
Metabolomics-Based Analyses of Dynamic Changes in Flavonoid Profiles in the Black Mulberry Winemaking Process


Abstract
1. Introduction
2. Materials and Methods
2.1. Chemicals and Reagents
2.2. Collection of Samples and Metabolite Extraction
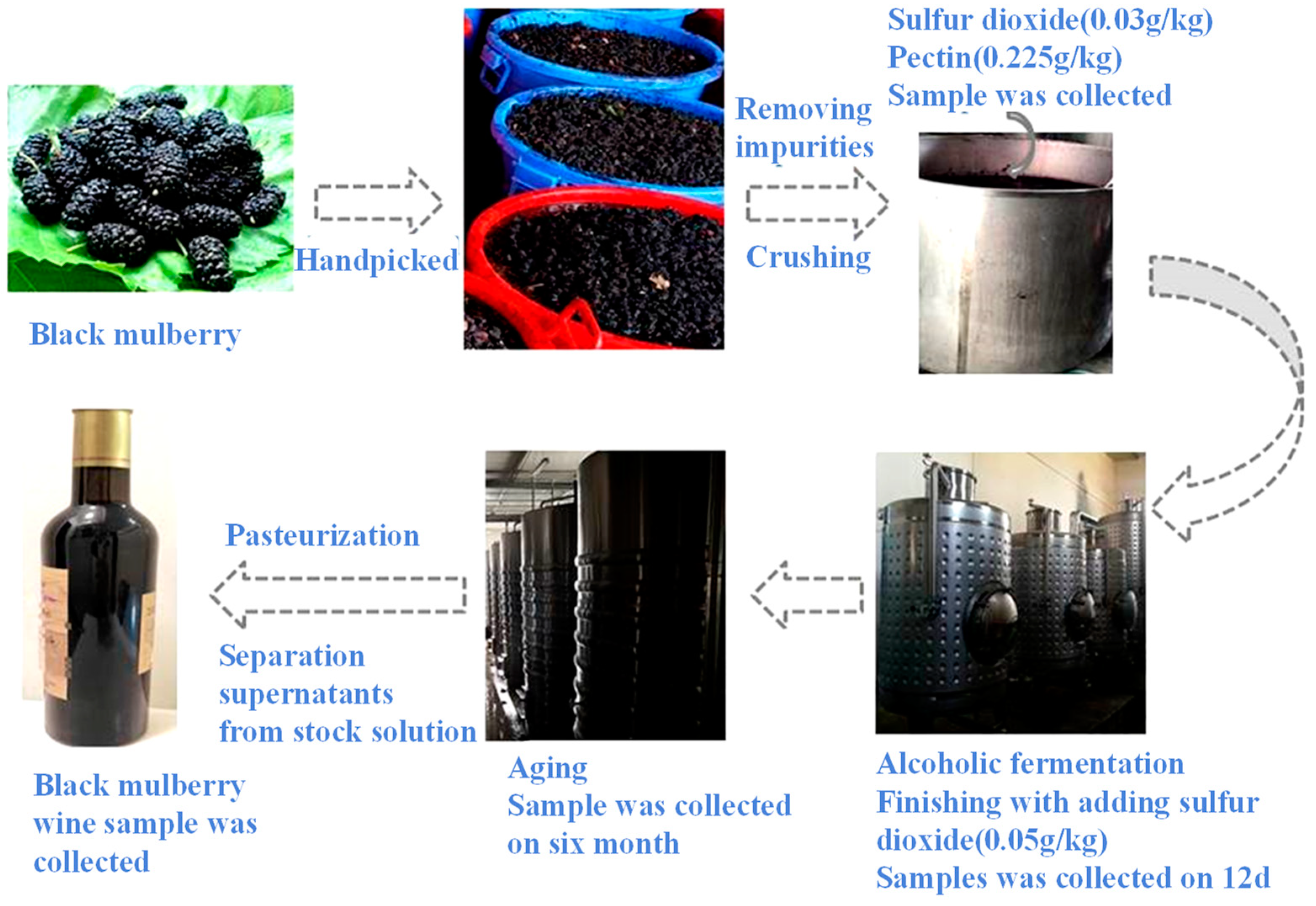
2.3. Mulberry Winemaking
2.4. Analysis of Physicochemical Property
2.5. UHPLC-QE-MS/MS Analyses and Data Processing
2.6. Enrichment Analysis Based on Metabolic Pathways
2.7. Statistical Analysis
3. Results and Discussion
3.1. Physicochemical Parameter Analysis
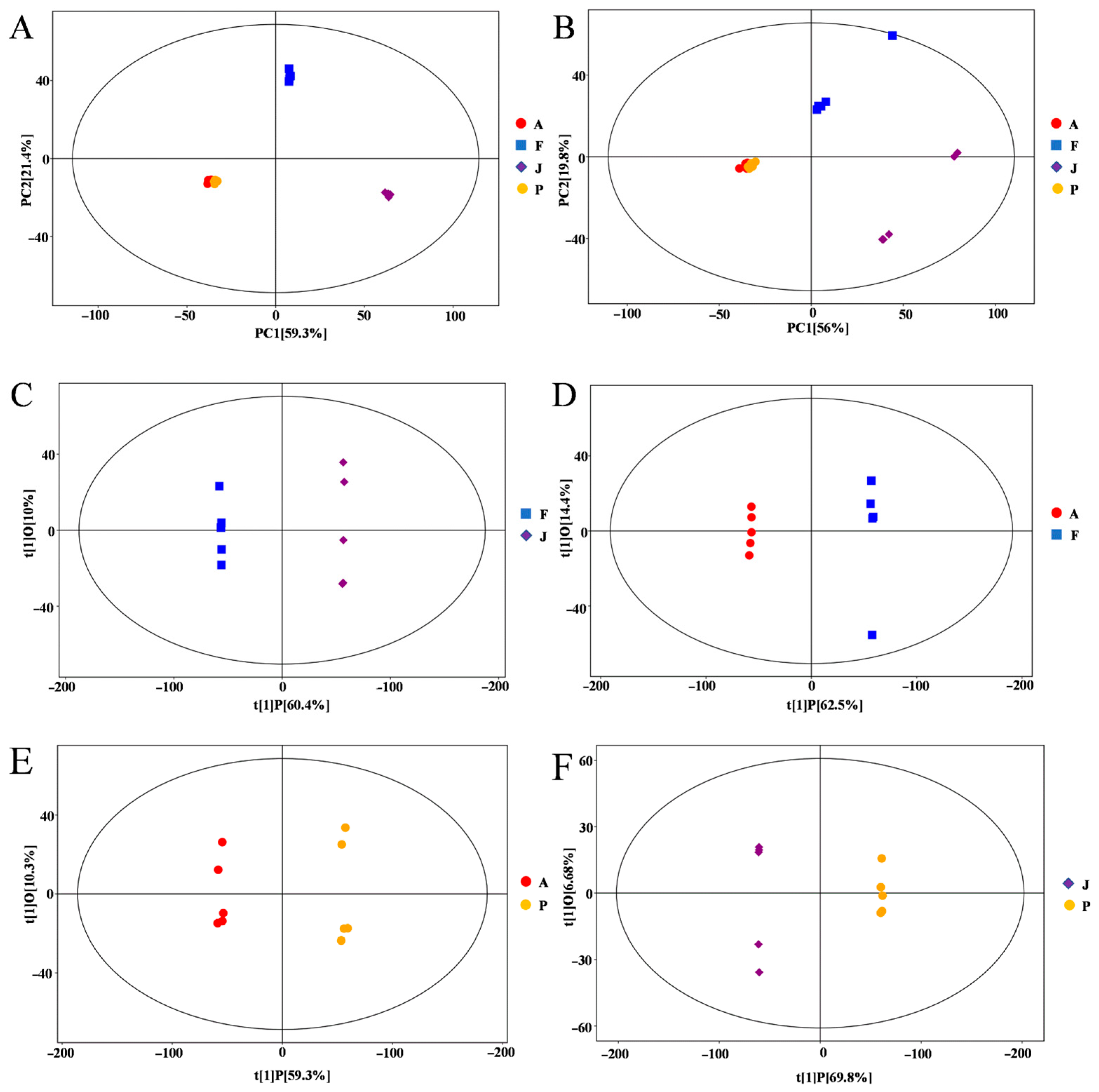
3.2. Multivariate Statistical Analysis
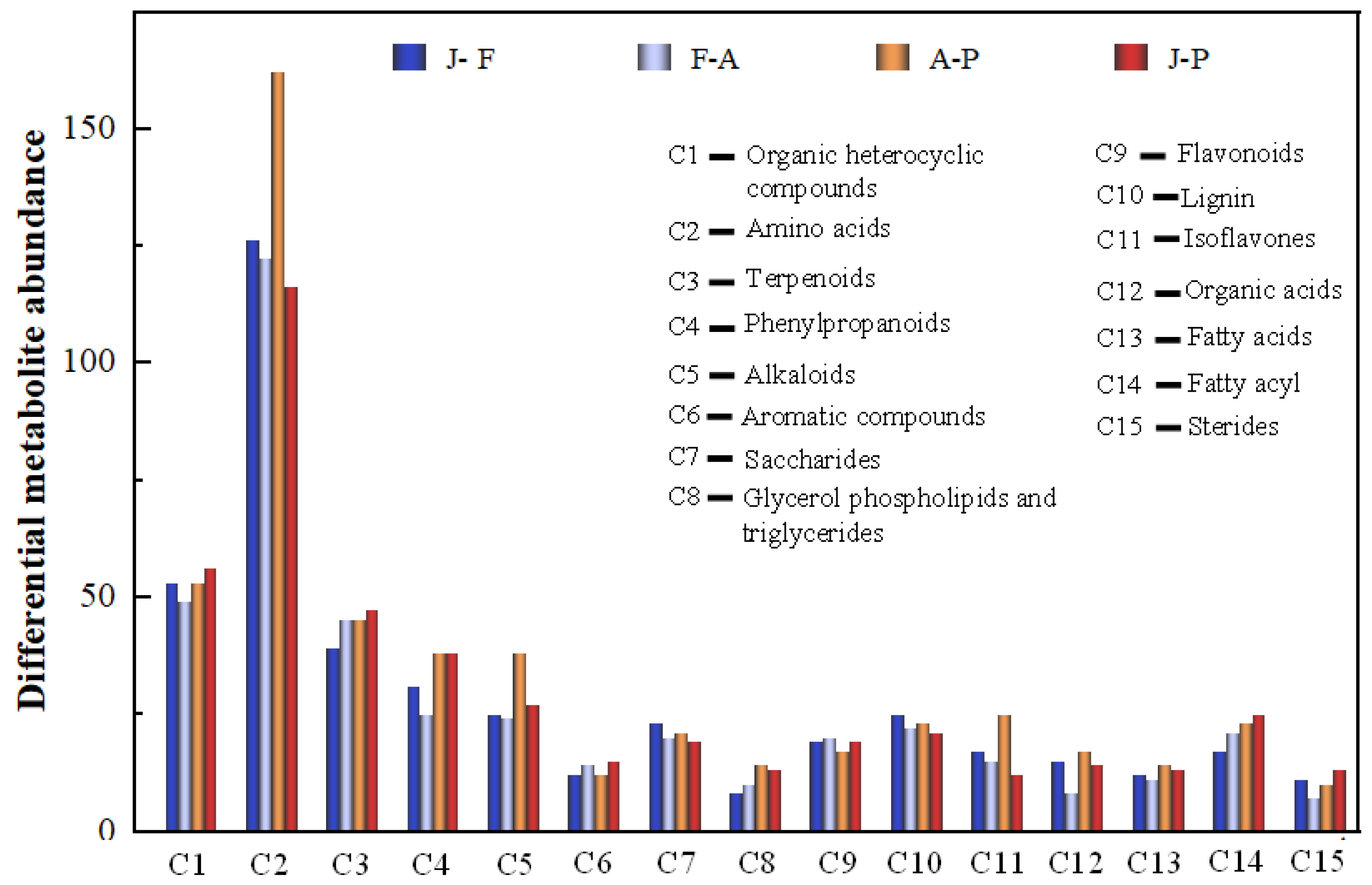
3.3. Screening of Differential Metabolites
3.3.1. Determination of Differential Markers
3.3.2. Relationships between Major Metabolites and Physicochemical Properties
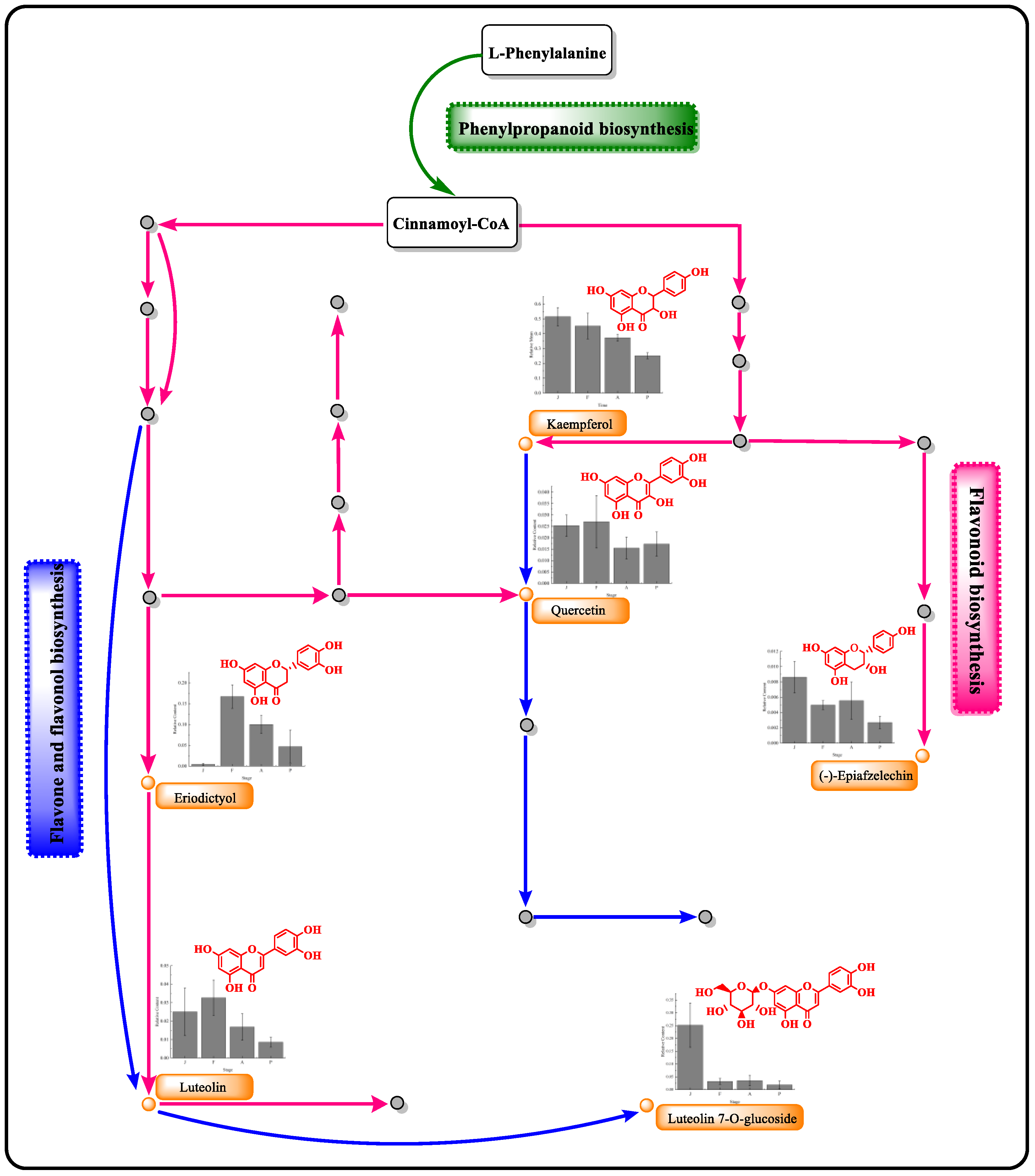
3.4. Flavonoid Metabolites and Flavonoid Metabolic Pathway Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Jiang, Y.; Nie, W.-J. Chemical properties in fruits of mulberry species from the Xinjiang province of China. Food Chem. 2015, 174, 460–466. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Yang, Z.; Zeng, Q.; Wang, S.; Luo, Y.; Huang, Y.; Xin, Y.; He, N. Abnormal expression of bHLH3 disrupts a flavonoid homeostasis network, causing differences in pigment composition among mulberry fruits. Hortic. Res. 2020, 7, 83. [Google Scholar] [CrossRef] [PubMed]
- Tomas, M.; Toydemir, G.; Boyacioglu, D.; Hall, R.D.; Beekwilder, J.; Capanoglu, E. Processing black mulberry into jam: Effects on antioxidant potential and in vitro bioaccessibility. J. Sci. Food Agric. 2017, 97, 3106–3113. [Google Scholar] [CrossRef] [PubMed]
- Kwaw, E.; Ma, Y.; Tchabo, W.; Apaliya, M.T.; Wu, M.; Sackey, A.S.; Xiao, L.; Tahir, H.E. Effect of lactobacillus strains on phenolic profile, color attributes and antioxidant activities of lactic-acid-fermented mulberry juice. Food Chem. 2018, 250, 148–154. [Google Scholar] [CrossRef] [PubMed]
- Hussain, A.; Bose, S.; Wang, J.-H.; Yadav, M.K.; Mahajan, G.B.; Kim, H. Fermentation, a feasible strategy for enhancing bioactivity of herbal medicines. Food Res. Int. 2016, 81, 1–16. [Google Scholar] [CrossRef]
- Hur, S.J.; Lee, S.Y.; Kim, Y.-C.; Choi, I.; Kim, G.-B. Effect of fermentation on the antioxidant activity in plant-based foods. Food Chem. 2014, 160, 346–356. [Google Scholar] [CrossRef]
- Hu, S.; He, C.; Li, Y.; Yu, Z.; Chen, Y.; Wang, Y.; Ni, D. Changes of fungal community and non-volatile metabolites during pile-fermentation of dark green tea. Food Res. Int. 2021, 147, 110472. [Google Scholar] [CrossRef]
- Huynh, N.T.; Van Camp, J.; Smagghe, G.; Raes, K. Improved Release and Metabolism of Flavonoids by Steered Fermentation Processes: A Review. Int. J. Mol. Sci. 2014, 15, 19369–19388. [Google Scholar] [CrossRef]
- Wang, R.; Sun, J.; Lassabliere, B.; Yu, B.; Liu, S.Q. UPLC-Q-TOF-MS Mased Metabolomics and Chemometric Analyses for Green Tea Fermented with Saccharomycesboulardii CNCM I-745 and Lactiplantibacillus plantarum 299V. Curr. Res. Food Sci. 2022, 5, 471–478. [Google Scholar] [CrossRef]
- Wang, D.; Wang, M.; Cao, L.; Wang, X.; Sun, J.; Yuan, J.; Gu, S. Changes and correlation of microorganism and flavor substances during persimmon vinegar fermentation. Food Biosci. 2022, 46, 101565. [Google Scholar] [CrossRef]
- Zhong, J.; Ren, D.; Shang, Y.; Huang, S.; Li, Y.; Hu, Y.; Yi, L. Targeted identification of glycosylated flavones and isomers in green tea through integrated ion-filtering strategy and mass-fragmentation characteristics based on the UPLC–Q–Orbitrap–MS/MS platform. Food Chem. 2022, 377, 131901. [Google Scholar] [CrossRef]
- Wilkowska, A.; Pogorzelski, E. Aroma enhancement of cherry juice and wine using exogenous glycosidases from mould, yeast and lactic acid bacteria. Food Chem. 2017, 237, 282–289. [Google Scholar] [CrossRef]
- Echeverría, J.; Opazo, J.; Mendoza, L.; Urzúa, A.; Wilkens, M. Structure-Activity and Lipophilicity Relationships of Selected Antibacterial Natural Flavones and Flavanones of Chilean Flora. Molecules 2017, 22, 608. [Google Scholar] [CrossRef]
- Kubiak-Tomaszewska, G.; Roszkowski, P.; Grosicka-Maciąg, E.; Strzyga-Łach, P.; Struga, M. Effect of Hydroxyl Groups Esterification with Fatty Acids on the Cytotoxicity and Antioxidant Activity of Flavones. Molecules 2022, 27, 420. [Google Scholar] [CrossRef]
- Hu, F.; Qin, Y.; Zhou, Y.; Li, L.; Wang, Y.; Deng, Z. Characterization of precipitation from citrus vinegar during ageing: Chemical constituents, formation mechanism and anti-proliferative effect. Food Funct. 2022, 13, 4930–4940. [Google Scholar] [CrossRef]
- Dai, W.; Qi, D.; Yang, T.; Lv, H.; Guo, L.; Zhang, Y.; Zhu, Y.; Peng, Q.; Xie, D.; Tan, J.; et al. Nontargeted Analysis Using Ultraperformance Liquid Chromatography-Quadrupole Time-of-Flight Mass Spectrometry Uncovers the Effects of Harvest Sea-son on the Metabolites and Taste Quality of Tea (Camellia sinensis L.). J. Agric. Food Chem. 2015, 63, 9869–9878. [Google Scholar] [CrossRef]
- López, R.; Tenorio, C.; Gutiérrez, A.R.; Garde-Cerdán, T.; Garijo, P.; González-Arenzana, L.; López-Alfaro, I.; Santamaría, P. Elaboration of Tempranillo Wines at Two Different pHs. Influence on Biogenic Amine Contents. Food Control 2012, 25, 583–590. [Google Scholar] [CrossRef]
- Scutarașu, E.C.; Teliban, I.V.; Zamfir, C.I.; Luchian, C.E.; Colibaba, L.C.; Niculaua, M.; Cotea, V.V. Effect of Different Winemaking Conditions on Organic Acids Compounds of White Wines. Foods 2021, 10, 2569. [Google Scholar] [CrossRef]
- Sun, S.Y.; Chen, Z.X.; Jin, C.W. Combined influence of lactic acid bacteria starter and final pH on the induction of malolactic fermentation and quality of cherry wines. LWT 2018, 89, 449–456. [Google Scholar] [CrossRef]
- Qian, M.; Ruan, F.; Zhao, W.; Dong, H.; Bai, W.; Li, X.; Huang, X.; Li, Y. The dynamics of physicochemical properties, microbial community, and flavor metabolites during the fermentation of semi-dry Hakka rice wine and traditional sweet rice wine. Food Chem. 2023, 416, 135844. [Google Scholar] [CrossRef]
- Yang, L.; Fan, W.; Xu, Y. GC × GC-TOF/MS and UPLC-Q-TOF/MS Based Untargeted Metabolomics Coupled with Physicochemical Properties to Reveal the Characteristics of Different Type Daqus for Making Soy Sauce Aroma and Flavor Type Baijiu. LWT 2021, 146, 111416. [Google Scholar] [CrossRef]
- Li, J.; Wu, J.; Xu, N.; Yu, Y.; Brake, J.; Xu, R.; Wu, X. Dynamic Evolution and Correlation between Microorganisms and Metabolites during Manufacturing Process and Storage of Pu-erh Tea. LWT 2022, 158, 113128. [Google Scholar] [CrossRef]
- Li, Z.; Howell, K.; Fang, Z.; Zhang, P. Sesquiterpenes in grapes and wines: Occurrence, biosynthesis, functionality, and influence of winemaking processes. Compr. Rev. Food Sci. Food Saf. 2020, 19, 247–281. [Google Scholar] [CrossRef] [PubMed]
- Cadi, H.E.; Bouzidi, H.E.; Selama, G.; Ramdan, B.; Majdoub, Y.O.E.; Alibrando, F.; Arena, K.; Lovillo, M.P.; Brigui, J.; Mondello, L.; et al. Elucidation of Antioxidant Compounds in Moroccan Chamaerops humilis L. Fruits by GC–MS and HPLC–MS Techniques. Molecules 2021, 26, 2710. [Google Scholar] [CrossRef]
- Salas-Arias, K.; Irías-Mata, A.; Sánchez-Kopper, A.; Hernández-Moncada, R.; Salas-Morgan, B.; Villalta-Romero, F.; Calvo-Castro, L.A. Strawberry Fragaria x ananassa cv. Festival: A Polyphenol-Based Phytochemical Characterization in Fruit and Leaf Extracts. Molecules 2023, 28, 1865. [Google Scholar] [CrossRef]
- Kocaefe-Özşen, N.; Yilmaz, B.; Alkım, C.; Arslan, M.; Topaloğlu, A.; Kısakesen, H.L.; Gülsev, E.; Çakar, Z.P. Physiological and Molecular Characterization of an Oxidative Stress-Resistant Saccharomyces cerevisiae Strain Obtained by Evolutionary Engineering. Front. Microbiol. 2022, 13, 822864. [Google Scholar] [CrossRef]
- Zhang, L.; Qin, Z.; Zhang, L.; Jiang, Y.; Zhu, J. Dynamic changes of quality and flavor characterization of Zhejiang rosy vinegar during fermentation and aging based on untargeted metabolomics. Food Chem. 2023, 404, 134702. [Google Scholar] [CrossRef]
- Velmurugan, B.K.; Lin, J.T.; Mahalakshmi, B.; Chuang, Y.C.; Lin, C.C.; Lo, Y.S.; Hsieh, M.J.; Chen, M.K. Luteolin-7-O-Glucoside Inhibits Oral Cancer Cell Migration and Invasion by Regulating Matrix Metalloproteinase-2 Expression and Extracellular Signal-Regulated Kinase Pathway. Biomolecules 2020, 10, 502. [Google Scholar] [CrossRef]
- Li, N.; Zhu, H.T.; Wang, D.; Zhang, M.; Yang, C.R.; Zhang, Y.J. New Flavoalkaloids with Potent α-Glucosidase and Acetylcholinesterase Inhibitory Activities from Yunnan Black Tea ‘Jin-Ya’. J. Agric. Food Chem. 2020, 68, 7955–7963. [Google Scholar] [CrossRef]
- Devi, K.P.; Malar, D.S.; Nabavi, S.F.; Sureda, A.; Xiao, J.; Nabavi, S.M.; Daglia, M. Kaempferol and inflammation: From chemistry to medicine. Pharmacol. Res. 2015, 99, 1–10. [Google Scholar] [CrossRef]
- Wang, J.; Fang, X.; Ge, L.; Cao, F.; Zhao, L.; Wang, Z.; Xiao, W. Antitumor, antioxidant and anti-inflammatory activities of kaempferol and its corresponding glycosides and the enzymatic preparation of kaempferol. PLoS ONE 2018, 13, e0197563. [Google Scholar] [CrossRef]
- Du, H.; Liu, M.; Yang, X.; Zhai, G. The Role of Glycyrrhetinic Acid Modification on Preparation and Evaluation of Quercetin-Loaded Chitosan-Based Self-Aggregates. J. Colloid Interface Sci. 2015, 460, 87–96. [Google Scholar] [CrossRef]
- Kandemir, K.; Tomas, M.; McClements, D.J.; Capanoglu, E. Recent Advances on the Improvement of Quercetin Bioavailability. Trends Food Sci. Technol. 2022, 119, 192–200. [Google Scholar] [CrossRef]
- Baiano, A.; Mentana, A.; Quinto, M.; Centonze, D.; Longobardi, F.; Ventrella, A.; Agostiano, A.; Varva, G.; De Gianni, A.; Terracone, C.; et al. The effect of in-amphorae aging on oenological parameters, phenolic profile and volatile composition of Minutolo white wine. Food Res. Int. 2015, 74, 294–305. [Google Scholar] [CrossRef]
- Liu, Y.; Duan, C.; Shi, Y. Evolution of Malvidin-3-Glucoside and Color Characteristics of Red Wines during Forced Aging Processes: Effect of (+)-Catechin and (−)-Epicatechin Addition. J. Food Process. Preserv. 2021, 45, e15177. [Google Scholar] [CrossRef]
- Rajan, V.K.; Ahamed, T.S.; Muraleedharan, K. Studies on the UV filtering and radical scavenging capacity of the bitter masking flavanone Eriodictyol. J. Photochem. Photobiol. B Biol. 2018, 185, 254–261. [Google Scholar] [CrossRef]
- Shafreen, R.M.B.; Lakshmi, S.A.; Pandian, S.K.; Kim, Y.-M.; Deutsch, J.; Katrich, E.; Gorinstein, S. In Vitro and In Silico Interaction Studies with Red Wine Polyphenols against Different Proteins from Human Serum. Molecules 2021, 26, 6686. [Google Scholar] [CrossRef]
- Carrascón, V.; Vallverdu-Queralt, A.; Meudec, E.; Sommerer, N.; Fernandez-Zurbano, P.; Ferreira, V. The kinetics of oxygen and SO2 consumption by red wines. What do they tell about oxidation mechanisms and about changes in wine composition? Food Chem. 2018, 241, 206–214. [Google Scholar] [CrossRef]
- Gutiérrez-Escobar, R.; Aliaño-González, M.J.; Cantos-Villar, E. Wine Polyphenol Content and Its Influence on Wine Quality and Properties: A Review. Molecules 2021, 26, 718. [Google Scholar] [CrossRef]
- Tohge, T.; de Souza, L.P.; Fernie, A.R. Current understanding of the pathways of flavonoid biosynthesis in model and crop plants. J. Exp. Bot. 2017, 68, 4013–4028. [Google Scholar] [CrossRef]
- Popović, B.; Štajner, D.; Ždero-Pavlović, R.; Tumbas-Šaponjac, V.; Čanadanović-Brunet, J.; Orlović, S. Water stress induces changes in polyphenol profile and antioxidant capacity in poplar plants (Populus spp.). Plant Physiol. Biochem. 2016, 105, 242–250. [Google Scholar] [CrossRef] [PubMed]
- Marín, L.; Gutiérrez-del-Río, I.; Yagüe, P.; Manteca, Á.; Villar, C.J.; Lombó, F. De Novo Biosynthesis of Apigenin, Luteolin, and Eriodictyol in the Actinomycete Streptomyces albus and Production Improvement by Feeding and Spore Conditioning. Front. Microbiol. 2017, 8, 921. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Physicochemical Indices | Black Mulberry Winemaking Process | |||
|---|---|---|---|---|
| J | F | A | P | |
| pH | 5.48 ± 0.006 a | 3.92 ± 0.006 c | 3.99 ± 0.006 d | 3.95 ± 0.010 b |
| TA (g/L tartaric acid) | 2.793 ± 0.326 c | 12.757 ± 2.295 a | 6.597 ± 0.078 b | 8.443 ± 0.078 b |
| TS (g/L) | 139.152 ± 26.492 a | 58.889 ± 1.111 b | 21.783 ± 0.729 c | 39.974 ± 2.884 cb |
| TSS (%) | 16.9 ± 0.100 b | 19.97 ± 0.153 a | 11.33 ± 0.578 d | 12.43 ± 0.252 c |
| Alcohol (v/v, %) | 0 c | 8.0 ± 0.1 b | 10.0 a | 10.0 ± 0.1 a |
| Fermentation Stage | Marker Compound | RT (min) | m/z | Mass Error | Ion Model | VIP | p-Value | FC | log2 (FC) |
|---|---|---|---|---|---|---|---|---|---|
| J-F | Luteolin | 5.0729 | 287.0547 | 0.9783 | pos | 1.1786 | 0.0030 | 2.0701 | 1.0497 |
| Luteolin-7-O-glucoside | 8.9374 | 449.1077 | 0.9403 | pos | 1.2407 | 0.0044 | 7.8548 | 2.9736 | |
| (−)-Epiafzelechin | 2.4847 | 275.0911 | 0.8052 | pos | 1.0672 | 0.0145 | 1.7339 | 0.7940 | |
| Eriodictyol | 6.6508 | 287.0538 | 0.7186 | neg | 1.4765 | 0.2353 | 0.0357 | −4.8081 | |
| F-A | Luteolin | 5.0729 | 287.0547 | 0.9783 | pos | 1.1847 | 0.0001 | 0.5665 | −0.8198 |
| Kaempferol | 0.7897 | 285.0407 | 0.9975 | neg | 1.1712 | 0.2597 | 8.5817 | 3.1013 | |
| Quercetin | 1.0179 | 301.0354 | 0.9579 | neg | 1.0688 | 0.2721 | 7.5019 | 2.9072 | |
| A-P | Kaempferol | 2.9898 | 287.0545 | 0.9975 | pos | 1.2480 | 0.0000 | 1.4801 | 0.5657 |
| Luteolin | 5.0624 | 287.0547 | 0.9783 | pos | 1.2650 | 0.0000 | 4.7709 | 2.2543 | |
| J-P | Kaempferol | 2.9898 | 287.0545 | 0.9975 | pos | 1.1606 | 0.0000 | 2.0471 | 1.0336 |
| Luteolin | 5.0729 | 287.0547 | 0.9783 | pos | 1.1650 | 0.0005 | 5.5954 | 2.4842 | |
| Luteolin-7-O-glucoside | 8.9374 | 449.1077 | 0.9403 | pos | 1.1403 | 0.0033 | 13.5785 | 3.7632 | |
| (−)-Epiafzelechin | 2.4847 | 275.0911 | 0.8052 | pos | 1.1044 | 0.0003 | 3.2103 | 1.6827 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Qin, Y.; Xu, H.; Chen, Y.; Lei, J.; Sun, J.; Zhao, Y.; Lian, W.; Zhang, M. Metabolomics-Based Analyses of Dynamic Changes in Flavonoid Profiles in the Black Mulberry Winemaking Process. Foods 2023, 12, 2221. https://doi.org/10.3390/foods12112221
Qin Y, Xu H, Chen Y, Lei J, Sun J, Zhao Y, Lian W, Zhang M. Metabolomics-Based Analyses of Dynamic Changes in Flavonoid Profiles in the Black Mulberry Winemaking Process. Foods. 2023; 12(11):2221. https://doi.org/10.3390/foods12112221
Chicago/Turabian StyleQin, Yanan, Haotian Xu, Ya Chen, Jing Lei, Jingshuai Sun, Yan Zhao, Weijia Lian, and Minwei Zhang. 2023. "Metabolomics-Based Analyses of Dynamic Changes in Flavonoid Profiles in the Black Mulberry Winemaking Process" Foods 12, no. 11: 2221. https://doi.org/10.3390/foods12112221
APA StyleQin, Y., Xu, H., Chen, Y., Lei, J., Sun, J., Zhao, Y., Lian, W., & Zhang, M. (2023). Metabolomics-Based Analyses of Dynamic Changes in Flavonoid Profiles in the Black Mulberry Winemaking Process. Foods, 12(11), 2221. https://doi.org/10.3390/foods12112221