
Genome-Wide Comparative Analysis of Lactiplantibacillus pentosus Isolates Autochthonous to Cucumber Fermentation Reveals Subclades of Divergent Ancestry

Abstract
1. Introduction
2. Materials and Methods
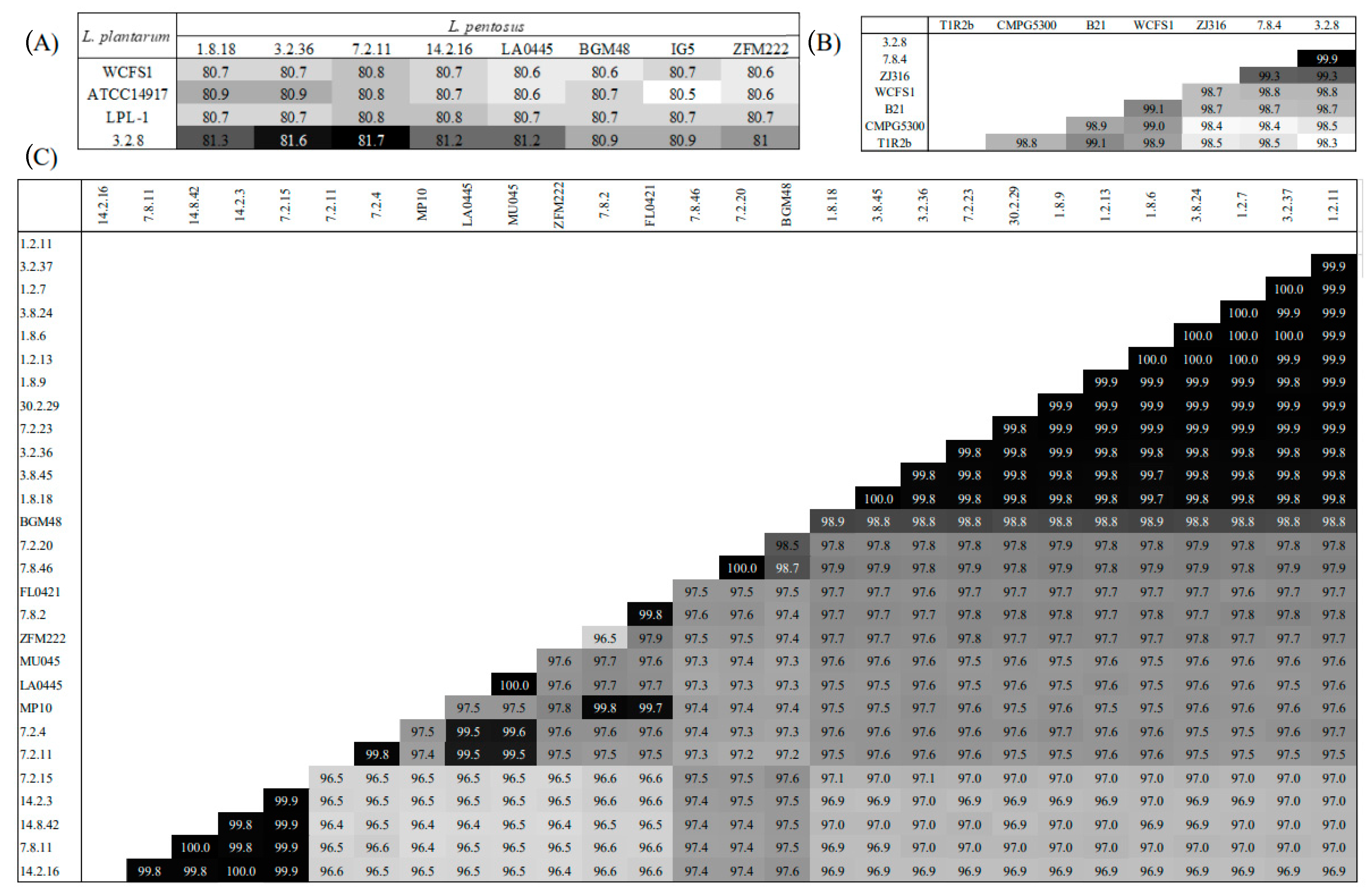
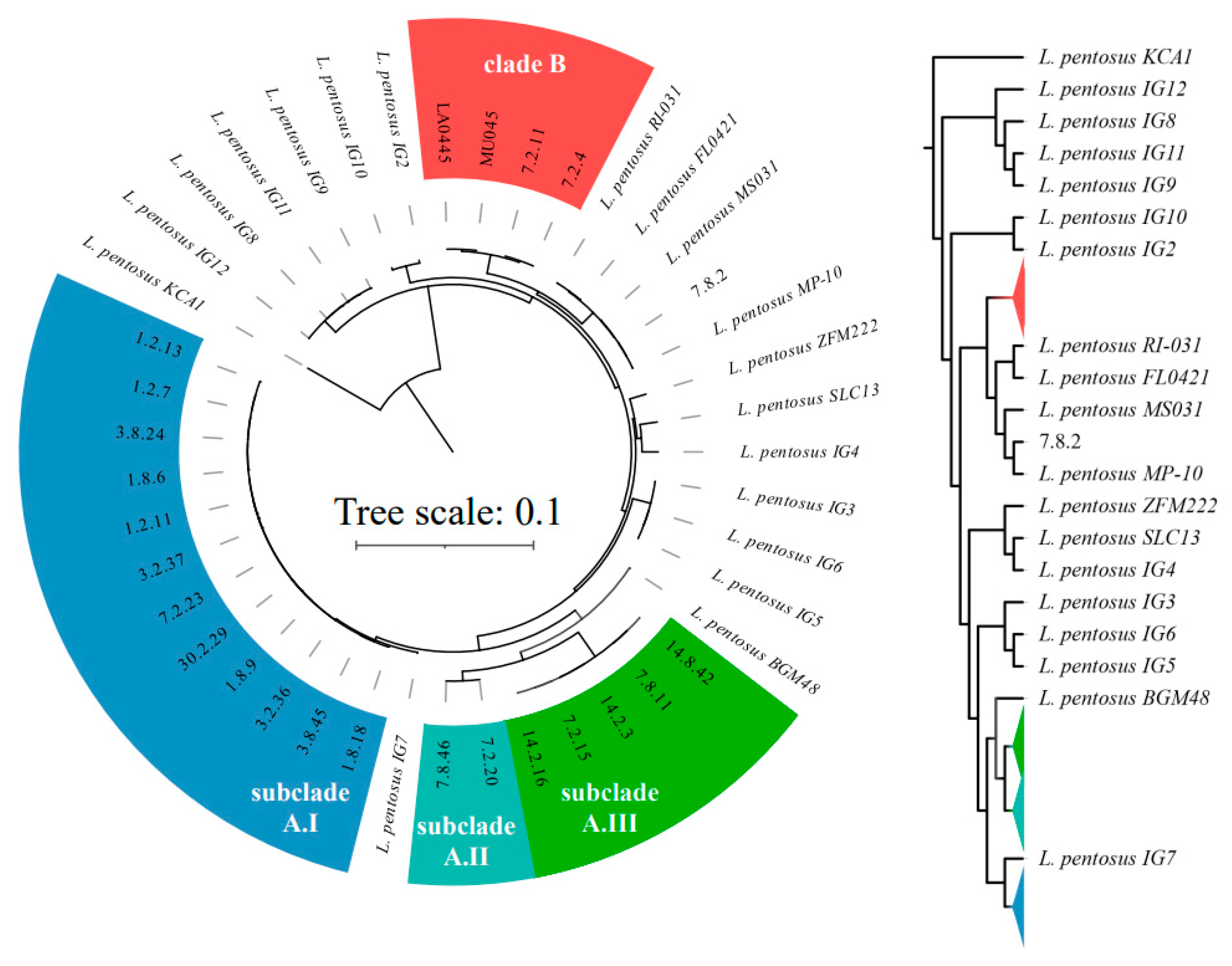
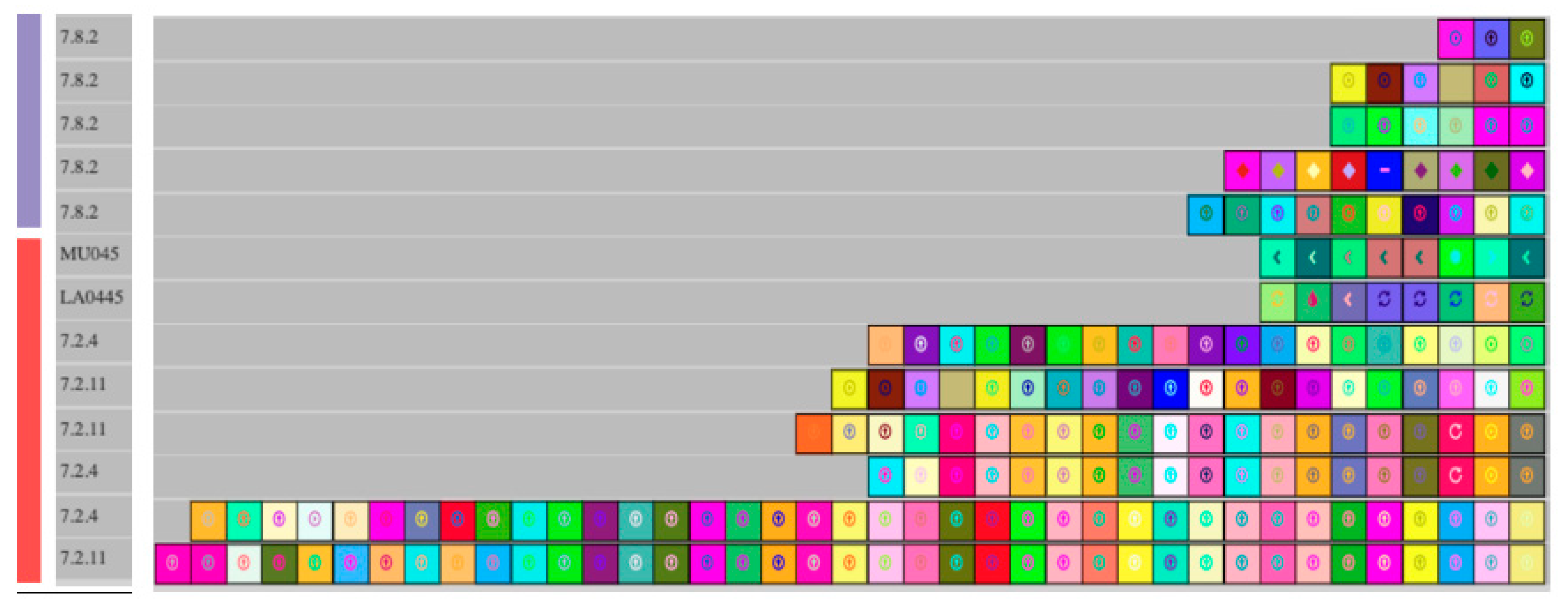
3. Results and Discussion
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Al-Sahlany, S.T.; Niamah, A.K. Bacterial viability, antioxidant stability, antimutagenicity and sensory properties of onion types fermentation by using probiotic starter during storage. Nutr. Food Sci. 2022, 52, 901–916. [Google Scholar] [CrossRef]
- Benítez-Cabello, A.; Bautista-Gallego, J.; Garrido-Fernández, A.; Rantsiou, K.; Cocolin, L.; Jiménez-Díaz, R.; Arroyo-López, F.N. RT-PCR–DGGE Analysis to Elucidate the Dominant Bacterial Species of Industrial Spanish-Style Green Table Olive Fermentations. Front. Microbiol. 2016, 7, 1291. [Google Scholar] [CrossRef]
- Chen, K.-H.; McFEETERS, R.F.; Fleming, H.P. Stability of Mannitol to Lactobacillus plantarum Degradation in Green Beans Fermented with Lactobacillus cellobiosus. J. Food Sci. 1983, 48, 972–974. [Google Scholar] [CrossRef]
- Di Cagno, R.; Surico, R.F.; Siragusa, S.; De Angelis, M.; Paradiso, A.; Minervini, F.; De Gara, L.; Gobbetti, M. Selection and use of autochthonous mixed starter for lactic acid fermentation of carrots, French beans or marrows. Int. J. Food Microbiol. 2008, 127, 220–228. [Google Scholar] [CrossRef] [PubMed]
- Jung, J.Y.; Lee, S.H.; Kim, J.M.; Park, M.S.; Bae, J.W.; Hahn, Y.; Madsen, E.L.; Jeon, C.O. Metagenomic Analysis of Kimchi, a Traditional Korean Fermented Food. Appl. Environ. Microbiol. 2011, 77, 2264–2274. [Google Scholar] [CrossRef]
- Kotzekidou, P.; Roukas, T. Characterization and Distribution of Lactobacilli during Lactic Fermentation of Okra (Hibiscus esculentus). J. Food Sci. 1986, 51, 623–625. [Google Scholar] [CrossRef]
- Lee, Y.G.; Cho, J.-Y.; Kim, Y.-M.; Moon, J.-H. Change in Flavonoid Composition and Antioxidative Activity during Fermentation of Onion (Allium cepa L.) by Leuconostoc mesenteroides with Different Salt Concentrations. J. Food Sci. 2016, 81, C1385–C1393. [Google Scholar] [CrossRef]
- Paramithiotis, S.; Kouretas, K.; Drosinos, E.H. Effect of ripening stage on the development of the microbial community during spontaneous fermentation of green tomatoes. J. Sci. Food Agric. 2014, 94, 1600–1606. [Google Scholar] [CrossRef]
- Pérez-Díaz, I.M.; Dickey, A.; Fitria, R.; Ravishankar, N.; Hayes, J.S.; Campbell, K.; Arritt, F. Modulation of the bacterial population in commercial cucumber fermentations by brining salt type. J. Appl. Microbiol. 2020, 128, 1678–1693. [Google Scholar] [CrossRef]
- Pérez-Díaz, I.M.; Hayes, J.; Medina, E.; Anekella, K.; Daughtry, K.; Dieck, S.; Levi, M.; Price, R.; Butz, N.; Lu, Z.; et al. Reassessment of the succession of lactic acid bacteria in commercial cucumber fermentations and physiological and genomic features associated with their dominance. Food Microbiol. 2017, 63, 217–227. [Google Scholar] [CrossRef]
- Plengvidhya, V.; Breidt, F.; Lu, Z.; Fleming, H.P. DNA Fingerprinting of Lactic Acid Bacteria in Sauerkraut Fermentations. Appl. Environ. Microbiol. 2007, 73, 7697–7702. [Google Scholar] [CrossRef] [PubMed]
- Ashaolu, T.J.; Reale, A. A Holistic Review on Euro-Asian Lactic Acid Bacteria Fermented Cereals and Vegetables. Microorganisms 2020, 8, 1176. [Google Scholar] [CrossRef] [PubMed]
- De Castro, A.; Montaño, A.; Sánchez, A.H.; Rejano, L. Lactic acid fermentation and storage of blanched garlic. Int. J. Food Microbiol. 1998, 39, 205–211. [Google Scholar] [CrossRef] [PubMed]
- Jung, J.H.; Kim, S.J.; Lee, J.Y.; Yoon, S.R.; You, S.Y.; Kim, S.H. Multifunctional properties of Lactobacillus plantarum strains WiKim83 and WiKim87 as a starter culture for fermented food. Food Sci. Nutr. 2019, 7, 2505–2516. [Google Scholar] [CrossRef] [PubMed]
- Lorn, D.; Nguyen, T.-K.-C.; Ho, P.-H.; Tan, R.; Licandro, H.; Waché, Y. Screening of lactic acid bacteria for their potential use as aromatic starters in fermented vegetables. Int. J. Food Microbiol. 2021, 350, 109242. [Google Scholar] [CrossRef] [PubMed]
- Maoloni, A.; Ferrocino, I.; Milanović, V.; Cocolin, L.; Corvaglia, M.R.; Ottaviani, D.; Bartolini, C.; Talevi, G.; Belleggia, L.; Cardinali, F.; et al. The Microbial Diversity of Non-Korean Kimchi as Revealed by Viable Counting and Metataxonomic Sequencing. Foods 2020, 9, 1568. [Google Scholar] [CrossRef]
- Montaño, A.; Sánchez, A.H.; Rejano, L.; de Castro, A. Processing and storage of lye-treated carrots fermented by a mixed starter culture. Int. J. Food Microbiol. 1997, 35, 83–90. [Google Scholar] [CrossRef]
- Oguntoyinbo, F.A.; Cho, G.S.; Trierweiler, B.; Kabisch, J.; Rösch, N.; Neve, H.; Bockelmann, W.; Frommherz, L.; Nielsen, D.S.; Krych, L.; et al. Fermentation of African kale (Brassica carinata) using L. plantarum BFE 5092 and L. fermentum BFE 6620 starter strains. Int. J. Food Microbiol. 2016, 238, 103–112. [Google Scholar] [CrossRef]
- Di Cagno, R.; Surico, R.F.; Paradiso, A.; De Angelis, M.; Salmon, J.-C.; Buchin, S.; De Gara, L.; Gobbetti, M. Effect of autochthonous lactic acid bacteria starters on health-promoting and sensory properties of tomato juices. Int. J. Food Microbiol. 2009, 128, 473–483. [Google Scholar] [CrossRef]
- Cheng, L.; Luo, J.; Li, P.; Yu, H.; Huang, J.; Luo, L. Microbial diversity and flavor formation in onion fermentation. Food Funct. 2014, 5, 2338–2347. [Google Scholar] [CrossRef]
- Zhai, Y.; Pérez-Díaz, I.M. Identification of potential causative agents of the CO2-mediated bloater defect in low salt cucumber fermentation. Int. J. Food Microbiol. 2021, 344, 109115. [Google Scholar] [CrossRef] [PubMed]
- McDonald, L.C.; Fleming, H.P.; Hassan, H.M. Acid Tolerance of Leuconostoc mesenteroides and Lactobacillus plantarum. Appl. Environ. Microbiol. 1990, 56, 2120–2124. [Google Scholar] [CrossRef] [PubMed]
- Ucar, R.A.; Pérez-Díaz, I.M.; Dean, L.L. Content of xylose, trehalose and l-citrulline in cucumber fermentations and utilization of such compounds by certain lactic acid bacteria. Food Microbiol. 2020, 91, 103454. [Google Scholar] [CrossRef] [PubMed]
- Fleming, H.P.; Daeschel, M.A.; McFeeters, R.F.; Pierson, M.D. Butyric Acid Spoilage of Fermented Cucumbers. J. Food Sci. 1989, 54, 636–639. [Google Scholar] [CrossRef]
- Franco, W.; Pérez-Díaz, I.M.; Johanningsmeier, S.D.; McFeeters, R.F. Characteristics of Spoilage-Associated Secondary Cucumber Fermentation. Appl. Environ. Microbiol. 2012, 78, 1273–1284. [Google Scholar] [CrossRef]
- Pérez-Díaz, I.M.; Medina, E.; Page, C.A.; Johanningsmeier, S.D.; Daughtry, K.V.; Moeller, L. Prevention of microbes-induced spoilage in sodium chloride–free cucumber fermentations employing preservatives. J. Food Sci. 2022, 87, 5054–5069. [Google Scholar] [CrossRef]
- Fred, E.B.; Peterson, W.H.; Anderson, J.A. The characteristics of certain pentose-destroying bacteria, especially as it concerns their action on arabinose. J. Biol. Chem. 1921, 48, 385–412. [Google Scholar] [CrossRef]
- Zanoni, P.; Farrow, J.A.E.; Phillips, B.A.; Collins, M.D. Lactobacillus pentosus (Fred, Peterson, and Anderson) sp. nov., nom. rev. Int. J. Syst. Bacteriol. 1987, 37, 339–341. [Google Scholar] [CrossRef]
- Bringel, F.; Curk, M.-C.; Hubert, J.-C. Characterization of Lactobacilli by Southern-Type Hybridization with a Lactobacillus plantarum pyrDFE Probe. Int. J. Syst. Bacteriol. 1996, 46, 588–594. [Google Scholar] [CrossRef]
- Dellaglio, F.; Bottazzi, V.; Vescovo, M. Deoxyribonucleic Acid Homology Among Lactobacillus Species of the Subgenus Streptobacterium Orla-Jensen. Int. J. Syst. Bacteriol. 1975, 25, 160–172. [Google Scholar] [CrossRef]
- Torriani, S.; Felis, G.E.; Dellaglio, F. Differentiation of Lactobacillus plantarum, L. pentosus, and L. paraplantarum by recA Gene Sequence Analysis and Multiplex PCR Assay with recA Gene-Derived Primers. Appl. Environ. Microbiol. 2001, 67, 3450–3454. [Google Scholar] [CrossRef] [PubMed]
- Pérez-Díaz, I.M.; Johanningsmeier, S.D.; Anekella, K.; Pagán-Medina, C.G.; Méndez-Sandoval, L.; Arellano, C.; Price, R.; Daughtry, K.V.; Borges, M.; Bream, C.; et al. Genotypic and phenotypic diversity among Lactobacillus plantarum and Lactobacillus pentosus isolated from industrial scale cucumber fermentations. Food Microbiol. 2021, 94, 103652. [Google Scholar] [CrossRef] [PubMed]
- Figueras, M.J.; Beaz-Hidalgo, R.; Hossain, M.J.; Liles, M.R. Taxonomic Affiliation of New Genomes Should Be Verified Using Average Nucleotide Identity and Multilocus Phylogenetic Analysis. Genome Announc. 2014, 2, e00927-14. [Google Scholar] [CrossRef] [PubMed]
- Richter, M.; Rosselló-Móra, R. Shifting the genomic gold standard for the prokaryotic species definition. Proc. Natl. Acad. Sci. USA 2009, 106, 19126–19131. [Google Scholar] [CrossRef]
- Anekella, K.; Pérez-Díaz, I.M. Characterization of robust Lactobacillus plantarum and Lactobacillus pentosus starter cultures for environmentally friendly low-salt cucumber fermentations. J. Food Sci. 2020, 85, 3487–3497. [Google Scholar] [CrossRef]
- Fleming, H.P.; McFeeters, R.F.; Daeschel, M.A.; Humphries, E.G.; Thompson, R.L. Fermentation of Cucumbers in Anaerobic Tanks. J. Food Sci. 1988, 53, 127–133. [Google Scholar] [CrossRef]
- McDonald, L.C.; Shieh, D.-H.; Fleming, H.P.; McFeeters, R.F.; Thompson, R.L. Evaluation of malolactic-deficient strains of Lactobacillus plantarum for use in cucumber fermentations. Food Microbiol. 1993, 10, 489–499. [Google Scholar] [CrossRef]
- Fideler Moore, J. Generation of Bioactive Peptides and ɣ-Aminobutyric Acid (GABA) during Natural Lactic Acid Fermentation of Cucumber. Ph.D. Thesis, North Carolina State University, Raleigh, NC, USA, 2021. Available online: https://www.lib.ncsu.edu/resolver/1840.20/38713 (accessed on 1 June 2023).
- Page, C.A.; Pérez-Díaz, I.M. Whole-Genome Sequencing and Annotation of Selected Lactobacillales Isolated from Commercial Cucumber Fermentation. Microbiol. Resour. Announc. 2021, 10, e0062521. [Google Scholar] [CrossRef]
- Markowitz, V.M.; Chen, I.-M.A.; Palaniappan, K.; Chu, K.; Szeto, E.; Grechkin, Y.; Ratner, A.; Jacob, B.; Huang, J.; Williams, P.; et al. IMG: The integrated microbial genomes database and comparative analysis system. Nucleic Acids Res. 2012, 40, D115–D122. [Google Scholar] [CrossRef]
- Jain, C.; Rodriguez-R, L.M.; Phillippy, A.M.; Konstantinidis, K.T.; Aluru, S. High throughput ANI analysis of 90K prokaryotic genomes reveals clear species boundaries. Nat. Commun. 2017, 9, 5114. [Google Scholar] [CrossRef]
- Versalovic, J.; Schneider, M.; De Bruijn, F.J.; Lupski, J.R. Genomic fingerprinting of bacteria using repetitive sequence-based polymerase chain reaction. Meth. Mol. Cell. Biol. 1994, 5, 25–40. [Google Scholar]
- Alanjary, M.; Steinke, K.; Ziemert, N. AutoMLST: An automated web server for generating multi-locus species trees highlighting natural product potential. Nucleic Acids Res. 2019, 47, W276–W282. [Google Scholar] [CrossRef] [PubMed]
- Davis, J.J.; Wattam, A.R.; Aziz, R.K.; Brettin, T.; Butler, R.; Butler, R.M.; Chlenski, P.; Conrad, N.; Dickerman, A.; Dietrich, E.M.; et al. The PATRIC Bioinformatics Resource Center: Expanding data and analysis capabilities. Nucleic Acids Res. 2020, 48, D606–D612. [Google Scholar] [CrossRef] [PubMed]
- Stamatakis, A. RAxML version 8: A tool for phylogenetic analysis and post-analysis of large phylogenies. Bioinformatics 2014, 30, 1312–1313. [Google Scholar] [CrossRef] [PubMed]
- Letunic, I.; Bork, P. Interactive Tree Of Life (iTOL) v5: An online tool for phylogenetic tree display and annotation. Nucleic Acids Res. 2021, 49, W293–W296. [Google Scholar] [CrossRef]
- Russel, J.; Pinilla-Redondo, R.; Mayo-Muñoz, D.; Shah, S.A.; Sørensen, S.J. CRISPRCasTyper: Automated Identification, Annotation, and Classification of CRISPR-Cas Loci. CRISPR J. 2020, 3, 462–469. [Google Scholar] [CrossRef]
- Nethery, M.A.; Barrangou, R. Predicting and visualizing features of CRISPR–Cas systems. Methods Enzymol. 2019, 616, 323–362. [Google Scholar] [CrossRef]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic local alignment search tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- Arndt, D.; Grant, J.R.; Marcu, A.; Sajed, T.; Pon, A.; Liang, Y.; Wishart, D.S. PHASTER: A better, faster version of the PHAST phage search tool. Nucleic Acids Res. 2016, 44, W16–W21. [Google Scholar] [CrossRef]
- Zhou, Y.; Liang, Y.; Lynch, K.H.; Dennis, J.J.; Wishart, D.S. PHAST: A Fast Phage Search Tool. Nucleic Acids Res. 2011, 39, W347–W352. [Google Scholar] [CrossRef]
- Davidson, B.E.; Kordias, N.; Dobos, M.; Hillier, A.J. Genomic organization of lactic acid bacteria. Antonie Leeuwenhoek 1996, 70, 161–183. [Google Scholar] [CrossRef] [PubMed]
- Yampolsky, L.Y. Mutation and genome evolution. In Encyclopedia of Evolutionary Biology; Kliman, E.D., Ed.; Academic Press: Cambridge, MA, USA, 2016; pp. 77–83. [Google Scholar] [CrossRef]
- Makarova, K.; Slesarev, A.; Wolf, Y.; Sorokin, A.; Mirkin, B.; Koonin, E.; Pavlov, A.; Pavlova, N.; Karamychev, V.; Polouchine, N.; et al. Comparative genomics of the lactic acid bacteria. Proc. Natl. Acad. Sci. USA 2006, 103, 15611–15616. [Google Scholar] [CrossRef] [PubMed]
- Bobay, L.-M.; Ochman, H. The Evolution of Bacterial Genome Architecture. Front. Genet. 2017, 8, 72. [Google Scholar] [CrossRef]
- Sela, I.; Wolf, Y.I.; Koonin, E.V. Theory of prokaryotic genome evolution. Proc. Natl. Acad. Sci. USA 2016, 113, 11399–11407. [Google Scholar] [CrossRef]
- Westoby, M.; Aagren, D.; Gillings, M.R.; Litchman, E.; Madin, J.S.; Paulsen, I.T.; Tetu, S.G. Cell size, genome size, and maximum growth rate are near-independent dimensions of ecological variation across bacteria and archaea. Ecol. Evol. 2021, 11, 3956–3976. [Google Scholar] [CrossRef] [PubMed]
- Goris, J.; Konstantinidis, K.T.; Klappenbach, J.A.; Coenye, T.; Vandamme, P.; Tiedje, J.M. DNA–DNA hybridization values and their relationship to whole-genome sequence similarities. Int. J. Syst. Evol. Microbiol. 2007, 57, 81–91. [Google Scholar] [CrossRef] [PubMed]
- Daeschel, M.A.; McFeeters, R.F.; Fleming, H.P. Modification of lactic acid bacteria for cucumber fermentations: Elimination of carbon dioxide production from malate. Devel. Ind. Microbiol. 1985, 26, 339–346. [Google Scholar]
- Suo, C.; Yin, Y.; Wang, X.; Lou, X.; Song, D.; Wang, X.; Gu, Q. Effects of Lactobacillus plantarum ZJ316 on pig growth and pork quality. BMC Vet. Res. 2012, 8, 89. [Google Scholar] [CrossRef]
- Parlindungan, E.; May, B.K.; Jones, O.A.H. Metabolic Insights into the Effects of Nutrient Stress on Lactobacillus plantarum B21. Front. Mol. Biosci. 2019, 6, 75. [Google Scholar] [CrossRef]
- Golneshin, A.; Gor, M.-C.; Williamson, N.; Vezina, B.; Van, T.T.H.; May, B.K.; Smith, A.T. Discovery and characterisation of circular bacteriocin plantacyclin B21AG from Lactiplantibacillus plantarum B21. Heliyon 2020, 6, e04715. [Google Scholar] [CrossRef]
- Barrangou, R.; Dudley, E.G. CRISPR-Based Typing and Next-Generation Tracking Technologies. Annu. Rev. Food Sci. Technol. 2016, 7, 395–411. [Google Scholar] [CrossRef]
- Horvath, P.; Romero, D.A.; Coûté-Monvoisin, A.C.; Richards, M.; Deveau, H.; Moineau, S.; Boyaval, P.; Fremaux, C.; Barrangou, R. Diversity, Activity, and Evolution of CRISPR Loci in Streptococcus thermophilus. J. Bacteriol. 2008, 190, 1401–1412. [Google Scholar] [CrossRef]
- Shariat, N.; DiMarzio, M.J.; Yin, S.; Dettinger, L.; Sandt, C.H.; Lute, J.R.; Barrangou, R.; Dudley, E.G. The combination of CRISPR-MVLST and PFGE provides increased discriminatory power for differentiating human clinical isolates of Salmonella enterica subsp. enterica serovar Enteritidis. Food Microbiol. 2013, 34, 164–173. [Google Scholar] [CrossRef] [PubMed]
- Toro, M.; Cao, G.; Ju, W.; Allard, M.; Barrangou, R.; Zhao, S.; Brown, E.; Meng, J. Association of Clustered Regularly Interspaced Short Palindromic Repeat (CRISPR) Elements with Specific Serotypes and Virulence Potential of Shiga Toxin-Producing Escherichia coli. Appl. Environ. Microbiol. 2014, 80, 1411–1420. [Google Scholar] [CrossRef] [PubMed]
- Barrangou, R.; Fremaux, C.; Deveau, H.; Richards, M.; Boyaval, P.; Moineau, S.; Romero, D.A.; Horvath, P. CRISPR Provides Acquired Resistance Against Viruses in Prokaryotes. Science 2007, 315, 1709–1712. [Google Scholar] [CrossRef] [PubMed]
- Lu, Z.; Pérez-Díaz, I.M.; Hayes, J.S.; Breidt, F. Bacteriophage Ecology in a Commercial Cucumber Fermentation. Appl. Environ. Microbiol. 2012, 78, 8571–8578. [Google Scholar] [CrossRef] [PubMed]
- Lu, Z.; Breidt, F.; Plengvidhya, V.; Fleming, H.P. Bacteriophage Ecology in Commercial Sauerkraut Fermentations. Appl. Environ. Microbiol. 2003, 69, 3192–3202. [Google Scholar] [CrossRef]
- Ramisetty, B.C.M.; Sudhakari, P.A. Bacterial “Grounded” Prophages: Hotspots for Genetic Renovation and Innovation. Front. Genet. 2019, 10, 65. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain | Accession Number | Strain | Accession Number |
|---|---|---|---|
| Lactiplantibacillus plantarum | |||
| CMPG5300 | GCA_000762955.1 | JDM1 | GCA_000023085.1 |
| B21 | GCA_000931425.2 | DOMLa | GCA_000604105.1 |
| WCFS1 | GCA_000203855. | 16 | GCA_000412205.1 |
| ZJ316 | GCA_000338115.2 | P-8 | GCA_000392485.2 |
| ATCC14917 | GCA_000143745.1 | 7.8.4 | GCA_018993285.1 |
| LPL-1 | GCA_002205775.2 | 3.2.8 | GCA_018991195.1 |
| ST-III | GCA_000148815.2 | T1R2b | GCA_018993325.1 |
| CGMCC1.557 | GCA_001272315.2 | ||
| Lactiplantibacillus pentosus | |||
| BGM48 | GCA_002850015.1 | 1.8.6 | GCA_018991355.1 |
| FL0421 | GCA_001188985.1 | 1.8.9 | GCA_018993945.1 |
| ZFM222 | GCA_003627295.1 | 1.8.18 | GCA_018993635.1 |
| MP-10 | GCA_900092635.1 | 3.2.37 | GCA_018993495.1 |
| KCA1 | GCA_000271445.1 | 3.2.36 | GCA_018993725.1 |
| RI-031 | GCA_002751855.1 | 3.8.24 | GCA_018993345.1 |
| MS031 | GCA_016804305.1 | 3.8.45 | GCA_018991565.1 |
| SLC13 | GCA_002211885.1 | 7.2.4 | GCA_018991185.1 |
| IG2 | GCA_002993465.1 | 7.2.11 | GCA_018991375.1 |
| IG3 | GCA_002993425.1 | 7.2.15 | GCA_018993445.1 |
| IG4 | GCA_002993385.1 | 7.2.20 | GCA_018993665.1 |
| IG5 | GCA_002993435.1 | 7.2.23 | GCA_018993485.1 |
| IG6 | GCA_002993485.1 | 7.8.2 | GCA_018991535.1 |
| IG7 | GCA_002993395.1 | 7.8.11 | GCA_018993275.1 |
| IG8 | GCA_003702625.1 | 7.8.46 | GCA_018991465.1 |
| IG9 | GCA_003702665.1 | 14.8.42 | GCA_018993525.1 |
| IG10 | GCA_003702635.1 | 14.2.3 | GCA_018991425.1 |
| IG11 | GCA_003702605.1 | 14.2.16 | GCA_018993625.1 |
| IG12 | GCA_003702565.1 | 30.2.29 | GCA_018991255.1 |
| 1.2.7 | GCA_018991365.1 | LA0445 | GCA_018993965.1 |
| 1.2.11 | GCA_018991285.1 | MU045 | GCA_018993585.1 |
| 1.2.13 | GCA_018991345.1 | ||
| Weissella spp. | Leuconostoc spp. | ||
| W. paramesenteroides 3.2.24 | GCA_018991665.1 | Leu. citreum 3.8.12 | GCA_018991475.1 |
| W. cibaria 7.8.34 | GCA_025770435.1 | Leu. lactis 1.2.50 | GCA_018993865.1 |
| W. cibaria 3.8.44 | GCA_018993825.1 | Leu. fallax 1.2.22 | GCA_018993745.1 |
| W. sagaensis 1.2.50 | GCA_018993865.1 | Leu. mesenteroides 1.2.47 | GCA_018991785.1 |
| Pediococcus spp. | Lactococcus spp. | ||
| P. ethanolidurans 90.8.30 | GCA_018993685.1 | Lc. lactis LA0312 | GCA_018991605.1 |
| P. damnosus DSM 20331 | GCA_001437255.1 | Lc. lactis 1.8.12 | GCA_018991645.1 |
| Levilactobacillus spp. | |||
| Lev. brevis 14.2.24 | GCA_018993885.1 | Lactiplantibacillus paraplantarum L-ZS9 | GCA_001443645.1 |
| Lev. brevis 14.2.10 | GCA_018991505.1 | ||
| Lev. brevis 3.2.41 | GCA_018991675.1 | ||
| Genomic DNA Sequence Identification | ||||||||
|---|---|---|---|---|---|---|---|---|
| L. plantarum | L. pentosus | |||||||
| 3.2.8 | 7.8.4 | T1R2b | Subclade A.I | Subclade A.II | Subclade A.III | Clade B | 7.8.2 | |
| CRISPR loci | 1 | 1 | 0 | 4 ± 1 | 4 | 4 ± 1 | 2 ± 1 | 5 |
| Type I-E | 0 | 0 | 0 | 2 | 3 | 2 | 2 ± 1 | 4 |
| Type II-A | 1 | 1 | 0 | 2 ± 1 | 1 | 1 ± 1 | 0 | 1 |
| Closest Homolog Identity (>95%) | L. plantarum Isolates Autochthonous to Cucumber Fermentation | |||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 3.2.8 | 7.8.4 | T1R2b | ||||||||||||||||||||||
| Siphoviridae sp. ctu0P1 | X | |||||||||||||||||||||||
| Siphoviridae sp. ctyPQ2 | X | |||||||||||||||||||||||
| Siphoviridae sp. ct97I3 | X | |||||||||||||||||||||||
| Siphoviridae sp. ctk5O4 | ||||||||||||||||||||||||
| Siphoviridae sp. ctUZT2 | X | |||||||||||||||||||||||
| Closest Homolog Identity (>95%) | L. pentosus Isolates Autochthonous to Cucumber Fermentation | |||||||||||||||||||||||
| Subclade A.I | Subclade A.II | Subclade A.III | Clade B | |||||||||||||||||||||
| 1.2.7 | 1.2.13 | 3.8.24 | 1.8.6 | 1.2.11 | 3.2.37 | 30.2.29 | 1.8.9 | 7.2.23 | 3.2.36 | 3.8.45 | 1.8.18 | 7.8.46 | 7.2.20 | 14.2.3 | 7.2.15 | 14.2.16 | 14.8.42 | 7.8.11 | 7.2.11 | 7.2.4 | LA0445 | MU0045 | 7.8.2 | |
| Siphoviridae ctu0P1 | X | X | X | X | X | X | ||||||||||||||||||
| Siphoviridae ctyPQ2 | ||||||||||||||||||||||||
| Siphoviridae ct97I3 | ||||||||||||||||||||||||
| Siphoviridae ctk5O4 | X | |||||||||||||||||||||||
| Siphoviridae ctUZT2 | ||||||||||||||||||||||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Page, C.A.; Pérez-Díaz, I.M.; Pan, M.; Barrangou, R. Genome-Wide Comparative Analysis of Lactiplantibacillus pentosus Isolates Autochthonous to Cucumber Fermentation Reveals Subclades of Divergent Ancestry. Foods 2023, 12, 2455. https://doi.org/10.3390/foods12132455
Page CA, Pérez-Díaz IM, Pan M, Barrangou R. Genome-Wide Comparative Analysis of Lactiplantibacillus pentosus Isolates Autochthonous to Cucumber Fermentation Reveals Subclades of Divergent Ancestry. Foods. 2023; 12(13):2455. https://doi.org/10.3390/foods12132455
Chicago/Turabian StylePage, Clinton A., Ilenys M. Pérez-Díaz, Meichen Pan, and Rodolphe Barrangou. 2023. "Genome-Wide Comparative Analysis of Lactiplantibacillus pentosus Isolates Autochthonous to Cucumber Fermentation Reveals Subclades of Divergent Ancestry" Foods 12, no. 13: 2455. https://doi.org/10.3390/foods12132455
APA StylePage, C. A., Pérez-Díaz, I. M., Pan, M., & Barrangou, R. (2023). Genome-Wide Comparative Analysis of Lactiplantibacillus pentosus Isolates Autochthonous to Cucumber Fermentation Reveals Subclades of Divergent Ancestry. Foods, 12(13), 2455. https://doi.org/10.3390/foods12132455