen, Z.; Zhang, Y.; Li, G.; Wang, Y.; Cui, Q. Micronization Effects on Structural, Functional, and Antioxidant Properties of Wheat Bran. Foods 2023, 12, 98. [Google Scholar] [CrossRef] [PubMed]
- Kinsella, J.E.; Melachouris, N. Functional Properties of Proteins in Foods: A Survey. Crit. Rev. Food Sci. Nutr. 1976, 7, 219–280. [Google Scholar] [CrossRef]
- He, Y.; Wang, B.; Wen, L.; Wang, F.; Yu, H.; Chen, D.; Su, X.; Zhang, C. Effects of Dietary Fiber on Human Health. Food Sci. Hum. Wellness 2022, 11, 1–10. [Google Scholar] [CrossRef]
- Adiotomre, J.; Eastwood, M.; Edwards, C.; Brydon, W. Dietary Fiber: In Vitro Methods That Anticipate Nutrition and Metabolic Activity in Humans. Am. J. Clin. Nutr. 1990, 52, 128–134. [Google Scholar] [CrossRef]
- Chau, C.-F.; Huang, Y.-L.; Lee, M.-H. In Vitro Hypoglycemic Effects of Different Insoluble Fiber-Rich Fractions Prepared from the Peel of Citrus sinensis L. Cv. Liucheng. J. Agric. Food Chem. 2003, 51, 6623–6626. [Google Scholar] [CrossRef]
- Ou, S.; Gao, K.; Li, Y. An in Vitro Study of Wheat Bran Binding Capacity for Hg, Cd, and Pb. J. Agric. Food Chem. 1999, 47, 4714–4717. [Google Scholar] [CrossRef] [PubMed]
- Ou, S.; Kwok, K.; Li, Y.; Fu, L. In Vitro Study of Possible Role of Dietary Fiber in Lowering Postprandial Serum Glucose. J. Agric. Food Chem. 2001, 49, 1026–1029. [Google Scholar] [CrossRef]
- Ahmed, F.; Sairam, S.; Urooj, A. In Vitro Hypoglycemic Effects of Selected Dietary Fiber Sources. J. Food Sci. Technol. 2011, 48, 285–289. [Google Scholar] [CrossRef] [PubMed]
- Ma, M.; Mu, T.; Sun, H.; Zhang, M.; Chen, J.; Yan, Z. Optimization of Extraction Efficiency by Shear Emulsifying Assisted Enzymatic Hydrolysis and Functional Properties of Dietary Fiber from Deoiled Cumin (Cuminum cyminum L.). Food Chem. 2015, 179, 270–277. [Google Scholar] [CrossRef] [PubMed]
- Yan, J.-K.; Wu, L.-X.; Cai, W.-D.; Xiao, G.-S.; Duan, Y.; Zhang, H. Subcritical Water Extraction-Based Methods Affect the Physicochemical and Functional Properties of Soluble Dietary Fibers from Wheat Bran. Food Chem. 2019, 298, 124987. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Ye, R.; Yin, L.; Zhang, N. Novel Blasting Extrusion Processing Improved the Physicochemical Properties of Soluble Dietary Fiber from Soybean Residue and in Vivo Evaluation. J. Food Eng. 2014, 120, 1–8. [Google Scholar] [CrossRef]
- Hua, M.; Lu, J.; Qu, D.; Liu, C.; Zhang, L.; Li, S.; Chen, J.; Sun, Y. Structure, Physicochemical Properties and Adsorption Function of Insoluble Dietary Fiber from Ginseng Residue: A Potential Functional Ingredient. Food Chem. 2019, 286, 522–529. [Google Scholar] [CrossRef]
- Wu, T.; Li, Z.; Liu, R.; Sui, W.; Zhang, M. Effect of Extrusion, Steam Explosion and Enzymatic Hydrolysis on Functional Properties of Wheat Bran. Food Sci. Technol. Res. 2018, 24, 591–598. [Google Scholar] [CrossRef]
- Singh, J.; Metrani, R.; Shivanagoudra, S.R.; Jayaprakasha, G.K.; Patil, B.S. Review on Bile Acids: Effects of the Gut Microbiome, Interactions with Dietary Fiber, and Alterations in the Bioaccessibility of Bioactive Compounds. J. Agric. Food Chem. 2019, 67, 9124–9138. [Google Scholar] [CrossRef]
- Camire, M.E.; Zhao, J.; Violette, D.A. In Vitro Binding of Bile Acids by Extruded Potato Peels. J. Agric. Food Chem. 1993, 41, 2391–2394. [Google Scholar] [CrossRef]
- Kahlon, T.S.; Chow, F.I. In Vitro Binding of Bile Acids by Rice Bran, Oat Bran, Wheat Bran, and Corn Bran. Cereal Chem. J. 2000, 77, 518–521. [Google Scholar] [CrossRef]
- Xu, H.; Jiao, Q.; Yuan, F.; Gao, Y. In Vitro Binding Capacities and Physicochemical Properties of Soluble Fiber Prepared by Microfluidization Pretreatment and Cellulase Hydrolysis of Peach Pomace. LWT—Food Sci. Technol. 2015, 63, 677–684. [Google Scholar] [CrossRef]
- Chau, C.-F.; Cheung, P.C.-K. Effects of the Physico-Chemical Properties of Three Legume Fibers on Cholesterol Absorption in Hamsters. Nutr. Res. 1999, 19, 257–265. [Google Scholar] [CrossRef]
- Ghodrat, A.; Yaghobfar, A.; Ebrahimnezhad, Y.; Aghdam Shahryar, H.; Ghorbani, A. In Vitro Binding Capacity of Organic (Wheat Bran and Rice Bran) and Inorganic (Perlite) Sources for Mn, Zn, Cu, and Fe. J. Appl. Anim. Res. 2017, 45, 80–84. [Google Scholar] [CrossRef]
- Zhu, F.; Du, B.; Xu, B. Superfine Grinding Improves Functional Properties and Antioxidant Capacities of Bran Dietary Fibre from Qingke (Hull-Less Barley) Grown in Qinghai-Tibet Plateau, China. J. Cereal Sci. 2015, 65, 43–47. [Google Scholar] [CrossRef]
- Wang, Q.-H.; Yu, L.-J.; Liu, Y.; Lin, L.; Lu, R.; Zhu, J.; He, L.; Lu, Z.-L. Methods for the Detection and Determination of Nitrite and Nitrate: A Review. Talanta 2017, 165, 709–720. [Google Scholar] [CrossRef]
- Zhang, N.; Huang, C.; Ou, S. In Vitro Binding Capacities of Three Dietary Fibers and Their Mixture for Four Toxic Elements, Cholesterol, and Bile Acid. J. Hazard. Mater. 2011, 186, 236–239. [Google Scholar] [CrossRef]
- Jacobs, P.J.; Hemdane, S.; Dornez, E.; Delcour, J.A.; Courtin, C.M. Study of Hydration Properties of Wheat Bran as a Function of Particle Size. Food Chem. 2015, 179, 296–304. [Google Scholar] [CrossRef]
- Zhao, H.-M.; Guo, X.-N.; Zhu, K.-X. Impact of Solid State Fermentation on Nutritional, Physical and Flavor Properties of Wheat Bran. Food Chem. 2017, 217, 28–36. [Google Scholar] [CrossRef]
- Ma, J.; Yuan, M.; Liu, Y.; Duan, Y.; Zhang, D.; Shao, Y.; Wang, T.; Jiang, L. Effects of Steam Explosion on Yield and Properties of Soluble Dietary Fiber from Wheat Bran. Food Sci. Technol. Res. 2021, 27, 35–42. [Google Scholar] [CrossRef]
- Caprez, A.; Arrigoni, E.; Amadò, R.; Neukom, H. Influence of Different Types of Thermal Treatment on the Chemical Composition and Physical Properties of Wheat Bran. J. Cereal Sci. 1986, 4, 233–239. [Google Scholar] [CrossRef]
- Liu, X.; Suo, K.; Wang, P.; Li, X.; Hao, L.; Zhu, J.; Yi, J.; Kang, Q.; Huang, J.; Lu, J. Modification of Wheat Bran Insoluble and Soluble Dietary Fibers with Snail Enzyme. Food Sci. Hum. Wellness 2021, 10, 356–361. [Google Scholar] [CrossRef]
- Vuksan, V.; Jenkins, A.L.; Rogovik, A.L.; Fairgrieve, C.D.; Jovanovski, E.; Leiter, L.A. Viscosity Rather than Quantity of Dietary Fibre Predicts Cholesterol-Lowering Effect in Healthy Individuals. Br. J. Nutr. 2011, 106, 1349–1352. [Google Scholar] [CrossRef] [PubMed]
- Fardet, A. New hypotheses for the health-protective mechanisms of whole-grain cereals: What is beyond fibre? Nutr. Res. Rev. 2010, 23, 65–134. [Google Scholar] [CrossRef]
- Hussain, T.; Tan, B.; Yin, Y.; Blachier, F.; Tossou, M.C.B.; Rahu, N. Oxidative Stress and Inflammation: What Polyphenols Can Do for Us? Oxidative Med. Cell. Longev. 2016, 2016, e7432797. [Google Scholar] [CrossRef] [PubMed]
- Collin, S.; Crouzet, J. Polyphénols ET Procédés: Transformation Des Polyphénols AU Travers Des Procédés Appliqués à L’Agro-Alimentaire; Collection de L’Agence Universitaire de la Francophonie; Tec & Doc Lavoisier, Ed.; Agence Universitaire de la Francophonie: Paris, France; Montréal, QC, Canada, 2011; ISBN 978-2-7430-1338-7. [Google Scholar]
- Singleton, V.L.; Orthofer, R.; Lamuela-Raventós, R.M. Analysis of Total Phenols and Other Oxidation Substrates and Antioxidants by Means of Folin-Ciocalteu Reagent. In Methods in Enzymology; Oxidants and Antioxidants Part A; Academic Press: Cambridge, MA, USA, 1999; Volume 299, pp. 152–178. [Google Scholar]
- Bibi Sadeer, N.; Montesano, D.; Albrizio, S.; Zengin, G.; Mahomoodally, M.F. The Versatility of Antioxidant Assays in Food Science and Safety—Chemistry, Applications, Strengths, and Limitations. Antioxidants 2020, 9, 709. [Google Scholar] [CrossRef]
- López-Fernández, O.; Domínguez, R.; Pateiro, M.; Munekata, P.E.S.; Rocchetti, G.; Lorenzo, J.M. Determination of Polyphenols Using Liquid Chromatography-Tandem Mass Spectrometry Technique (LC-MS/MS): A Review. Antioxidants 2020, 9, 479. [Google Scholar] [CrossRef] [PubMed]
- Alam, M.N.; Bristi, N.J.; Rafiquzzaman, M. Review on in Vivo and in Vitro Methods Evaluation of Antioxidant Activity. Saudi Pharm. J. 2013, 21, 143–152. [Google Scholar] [CrossRef]
- Ross, K.A.; Beta, T.; Arntfield, S.D. A Comparative Study on the Phenolic Acids Identified and Quantified in Dry Beans Using HPLC as Affected by Different Extraction and Hydrolysis Methods. Food Chem. 2009, 113, 336–344. [Google Scholar] [CrossRef]
- Bruijnincx, P.; Weckhuysen, B.; Gruter, G.-J.; Westenbroek, A.; Engelen-Smeets, E. Lignin Valorisation: The Importance of a Full Value Chain Approach; Utrecht University: Utrecht, The Netherlands, 2016; p. 22. [Google Scholar]
- Tanasković, S.J.; Šekuljica, N.; Jovanović, J.; Gazikalović, I.; Grbavčić, S.; Đorđević, N.; Sekulić, M.V.; Hao, J.; Luković, N.; Knežević-Jugović, Z. Upgrading of Valuable Food Component Contents and Anti-Nutritional Factors Depletion by Solid-State Fermentation: A Way to Valorize Wheat Bran for Nutrition. J. Cereal Sci. 2021, 99, 103159. [Google Scholar] [CrossRef]
- Santos Sánchez, N.; Salas-Coronado, R.; Villanueva, C.; Hernández-Carlos, B. Antioxidant Compounds and Their Antioxidant Mechanism; IntechOpen: London, UK, 2019; ISBN 978-1-78923-919-5. [Google Scholar]
- Abozed, S.S.; El-kalyoubi, M.; Abdelrashid, A.; Salama, M.F. Total Phenolic Contents and Antioxidant Activities of Various Solvent Extracts from Whole Wheat and Bran. Ann. Agric. Sci. 2014, 59, 63–67. [Google Scholar] [CrossRef]
- López-Perea, P.; Guzmán-Ortiz, F.A.; Román-Gutiérrez, A.D.; Castro-Rosas, J.; Gómez-Aldapa, C.A.; Rodríguez-Marín, M.L.; Falfán-Cortés, R.N.; González-Olivares, L.G.; Torruco-Uco, J.G. Bioactive Compounds and Antioxidant Activity of Wheat Bran and Barley Husk in the Extracts with Different Polarity. Int. J. Food Prop. 2019, 22, 646–658. [Google Scholar] [CrossRef]
- Vaher, M.; Matso, K.; Levandi, T.; Helmja, K.; Kaljurand, M. Phenolic Compounds and the Antioxidant Activity of the Bran, Flour and Whole Grain of Different Wheat Varieties. Procedia Chem. 2010, 2, 76–82. [Google Scholar] [CrossRef]
- Floegel, A.; Kim, D.-O.; Chung, S.-J.; Koo, S.I.; Chun, O.K. Comparison of ABTS/DPPH Assays to Measure Antioxidant Capacity in Popular Antioxidant-Rich US Foods. J. Food Compos. Anal. 2011, 24, 1043–1048. [Google Scholar] [CrossRef]
- Esposito, F.; Arlotti, G.; Maria Bonifati, A.; Napolitano, A.; Vitale, D.; Fogliano, V. Antioxidant Activity and Dietary Fibre in Durum Wheat Bran By-Products. Food Res. Int. 2005, 38, 1167–1173. [Google Scholar] [CrossRef]
- Graf, E. Antioxidant Potential of Ferulic Acid. Free Radic. Biol. Med. 1992, 13, 435–448. [Google Scholar] [CrossRef]
- Brewer, L.R.; Kubola, J.; Siriamornpun, S.; Herald, T.J.; Shi, Y.-C. Wheat Bran Particle Size Influence on Phytochemical Extractability and Antioxidant Properties. Food Chem. 2014, 152, 483–490. [Google Scholar] [CrossRef]
- Zhu, K.; Huang, S.; Peng, W.; Qian, H.; Zhou, H. Effect of Ultrafine Grinding on Hydration and Antioxidant Properties of Wheat Bran Dietary Fiber. Food Res. Int. 2010, 43, 943–948. [Google Scholar] [CrossRef]
- Benguella, R.; Meziani, S.; Zohra, C.F.; Barek, S.; Aissaoui, M.; Nadjib Rahmoun, M.; Demmouche, A. Comparison of the Nutritional and Antioxidant Values of the Peripheral Layers in Two Species of Wheat (Soft and Hard) Grown in Algeria. Chil. J. Agric. Anim. Sci. 2022, 38, 15–25. [Google Scholar] [CrossRef]
- Sparkman, O.D.; Penton, Z.E.; Kitson, F.G. Chapter 2—Gas Chromatography. In Gas Chromatography and Mass Spectrometry, 2nd ed.; Sparkman, O.D., Penton, Z.E., Kitson, F.G., Eds.; Academic Press: Amsterdam, The Netherlands, 2011; pp. 15–83. ISBN 978-0-12-373628-4. [Google Scholar]
- Spaggiari, M.; Ricci, A.; Calani, L.; Bresciani, L.; Neviani, E.; Dall’Asta, C.; Lazzi, C.; Galaverna, G. Solid State Lactic Acid Fermentation: A Strategy to Improve Wheat Bran Functionality. LWT 2020, 118, 108668. [Google Scholar] [CrossRef]
- Hromádková, Z.; Paulsen, B.S.; Polovka, M.; Košťálová, Z.; Ebringerová, A. Structural Features of Two Heteroxylan Polysaccharide Fractions from Wheat Bran with Anti-Complementary and Antioxidant Activities. Carbohydr. Polym. 2013, 93, 22–30. [Google Scholar] [CrossRef] [PubMed]
- Gidley, M.J.; Yakubov, G.E. Functional Categorisation of Dietary Fibre in Foods: Beyond ‘Soluble’ vs. ‘Insoluble’. Trends Food Sci. Technol. 2019, 86, 563–568. [Google Scholar] [CrossRef]
- Gunness, P.; Gidley, M.J. Mechanisms Underlying the Cholesterol-Lowering Properties of Soluble Dietary Fibre Polysaccharides. Food Funct. 2010, 1, 149. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Du, J. Molecular Characterization of Arabinoxylan from Wheat Beer, Beer Foam and Defoamed Beer. Molecules 2019, 24, 1230. [Google Scholar] [CrossRef] [PubMed]
- Thondre, P.S.; Shafat, A.; Clegg, M.E. Molecular Weight of Barley β-Glucan Influences Energy Expenditure, Gastric Emptying and Glycaemic Response in Human Subjects. Br. J. Nutr. 2013, 110, 2173–2179. [Google Scholar] [CrossRef]
- Ninomiya, K.; Ina, S.; Nakamura, H.; Yamaguchi, Y.; Kumagai, H.; Kumagai, H. Evaluation of the Amount of Glucose Adsorbed on Water-Soluble Dietary Fibres by the Analysis of Its Diffusion Rate through a Dialysis Membrane. Food Hydrocoll. 2022, 129, 107626. [Google Scholar] [CrossRef]
- Shen, X.L.; Zhao, T.; Zhou, Y.; Shi, X.; Zou, Y.; Zhao, G. Effect of Oat β-Glucan Intake on Glycaemic Control and Insulin Sensitivity of Diabetic Patients: A Meta-Analysis of Randomized Controlled Trials. Nutrients 2016, 8, 39. [Google Scholar] [CrossRef]
- Pino, J.L.; Mujica, V.; Arredondo, M. Effect of Dietary Supplementation with Oat β-Glucan for 3 months in Subjects with Type 2 Diabetes: A Randomized, Double-Blind, Controlled Clinical Trial. J. Funct. Foods 2021, 77, 104311. [Google Scholar] [CrossRef]
- Cabou, C.; Burcelin, R. GLP-1, the Gut-Brain, and Brain-Periphery Axes. Rev. Diabet. Stud. RDS 2011, 8, 418–431. [Google Scholar] [CrossRef]
- Mio, K.; Ogawa, R.; Tadenuma, N.; Aoe, S. Arabinoxylan as Well as β-Glucan in Barley Promotes GLP-1 Secretion by Increasing Short-Chain Fatty Acids Production. Biochem. Biophys. Rep. 2022, 32, 101343. [Google Scholar] [CrossRef]
- Bays, H.; Frestedt, J.L.; Bell, M.; Williams, C.; Kolberg, L.; Schmelzer, W.; Anderson, J.W. Reduced Viscosity Barley β-Glucan versus Placebo: A Randomized Controlled Trial of the Effects on Insulin Sensitivity for Individuals at Risk for Diabetes Mellitus. Nutr. Metab. 2011, 8, 58. [Google Scholar] [CrossRef] [PubMed]
- Eberlé, D.; Hegarty, B.; Bossard, P.; Ferré, P.; Foufelle, F. SREBP Transcription Factors: Master Regulators of Lipid Homeostasis. Biochimie 2004, 86, 839–848. [Google Scholar] [CrossRef] [PubMed]
- Garcia, A.L.; Otto, B.; Reich, S.-C.; Weickert, M.O.; Steiniger, J.; Machowetz, A.; Rudovich, N.N.; Möhlig, M.; Katz, N.; Speth, M.; et al. Arabinoxylan Consumption Decreases Postprandial Serum Glucose, Serum Insulin and Plasma Total Ghrelin Response in Subjects with Impaired Glucose Tolerance. Eur. J. Clin. Nutr. 2007, 61, 334–341. [Google Scholar] [CrossRef] [PubMed]
- Broglio, F.; Gottero, C.; Prodam, F.; Gauna, C.; Muccioli, G.; Papotti, M.; Abribat, T.; van der Lely, A.J.; Ghigo, E. Non-Acylated Ghrelin Counteracts the Metabolic But Not the Neuroendocrine Response to Acylated Ghrelin in Humans. J. Clin. Endocrinol. Metab. 2004, 89, 3062–3065. [Google Scholar] [CrossRef] [PubMed]
- Miao, H.; Pan, H.; Wang, L.; Yang, H.; Zhu, H.; Gong, F. Ghrelin Promotes Proliferation and Inhibits Differentiation of 3T3-L1 and Human Primary Preadipocytes. Front. Physiol. 2019, 10, 1296. [Google Scholar] [CrossRef]
- Lafond, D.; Greaves, K.; Maki, K.; Leidy, H.; Romsos, D. Effects of Two Dietary Fibers as Part of Ready-to-Eat Cereal (RTEC) Breakfasts on Perceived Appetite and Gut Hormones in Overweight Women. Nutrients 2015, 7, 1245–1266. [Google Scholar] [CrossRef]
- Habib, A.M.; Richards, P.; Rogers, G.J.; Reimann, F.; Gribble, F.M. Co-Localisation and Secretion of Glucagon-like Peptide 1 and Peptide YY from Primary Cultured Human L Cells. Diabetologia 2013, 56, 1413–1416. [Google Scholar] [CrossRef]
- Gulati, S.; Misra, A.; Pandey, R.M. Effects of 3 g of Soluble Fiber from Oats on Lipid Levels of Asian Indians—A Randomized Controlled, Parallel Arm Study. Lipids Health Dis. 2017, 16, 71. [Google Scholar] [CrossRef]
- Ortiz de Erive, M.; He, F.; Wang, T.; Chen, G. Development of β-Glucan Enriched Wheat Bread Using Soluble Oat Fiber. J. Cereal Sci. 2020, 95, 103051. [Google Scholar] [CrossRef]
- Sivieri, K.; de Oliveira, S.M.; de Souza Marquez, A.; Pérez-Jiménez, J.; Diniz, S.N. Insights on β-Glucan as a Prebiotic Coadjuvant in the Treatment of Diabetes Mellitus: A Review. Food Hydrocoll. Health 2022, 2, 100056. [Google Scholar] [CrossRef]
- Keenan, J.M.; Goulson, M.; Shamliyan, T.; Knutson, N.; Kolberg, L.; Curry, L. The Effects of Concentrated Barley Beta-Glucan on Blood Lipids in a Population of Hypercholesterolaemic Men and Women. Br. J. Nutr. 2007, 97, 1162–1168. [Google Scholar] [CrossRef]
- Wolever, T.M.S.; Tosh, S.M.; Gibbs, A.L.; Brand-Miller, J.; Duncan, A.M.; Hart, V.; Lamarche, B.; Thomson, B.A.; Duss, R.; Wood, P.J. Physicochemical Properties of Oat β-Glucan Influence Its Ability to Reduce Serum LDL Cholesterol in Humans: A Randomized Clinical Trial. Am. J. Clin. Nutr. 2010, 92, 723–732. [Google Scholar] [CrossRef]
- Thandapilly, S.J.; Ndou, S.P.; Wang, Y.; Nyachoti, C.M.; Ames, N.P. Barley β-Glucan Increases Fecal Bile Acid Excretion and Short Chain Fatty Acid Levels in Mildly Hypercholesterolemic Individuals. Food Funct. 2018, 9, 3092–3096. [Google Scholar] [CrossRef] [PubMed]
- Neyrinck, A.M.; Possemiers, S.; Druart, C.; Van de Wiele, T.; De Backer, F.; Cani, P.D.; Larondelle, Y.; Delzenne, N.M. Prebiotic Effects of Wheat Arabinoxylan Related to the Increase in Bifidobacteria, Roseburia and Bacteroides/Prevotella in Diet-Induced Obese Mice. PLoS ONE 2011, 6, e20944. [Google Scholar] [CrossRef] [PubMed]
- Salden, B.N.; Troost, F.J.; Wilms, E.; Truchado, P.; Vilchez-Vargas, R.; Pieper, D.H.; Jáuregui, R.; Marzorati, M.; van de Wiele, T.; Possemiers, S.; et al. Reinforcement of Intestinal Epithelial Barrier by Arabinoxylans in Overweight and Obese Subjects: A Randomized Controlled Trial: Arabinoxylans in Gut Barrier. Clin. Nutr. 2018, 37, 471–480. [Google Scholar] [CrossRef]
- Cloetens, L.; Broekaert, W.F.; Delaedt, Y.; Ollevier, F.; Courtin, C.M.; Delcour, J.A.; Rutgeerts, P.; Verbeke, K. Tolerance of Arabinoxylan-Oligosaccharides and Their Prebiotic Activity in Healthy Subjects: A Randomised, Placebo-Controlled Cross-over Study. Br. J. Nutr. 2010, 103, 703–713. [Google Scholar] [CrossRef] [PubMed]
- François, I.E.J.A.; Lescroart, O.; Veraverbeke, W.S.; Marzorati, M.; Possemiers, S.; Evenepoel, P.; Hamer, H.; Houben, E.; Windey, K.; Welling, G.W.; et al. Effects of a Wheat Bran Extract Containing Arabinoxylan Oligosaccharides on Gastrointestinal Health Parameters in Healthy Adult Human Volunteers: A Double-Blind, Randomised, Placebo-Controlled, Cross-over Trial. Br. J. Nutr. 2012, 108, 2229–2242. [Google Scholar] [CrossRef] [PubMed]
- Kjølbæk, L.; Benítez-Páez, A.; Gómez del Pulgar, E.M.; Brahe, L.K.; Liebisch, G.; Matysik, S.; Rampelli, S.; Vermeiren, J.; Brigidi, P.; Larsen, L.H.; et al. Arabinoxylan Oligosaccharides and Polyunsaturated Fatty Acid Effects on Gut Microbiota and Metabolic Markers in Overweight Individuals with Signs of Metabolic Syndrome: A Randomized Cross-over Trial. Clin. Nutr. 2020, 39, 67–79. [Google Scholar] [CrossRef]
- Van Craeyveld, V.; Swennen, K.; Dornez, E.; Van de Wiele, T.; Marzorati, M.; Verstraete, W.; Delaedt, Y.; Onagbesan, O.; Decuypere, E.; Buyse, J.; et al. Structurally Different Wheat-Derived Arabinoxylooligosaccharides Have Different Prebiotic and Fermentation Properties in Rats. J. Nutr. 2008, 138, 2348–2355. [Google Scholar] [CrossRef]
- Hosseini, E.; Grootaert, C.; Verstraete, W.; Van de Wiele, T. Propionate as a Health-Promoting Microbial Metabolite in the Human Gut. Nutr. Rev. 2011, 69, 245–258. [Google Scholar] [CrossRef]
- Kibbie, J.J.; Dillon, S.M.; Thompson, T.A.; Purba, C.M.; McCarter, M.D.; Wilson, C.C. Butyrate Directly Decreases Human Gut Lamina Propria CD4 T Cell Function through Histone Deacetylase (HDAC) Inhibition and GPR43 Signaling. Immunobiology 2021, 226, 152126. [Google Scholar] [CrossRef]
- Yang, J.; Martínez, I.; Walter, J.; Keshavarzian, A.; Rose, D.J. In Vitro Characterization of the Impact of Selected Dietary Fibers on Fecal Microbiota Composition and Short Chain Fatty Acid Production. Anaerobe 2013, 23, 74–81. [Google Scholar] [CrossRef]
- Paesani, C.; Sciarini, L.S.; Moiraghi, M.; Salvucci, E.; Prado, S.B.R.; Pérez, G.T.; Fabi, J.P. Human Colonic in Vitro Fermentation of Water-Soluble Arabinoxylans from Hard and Soft Wheat Alters Bifidobacterium Abundance and Short-Chain Fatty Acids Concentration. LWT 2020, 134, 110253. [Google Scholar] [CrossRef]
- Hughes, R.; Magee, E.A.M.; Bingham, S. Protein Degradation in the Large Intestine: Relevence to Colorectal Cancer. Curr. Issues Intestig. Microbiol. 2000, 1, 51–58. [Google Scholar]
- Shimada, R.; Yoshimura, M.; Murakami, K.; Ebihara, K. Plasma Concentrations of GLP-1 and PYY in Rats Fed Dietary Fiber Depend on the Fermentability of Dietary Fiber and Respond to an Altered Diet. Int. J. Clin. Nutr. Diet 2015, 1, 103. [Google Scholar] [CrossRef] [PubMed]
- Jefferson, A.; Adolphus, K. The Effects of Intact Cereal Grain Fibers, Including Wheat Bran on the Gut Microbiota Composition of Healthy Adults: A Systematic Review. Front. Nutr. 2019, 6, 33. [Google Scholar] [CrossRef] [PubMed]
- Hughes, S.A.; Shewry, P.R.; Gibson, G.R.; McCleary, B.V.; Rastall, R.A. In Vitro Fermentation of Oat and Barley Derived Beta-Glucans by Human Faecal Microbiota. FEMS Microbiol. Ecol. 2008, 64, 482–493. [Google Scholar] [CrossRef] [PubMed]
- Aoe, S.; Mio, K.; Yamanaka, C.; Kuge, T. Low Molecular Weight Barley β-Glucan Affects Glucose and Lipid Metabolism by Prebiotic Effects. Nutrients 2020, 13, 130. [Google Scholar] [CrossRef]
- Wang, Y.; Ames, N.; Li, S.; Jones, P.; Khafipour, E. High Molecular Weight Barley β-Glucan Supports Bacterial Populations Beneficial for Gut Health (647.45). FASEB J. 2014, 28, 647.45. [Google Scholar] [CrossRef]
- Gabriel, F.C.; Fantuzzi, G. The Association of Short-Chain Fatty Acids and Leptin Metabolism: A Systematic Review. Nutr. Res. 2019, 72, 18–35. [Google Scholar] [CrossRef]
- Velikonja, A.; Lipoglavšek, L.; Zorec, M.; Orel, R.; Avguštin, G. Alterations in Gut Microbiota Composition and Metabolic Parameters after Dietary Intervention with Barley Beta Glucans in Patients with High Risk for Metabolic Syndrome Development. Anaerobe 2019, 55, 67–77. [Google Scholar] [CrossRef]
- Snelders, J.; Olaerts, H.; Dornez, E.; Van de Wiele, T.; Aura, A.-M.; Vanhaecke, L.; Delcour, J.A.; Courtin, C.M. Structural Features and Feruloylation Modulate the Fermentability and Evolution of Antioxidant Properties of Arabinoxylanoligosaccharides during in Vitro Fermentation by Human Gut Derived Microbiota. J. Funct. Foods 2014, 10, 1–12. [Google Scholar] [CrossRef]
- Lecerf, J.-M.; Dépeint, F.; Clerc, E.; Dugenet, Y.; Niamba, C.N.; Rhazi, L.; Cayzeele, A.; Abdelnour, G.; Jaruga, A.; Younes, H.; et al. Xylo-Oligosaccharide (XOS) in Combination with Inulin Modulates Both the Intestinal Environment and Immune Status in Healthy Subjects, While XOS Alone Only Shows Prebiotic Properties. Br. J. Nutr. 2012, 108, 1847–1858. [Google Scholar] [CrossRef]
- Damen, B.; Cloetens, L.; Broekaert, W.F.; François, I.; Lescroart, O.; Trogh, I.; Arnaut, F.; Welling, G.W.; Wijffels, J.; Delcour, J.A.; et al. Consumption of Breads Containing In Situ–Produced Arabinoxylan Oligosaccharides Alters Gastrointestinal Effects in Healthy Volunteers. J. Nutr. 2012, 142, 470–477. [Google Scholar] [CrossRef]
- Maki, K.C.; Gibson, G.R.; Dickmann, R.S.; Kendall, C.W.C.; Chen, C.-Y.O.; Costabile, A.; Comelli, E.M.; McKay, D.L.; Almeida, N.G.; Jenkins, D.; et al. Digestive and Physiologic Effects of a Wheat Bran Extract, Arabino-Xylan-Oligosaccharide, in Breakfast Cereal. Nutrition 2012, 28, 1115–1121. [Google Scholar] [CrossRef] [PubMed]
- Walton, G.E.; Lu, C.; Trogh, I.; Arnaut, F.; Gibson, G.R. A Randomised, Double-Blind, Placebo Controlled Cross-over Study to Determine the Gastrointestinal Effects of Consumption of Arabinoxylan-Oligosaccharides Enriched Bread in Healthy Volunteers. Nutr. J. 2012, 11, 36. [Google Scholar] [CrossRef] [PubMed]
- Hamer, H.M.; Jonkers, D.; Venema, K.; Vanhoutvin, S.; Troost, F.J.; Brummer, R.-J. Review Article: The Role of Butyrate on Colonic Function. Aliment. Pharmacol. Ther. 2007, 27, 104–119. [Google Scholar] [CrossRef]
- Mendez-Encinas, M.A.; Valencia-Rivera, D.E.; Carvajal-Millan, E.; Astiazaran-Garcia, H.; Micard, V.; Rascón-Chu, A. Fermentation of Ferulated Arabinoxylan Recovered from the Maize Bioethanol Industry. Processes 2021, 9, 165. [Google Scholar] [CrossRef]
- Estrada, A.; Yun, C.; Van Kessel, A.; Li, B.; Hauta, S.; Laarveld, B. Immunomodulatory Activities of Oat β-Glucan In Vitro and In Vivo. Microbiol. Immunol. 1997, 41, 991–998. [Google Scholar] [CrossRef]
- Volman, J.J.; Ramakers, J.D.; Plat, J. Dietary Modulation of Immune Function by β-Glucans. Physiol. Behav. 2008, 94, 276–284. [Google Scholar] [CrossRef] [PubMed]
- Song, G.; Du, Q. Structure Characterization and Antitumor Activity of an α β-Glucan Polysaccharide from Auricularia Polytricha. Food Res. Int. 2012, 45, 381–387. [Google Scholar] [CrossRef]
- Wani, S.M.; Gani, A.; Mir, S.A.; Masoodi, F.A.; Khanday, F.A. β-Glucan: A Dual Regulator of Apoptosis and Cell Proliferation. Int. J. Biol. Macromol. 2021, 182, 1229–1237. [Google Scholar] [CrossRef] [PubMed]
- Vetvicka, V.; Richter, J.; Král, V.; Rajnohová Dobiášová, L.; Stiborova, I.; Pohorska, J. Regulation of Hematopoiesis in Cancer Patients: Placebo-Driven, Double-Blind Clinical Trials of Beta-Glucan. J. Tumor 2015, 18, 305–308. [Google Scholar]
- Vetvicka, V.; Vannucci, L.; Sima, P.; Richter, J. Beta Glucan: Supplement or Drug? From Laboratory to Clinical Trials. Molecules 2019, 24, 1251. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Smith, C.; Ashworth, J.; Li, W. Regulation of Inducible Nitric Oxide Synthase by Arabinoxylans with Molecular Characterisation from Wheat Flour in Cultured Human Monocytes. Int. J. Food Sci. Technol. 2018, 53, 1294–1302. [Google Scholar] [CrossRef]
- Akhtar, M.; Tariq, A.F.; Awais, M.M.; Iqbal, Z.; Muhammad, F.; Shahid, M.; Hiszczynska-Sawicka, E. Studies on Wheat Bran Arabinoxylan for Its Immunostimulatory and Protective Effects against Avian Coccidiosis. Carbohydr. Polym. 2012, 90, 333–339. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Wang, J.; Liu, Y.; Sun, B. Wheat Bran Feruloyl Oligosaccharides Modulate the Phase II Detoxifying/Antioxidant Enzymes via Nrf2 Signaling. Int. J. Biol. Macromol. 2015, 74, 150–154. [Google Scholar] [CrossRef]
- Shah, A.; Gani, A.; Masoodi, F.A.; Wani, S.M.; Ashwar, B.A. Structural, Rheological and Nutraceutical Potential of β-Glucan from Barley and Oat. Bioact. Carbohydr. Diet. Fibre 2017, 10, 10–16. [Google Scholar] [CrossRef]
- Paesani, C.; Degano, A.L.; Zalosnik, M.I.; Fabi, J.P.; Pérez, G.T. Enzymatic Modification of Arabinoxylans from Soft and Hard Argentinian Wheat Inhibits the Viability of HCT-116 Cells. Food Res. Int. 2021, 147, 110466. [Google Scholar] [CrossRef]
- Femia, A.P.; Salvadori, M.; Broekaert, W.F.; François, I.E.J.A.; Delcour, J.A.; Courtin, C.M.; Caderni, G. Arabinoxylan-Oligosaccharides (AXOS) Reduce Preneoplastic Lesions in the Colon of Rats Treated with 1,2-Dimethylhydrazine (DMH). Eur. J. Nutr. 2010, 49, 127–132. [Google Scholar] [CrossRef]
- Glei, M.; Hofmann, T.; Küster, K.; Hollmann, J.; Lindhauer, M.G.; Pool-Zobel, B.L. Both Wheat (Triticum Aestivum) Bran Arabinoxylans and Gut Flora-Mediated Fermentation Products Protect Human Colon Cells from Genotoxic Activities of 4-Hydroxynonenal and Hydrogen Peroxide. J. Agric. Food Chem. 2006, 54, 2088–2095. [Google Scholar] [CrossRef]
- Stevenson, L.; Phillips, F.; O’sullivan, K.; Walton, J. Wheat Bran: Its Composition and Benefits to Health, a European Perspective. Int. J. Food Sci. Nutr. 2012, 63, 1001–1013. [Google Scholar] [CrossRef]
- Vauzour, D.; Rodriguez-Mateos, A.; Corona, G.; Oruna-Concha, M.J.; Spencer, J.P.E. Polyphenols and Human Health: Prevention of Disease and Mechanisms of Action. Nutrients 2010, 2, 1106–1131. [Google Scholar] [CrossRef] [PubMed]
- Jakobek, L.; Matić, P. Non-Covalent Dietary Fiber—Polyphenol Interactions and Their Influence on Polyphenol Bioaccessibility. Trends Food Sci. Technol. 2019, 83, 235–247. [Google Scholar] [CrossRef]
- Zhang, L.; Wu, T.; Zhang, Y.; Chen, Y.; Ge, X.; Sui, W.; Zhu, Q.; Geng, J.; Zhang, M. Release of Bound Polyphenols from Wheat Bran Soluble Dietary Fiber during Simulated Gastrointestinal Digestion and Colonic Fermentation in Vitro. Food Chem. 2023, 402, 134111. [Google Scholar] [CrossRef] [PubMed]
- Konishi, Y.; Zhao, Z.; Shimizu, M. Phenolic Acids Are Absorbed from the Rat Stomach with Different Absorption Rates. J. Agric. Food Chem. 2006, 54, 7539–7543. [Google Scholar] [CrossRef]
- Zhang, H.; Sun, S.; Ai, L. Physical Barrier Effects of Dietary Fibers on Lowering Starch Digestibility. Curr. Opin. Food Sci. 2022, 48, 100940. [Google Scholar] [CrossRef]
- Sharma, P.; Hajam, Y.A.; Kumar, R.; Rai, S. Complementary and Alternative Medicine for the Treatment of Diabetes and Associated Complications: A Review on Therapeutic Role of Polyphenols. Phytomed. Plus 2022, 2, 100188. [Google Scholar] [CrossRef]
- Singh, M.; Thrimawithana, T.; Shukla, R.; Adhikari, B. Managing Obesity through Natural Polyphenols: A Review. Future Foods 2020, 1–2, 100002. [Google Scholar] [CrossRef]
- Santhakumar, A.B.; Battino, M.; Alvarez-Suarez, J.M. Dietary Polyphenols: Structures, Bioavailability and Protective Effects against Atherosclerosis. Food Chem. Toxicol. 2018, 113, 49–65. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Tu, Y.-C.; Lian, T.-W.; Hung, J.-T.; Yen, J.-H.; Wu, M.-J. Distinctive Antioxidant and Antiinflammatory Effects of Flavonols. J. Agric. Food Chem. 2006, 54, 9798–9804. [Google Scholar] [CrossRef] [PubMed]
- Brenelli de Paiva, L.B.; Goldbeck, R.; dos Santos, W.D.; Squina, F.M. Ferulic Acid and Derivatives: Molecules with Potential Application in the Pharmaceutical Field. Braz. J. Pharm. Sci. 2013, 49, 395–411. [Google Scholar] [CrossRef]
- Smith, K.N.; Queenan, K.M.; Thomas, W.; Fulcher, R.G.; Slavin, J.L. Physiological Effects of Concentrated Barley Beta-Glucan in Mildly Hypercholesterolemic Adults. J. Am. Coll. Nutr. 2008, 27, 434–440. [Google Scholar] [CrossRef] [PubMed]
- Official Journal of the European Union. Annex Nutrition Claims and Conditions Applying to Them; OPOCE: Luxembourg, 2006; Volume 1924, p. 17. [Google Scholar]
- European Comission. EU Register on Nutrition and Health Claims. 2013. Available online: https://ec.europa.eu/food/food-feed-portal/backend/claims/files/euregister.pdf (accessed on 21 December 2022).
- Food and Drug Administration. Food Labelling; Food and Drug Administration: Silver Spring, ML, USA, 2016. [Google Scholar]
- Moshawih, S.; Abdullah Juperi, R.N.A.; Paneerselvam, G.S.; Ming, L.C.; Liew, K.B.; Goh, B.H.; Al-Worafi, Y.M.; Choo, C.-Y.; Thuraisingam, S.; Goh, H.P.; et al. General Health Benefits and Pharmacological Activities of Triticum Aestivum L. Molecules 2022, 27, 1948. [Google Scholar] [CrossRef] [PubMed]
- Wrigley, C.; Corke, H.; Seetharaman, K.; Faubion, J. Encyclopedia of Food Grains, 2nd ed.; Academic Press: Cambridge, MA, USA, 2016; Volume 1. [Google Scholar]
- Müller-Lissner, S.A. Effect of Wheat Bran on Weight of Stool and Gastrointestinal Transit Time: A Meta Analysis. Br. Med. J. 1988, 296, 615–617. [Google Scholar] [CrossRef]
- Ho, K.-S.; Tan, C.Y.M.; Daud, M.A.M.; Seow-Choen, F. Stopping or Reducing Dietary Fiber Intake Reduces Constipation and Its Associated Symptoms. World J. Gastroenterol. 2012, 18, 4593–4596. [Google Scholar] [CrossRef]
- Gonzalez-Anton, C.; Rico, M.; Sanchez-Rodriguez, E.; Ruiz-Lopez, M.; Gil, A.; Mesa, M. Glycemic Responses, Appetite Ratings and Gastrointestinal Hormone Responses of Most Common Breads Consumed in Spain. A Randomized Control Trial in Healthy Humans. Nutrients 2015, 7, 4033–4053. [Google Scholar] [CrossRef]
- Gómez, M.; Ronda, F.; Blanco, C.; Caballero, P.A.; Apesteguía, A. Effect of Dietary Fibre on Dough Rheology and Bread Quality. Eur. Food Res. Technol. 2003, 216, 51–56. [Google Scholar] [CrossRef]
- Han, H.; Dye, L.; Mackie, A. The Impact of Processing on the Release and Antioxidant Capacity of Ferulic Acid from Wheat: A Systematic Review. Food Res. Int. 2023, 164, 112371. [Google Scholar] [CrossRef]
- Zhang, L.; Gao, W.; Chen, X.; Wang, H. The Effect of Bioprocessing on the Phenolic Acid Composition and Antioxidant Activity of Wheat Bran. Cereal Chem. 2014, 91, 255–261. [Google Scholar] [CrossRef]
- Călinoiu, L.; Vodnar, D. Thermal Processing for the Release of Phenolic Compounds from Wheat and Oat Bran. Biomolecules 2019, 10, 21. [Google Scholar] [CrossRef]
- Tu, J.; Zhao, J.; Liu, G.; Tang, C.; Han, Y.; Cao, X.; Jia, J.; Ji, G.; Xiao, H. Solid State Fermentation by Fomitopsis Pinicola Improves Physicochemical and Functional Properties of Wheat Bran and the Bran-Containing Products. Food Chem. 2020, 328, 127046. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Lei, M.; Samina, N.; Chen, L.; Liu, C.; Yin, T.; Yan, X.; Wu, C.; He, H.; Yi, C. Impact of Lactobacillus Plantarum 423 Fermentation on the Antioxidant Activity and Flavor Properties of Rice Bran and Wheat Bran. Food Chem. 2020, 330, 127156. [Google Scholar] [CrossRef]
- Bartkiene, E.; Jomantaite, I.; Mockus, E.; Ruibys, R.; Baltusnikiene, A.; Santini, A.; Zokaityte, E. The Contribution of Extruded and Fermented Wheat Bran to the Quality Parameters of Wheat Bread, Including the Profile of Volatile Compounds and Their Relationship with Emotions Induced for Consumers. Foods 2021, 10, 2501. [Google Scholar] [CrossRef] [PubMed]
- Reisinger, M.; Tirpanalan, Ö.; Prückler, M.; Huber, F.; Kneifel, W.; Novalin, S. Wheat Bran Biorefinery—A Detailed Investigation on Hydrothermal and Enzymatic Treatment. Bioresour. Technol. 2013, 144, 179–185. [Google Scholar] [CrossRef]
- Chen, Z.; Zha, B.; Wang, L.; Wang, R.; Chen, Z.; Tian, Y. Dissociation of Aleurone Cell Cluster from Wheat Bran by Centrifugal Impact Milling. Food Res. Int. 2013, 54, 63–71. [Google Scholar] [CrossRef]
- Lebert, L.; Buche, F.; Sorin, A.; Aussenac, T. The Wheat Aleurone Layer: Optimisation of Its Benefits and Application to Bakery Products. Foods 2022, 11, 3552. [Google Scholar] [CrossRef]
- Lin, S.; Jin, X.; Gao, J.; Qiu, Z.; Ying, J.; Wang, Y.; Dong, Z.; Zhou, W. Impact of Wheat Bran Micronization on Dough Properties and Bread Quality: Part I—Bran Functionality and Dough Properties. Food Chem. 2021, 353, 129407. [Google Scholar] [CrossRef] [PubMed]
- Ye, J.; Hu, X.; Luo, S.; Liu, W.; Chen, J.; Zeng, Z.; Liu, C. Properties of Starch after Extrusion A Review. Starch–Stärke 2018, 70, 1700110. [Google Scholar] [CrossRef]
- Iwai, K.; Yoshikawa, Y.; Miyoshi, N.; Fukutomi, R.; Asada, K.; Ohashi, N. Effects of Short-Term Intake of Wheat Bran with Different Particle Sizes on the Murine Intestinal Environment. Food Sci. Technol. Res. 2017, 23, 733–742. [Google Scholar] [CrossRef]
- Dziedzic, K.; Szwengiel, A.; Górecka, D.; Gujska, E.; Kaczkowska, J.; Drożdżyńska, A.; Walkowiak, J. Effect of Wheat Dietary Fiber Particle Size during Digestion In Vitro on Bile Acid, Faecal Bacteria and Short-Chain Fatty Acid Content. Plant Foods Hum. Nutr. 2016, 71, 151–157. [Google Scholar] [CrossRef]
- Schutyser, M.A.I.; van der Goot, A.J. The Potential of Dry Fractionation Processes for Sustainable Plant Protein Production. Trends Food Sci. Technol. 2011, 22, 154–164. [Google Scholar] [CrossRef]
- Aktas-Akyildiz, E.; Mattila, O.; Sozer, N.; Poutanen, K.; Koksel, H.; Nordlund, E. Effect of Steam Explosion on Enzymatic Hydrolysis and Baking Quality of Wheat Bran. J. Cereal Sci. 2017, 78, 25–32. [Google Scholar] [CrossRef]
- Wang, T.; Sun, X.; Zhou, Z.; Chen, G. Effects of Microfluidization Process on Physicochemical Properties of Wheat Bran. Food Res. Int. 2012, 48, 742–747. [Google Scholar] [CrossRef]
- Wang, T.; Raddatz, J.; Chen, G. Effects of Microfluidization on Antioxidant Properties of Wheat Bran. J. Cereal Sci. 2013, 58, 380–386. [Google Scholar] [CrossRef]
- Kaur, S.; Sharma, S.; Singh, B.; Dar, B.N. Effect of Extrusion Variables (Temperature, Moisture) on the Antinutrient Components of Cereal Brans. J. Food Sci. Technol. 2015, 52, 1670–1676. [Google Scholar] [CrossRef] [PubMed]
- Gualberto, D.G.; Bergman, C.J.; Kazemzadeh, M.; Weber, C.W. Effect of Extrusion Processing on the Soluble and Insoluble Fiber, and Phytic Acid Contents of Cereal Brans|SpringerLink. Available online: https://link.springer.com/article/10.1023/A:1007941032726 (accessed on 29 October 2021).
- Liu, L.; Zhao, M.; Liu, X.; Zhong, K.; Tong, L.; Zhou, X.; Zhou, S. Effect of Steam Explosion-Assisted Extraction on Phenolic Acid Profiles and Antioxidant Properties of Wheat Bran: Effect of Steam Explosion on Wheat Bran Phenolics. J. Sci. Food Agric. 2016, 96, 3484–3491. [Google Scholar] [CrossRef]
- Bader Ul Ain, H.; Saeed, F.; Khan, M.A.; Niaz, B.; Khan, S.G.; Anjum, F.M.; Tufail, T.; Hussain, S. Comparative Study of Chemical Treatments in Combination with Extrusion for the Partial Conversion of Wheat and Sorghum Insoluble Fiber into Soluble. Food Sci. Nutr. 2019, 7, 2059–2067. [Google Scholar] [CrossRef]
- Yan, J.; Lv, Y.; Ma, S. Wheat Bran Enrichment for Flour Products: Challenges and Solutions. J. Food Process. Preserv. 2022, 46, e16977. [Google Scholar] [CrossRef]
- Park, K.H.; Lee, K.Y.; Lee, H.G. Chemical Composition and Physicochemical Properties of Barley Dietary Fiber by Chemical Modification. Int. J. Biol. Macromol. 2013, 60, 360–365. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.-Y.; Liao, A.-M.; Thakur, K.; Huang, J.-H.; Zhang, J.-G.; Wei, Z.-J. Modification of Wheat Bran Insoluble Dietary Fiber with Carboxymethylation, Complex Enzymatic Hydrolysis and Ultrafine Comminution. Food Chem. 2019, 297, 124983. [Google Scholar] [CrossRef]
- Chakka, V.P.; Zhou, T. Carboxymethylation of Polysaccharides: Synthesis and Bioactivities. Int. J. Biol. Macromol. 2020, 165, 2425–2431. [Google Scholar] [CrossRef] [PubMed]
- Cheng, Y.; Xue, F.; Yu, S.; Du, S.; Yang, Y. Subcritical Water Extraction of Natural Products. Molecules 2021, 26, 4004. [Google Scholar] [CrossRef]
- Malunga, L.N.; Beta, T. Antioxidant Capacity of Arabinoxylan Oligosaccharide Fractions Prepared from Wheat Aleurone Using Trichoderma Viride or Neocallimastix Patriciarum Xylanase. Food Chem. 2015, 167, 311–319. [Google Scholar] [CrossRef]
- Lo, T.C.-T.; Chang, C.A.; Chiu, K.-H.; Tsay, P.-K.; Jen, J.-F. Correlation Evaluation of Antioxidant Properties on the Monosaccharide Components and Glycosyl Linkages of Polysaccharide with Different Measuring Methods. Carbohydr. Polym. 2011, 86, 320–327. [Google Scholar] [CrossRef]
- Li, C.; Wang, L.; Chen, Z.; Li, Y.; Luo, X.; Zhao, F. Ozonolysis of Wheat Bran in Subcritical Water for Enzymatic Saccharification and Polysaccharide Recovery. J. Supercrit. Fluids 2021, 168, 105092. [Google Scholar] [CrossRef]
- Monsalve-Gonzalez, A.; Prakash, A. Bran and Bran Containing Products of Improved Flavor and Methods of Preparation. U.S. Patent 8,053,010, 8 November 2011. [Google Scholar]
- Zhang, L.; Meng, C.; Fu, J.; Lou, J.; Zhang, X.; Gao, W.; Fan, X. Effect of Ozone Treatment on the Chemical and Mechanical Properties of Flax Fibers. Ind. Crops Prod. 2022, 189, 115694. [Google Scholar] [CrossRef]
- Lemeune, S.; Jameel, H.; Chang, H.-M.; Kadla, J.F. Effects of Ozone and Chlorine Dioxide on the Chemical Properties of Cellulose Fibers. J. Appl. Polym. Sci. 2004, 93, 1219–1223. [Google Scholar] [CrossRef]
- Légifrance List of Food Enzymes Permitted in France. Arrêté Du 19 Octobre 2006 Relatif à l’emploi d’auxiliaires Technologiques Dans La Fabrication de Certaines Denrées Alimentaires, Annexe IC. 2020. Available online: legifrance.gouv.fr (accessed on 12 April 2023).
- Zhu, F. Effect of Ozone Treatment on the Quality of Grain Products. Food Chem. 2018, 264, 358–366. [Google Scholar] [CrossRef] [PubMed]



| Nutrient, Substance, Food | Claim | Conditions | Health Relationship | EFSA Opinion Reference/Journal Reference | Commission Regulation |
|---|---|---|---|---|---|
| Wheat bran fibre | Wheat bran fibre contributes to an increase in faecal bulk | The claim may be used only for food which is high in that fibre as referred to in the claim HIGH FIBRE as listed in the Annex to Regulation (EC) No 1924/2006 | Increase in faecal bulk | 2010;8(10):1817 | Commission Regulation (EU) 432/2012 of 16 May 2012 |
| Wheat bran fibre | Wheat bran fibre contributes to an acceleration of intestinal transit | The claim may be used only for food which is high in that fibre as referred to in the claim HIGH FIBRE as listed in the Annex to Regulation (EC) No 1924/2006. In order to bear the claim, information shall be given to the consumer that the claimed effect is obtained with a daily intake of at least 10 g of wheat bran fibre | Reduction in intestinal transit time | 2010;8(10):1817 | Commission Regulation (EU) 432/2012 of 16 May 2012 |
| Arabinoxylan produced from wheat endosperm | Consumption of arabinoxylan as a part of a meal contributes to a reduction of the blood glucose rise after that meal | The claim may be used only for food which contains at least 8 g of arabinoxylan (AX)-rich fibre produced from wheat endosperm (at least 60% AX by weight) per 100 g of available carbohydrates in a quantified portion as part of the meal. In order to bear the claim, information shall be given to the consumer that the beneficial effect is obtained by consuming the arabinoxylan (AX)-rich fibre produced from wheat endosperm as part of the meal. | Reduction of post-prandial glycemic responses | 2011;9(6):2205 | Commission Regulation (EU) 432/2012 of 16 May 2012 |
| Microorganism/Enzyme | Matrix | Objective and Perspectives | Effects in Comparison with Control Sample | References |
|---|---|---|---|---|
| Eurotium cristatum (Fungi) | Wheat bran | To study if E. cristatum has a potential to produce wheat bran food products more nutritional, flavourful and functional. | ↗ Ferulic acid content ↗ SDF ↗ Bindings capacities (WSC, WHC, OBC) ↗ TPC, anthocyanins, phenolic acids ↗ DPPH and ABTS ↗ Pancreatic lipase inhibition activity ↗ phenylethyl alcohol Different flavour | [130] |
| Fomitopsis pinicola (Fungi) | Wheat bran | Evaluate the potential application of F. pinicola to improve the physicochemical and functional properties of wheat bran. | ↗ Polyphenols ↗ Alkylresorcinols ↗ AO activity ↗ Swelling capacity ↗ Protein ↘ Phytic acid | [265] |
| Lactobacillus plantarum 423 (Bacteria) | Rice bran and wheat bran | To study the potential application of rice and wheat bran in health foods and nutraceuticals. | ↗ odour intensity ↗ AO activity | [266] |
| Bacillus species, yeasts, filamentous fungi (Bacteria, fungi, yeast) | Wheat bran | Improving phenolic acid composition and antioxidant activity of wheat bran. | ↗ free phenolic content ↗ DPPH AO activity correlated with TPC | [263] |
| Bacillus sp. TMF-2 (strain which produces several enzymes) | Wheat bran | To produce wheat bran with higher nutritional quality. | ↗ Total phenolic content (×3) ↗ Antioxidant capacity (FRAP ×10) ↗ hydrolytic enzymes ↘ phytic acid | [167] |
| Lactobacillus rhamnosus (Bacteria) | Wheat bran | To study the ability of lactic acid bacteria to modify the overall characteristics of wheat bran as a pre-treatment to potentially enhance its health and sensory properties. | ↗ WEAX ↗ Fruity note at volatile profile ↘ 37% phytic acid | [179] |
| L. plantarum (Mixed acid lactic bacteria) | Wheat bran IDF | To study structure, physiochemical, functional properties and antioxidant activity of wheat bran modified IDF by fermentation in order to provide high-quality functional IDF for food processing in human health management. | ↗ WRC, Oil retention Capacity (ORC), WSC ↗ NIAC, TPC ↘ CAC | [73] |
| L. acidophilus (Bacteria) | ↗ SCAC CEC ↗ NIAC, TPC ↘ CAC | |||
| Yeast and lactic acid bacteria | Wheat bran | To improve the nutritional, physical and flavour properties of wheat bran. | ↗ WEAX ↗ SDF ↗ Alkylresorcinols ↗ Binding/hydration capacities ↗ Phenolic content ≠ flavour ↘ 20% phytic acid | [153] |
| Baker’s yeast | Wheat bran | To compare different treatments and evaluate their effects on wheat bran properties in order to improve its quality. | ↗ WEAX (+46.4%) ↗ SDF ↗ Free Phenolic content ↗ WHC ↗ Vitamin B2 ↘ Vitamin B1 ↘ OHC | [25] |
| Enzymatic treatment with xylanase | Wheat bran | ↗ Folic acid ↘ Vitamin C, B1, B2 ↘ Phytic acid ↗ Bound phenolic content ↘ Free phenolic content↘ DPPH scavenging activity (−16.9%) ↘ WHC, OHC | [25] | |
| Enzymatic hydrolysis with cellulase and xylanase (1:3) | Wheat bran | Improve the value of wheat bran to provide a reference for the development of WB treatment | = WHC (↘ WHC with particle size reduction) ↘ SCAC (and with particle size reduction) ↘ CEC = Phytate content | [142] |
| Snail enzymes | Wheat bran | To modify IDF and SDF from wheat bran in order to improve their functional and physico-chemical properties for potential application in the food industry as a functional ingredient. | ↗ IDF Oil retention capacity ↗ Glucose adsorption ↗ Cholesterol adsorption ↗ Radical scavenging activity (DPPH) Change in microstructure (SEM) Hemicellulose and cellulose degradation | [156] |
| Hydrothermal and enzymatic treatment | Wheat bran | To investigate a wheat bran pre-treatment for it use as a feedstock for biorefineries. | Sugars dissolution Transformation of carbohydrates in free sugars Partial dissolution of hemicellulose | [268] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sztupecki, W.; Rhazi, L.; Depeint, F.; Aussenac, T. Functional and Nutritional Characteristics of Natural or Modified Wheat Bran Non-Starch Polysaccharides: A Literature Review. Foods 2023, 12, 2693. https://doi.org/10.3390/foods12142693
Sztupecki W, Rhazi L, Depeint F, Aussenac T. Functional and Nutritional Characteristics of Natural or Modified Wheat Bran Non-Starch Polysaccharides: A Literature Review. Foods. 2023; 12(14):2693. https://doi.org/10.3390/foods12142693
Chicago/Turabian StyleSztupecki, Wendy, Larbi Rhazi, Flore Depeint, and Thierry Aussenac. 2023. "Functional and Nutritional Characteristics of Natural or Modified Wheat Bran Non-Starch Polysaccharides: A Literature Review" Foods 12, no. 14: 2693. https://doi.org/10.3390/foods12142693
APA StyleSztupecki, W., Rhazi, L., Depeint, F., & Aussenac, T. (2023). Functional and Nutritional Characteristics of Natural or Modified Wheat Bran Non-Starch Polysaccharides: A Literature Review. Foods, 12(14), 2693. https://doi.org/10.3390/foods12142693