
Cryoprotective Effects of Carrageenan Oligosaccharides on Crayfish (Procambarus clarkii) during Superchilling

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Crayfish Samples and Treatments
2.2. Determination of pH
2.3. Determination of TVB-N
2.4. Microbiological Analysis
2.5. Determination of Thawing Loss
2.6. Determination of Water Holding Capacity
2.7. Low Field 1H Nuclear Magnetic Resonance (LF 1H NMR) Spectroscopy
2.8. Extraction of Myofibrillar Protein
2.9. Determination of Total Sulfhydryl Content
2.10. Determination of Disulfide Bond Content
2.11. Determination of Carbonyl Content
2.12. Determination of Surface Hydrophobicity
2.13. Sodium Dodecyl Sulfate-Polyacrylamide Gel Electrophoresis (SDS-PAGE) Analysis
2.14. Statistical Analysis
3. Results and Discussion
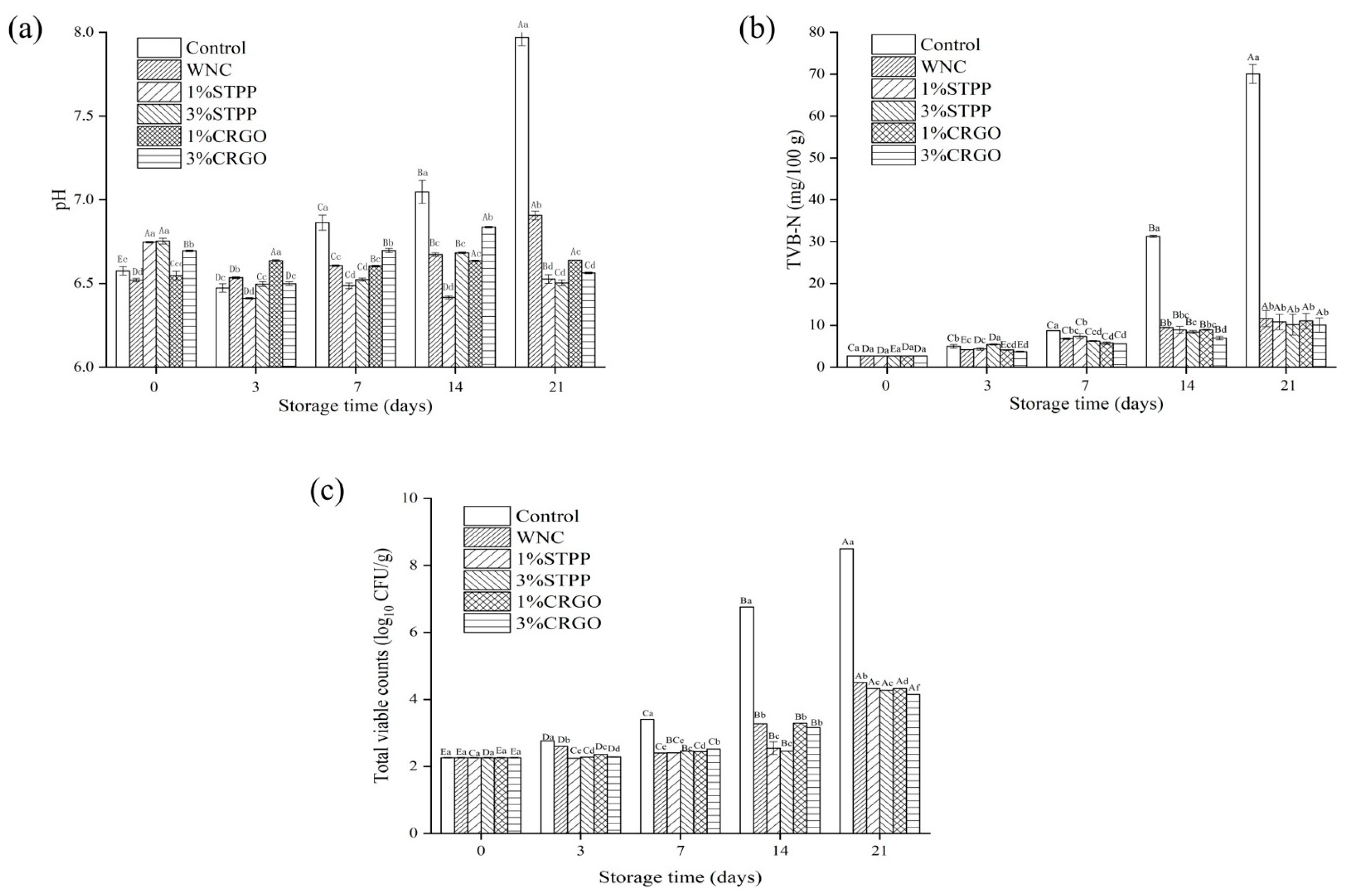
3.1. The pH, TVB-N, and TVC Analysis
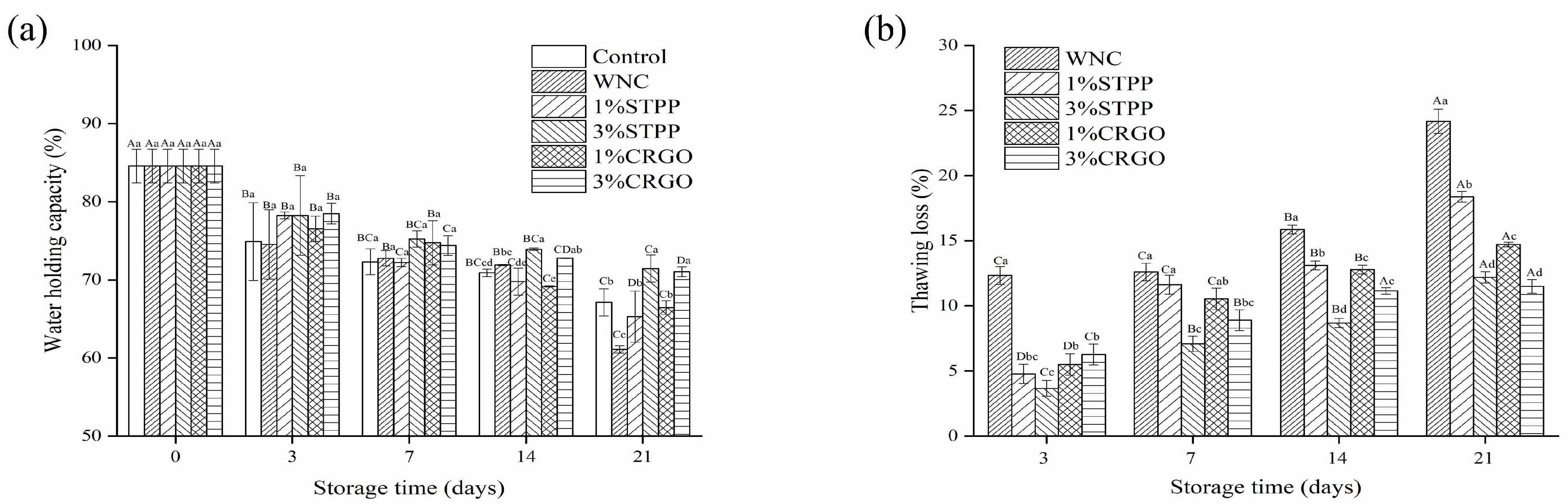
3.2. WHC and Thawing Loss Analysis
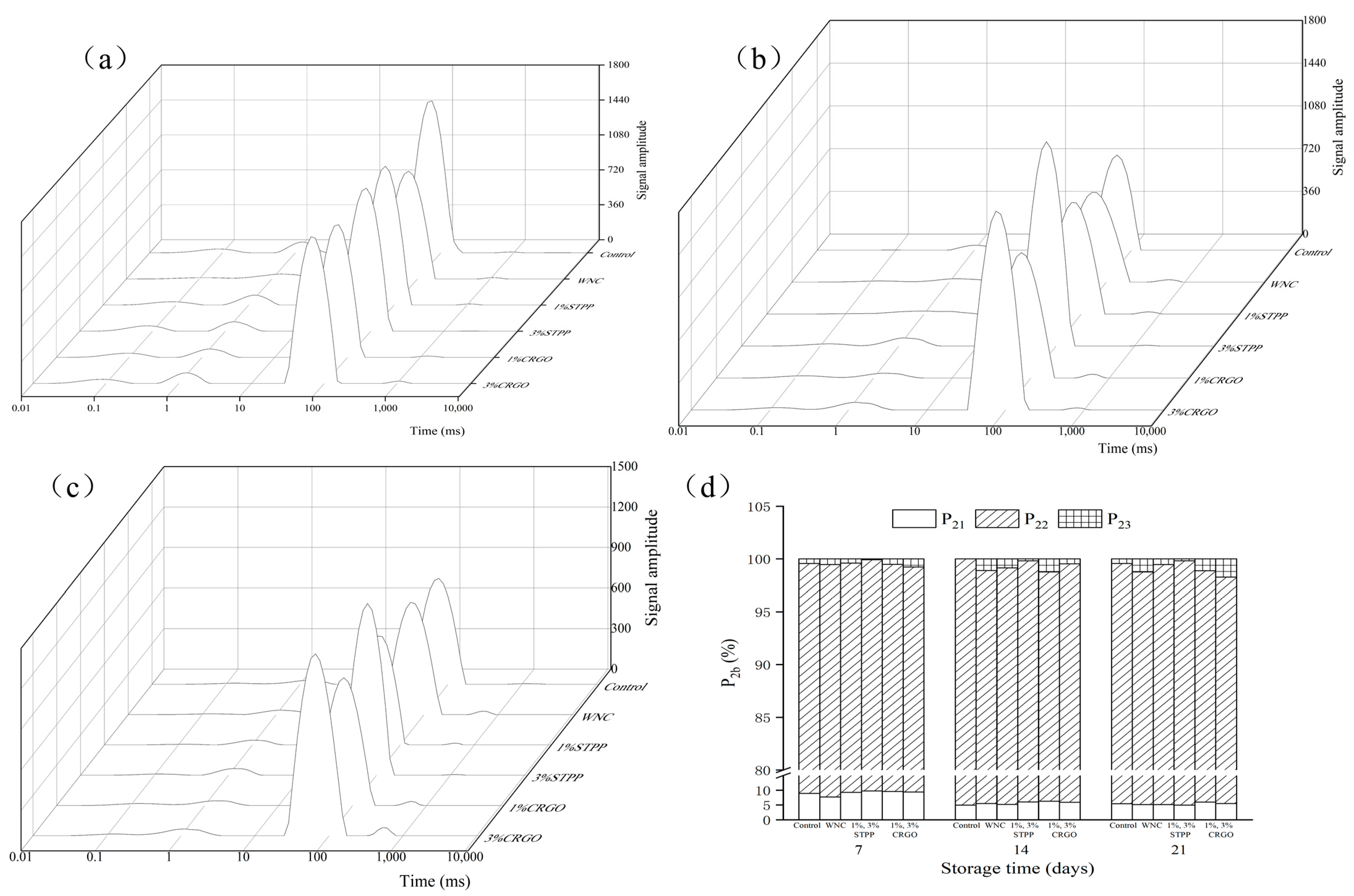
3.3. Water Mobility and Redistribution
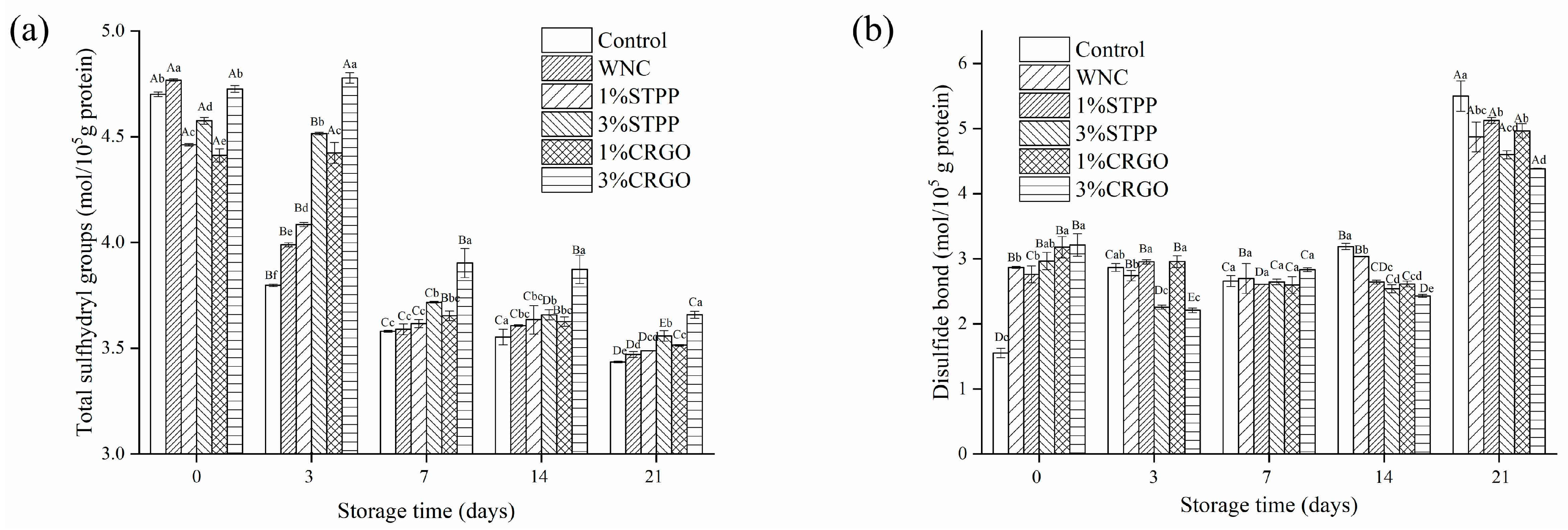
3.4. Total Sulfhydryl and Disulfide Bond Analysis
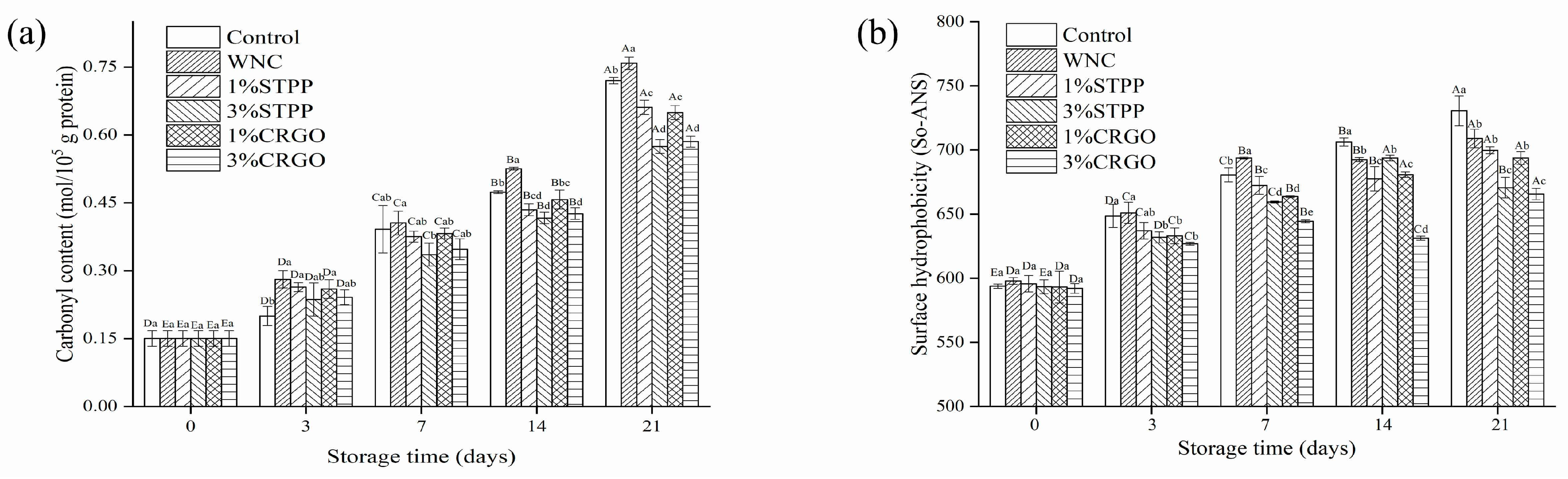
3.5. Carbonyl Content and Surface Hydrophobicity Analysis
3.6. SDS-PAGE
4. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Liu, D.; Liang, L.; Xia, W.; Regenstein, J.; Zhou, P. Biochemical and physical changes of grass carp (Ctenopharyngodon idella) fillets stored at −3 and 0 °C. Food Chem. 2013, 140, 105–114. [Google Scholar] [CrossRef] [PubMed]
- Pomponio, L.; Ruiz-Carrascal, J. Oxidative deterioration of pork during superchilling storage. J. Sci. Food Agric. 2017, 97, 5211–5215. [Google Scholar] [CrossRef] [PubMed]
- Qin, L.; Wu, Y.; Chen, J.; Xia, W.; Liao, E.; Wang, H. Effects of superchilling on quality of crayfish (Procambarus clarkii): Water migration, biogenic amines accumulation, and nucleotides catabolism. Int. J. Food Sci. Technol. 2021, 57, 506–515. [Google Scholar] [CrossRef]
- Hong, H.; Regenstein, J.; Luo, Y. The importance of ATP-related compounds for the freshness and flavor of post-mortem fish and shellfish muscle: A review. Crit. Rev. Food Sci. Nutr. 2017, 57, 1787–1798. [Google Scholar] [CrossRef] [PubMed]
- Etemadian, Y.; Shabanpour, B.; Mahoonak, A.; Shabani, A.; Alami, M. Cryoprotective effects of polyphosphates on Rutilus frisii kutum fillets during ice storage. Food Chem. 2011, 129, 1544–1551. [Google Scholar] [CrossRef]
- Ying, X.; Wu, Q.; Shui, S.; Zhang, B.; Benjakul, S. Insights into the similarities and differences of whiteleg shrimp pre-soaked with sodium tripolyphosphate and sodium trimetaphosphate during frozen storage. Food Chem. 2021, 348, 129134. [Google Scholar] [CrossRef]
- Sekercioglu, N.; Veroniki, A.; Thabane, L.; Busse, J.; Akhtar-Danesh, N.; Iorio, A.; Lopes, L.; Guyatt, G. Effects of different phosphate lowering strategies in patients with CKD on laboratory outcomes: A systematic review and NMA. PLoS ONE 2017, 12, e0171028. [Google Scholar] [CrossRef]
- Zhang, B.; Cao, H.; Lin, H.; Deng, S.; Wu, H. Insights into ice-growth inhibition by trehalose and alginate oligosaccharides in peeled Pacific white shrimp (Litopenaeus vannamei) during frozen storage. Food Chem. 2019, 278, 482–490. [Google Scholar] [CrossRef]
- Chouljenko, A.; Chotiko, A.; Bonilla, F.; Moncada, M.; Reyes, V.; Sathivel, S. Effects of vacuum tumbling with chitosan nanoparticles on the quality characteristics of cryogenically frozen shrimp. LWT 2017, 75, 114–123. [Google Scholar] [CrossRef]
- Xiong, G.; Cheng, W.; Ye, L.; Du, X.; Zhou, M.; Lin, R.; Geng, S.; Chen, M.; Corke, H.; Cai, Y. Effects of konjac glucomannan on physicochemical properties of myofibrillar protein and surimi gels from grass carp (Ctenopharyngodon idella). Food Chem. 2009, 116, 413–418. [Google Scholar] [CrossRef]
- Zhang, B.; Hao, G.; Cao, H.; Tang, H.; Zhang, Y.; Deng, S. The cryoprotectant effect of xylooligosaccharides on denaturation of peeled shrimp (Litopenaeus vannamei) protein during frozen storage. Food Hydrocoll. 2018, 77, 228–237. [Google Scholar] [CrossRef]
- Sun, Y.; Yang, B.; Wu, Y.; Liu, Y.; Gu, X.; Zhang, H.; Wang, C.; Cao, H.; Huang, L.; Wang, Z. Structural characterization and antioxidant activities of kappa-carrageenan oligosaccharides degraded by different methods. Food Chem. 2015, 178, 311–318. [Google Scholar] [CrossRef] [PubMed]
- Shui, S.; Qi, H.; Shaimaa, H.; Aubourg, S.; Zhang, B. Kappa-carrageenan and its oligosaccharides maintain the physicochemical properties of myofibrillar proteins in shrimp mud (Xia-Hua) during frozen storage. J. Food Sci. 2021, 86, 140–148. [Google Scholar] [CrossRef] [PubMed]
- Zhang, B.; Fang, C.; Hao, G.; Zhang, Y. Effect of kappa-carrageenan oligosaccharides on myofibrillar protein oxidation in peeled shrimp (Litopenaeus vannamei) during long-term frozen storage. Food Chem. 2018, 245, 254–261. [Google Scholar] [CrossRef]
- Huang, S.; Xiong, Y.; Zou, Y.; Dong, Q.; Ding, F.; Liu, X.; Li, H. A novel colorimetric indicator based on agar incorporated with Arnebia euchroma root extracts for monitoring fish freshness. Food Hydrocoll. 2019, 90, 198–205. [Google Scholar] [CrossRef]
- Yu, D.; Xu, Y.; Jiang, Q.; Yang, F.; Xia, W. Freshness assessment of grass carp (Ctenopharyngodon idellus) fillets during stroage at 4 °C by physicochemical, microbiological and sensorial evaluations. J. Food Saf. 2017, 37, e12305. [Google Scholar] [CrossRef]
- Ma, L.; Zhang, B.; Deng, S.; Xie, C. Comparison of the cryoprotective effects of trehalose, alginate, and its oligosaccharides on peeled shrimp (Litopenaeus vannamei) during frozen storage. J. Food Sci. 2015, 80, C540–C546. [Google Scholar] [CrossRef]
- Gao, W.; Hou, R.; Zeng, X. Synergistic effects of ultrasound and soluble soybean polysaccharide on frozen surimi from grass carp. J. Food Eng. 2019, 240, 1–8. [Google Scholar] [CrossRef]
- Li, D.; Jia, S.; Zhang, L.; Li, Q.; Pan, J.; Zhu, B.; Prinyawiwatkul, W.; Luo, Y. Post-thawing quality changes of common carp (Cyprinus carpio) cubes treated by high voltage electrostatic field (HVEF) during chilled storage. Innov. Food Sci. Emerg. Technol. 2017, 42, 25–32. [Google Scholar] [CrossRef]
- Jiang, S.; Zhao, D.; Nian, Y.; Wu, J.; Zhang, M.; Li, Q.; Li, C. Ultrasonic treatment increased functional properties and in vitro digestion of actomyosin complex during meat storage. Food Chem. 2021, 352, 129398. [Google Scholar] [CrossRef] [PubMed]
- Riebroy, S.; Benjakul, S.; Visessanguan, W.; Erikson, U.; Rustad, T. Acid-induced gelation of natural actomyosin from Atlantic cod (Gadus morhua) and burbot (Lota lota). Food Hydrocoll. 2009, 23, 26–39. [Google Scholar] [CrossRef]
- Kong, B.; Guo, Y.; Xia, X.; Liu, Q.; Li, Y.; Chen, H. Cryoprotectants reduce protein oxidation and structure deterioration induced by freeze-thaw cycles in common carp (Cyprinus carpio) Surimi. Food Biophys. 2013, 8, 104–111. [Google Scholar] [CrossRef]
- Li, Y.; Kong, B.; Xia, X.; Liu, Q.; Li, P. Inhibition of frozen storage-induced oxidation and structural changes in myofibril of common carp (Cyprinus carpio) surimi by cryoprotectant and hydrolysed whey protein addition. Int. J. Food Sci. Technol. 2013, 48, 1916–1923. [Google Scholar] [CrossRef]
- Zhang, Z.; Xiong, Z.; Walayat, N.; Lorenzo, J.; Liu, J.; Nawaz, A.; Xiong, H. Influence of the mixture of carrageenan oligosaccharides and egg white protein on the gelation properties of culter alburnus myofibrillar protein under repeated freezing-thawing cycles. Antioxidants 2022, 11, 32. [Google Scholar] [CrossRef] [PubMed]
- Qi, X.; Liao, E.; Zhao, K.; Regenstein, J.; Mao, X. Multi-stage countercurrent process for extracting protein from Antarctic Krill (Euphausia superba). J. Food Sci. Technol. 2018, 55, 4450–4457. [Google Scholar] [CrossRef]
- Nirmal, N.; Benjakul, S. Retardation of quality changes of Pacific white shrimp by green tea extract treatment and modified atmosphere packaging during refrigerated storage. Int. J. Food Microbiol. 2011, 149, 247–253. [Google Scholar] [CrossRef]
- Dong, H.; Ling, Z.; Zhang, X.; Zhang, X.; Ramaswamy, S.; Xu, F. Smart colorimetric sensing films with high mechanical strength and hydrophobic properties for visual monitoring of shrimp and pork freshness. Sens. Actuators B Chem. 2020, 309, 127752. [Google Scholar] [CrossRef]
- Na, S.; Kim, J.; Jang, H.; Park, H.; Oh, S. Shelf life extension of Pacific white shrimp (Litopenaeus vannamei) using chitosan and ε-polylysine during cold storage. Int. J. Biol. Macromol. 2018, 115, 1103–1108. [Google Scholar] [CrossRef] [PubMed]
- Nirmal, N.; Benjakul, S. Effect of ferulic acid on inhibition of polyphenoloxidase and quality changes of Pacific white shrimp (Litopenaeus vannamei) during iced storage. Food Chem. 2009, 116, 323–331. [Google Scholar] [CrossRef]
- Li, Y.; Yang, Z.; Li, J. Shelf-life extension of Pacific white shrimp using algae extracts during refrigerated storage. J. Sci. Food Agric. 2017, 97, 291–298. [Google Scholar] [CrossRef]
- Thangavelu, K.; Kerry, J.; Tiwari, B.; McDonnell, C. Novel processing technologies and ingredient strategies for the reduction of phosphate additives in processed meat. Trends Food Sci. Technol. 2019, 94, 43–53. [Google Scholar] [CrossRef]
- Sun, Y.; Cui, X.; Duan, M.; Ai, C.; Song, S.; Chen, X. In vitro fermentation of κ-carrageenan oligosaccharides by human gut microbiota and its inflammatory effect on HT29 cells. J. Funct. Foods 2019, 59, 80–91. [Google Scholar] [CrossRef]
- Olsson, G.; Ofstad, R.; Lødemel, J.; Olsen, R. Changes in water-holding capacity of halibut muscle during cold storage. LWT 2003, 36, 771–778. [Google Scholar] [CrossRef]
- Kilinc, B.; Cakli, S.; Dincer, T.; Cadun, A. Effects of phosphates treatment on the quality of frozen-thawed fish species. J. Muscle Foods 2009, 20, 377–391. [Google Scholar] [CrossRef]
- Chantarasuwan, C.; Benjakul, S.; Visessanguan, W. The effects of sodium bicarbonate on conformational changes of natural actomyosin from Pacific white shrimp (Litopenaeus vannamei). Food Chem. 2011, 129, 1636–1643. [Google Scholar] [CrossRef]
- Xia, X.; Kong, B.; Liu, Q.; Liu, J. Physicochemical change and protein oxidation in porcine longissimus dorsi as influenced by different freeze-thaw cycles. Meat Sci. 2009, 83, 239–245. [Google Scholar] [CrossRef] [PubMed]
- Zhang, T.; Xue, Y.; Li, Z.; Wang, Y.; Yang, W.; Xue, C. Effects of ozone-induced oxidation on the physicochemical properties of myofibrillar proteins recovered from bighead carp (Hypophthalmichthys nobilis). Food Bioproc. Tech. 2014, 8, 181–190. [Google Scholar] [CrossRef]
- Zhang, B.; Yan, H.; Su, L.; Chen, X. Kappa-carrageenan oligosaccharides retard the progression of protein and lipid oxidation in mackerel (Scomber japonicus) fillets during frozen storage. RSC Adv. 2020, 10, 20827–20836. [Google Scholar] [CrossRef]
- Yu, D.; Regenstein, J.; Zang, J.; Xia, W.; Xu, Y.; Jiang, Q.; Yang, F. Inhibitory effects of chitosan-based coatings on endogenous enzyme activities, proteolytic degradation and texture softening of grass carp (Ctenopharyngodon idellus) fillets stored at 4 °C. Food Chem. 2018, 262, 1–6. [Google Scholar] [CrossRef]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liao, E.; Wu, Y.; Pan, Y.; Zhang, Y.; Zhang, P.; Chen, J. Cryoprotective Effects of Carrageenan Oligosaccharides on Crayfish (Procambarus clarkii) during Superchilling. Foods 2023, 12, 2258. https://doi.org/10.3390/foods12112258
Liao E, Wu Y, Pan Y, Zhang Y, Zhang P, Chen J. Cryoprotective Effects of Carrageenan Oligosaccharides on Crayfish (Procambarus clarkii) during Superchilling. Foods. 2023; 12(11):2258. https://doi.org/10.3390/foods12112258
Chicago/Turabian StyleLiao, E, Yuxin Wu, Yang Pan, Ying Zhang, Peng Zhang, and Jiwang Chen. 2023. "Cryoprotective Effects of Carrageenan Oligosaccharides on Crayfish (Procambarus clarkii) during Superchilling" Foods 12, no. 11: 2258. https://doi.org/10.3390/foods12112258
APA StyleLiao, E., Wu, Y., Pan, Y., Zhang, Y., Zhang, P., & Chen, J. (2023). Cryoprotective Effects of Carrageenan Oligosaccharides on Crayfish (Procambarus clarkii) during Superchilling. Foods, 12(11), 2258. https://doi.org/10.3390/foods12112258