
Total Protein Content, Amino Acid Composition and Eating-Quality Evaluation of Foxtail Millet (Setaria italica (L.) P. Beauv)

Abstract
1. Introduction
2. Materials and Methods
2.1. Plant Materials and Growth Environment
2.2. Total Protein Content (TPC) Determination
2.3. Amino Acid Composition and Evaluation of Nutritional Parameters
2.4. Starch Content Analysis
2.5. Gel Consistency
2.6. Grain Hardness and Color Index Determination
2.7. Data Collection and Statistical Analysis
3. Results and Discussion
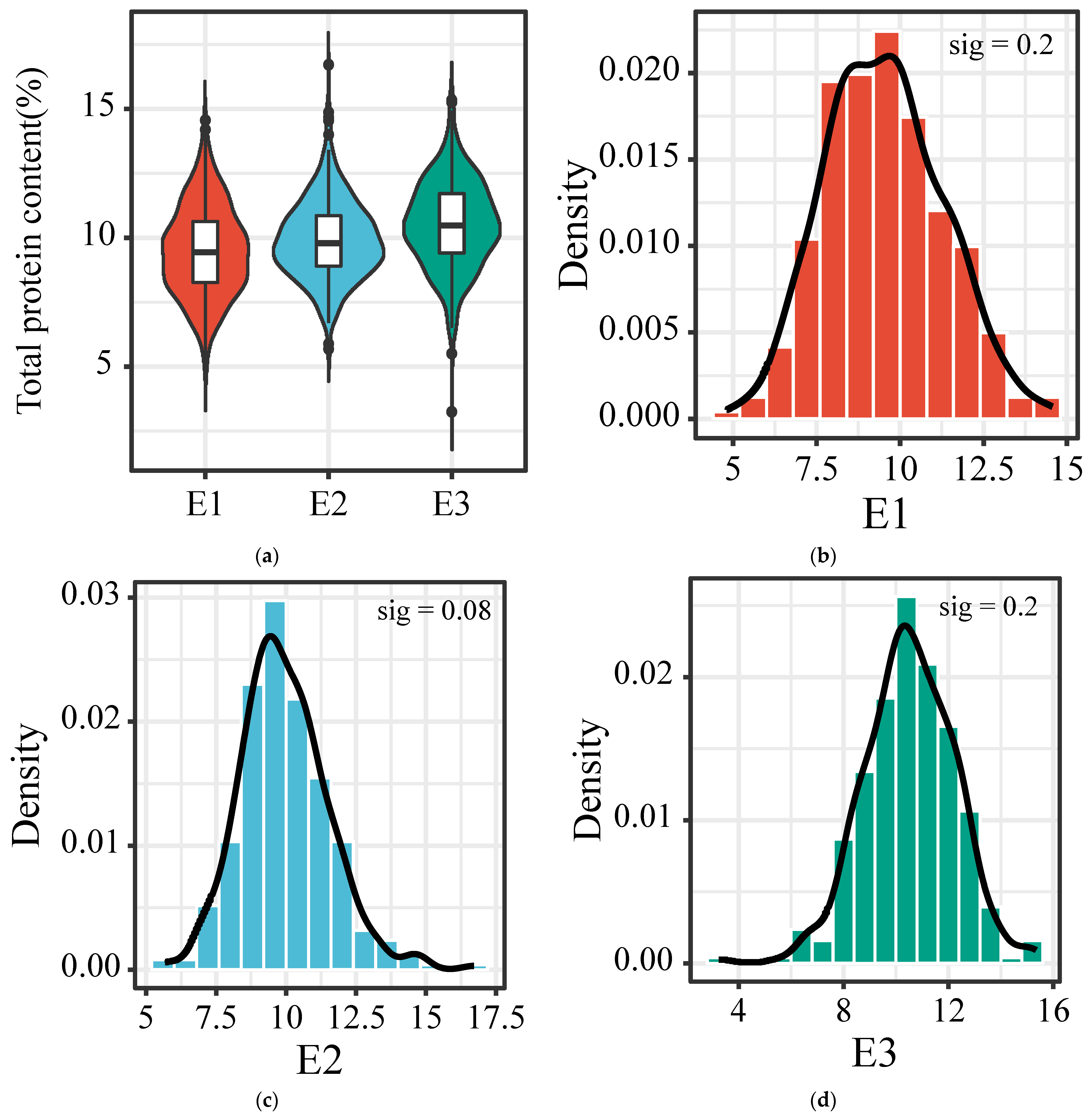
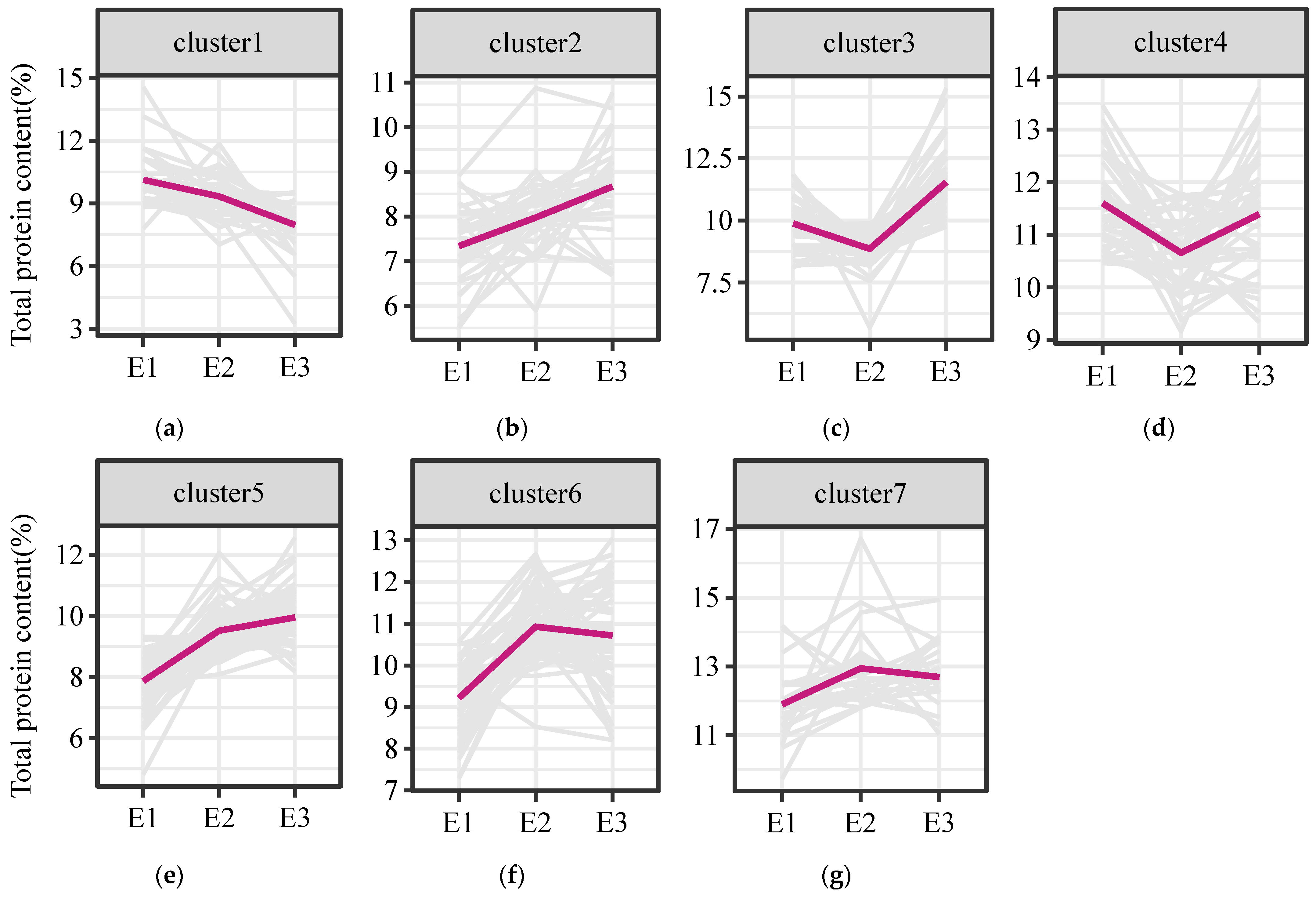
3.1. Characteristics of Total Protein Content Variation in Foxtail Millet
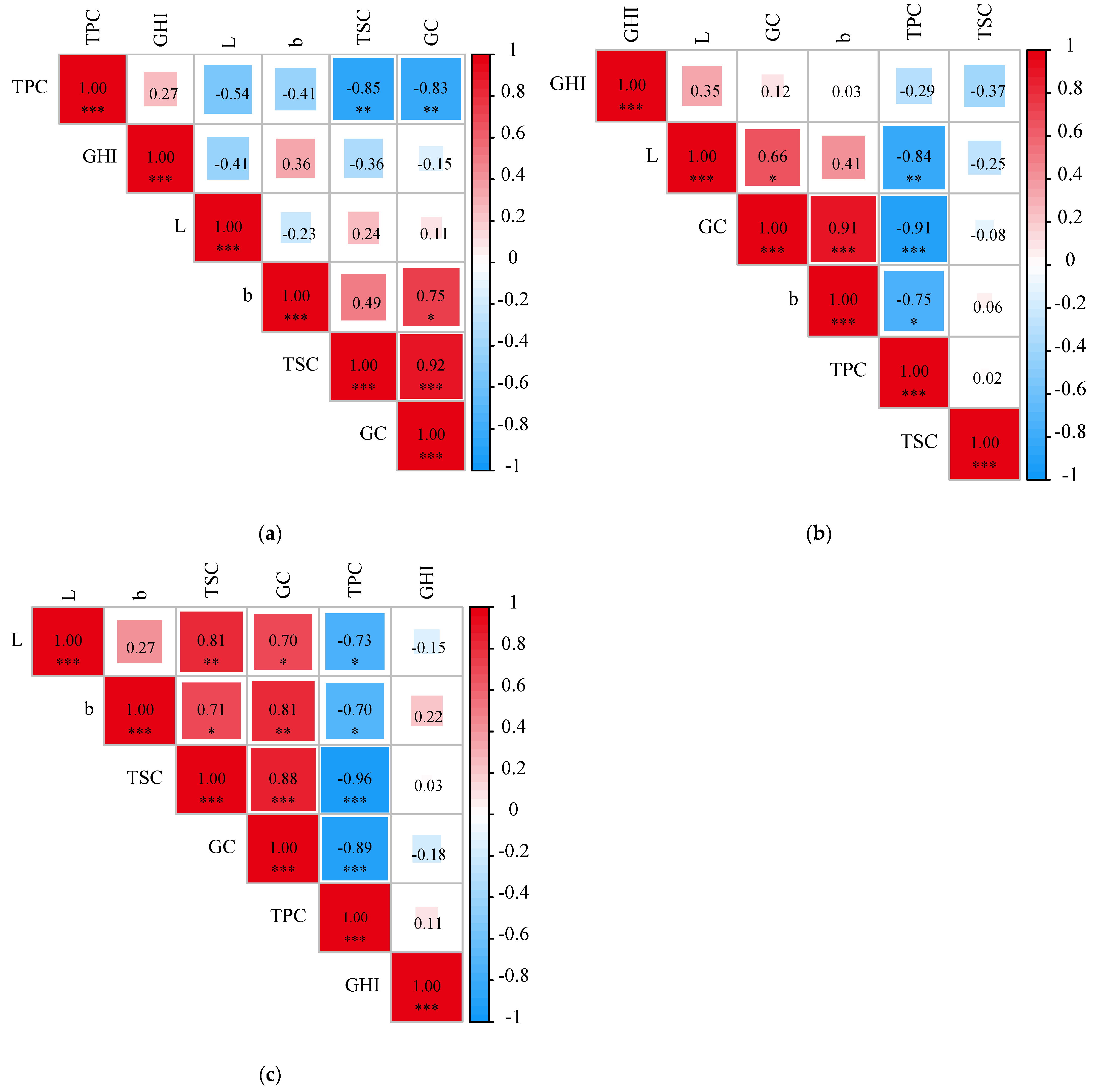
3.2. Correlation Analysis of TPC and Climate Factors
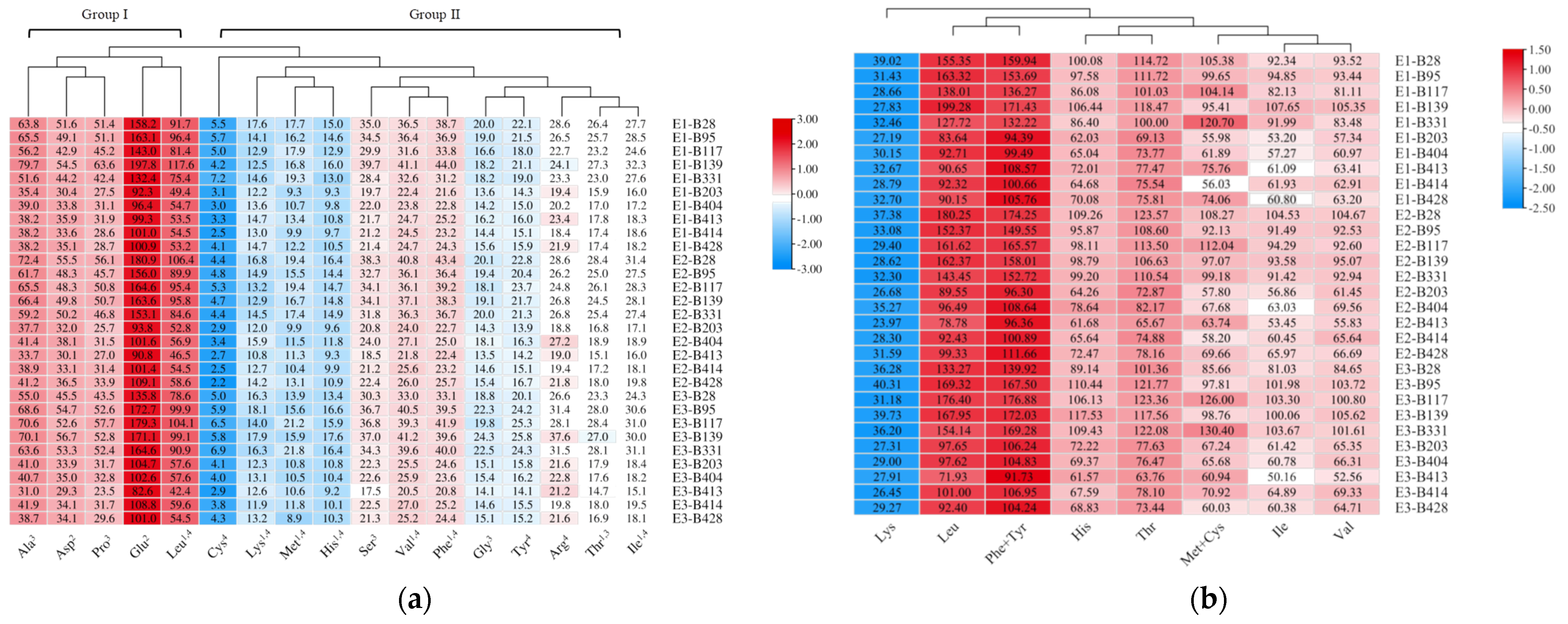
3.3. Amino Acid Composition and Evaluation in Foxtail Millet Grains
3.4. Eating-Quality Evaluation and Correlation Analysis of Foxtail Millet
3.5. Identification of Superior Eating and Protein Quality Variety
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Yang, X.; Wan, Z.; Perry, L.; Lu, H.; Wang, Q.; Zhao, C.; Li, J.; Xie, F.; Yu, J.; Cui, T.; et al. Early millet use in northern China. Proc. Natl. Acad. Sci. USA 2012, 109, 3726–3730. [Google Scholar] [CrossRef]
- Sharma, N.; Niranjan, K. Foxtail millet: Properties, processing, health benefits, and uses. Food Rev. Int. 2018, 34, 329–363. [Google Scholar] [CrossRef]
- Peng, R.; Zhang, B. Foxtail Millet: A new model for C4 plants. Trends Plant Sci. 2021, 26, 199–201. [Google Scholar] [CrossRef]
- Yang, Z.; Zhang, H.; Li, X.; Shen, H.; Gao, J.; Hou, S.; Zhang, B.; Mayes, S.; Bennett, M.; Ma, J.; et al. A mini foxtail millet with an Arabidopsis-like life cycle as a C4 model system. Nat. Plants 2020, 6, 1167–1178. [Google Scholar] [CrossRef]
- Hermes, T.R.; Frachetti, M.D.; Doumani, D.P.; MarYashev, A.; Nebel, A.; Makarewicz, C.A. Early integration of pastoralism and millet cultivation in Bronze Age Eurasia. Proc. Biol Sci 2019, 286, 20191273. [Google Scholar] [CrossRef]
- Li, P.; He, W.; Wu, G. Composition of amino acids in foodstuffs for humans and animals. Adv. Exp. Med. Biol. 2021, 1332, 189–210. [Google Scholar]
- Li, H.; Han, S.; Huo, Y.; Ma, G.; Sun, Z.; Li, H.; Hou, S.; Han, Y. Comparative metabolomic and transcriptomic analysis reveals a coexpression network of the carotenoid metabolism pathway in the panicle of Setaria italica. BMC Plant Biol. 2022, 22, 105. [Google Scholar] [CrossRef]
- Hou, S.; Man, X.; Lian, B.; Ma, G.; Sun, Z.; Han, L.; Yan, L.; Gao, H.; Du, W.; Wang, X.; et al. Folate metabolic profiling and expression of folate metabolism-related genes during panicle development in foxtail millet (Setaria italica (L.) P. Beauv). J. Sci. Food Agric. 2022, 102, 268–279. [Google Scholar] [CrossRef]
- Yang, X.; Wang, L.; Zhou, X.; Shuang, S.; Zhu, Z.; Li, N.; Li, Y.; Liu, F.; Liu, S.; Lu, P.; et al. Determination of protein, fat, starch, and amino acids in foxtail millet [Setaria italica (L.) Beauv.] by Fourier transform near-infrared reflectance spectroscopy. Food Sci. Biotechnol. 2013, 22, 1495–1500. [Google Scholar] [CrossRef]
- Sachdev, N.; Goomer, S.; Sing, L.R. Foxtail millet: A potential crop to meet future demand scenario for alternative sustainable protein. J. Sci. Food Agric. 2021, 101, 831–842. [Google Scholar] [CrossRef] [PubMed]
- Lynch, J.M.; Barbano, D.M. Kjeldahl nitrogen analysis as a reference method for protein determination in dairy products. J. AOAC Int. 1999, 82, 1389–1398. [Google Scholar] [CrossRef] [PubMed]
- Shankaramurthy, K.N.; Somannavar, M.S. Moisture, carbohydrate, protein, fat, calcium, and zinc content in finger, foxtail, pearl, and proso millets. Indian J. Health Sci. Biomed. Res. (KLEU) 2019, 12, 228–232. [Google Scholar] [CrossRef]
- Zhang, H.; Wang, X.; Wang, F.; Zhao, F.; Li, X.; Fan, G.; Zhao, Z.; Guo, P. Rapid prediction of Apparent Amylose, total starch, and crude protein by near-infrared reflectance spectroscopy for foxtail millet (Setaria italica). Cereal Chem. 2020, 97, 653–660. [Google Scholar] [CrossRef]
- Nandini, K.M.; Sridhara, S. Response of growth yield and quality parameters of foxtail millet genotypes to different planting density. Int. J. Curr. Microbiol. Appl. Sci. 2019, 8, 1765–1773. [Google Scholar] [CrossRef]
- Gaur, V.S.; Sood, S.; Tiwari, S.; Kuma, A. Genome-wide identification and characterization of seed storage proteins (SSPs) of foxtail millet (Setaria italica (L.) P. Beauv.). 3 Biotech 2018, 8, 415. [Google Scholar] [CrossRef] [PubMed]
- Zhang, F.; Fu, Y.; Liu, Z.; Shen, Q. Comparison of the characteristics of prolamins among foxtail millet varieties with different palatability: Structural, morphological, and physicochemical properties. Int. J. Biol. Macromol. 2021, 186, 194–205. [Google Scholar] [CrossRef] [PubMed]
- Pan, J.; Li, Z.; Wang, Q.; Garrell, A.K.; Liu, M.; Guan, Y.; Zhou, W.; Liu, W. Comparative proteomic investigation of drought responses in foxtail millet. BMC Plant Biol. 2018, 18, 315. [Google Scholar] [CrossRef]
- Bohn, H.L.; Myer, R.A.; O’Connor, G.A. Soil Chemistry; John Wiley & Sons, Inc.: New York, NY, USA, 2002. [Google Scholar]
- Sui, Y.; Harvey, P.J. Effect of light intensity and wavelength on biomass growth and protein and amino acid composition of Dunaliella salina. Foods 2021, 10, 1018. [Google Scholar] [CrossRef]
- Boye, J.; Wijesinha-Bettoni, R.; Burlingame, B. Protein quality evaluation twenty years after the introduction of the protein digestibility corrected amino acid score method. Br. J. Nutr. 2012, 108, S183–S211. [Google Scholar] [CrossRef]
- Hall, M.B. Determination of dietary starch in animal feeds and pet food by an enzymatic-colorimetric method: Collaborative study. J. AOAC Int. 2015, 98, 397–409. [Google Scholar] [CrossRef]
- Cagampang, G.B.; Perez, C.M.; Juliano, B.O. A gel consistency test for eating quality of rice. J. Sci. Food Agric. 1973, 24, 1589–1594. [Google Scholar] [CrossRef]
- Langfelder, P.; Horvath, S. WGCNA: An R package for weighted correlation network analysis. BMC Bioinform. 2008, 9, 1–13. [Google Scholar] [CrossRef]
- Wickham, H. Ggplot2: Elegant Graphics for Data Analysis; Springer: New York, NY, USA, 2016; ISBN 978-3-319-24277-4. [Google Scholar]
- Cao, X.; Jiang, F.; Wang, X.; Zang, Y.; Wu, Z. Comprehensive evaluation and screening for chilling-tolerance in tomato lines at the seedling stage. Euphytica 2015, 205, 569–584. [Google Scholar] [CrossRef]
- Martre, P.; He, J.; Le Gouis, J.; Semenov, M.A. In silico system analysis of physiological traits determining grain yield and protein concentration for wheat as influenced by climate and crop management. J. Exp. Bot. 2015, 66, 3581–3598. [Google Scholar] [CrossRef]
- Bertholdsson, N.O. Characterization of malting barley cultivars with more or less stable grain protein content under varying environmental conditions. Eur. J. Agron. 1999, 10, 1–8. [Google Scholar] [CrossRef]
- Gu, S.; Gu, X.; Geng, J. Effects of different soils and altitudes on the compositioj of amino acids in foxtil millet. Eco-Agric. Res. 2000, 8, 32–35. [Google Scholar]
- Chuma, B.A.; Cotter, M.; Kalisa, A.; Rajaona, A.; Senthilkumar, K.; Stuerz, S.; Vincent, I.; Asch, F. Altitude, temperature, and N Management effects on yield and yield components of contrasting lowland rice cultivars. J. Agron. Crop Sci. 2020, 206, 456–465. [Google Scholar] [CrossRef]
- Song, W.; Yang, R.; Wu, T.; Wu, C.; Sun, S.; Zhang, S.; Jiang, B.; Tian, S.; Liu, X.; Han, T. Analyzing the effects of climate factors on soybean protein, oil contents, and composition by extensive and high-density sampling in China. J. Agric. Food Chem. 2016, 64, 4121–4130. [Google Scholar] [CrossRef]
- Bosaz, L.B.; Gerde, J.A.; Borrás, L.; Cipriotti, P.A.; Ascheri, L.; Campos, M.; Gallo, S.; Rotundo, J.L. Management and environmental factors explaining soybean seed protein variability in central Argentina. Field Crops Res. 2019, 240, 34–43. [Google Scholar] [CrossRef]
- Sharma, M.; Saini, C.S. Amino acid composition, nutritional profiling, mineral content and physicochemical properties of protein isolate from flaxseeds (Linum usitatissimum). J. Food Meas. Charact. 2022, 16, 829–839. [Google Scholar] [CrossRef]
- Taira, H. Amino acid composition of different varieties of foxtain millet (Setaria italica). J. Agric. Food Chem. 1968, 16, 1025–1027. [Google Scholar] [CrossRef]
- Milinovic, J.; Campos, B.; Mata, P.; Diniz, M.; Noronha, J.P. Umami free amino acids in edible green, red, and brown seaweeds from the Portuguese seashore. J. Appl. Phycol. 2020, 32, 3331–3339. [Google Scholar] [CrossRef]
- Muqaddasi, Q.H.; Brassac, J.; Ebmeyer, E.; Kollers, S.; Korzun, V.; Argillier, O.; Stiewe, G.; Plieske, J.; Ganal, M.W.; Röder, M.S. Prospects of GWAS and predictive breeding for European winter wheat’s grain protein content, grain starch content, and grain hardness. Sci. Rep. 2020, 10, 12541. [Google Scholar] [CrossRef]
- Liu, Q.; Tao, Y.; Cheng, S.; Zhou, L.; Tian, J.; Xing, Z.; Liu, G.; Wei, H.; Zhang, H. Relating amylose and protein contents to eating quality in 105 varieties of Japonica rice. Cereal Chem. 2020, 97, 1303–1312. [Google Scholar] [CrossRef]
- Gao, Z.; Zeng, D.; Cheng, F.; Tian, Z.; Guo, L.; Su, Y.; Yan, M.; Jiang, H.; Dong, G.; Huang, Y.; et al. ALK, the key gene for gelatinization temperature, is a modifier gene for gel consistency in rice. J. Integr. Plant Biol. 2011, 53, 756–765. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Environment | ATG | AAT | MDTRG | MDTR | TRF | DASSH | ALT |
|---|---|---|---|---|---|---|---|
| E1 | 2186.30 | 1871.50 | 26.41 | 11.43 | 306.90 | 7.49 | 10,343.00 |
| E2 | 2454.70 | 2031.70 | 27.99 | 10.81 | 279.80 | 7.17 | 7858.00 |
| E3 | 2559.90 | 2048.60 | 28.55 | 10.60 | 299.40 | 6.80 | 8373.00 |
| Environment | Min (%) | Max (%) | Med | AV | SD | CV (%) | SC | KC | Sig (p > 0.05) |
|---|---|---|---|---|---|---|---|---|---|
| E1 (n = 301) | 4.81 | 14.56 | 9.44 | 9.51 c | 1.76 | 18.50 | 0.193 | −0.233 | 0.2 |
| E2 (n = 314) | 5.68 | 16.71 | 9.78 | 9.94 b | 1.58 | 15.89 | 0.557 | 1.169 | 0.08 |
| E3 (n = 316) | 5.50 | 15.34 | 10.49 | 10.53 a | 1.67 | 15.86 | 0.003 | 0.078 | 0.2 |
| Variety | Fresh Amino Acids (%) | Sweet Amino Acids (%) | Bitter Amino Acids (%) | Difference Value between Fresh Amino Acids + Sweet Amino Acids and Bitter Amino Acids (%) |
|---|---|---|---|---|
| B28 | 29.79 | 27.67 | 42.53 | 14.93 |
| B95 | 30.12 | 27.53 | 42.35 | 15.3 |
| B117 | 30.06 | 27.60 | 42.34 | 15.32 |
| B139 | 30.35 | 27.77 | 41.88 | 16.24 |
| B331 | 29.72 | 27.21 | 43.07 | 13.86 |
| B203 | 29.68 | 27.25 | 43.08 | 13.85 |
| B404 | 29.10 | 27.57 | 43.34 | 13.33 |
| B413 | 29.47 | 26.77 | 43.76 | 12.48 |
| B414 | 30.11 | 27.16 | 42.74 | 14.53 |
| B428 | 29.87 | 26.79 | 43.33 | 13.33 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hou, S.; Men, Y.; Wei, M.; Zhang, Y.; Li, H.; Sun, Z.; Han, Y. Total Protein Content, Amino Acid Composition and Eating-Quality Evaluation of Foxtail Millet (Setaria italica (L.) P. Beauv). Foods 2023, 12, 31. https://doi.org/10.3390/foods12010031
Hou S, Men Y, Wei M, Zhang Y, Li H, Sun Z, Han Y. Total Protein Content, Amino Acid Composition and Eating-Quality Evaluation of Foxtail Millet (Setaria italica (L.) P. Beauv). Foods. 2023; 12(1):31. https://doi.org/10.3390/foods12010031
Chicago/Turabian StyleHou, Siyu, Yihan Men, Min Wei, Yijuan Zhang, Hongying Li, Zhaoxia Sun, and Yuanhuai Han. 2023. "Total Protein Content, Amino Acid Composition and Eating-Quality Evaluation of Foxtail Millet (Setaria italica (L.) P. Beauv)" Foods 12, no. 1: 31. https://doi.org/10.3390/foods12010031
APA StyleHou, S., Men, Y., Wei, M., Zhang, Y., Li, H., Sun, Z., & Han, Y. (2023). Total Protein Content, Amino Acid Composition and Eating-Quality Evaluation of Foxtail Millet (Setaria italica (L.) P. Beauv). Foods, 12(1), 31. https://doi.org/10.3390/foods12010031