
Chemical Composition and Agronomic Traits of Allium sativum and Allium ampeloprasum Leaves and Bulbs and Their Action against Listeria monocytogenes and Other Food Pathogens

 ,
,  , ,
, ,  and
and 
Abstract
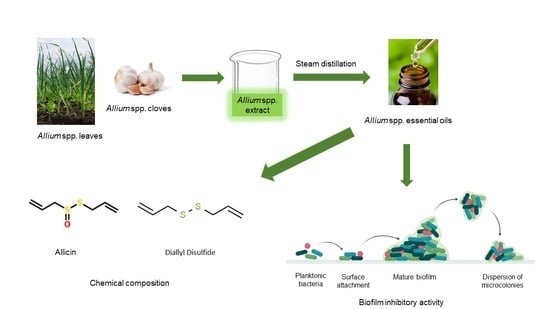
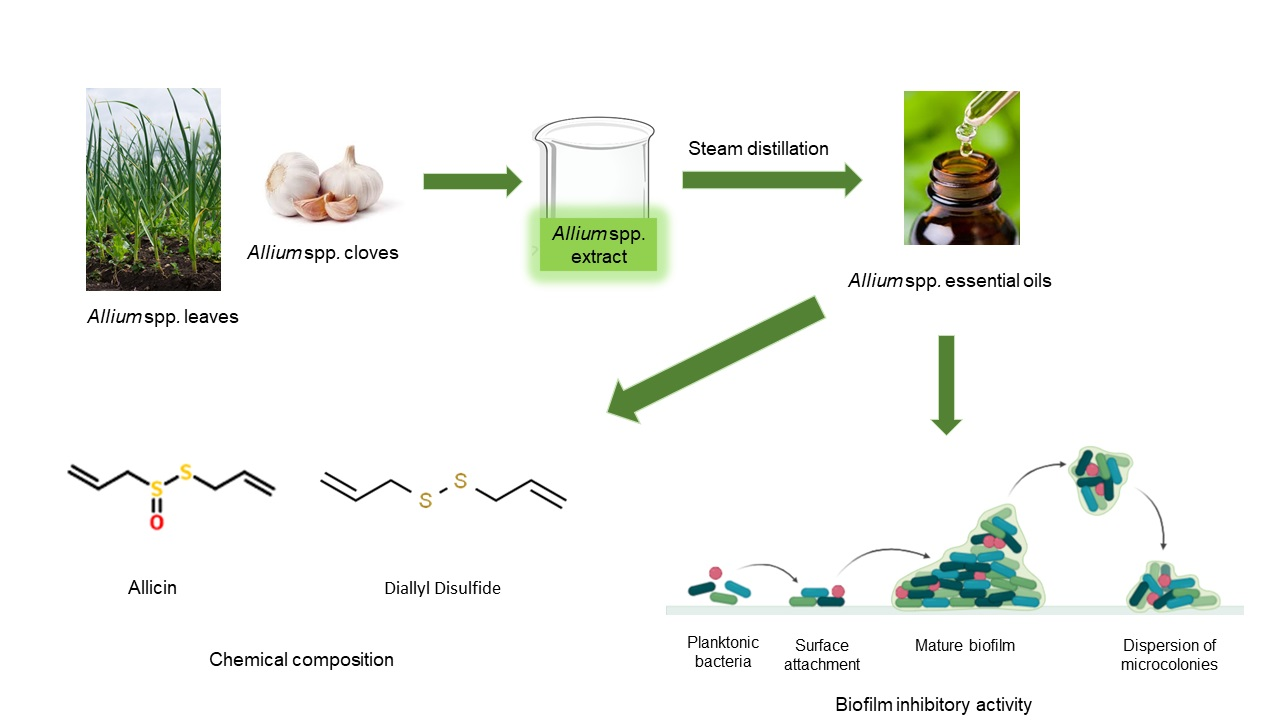
:
1. Introduction
2. Materials and Methods
2.1. Plant Material
2.2. Extraction of Essential Oils
2.3. Composition of the Essential Oils
2.4. Antibacterial Properties of the Oils
Microorganisms and Culture Conditions
2.5. Minimal Inhibitory Concentration (MIC)
2.6. Biofilm Inhibitory Action of the EOs
2.7. Inhibition of Cell Metabolic Activity within the Biofilm
2.8. Statistical Analysis
3. Results and Discussion
3.1. Morphological Traits of Bulbs and Cloves of Garlic Cultivars
3.2. Chemical Composition
3.3. Biofilm Inhibitory Capacity of the EOs
Action of EOs against Bacterial Metabolism
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Friesen, N.; Fritsch, R.M.; Blattner, F.R. Phylogeny and new intrageneric classification of Allium (Alliaceae) based on nuclear ribosomal DNA ITS sequences. J. Syst. Evol. Bot. 2006, 22, 372–395. [Google Scholar] [CrossRef]
- Hanelt, P.; Schultze-Motel, J.; Fritsch; Kruse, J.R.; Maass, H.I.; Ohle, H.; Pistrick, K. Infrageneric grouping of Allium—The Gatersleben approach. In Proceedings of the International Sympsium, Gatersleben, Germany, 11–13 June 1991; pp. 107–123. [Google Scholar]
- Mathew, B. A Review of Allium Section Allium; Royal Botanic Gardens: Kew, UK, 1996. [Google Scholar]
- Londhe, V.P. Role of garlic (Allium sativum) in various diseases: An overview. Angiogenesis 2011, 12, 13. [Google Scholar]
- Griffiths, G.; Trueman, L.; Crowther, T.; Thomas, B.; Smith, B. Onions—A global benefit to health. Phytother. Res. 2002, 16, 603–615. [Google Scholar] [CrossRef] [PubMed]
- Chauhan, N.B. Multiplicity of garlic health effects and Alzheimer’s disease. J. Nutr. Health Aging 2005, 9, 421–432. [Google Scholar]
- Morihara, N.; Nishihama, T.; Ushijima, M.; Ide, N.; Takeda, H.; Hayama, M. Garlic as an antifatigue agent. Mol. Nutr. Food Res. 2007, 51, 1329–1334. [Google Scholar] [CrossRef] [PubMed]
- Iciek, M.; Kwiecień, I.; Włodek, L. Biological properties of garlic and garlic-derived organosulfur compounds. Environ. Mol. Mutagen. 2009, 50, 247–265. [Google Scholar] [CrossRef]
- Nicastro, H.L.; Ross, S.A.; Milner, J.A. Garlic and onions: Their cancer prevention properties. Cancer Prev. Res. 2015, 8, 181–189. [Google Scholar] [CrossRef] [Green Version]
- Guan, M.J.; Zhao, N.; Xie, K.Q.; Zeng, T. Hepatoprotective effects of garlic against ethanol-induced liver injury: A mini-review. Food Chem. Toxicol. 2018, 111, 467–473. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Y.; Anand, R.; Geng, X.; Ding, Y. A mini review: Garlic extract and vascular diseases. Neurol. Res. 2018, 40, 421–425. [Google Scholar] [CrossRef] [PubMed]
- Bhandari, P.R. Garlic (Allium sativum L.): A review of potential therapeutic applications. Int. J. Green Pharm. 2012, 6, 118–129. [Google Scholar] [CrossRef]
- Pignatti, S. Flora d’Italia; Edagricole: Bologna, Italy, 1982. [Google Scholar]
- Agusti, K.T. Therapeutic and medicinal values of onions and garlic. In Onions and Allied Crops; Rabinovitch, H.D., Ed.; Routledge: Milton Park, UK, 2017; pp. 99–104. [Google Scholar]
- Kim, J.H. Anti-bacterial action of onion (Allium cepa L.) extracts against oral pathogenic bacteria. J. Nihon Univ. Sch. Dent. 1997, 39, 136–141. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yin, M.C.; Cheng, W.S. Antioxidant activity of several Allium members. J. Agric. Food Chem. 1998, 46, 4097–4101. [Google Scholar] [CrossRef]
- Siegers, C.P.; Röbke, A.; Pentz, R. Effects of garlic preparations on superoxide production by phorbol ester activated granulocytes. Phytomedicine 1999, 6, 13–16. [Google Scholar] [CrossRef]
- Hobauer, R.; Frass, M.; Gmeiner, B.; Kaye, A.D.; Frost, E.A. Garlic extract (Allium sativum) reduces migration of neutrophils through endothelial cell monolayers. Middle East J. Anesthesiol. 2000, 15, 649–658. [Google Scholar] [PubMed]
- Sela, U.R.I.; Ganor, S.; Hecht, I.; Brill, A.; Miron, T.; Rabinkov, A.; Wilchek, M.; Mirelman, D.; Lider, O.; Hershkoviz, R. Allicin inhibits SDF–1α-induced T cell interactions with fibronectin and endothelial cells by down-regulating cytoskeleton rearrangement, Pyk-2 phosphorylation and VLA-4 expression. Immunology 2004, 111, 391–399. [Google Scholar] [CrossRef]
- Jin, P.; Kim, J.A.; Choi, D.Y.; Lee, Y.J.; Jung, H.S.; Hong, J.T. Anti-inflammatory and anti-amyloidogenic effects of a small molecule, 2, 4-bis (p-hydroxyphenyl)-2-butenal in Tg2576 Alzheimer’s disease mice model. J. Neuroinflamm. 2013, 10, 767. [Google Scholar] [CrossRef] [Green Version]
- Jeong, Y.Y.; Ryu, J.H.; Shin, J.H.; Kang, M.J.; Kang, J.R.; Han, J.; Kang, D. Comparison of anti-oxidant and anti-inflammatory effects between fresh and aged black garlic extracts. Molecules 2016, 21, 430. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abdel-Daim, M.M.; Abdel-Rahman, H.G.; Dessouki, A.A.; Ali, H.; Khodeer, D.M.; Bin-Jumah, M.; Alhader, M.S.; Alkahtani, S.; Aleya, L. Impact of garlic (Allium sativum) oil on cisplatin-induced hepatorenal biochemical and histopathological alterations in rats. Sci. Total Environ. 2020, 710, 136338. [Google Scholar] [CrossRef] [PubMed]
- Bayan, L.; Koulivand, P.H.; Gorji, A. Garlic: A review of potential therapeutic effects. Avicenna J. Phytomed. 2014, 4, 1. [Google Scholar]
- Iweala, E.E.J.; Akubugwo, E.I.; Okeke, C.U. Effects of ethanolic extracts of Allium sativum Linn. Liliaceae (Garlic) on serum cholestrol and blood sugar levels of albino rabbits. J. Plant Res. 2005, 9, 14–18. [Google Scholar]
- Qidwai, W.; Ashfaq, T. Role of garlic usage in cardiovascular disease prevention: An evidence-based approach. Evid. Based Complement. Altern. Med. 2013, 2013, 125649. [Google Scholar] [CrossRef] [PubMed]
- Patel, D.K.; Prasad, S.K.; Kumar, R.; Hemalatha, S. An overview on antidiabetic medicinal plants having insulin mimetic property. Asian Pac. J. Trop. Biomed. 2012, 2, 320–330. [Google Scholar] [CrossRef] [Green Version]
- Faroughi, F.; Mohammad-Alizadeh Charandabi, S.; Javadzadeh, Y.; Mirghafourvand, M. Effects of garlic pill on blood glucose level in borderline gestational diabetes mellitus: A triple blind, randomized clinical trial. Iran. Red Crescent Med. J. 2018, 20, 1–7. [Google Scholar] [CrossRef]
- Zhai, B.; Zhang, C.; Sheng, Y.; Zhao, C.; He, X.; Xu, W.; Huang, K.; Luo, Y. Hypoglycemic and hypolipidemic effect of S-allyl-cysteine sulfoxide (alliin) in DIO mice. Sci. Rep. 2018, 8, 3527. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Keophiphath, M.; Priem, F.; Jacquemond-Collet, I.; Clément, K.; Lacasa, D. 1, 2-vinyldithiin from garlic inhibits differentiation and inflammation of human preadipocytes. J. Nutr. 2009, 139, 2055–2060. [Google Scholar] [CrossRef]
- Apitz-Castro, R.; Escalante, J.; Vargas, R.; Jain, M.K. Ajoene, the antiplatelet principle of garlic, synergistically potentiates the antiaggregatory action of prostacyclin, forskolin, indomethacin and dypiridamole on human platelets. Thromb. Res. 1986, 42, 303–311. [Google Scholar] [CrossRef]
- Bordia, A.; Verma, S.K.; Srivastava, K.C. Effect of garlic (Allium sativum) on blood lipids, blood sugar, fibrinogen and fibrinolytic activity in patients with coronary artery disease. Prostaglandins Leukot. Essent. Fat. Acids 1998, 58, 257–263. [Google Scholar] [CrossRef]
- Ariga, T.; Tsuji, K.; Seki, T.; Moritomo, T.; Yamamoto, J.I. Antithrombotic and antineoplastic effects of phyto-organosulfur compounds. Biofactors 2000, 13, 251–255. [Google Scholar] [CrossRef] [PubMed]
- Ascrizzi, R.; Flamini, G. Leek or garlic? A chemical evaluation of elephant garlic volatiles. Molecules 2020, 25, 2082. [Google Scholar] [CrossRef] [PubMed]
- Rendu, F.; Brohard-Bohn, B.; Pain, S.; Bachelot-Loza, C.; Auger, J. Thiosulfinates inhibit platelet aggregation and microparticle shedding at a calpain-dependent step. J. Thromb. Haemost. 2001, 86, 1284–1291. [Google Scholar]
- Gruhlke, M.C.; Nicco, C.; Batteux, F.; Slusarenko, A.J. The effects of allicin, a reactive sulfur species from garlic, on a selection of mammalian cell lines. Antioxidants 2017, 6, 1. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gu, X.; Wu, H.; Fu, P. Allicin attenuates inflammation and suppresses HLA-B27 protein expression in ankylosing spondylitis mice. Biomed Res. Int. 2013, 2013, 171573. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gallwitz, H.; Bonse, S.; Martinez-Cruz, A.; Schlichting, I.; Schumacher, K.; Krauth-Siegel, R.L. Ajoene is an inhibitor and subversive substrate of human glutathione reductase and Trypanosoma cruzi trypanothione reductase: Crystallographic, kinetic, and spectroscopic studies. J. Med. Chem. 1999, 42, 364–372. [Google Scholar] [CrossRef] [PubMed]
- Zhen, H.; Fang, F.; Ye, D.Y.; Shu, S.N.; Zhou, Y.F.; Dong, Y.S.; Nie, X.C.; Li, G. Experimental study on the action of allitridin against human cytomegalovirus in vitro: Inhibitory effects on immediate-early genes. Antivir. Res. 2006, 72, 68–74. [Google Scholar] [CrossRef] [PubMed]
- Pai, S.T.; Platt, M.W. Antifungal effects of Allium sativum (garlic) extract against the Aspergillus species involved in otomycosis. Lett. Appl. Microbiol. 1995, 20, 14–18. [Google Scholar] [CrossRef]
- Naganawa, R.; Iwata, N.; Ishikawa, K.; Fukuda, H.; Fujino, T.; Suzuki, A. Inhibition of microbial growth by ajoene, a sulfur-containing compound derived from garlic. Appl. Environ. Microbiol. 1996, 62, 4238–4242. [Google Scholar] [CrossRef] [Green Version]
- Yoshida, H.; Iwata, N.; Katsuzaki, H.; Naganawa, R.; Ishikawa, K.; Fukuda, H.; Fujino, T.; Suzuki, A. Antimicrobial activity of a compound isolated from an oil-macerated garlic extract. Biosci. Biotechnol. Biochem. 1998, 62, 1014–1017. [Google Scholar] [CrossRef] [Green Version]
- Ankri, S.; Mirelman, D. Antimicrobial properties of allicin from garlic. Microbes Infect. 1999, 1, 125–129. [Google Scholar] [CrossRef]
- Yoshida, H.; Katsuzaki, H.; Ohta, R.; Ishikawa, K.; Fukuda, H.; Fujino, T.; Suzuki, A. Antimicrobial activity of the thiosulfinates isolated from oil-macerated garlic extract. Biosci. Biotechnol. Biochem. 1999, 63, 591–594. [Google Scholar] [CrossRef]
- De Amorim, A.M.B.; dos Santos Nascimento, J. Acinetobacter: An underrated foodborne pathogen? J. Infect. Dev. Ctries 2017, 11, 111–114. [Google Scholar] [CrossRef] [Green Version]
- Abdallah, M.; Benoliel, C.; Drider, D.; Dhulster, P.; Chihib, N.E. Biofilm formation and persistence on abiotic surfaces in the context of food and medical environments. Arch. Microbiol. 2014, 196, 453–472. [Google Scholar] [CrossRef]
- Camargo, A.C.; Woodward, J.J.; Call, D.R.; Nero, L.A. Listeria monocytogenes in food-processing facilities, food contamination, and human listeriosis: The Brazilian scenario. Foodborne Pathog. Dis. 2017, 14, 623–636. [Google Scholar] [CrossRef] [PubMed]
- Carter, M.Q.; Louie, J.W.; Feng, D.; Zhong, W.; Brandl, M.T. Curli fimbriae are conditionally required in Escherichia coli O157: H7 for initial attachment and biofilm formation. Food Microbiol. 2016, 57, 81–89. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haindongo, N.; Anyogu, A.; Ekwebelem, O.; Anumudu, C.; Onyeaka, H. Antibacterial and antibiofilm effects of garlic (Allium sativum), ginger (Zingiber officinale) and mint (Mentha piperita) on Escherichia coli biofilms. Appl. Food Biotechnol. 2021, 4, 166–176. [Google Scholar] [CrossRef]
- Caputo, L.; Amato, G.; Fratianni, F.; Coppola, R.; Candido, V.; De Feo, V.; Nazzaro, F. Chemical characterization and antibiofilm activities of bulbs and leaves of two aglione (Allium ampeloprasum var. holmense Asch. et Graebn.) landraces grown in Southern Italy. Molecules 2020, 25, 5486. [Google Scholar] [CrossRef] [PubMed]
- Council of Europe. European Pharmacopeia, 5th ed.; Council of Europe: Strasbourg, France, 2004. [Google Scholar]
- Jennings, W.; Shibamoto, T. Qualitative Analysis of Flavour and Fragrance Volatiles by Glass Capillary Gas Chromatography; Academic Press: New York, NY, USA, 1980. [Google Scholar]
- Davies, N.W. Gas chromatographic retention indices of monoterpenes and sesquiterpenes on methyl silicon and Carbowax 20M phases. J. Chromatogr. A 1990, 503, 1–24. [Google Scholar] [CrossRef]
- Adams, R.P. Identification of Essential Oil Components by Gas Chromatography/Mass Spectrometry; Allured Publishing Co.: Carol Stream, IL, USA, 2007. [Google Scholar]
- Sarker, S.D.; Nahar, L.; Kumarasamy, Y. Microtitre plate-based antibacterial assay incorporating resazurin as an indicator of cell growth, and its application in the in vitro antibacterial screening of phytochemicals. Methods 2007, 42, 321–324. [Google Scholar] [CrossRef]
- Kairo, S.K.; Bedwell, J.; Tyler, P.C.; Carter, A.; Corbel, M.J. Development of a tetrazolium salt assay for rapid determination of viability of BCG vaccines. Vaccine 1999, 17, 2423–2428. [Google Scholar] [CrossRef]
- Jirovetz, L.; Jäger, W.; Koch, H.P.; Remberg, G. Investigations of volatile constituents of the essential oil of Egyptian garlic (Allium sativum L.) by means of GC-MS and GC-FTIR. Z. Lebensm. Unters. Forsch. 1992, 194, 363–365. [Google Scholar] [CrossRef]
- Satyal, P.; Craft, J.D.; Dosoky, N.S.; Setzer, W.N. The chemical compositions of the volatile oils of garlic (Allium sativum) and wild garlic (Allium vineale). Foods 2017, 6, 63. [Google Scholar] [CrossRef] [Green Version]
- Amagase, H. Clarifying the real bioactive constituents of garlic. J. Nutr. 2006, 136, 716S–725S. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stoll, A.; Seebeck, E. Allium compounds. I. Alliin the true mother compound of garlic oil. Helv. Chim. Acta 1948, 31, 189–210. [Google Scholar] [CrossRef] [PubMed]
- Amagase, H.; Petesch, B.L.; Matsuura, H.; Kasuga, S.; Itakura, Y. Intake of garlic and its bioactive components. J. Nutr. 2001, 131, 955S–962S. [Google Scholar] [CrossRef] [PubMed]
- Saxena, P.; Joshi, Y.; Rawat, K.; Bisht, R. Biofilms: Architecture, resistance, quorum sensing and control mechanisms. Indian J. Microbiol. 2019, 59, 3–12. [Google Scholar] [CrossRef] [PubMed]
- Schwalm III, N.D.; Groisman, E.A. Navigating the gut buffet: Control of polysaccharide utilization in Bacteroides spp. Trends Microbiol. 2017, 25, 1005–1015. [Google Scholar] [CrossRef] [PubMed]
- Kumar, C.G.; Anand, S.K. Significance of microbial biofilms in food industry: A review. Int. J. Food Microbiol. 1998, 42, 9–27. [Google Scholar] [CrossRef]
- Bhatwalkar, S.B.; Gound, S.S.; Mondal, R.; Srivastava, R.K.; Anupam, R. Anti-biofilm and antibacterial activity of Allium sativum against drug resistant shiga-toxin producing Escherichia coli (STEC) isolates from patient samples and food Sources. Indian J. Microbiol. 2019, 59, 171–179. [Google Scholar] [CrossRef]
- Borlinghaus, J.; Albrecht, F.; Gruhlke, M.C.; Nwachukwu, I.D.; Slusarenko, A.J. Allicin: Chemistry and biological properties. Molecules 2014, 19, 12591–12618. [Google Scholar] [CrossRef] [Green Version]
- Rattanachaikunsopon, P.; Phumkhachorn, P. Diallyl sulfide content and antimicrobial activity against food-borne pathogenic bacteria of chives (Allium schoenoprasum). Biosci. Biotechnol. Biochem. 2008, 72, 2987–2991. [Google Scholar] [CrossRef] [Green Version]
- Li, G.; Ma, X.; Deng, L.; Zhao, X.; Wei, Y.; Gao, Z.; Jia, J.; Xu, J.; Sun, C. Fresh garlic extract enhances the antimicrobial activities of antibiotics on resistant strains in vitro. Jundishapur J. Microbiol. 2015, 8, 14814. [Google Scholar] [CrossRef] [Green Version]
- Møretrø, T.; Langsrud, S. Listeria monocytogenes: Biofilm formation and persistence in food-processing environments. Biofilms 2004, 1, 107–121. [Google Scholar] [CrossRef]
- Nazzaro, F.; Fratianni, F.; De Martino, L.; Coppola, R.; De Feo, V. Effect of essential oils on pathogenic bacteria. Pharmaceuticals 2013, 6, 1451–1474. [Google Scholar] [CrossRef] [PubMed]
- Jadhav, S.; Shah, R.; Bhave, M.; Palombo, E.A. Inhibitory activity of yarrow essential oil on Listeria planktonic cells and biofilms. Food Control 2014, 29, 125–130. [Google Scholar] [CrossRef]
- Sandasi, M.; Leonard, C.M.; Viljoen, A.M. The effect of five common essential oil components on Listeria monocytogenes biofilms. Food Control 2008, 19, 1070–1075. [Google Scholar] [CrossRef]
- Somrani, M.; Inglés, M.C.; Debbabi, H.; Abidi, F.; Palop, A. Garlic, onion, and cinnamon essential oil anti-biofilms’ effect against Listeria monocytogenes. Foods 2020, 9, 567. [Google Scholar] [CrossRef] [PubMed]
- Martins, N.; Petropoulos, S.; Ferreira, I.C. Chemical composition and bioactive compounds of garlic (Allium sativum L.) as affected by pre-and post-harvest conditions: A review. Food Chem. 2016, 211, 41–50. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zarrilli, R.; Giannouli, M.; Tomasone, F.; Triassi, M.; Tsakris, A. Carbapenem resistance in Acinetobacter baumannii: The molecular epidemic features of an emerging problem in health care facilities. J. Infect. Dev. Ctries 2009, 3, 335–341. [Google Scholar] [CrossRef]
- Perez, F.; Hujer, A.M.; Hujer, K.M.; Decker, B.K.; Rather, P.N.; Bonomo, R.A. Global challenge of multidrug-resistant Acinetobacter baumannii. Antimicrob. Agents Chemother. 2007, 51, 3471–3484. [Google Scholar] [CrossRef] [Green Version]
- Yang, X.; Sha, K.; Xu, G.; Tian, H.; Wang, X.; Chen, S.; Wang, Y.; Li, J.; Chen, J.; Huang, N. Subinhibitory concentrations of allicin decrease uropathogenic Escherichia coli (UPEC) biofilm formation, adhesion ability, and swimming motility. Int. J. Mol. Sci. 2016, 17, 979. [Google Scholar] [CrossRef] [Green Version]
- De Martino, L.; Amato, G.; Caputo, L.; Nazzaro, F.; Scognamiglio, M.R.; De Feo, V. Variations in composition and bioactivity of Ocimum basilicum cv ‘Aroma 2′ essential oils. Ind. Crops Prod. 2021, 172, 114068. [Google Scholar] [CrossRef]
- Caputo, L.; Smeriglio, A.; Trombetta, D.; Cornara, L.; Trevena, G.; Valussi, M.; Fratianni, F.; De Feo, V.; Nazzaro, F. Chemical composition and biological activities of the essential oils of Leptospermum petersonii and Eucalyptus gunnii. Front. Microbiol. 2020, 11, 409. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, H.; Li, S.; Cheng, Y. Antibiofilm Activity of Allicin and Quercetin in Treating Biofilm-Associated Orthopaedics Infection. Appl. Biochem. Biotechnol. 2022. [Google Scholar] [CrossRef] [PubMed]
- Lihua, L.; Jianhui, W.; Jialin, Y.; Yayin, L.; Guanxin, L. Effects of allicin on the formation of Pseudomonas aeruginosa biofilm and the production of quorum-sensing controlled virulence factors. Pol. J. Microbiol. 2013, 62, 243–251. [Google Scholar] [CrossRef] [PubMed]
- Jin, Z.; Li, L.; Zheng, Y.; An, P. Diallyl disulfide, the antibacterial component of garlic essential oil, inhibits the toxicity of Bacillus cereus ATCC 14579 at sub-inhibitory concentrations. Food Control 2021, 126, 108090. [Google Scholar] [CrossRef]
- Li, W.R.; Ma, Y.K.; Shi, Q.S.; Xie, X.B.; Sun, T.L.; Peng, H.; Huang, X.M. Diallyl disulfide from garlic oil inhibits Pseudomonas aeruginosa virulence factors by inactivating key quorum sensing genes. Appl. Microbiol. Biotechnol. 2018, 102, 7555–7564. [Google Scholar] [CrossRef]

{kind=link}
| Cultivars 1 | Species | Bulb Skin Colour | Clove Skin Colour | Floral Stem | Bulb Mean Weight | Bulb Equatorial Diameter | Cloves per Bulb | Clove Mean Weight |
|---|---|---|---|---|---|---|---|---|
| (g) | (mm) | (n.) | (g) | |||||
| ‘Rosso di Sulmona’ | A. sativum | white | red | yes | 41.2 (±0.8) b | 48.1 (±0.9) b | 11.8 (±0.9) a | 3.1 (±0.7) a |
| ‘Rosso di Spagna’ | A. sativum | cream | red | yes | 50.3 (±1.4) a | 54.6 (±1.0) a | 11.3 (±0.7) a | 3.7 (±0.9) a |
| ‘Irsina’ | A. ampeloprasum | cream | l. brown 2 | yes | 68.1 (±0.4) b | 75.1 (±0.9) b | 5.3 (±0.6) a | 11.4 (±0.2) b |
| ‘Contursi T.’ | A. ampeloprasum | cream | l. brown 2 | yes | 75.0 (±0.9) a | 84.2 (±1.1) a | 5.1 (±0.5) a | 12.1 (±0.4) a |
| % | |||||||
|---|---|---|---|---|---|---|---|
| ‘Irsina’ | ‘Contursi T.’ | ||||||
| N. | Aerial Parts | Bulbs | Aerial Parts | Bulbs | RT | KIa | |
| 1 | 2,4-Dimethylhexane | - | 22.3 | - | - | 5.0 | 758 |
| 2 | 3-Methylthiophene | - | 3.4 | - | - | 7.4 | 788 |
| 3 | 2,2-bis (Methylthio)-1-propanol | - | - | 1.7 | - | 9.2 | 812 |
| 4 | 2,6-Dimethylnonane | 0.1 | - | - | - | 11.3 | 838 |
| 5 | 2,3,5,8-Tetramethyldecane | T | - | - | 1.3 | 12.2 | 851 |
| 6 | 4-Methyl-1-undecene | 0.1 | - | - | 0.3 | 12.3 | 852 |
| 7 | Diallyl disulfide | - | - | 0.1 | 0.2 | 12.9 | 860 |
| 8 | 2-Hydroxyethyl-disulfide | T | - | - | - | 15.2 | 889 |
| 9 | Borneol | 0.1 | - | - | - | 15.5 | 893 |
| 10 | Terpinen-4-ol | T | - | - | - | 15.9 | 898 |
| 11 | Tridecane | - | - | - | 0.3 | 16.6 | 907 |
| 12 | 1,1-Thiobis-1-butine | - | - | - | 0.3 | 16.8 | 910 |
| 13 | Dimethyl sulfide | 0.1 | - | - | 0.4 | 17.5 | 919 |
| 14 | (Z)-Methyl propenyl disulfide | T | - | - | 0.3 | 17.9 | 925 |
| 15 | 2,6,10-Trimethyldodecane | T | - | - | 1.0 | 18.2 | 928 |
| 16 | Dodecyl sulfide | - | - | - | 0.1 | 18.5 | 932 |
| 17 | Dodecyl-7-en disulfide | T | - | - | 2.5 | 18.7 | 934 |
| 18 | Dodecyl-8-en disulfide | - | - | - | 0.4 | 18.9 | 937 |
| 19 | Methyl octane | - | - | - | 0.4 | 19.0 | 939 |
| 20 | Carvacrol | 4.1 | - | - | - | 19.2 | 941 |
| 21 | n-Heptene | - | - | - | 0.3 | 19.4 | 942 |
| 22 | 2-Methoxy-4-vinylphenol | - | - | - | 1.1 | 20.0 | 950 |
| 23 | Hexanal | 0.1 | - | - | 0.5 | 20.6 | 959 |
| 24 | (E)-Allyl propyl disulfide | 0.2 | - | - | 0.2 | 21.0 | 964 |
| 25 | (Z)- Allyl propyl disulfide | 0.1 | - | - | 0.1 | 22.1 | 979 |
| 26 | Hexanol | - | - | - | 0.8 | 22.2 | 979 |
| 27 | Octane | - | - | - | 1.2 | 22.4 | 982 |
| 28 | Decane disulfide | - | - | - | 0.8 | 23.1 | 992 |
| 29 | Geranyl isovalerate | - | - | - | 0.3 | 23.3 | 994 |
| 30 | Nonanal | 0.1 | - | - | 2.8 | 23.5 | 997 |
| 31 | Nonene | - | - | - | 3.3 | 24.0 | 1000 |
| 32 | Decene | 0.3 | - | - | 6.1 | 24.4 | 1005 |
| 33 | 2,4-Bis(1,1-dimethyl-ethyl)-phenol | 0.1 | - | - | 1.1 | 24.7 | 1009 |
| 34 | 2-Butyl-1-octanol | 0.3 | - | 0.1 | 1.6 | 25.0 | 1013 |
| 35 | Butyl octene | - | - | - | 2.2 | 25.2 | 1017 |
| 36 | n-Nonane | - | - | - | 1.0 | 26.8 | 1038 |
| 37 | (Z)-9-Ottadecene | 0.1 | - | - | - | 26.5 | 1033 |
| 38 | Propyl trisulfure | - | - | - | 1.1 | 27.4 | 1046 |
| 39 | 1,3,5-Trithiane | - | - | - | 2.1 | 27.8 | 1050 |
| 40 | Undecane | - | - | - | 0.8 | 28.4 | 1059 |
| 41 | Undecene | - | - | - | 0.9 | 28.6 | 1061 |
| 42 | Methyl propenyl trisulfide | - | - | - | 7.7 | 29.1 | 1068 |
| 43 | Methyl 12-methyltridecanoate | 0.8 | - | 0.3 | 0.5 | 29.5 | 1073 |
| 44 | Methyl triacontanoate | - | - | - | 0.7 | 29.6 | 1075 |
| 45 | Ethyl 2-oxo-tetradecanoate | T | - | - | - | 29.9 | 1079 |
| 46 | Methyl pentadecanoate | 0.7 | - | - | - | 30.8 | 1088 |
| 47 | trans-Methyl -3-pentil-undadecanoate | 0.1 | - | - | - | 31.2 | 1097 |
| 48 | Propenyl trisulfide | 0.1 | - | 3.2 | 2.8 | 31.8 | 1098 |
| 49 | Propyl allyl disulfide | 0.9 | - | 34.4 | 14.7 | 31.9 | 1100 |
| 50 | Methyl 14-methyl-pentadecanoate | 0.4 | - | - | - | 32.8 | 1114 |
| 51 | Methyl (Z)- 9-esadecanoate | 0.8 | - | - | - | 33.1 | 1116 |
| 52 | Methyl 11-esadecanoate | 0.9 | - | - | - | 33.4 | 1120 |
| 53 | 2-Hexyl-1-octanol | 0.1 | - | - | - | 33.5 | 1122 |
| 54 | Diallyl disulfide | 15.2 | 42.5 | - | - | 33.8 | 1126 |
| 55 | Propyl allyl trisulfide | 0.2 | - | - | - | 34.0 | 1129 |
| 56 | Methy 14-methyl-esadecanoate | 0.4 | - | - | - | 34.9 | 1143 |
| 57 | Methyl 2-Hexyl-cyclopropan-octanoate | 0.2 | - | - | - | 35.0 | 1144 |
| 58 | Methyl Heptadecanoate | 0.4 | - | - | 0.5 | 35.5 | 1151 |
| 59 | Methyl (Z)-9-octadecenoate | 0.1 | - | - | 0.6 | 36.2 | 1160 |
| 60 | Allicin | 57.3 | 29.8 | 53.1 | 8.6 | 37.0 | 1171 |
| 61 | Methyl allicin | 7.0 | - | 3.2 | - | 37.3 | 1176 |
| 62 | Diallyl trisulfide | 2.6 | - | - | 1.0 | 37.6 | 1182 |
| 63 | Methyl 8,11-ottadienoate | - | - | - | 1.2 | 37.9 | 1185 |
| 64 | Methyl 10-oxo-octadecanoate | - | - | - | 0.6 | 38.1 | 1187 |
| 65 | Methyl alliy trisulfide | - | - | - | 0.2 | 38.3 | 1191 |
| 66 | Methyl diallyl trisulfide | 0.7 | - | - | 0.3 | 38.4 | 1192 |
| 67 | Ethyl allyl trisulfide | - | - | - | 0.1 | 39.3 | 1199 |
| 68 | Ethyl diallyl trisulfide | 1.9 | - | 1.5 | 2.9 | 39.4 | 1199 |
| 69 | Vinyl diallyl trisulfide | 0.1 | - | - | 2.7 | 40.8 | 1221 |
| 70 | Propenyl trisulfide | 0.2 | - | - | 1.1 | 41.3 | 1229 |
| 71 | Heptadecan trisulfide | - | - | - | 1.8 | 41.7 | 1235 |
| 72 | Di-tert-dodecyl disulfide | T | - | - | - | 42.0 | 1241 |
| 73 | Octadecan trisulfide | - | - | - | 0.2 | 42.4 | 1247 |
| 74 | Pentadecan tetrasulfide | - | - | 0.3 | 0.9 | 43.4 | 1262 |
| 75 | Methyl esacosanoate | T | - | - | 0.4 | 43.9 | 1269 |
| 76 | Methyl 9,12-epithio-9,11-octadecanoate | - | - | - | 2.7 | 44.2 | 1274 |
| 77 | Diallyl tetrasulfide | - | - | - | 2.6 | 44.9 | 1284 |
| 78 | Propyl allyl tetrasulfide | - | - | - | 0.9 | 46.4 | 1300 |
| 79 | Methyl tetracosanoate | - | - | - | 1.1 | 46.8 | 1307 |
| 80 | Propyl 3-(octadeciloxi)-oleate | - | - | - | 0.5 | 47.1 | 1313 |
| 81 | Propyl pentyl tetrasulfide | - | - | - | 2.0 | 47.3 | 1315 |
| 82 | Cyclo octasulfide | - | - | - | 2.7 | 50.1 | 1360 |
| Total | 96.9 | 98.0 | 97.9 | 97.8 | |||
| Rosso di Sulmona | Rosso di Spagna | ||||||
|---|---|---|---|---|---|---|---|
| % | |||||||
| N. | Aerial Parts | Bulbs | Aerial Parts | Bulbs | RT | KIa | |
| 1 | 2,4-Dimethylhexane | - | 1.7 | 17.1 | 0.1 | 5.0 | 757 |
| 2 | 3,31-Thiobis-1-propane | - | - | - | T | 6.4 | 776 |
| 3 | 3-Methyl-thiophene | - | - | - | T | 7.4 | 788 |
| 4 | 2,3-Dimethyl- thiophene | - | - | - | T | 7.7 | 792 |
| 5 | Methyl-2-propenyl-disulfide | - | - | - | T | 8.1 | 796 |
| 6 | α-Pinene | - | - | - | T | 8.5 | 803 |
| 7 | 2,2-Bis (Methylthio)-1-propanol | - | - | - | 0.6 | 9.2 | 812 |
| 8 | (-)-β-Pinene | - | - | - | T | 9.9 | 819 |
| 9 | 2,6-Dimethylnonane | - | - | - | T | 11.2 | 837 |
| 10 | D-Limonene | - | - | - | T | 11.4 | 840 |
| 11 | 1,1-Dimetoxi-cyclohexane | - | - | - | T | 12,2 | 850 |
| 12 | 2,3,5,8-Tetramethyl-decane | 0.7 | - | - | T | 12.3 | 851 |
| 13 | 4-Methyl-1-undecene | - | - | - | T | 12.4 | 853 |
| 14 | Butyl propenyl sulfide | - | - | - | T | 12.5 | 855 |
| 15 | Diallyl disulfide | - | - | - | 0.2 | 12.9 | 859 |
| 16 | 4-Etenyl-1,2-dimethyl-benzene | - | - | - | T | 14.3 | 877 |
| 17 | Allyl-1-propenyl sulfide | - | - | - | T | 14.7 | 882 |
| 18 | 9-Hydroxyethyl-ethyl-disulfide | - | - | - | 0.1 | 15.1 | 888 |
| 19 | 2- Hydroxyethyl- disulfide | 0.2 | - | - | - | 15.2 | 889 |
| 20 | Benzyl methyl sulphide | - | - | - | T | 15.3 | 889 |
| 21 | 3,4-Dimethyl-thiophene | - | - | - | 0.1 | 15.4 | 891 |
| 22 | 2-Ethyl-5-[(2-ethylbuthyl) thio]-thiophene | 0.1 | - | - | 0.2 | 16.1 | 901 |
| 23 | Bis(1,1-dimethylpropyl) -disulfide | - | - | - | 0.1 | 16.4 | 905 |
| 24 | Tridecane | 0.4 | - | - | - | 16.6 | 907 |
| 25 | 1,1-Thiobis-1-butine | - | - | - | T | 16.9 | 910 |
| 27 | Dimethyl disulfide | 0.2 | - | - | 0.1 | 17.6 | 921 |
| 28 | (Z)-Methyl propenyl disulfide | 2.9 | - | - | T | 17.9 | 925 |
| 29 | (E)- Methyl propenyl disulfide | - | - | - | T | 18.0 | 926 |
| 30 | 2,6,10-Trimethyl-dodecane | - | - | - | T | 18.2 | 928 |
| 31 | Dodecyl sulfide | - | - | - | T | 18.5 | 931 |
| 32 | Dodecyl-7-en disulfide | - | - | - | T | 18.6 | 934 |
| 33 | Dodecyl-8-en disulfide | - | - | - | T | 18.9 | 937 |
| 34 | 2-Methoxy-4-vinylphenol | - | - | - | 0.1 | 19.8 | 948 |
| 35 | (E)-Allyl propyl disulfide | 0.4 | - | - | 0.6 | 21.0 | 965 |
| 36 | (Z)-Allyl propyl disulfide | 3.9 | - | - | 0.3 | 22.1 | 979 |
| 37 | Methyl 9-oxo-nonanoate | - | - | - | T | 22.8 | 983 |
| 38 | Decane disulphide | - | - | - | T | 23.1 | 992 |
| 39 | Geranyl isovalerate | - | - | - | T | 23.3 | 994 |
| 40 | Nonene | 1.1 | - | 2.7 | - | 24 | 1000 |
| 41 | Decene | 1.3 | - | 0.4 | - | 24.2 | 1002 |
| 42 | 2,4-Bis(1,1-dimethyl-ethyl) -phenol | 1.1 | - | 0.7 | 0.3 | 24.7 | 1009 |
| 43 | 2-Butyl-1-octanol | 1.5 | - | - | 0.6 | 25 | 1013 |
| 44 | (E)-9-Octadecene | 0.2 | - | - | T | 26.5 | 1033 |
| 45 | 4-Methyl-1-undecene | 0.5 | - | - | 0.1 | 26.5 | 1034 |
| 46 | 1,3,5-Trithiane | 2.1 | - | - | 0.2 | 27.8 | 1051 |
| 47 | Methyl propenyl trisulfide | - | - | 0.6 | 3.4 | 28.8 | 1064 |
| 48 | Methyl 12-methyl-tridecanoate | 2.6 | - | - | 0.2 | 29.5 | 1073 |
| 49 | Methyl triacontanoate | - | - | - | 0.1 | 29.6 | 1074 |
| 50 | Ethyl 2-oxo-tetradecanoate | - | - | - | T | 30.0 | 1079 |
| 51 | Methyl pentadecanoate | - | - | - | 0.1 | 30.8 | 1091 |
| 52 | Methyl 12-methyl-tetradecanoate | - | - | - | 0.2 | 31.0 | 1093 |
| 53 | Methyl trans-3-pentil-oxiran-undecanoate | - | - | - | 0.3 | 31.6 | 1094 |
| 54 | Propenyl trisulfide | 0.8 | - | - | 0.2 | 31.8 | 1098 |
| 55 | Propyl allyl disulfide | - | - | - | 0.1 | 31.9 | 1099 |
| 56 | Propyl allyl trisulphide | 30.6 | - | 4.9 | - | 32.0 | 1100 |
| 57 | Vinyl trisulphide | - | - | - | 0.1 | 32.5 | 1108 |
| 58 | Methyl 14-methyl-pentadecanoate | - | - | - | 0.2 | 3.,8 | 1113 |
| 59 | Methyl (Z)-9-esadecanoate | - | - | - | 0.7 | 33.1 | 1116 |
| 60 | Diallyl disulfide | - | 34.5 | 48.5 | 12.6 | 33.7 | 1125 |
| 61 | Methyl 2-hexyl-cyclopropanoctanoate | - | - | - | 0.2 | 35.1 | 1144 |
| 62 | Tridecan trisulfide | 0.3 | - | - | 0.2 | 35.5 | 1151 |
| 63 | Sulfide cyclicoctatomic | - | - | - | T | 35.9 | 1156 |
| 64 | Methyl (Z)-11-octadecenoate | - | - | - | T | 36.2 | 1160 |
| 65 | Allicin | 36.8 | 61.8 | 21.1 | 52.9 | 36.9 | 1170 |
| 66 | Methyl allicin | 9.8 | - | 1.8 | - | 37.4 | 1177 |
| 67 | Methyl 8,11-octadecadienoate | - | - | - | 4.3 | 37.7 | 1182 |
| 68 | Diallyl trisulfide | - | - | - | 5.4 | 37.8 | 1183 |
| 69 | Methyl diallyl trisulfide | 0.4 | - | - | 0.8 | 39.0 | 1194 |
| 70 | Ethyl diallyl trisulfide | 0.1 | - | - | 5.1 | 39.6 | 1203 |
| 71 | Vinyl diallyl trisulfide | - | - | - | 0.2 | 40.8 | 1222 |
| 72 | Heptadecan trisulfide | - | - | - | 0.5 | 41.6 | 1234 |
| 73 | Di-tert-dodecyl disulfide | 0.2 | - | - | 0.8 | 42.0 | 1241 |
| 74 | Octadecan trisulfide | - | - | - | 1.7 | 42.5 | 1248 |
| 75 | Tridecan tetrasulfide | - | - | - | 0.1 | 43.1 | 1258 |
| 76 | Pentadecan tetrasulfide | - | - | - | 0.6 | 43.2 | 1262 |
| 77 | Methyl exacosanoate | - | - | - | 0.1 | 43.9 | 1269 |
| 78 | Methyl 9,12-epithio-9,11-octadecanoate | - | - | - | 0.7 | 44.0 | 1270 |
| 79 | Diallyl tetrasulfide | - | - | - | 0.3 | 44.9 | 1282 |
| 80 | Propyl allyl tetrasulfide | - | - | - | 0.1 | 45.4 | 1292 |
| 81 | Methyl triacontanoate | - | - | 0.4 | 0.3 | 45.5 | 1294 |
| 82 | Methyl tetracosanoate | - | - | - | 0.1 | 46.8 | 1308 |
| 83 | Propyl 3-octadeciloxi-oleate | - | - | - | 0.4 | 47.1 | 1312 |
| Total | 98.2 | 98.0 | 98.2 | 96.7 | |||
| A. baumannii | E. coli | L. monocytogenes | S. aureus | ||
|---|---|---|---|---|---|
| ‘Irsina’ | Aerial parts | 30 ± 2 | 40 a ± 3 | 30 ± 3 | 30 ± 2 |
| Bulbs | 30 ± 3 | 30 c ± 3 | 30 ± 2 | 30 ± 3 | |
| “Contursi T.” | Aerial parts | 30 ± 3 | 28 ± 2 | 40 ± 3 | 28 ± 2 |
| Bulbs | 40 b ± 2 | 35 a ± 3 | 30 ± 2 | 30 ± 2 | |
| Tetracycline | 31 ± 1 | 24 ± 3 | 39 ± 2 | 38 ± 2 |
| A. baumannii | E. coli | L. monocytogenes | S. aureus | ||
|---|---|---|---|---|---|
| ‘Rosso di Sulmona’ | Aerial parts | 30 ± 2 | 40 a ± 3 | 30 ± 3 | 40 ± 2 |
| Bulbs | 30 ± 2 | 30 b ± 3 | 30 ± 3 | 40 ± 2 | |
| ‘Rosso di Spagna’ | Aerial parts | 30 ± 4 | 30 b ± 2 | 30 ± 4 | 30 ± 2 |
| Bulbs | 30 ± 2 | 35 a ± 2 | 30 ± 3 | 28 ± 3 | |
| Tetracycline | 31 ± 1 | 24 ± 3 | 39 ± 2 | 38 ± 2 |
| A. baumannii | E. coli | L. monocytogenes | S. aureus | ||
|---|---|---|---|---|---|
| ‘Irsina’ | Aerial parts 10 µL/mL | 47.18 a ± 1.59 | 0 | 1.23 ± 0.18 | 0 |
| Aerial parts 20 µL/mL | 72.68 a ± 1.42 | 22.58 a ± 0.93 | 79.85 a ± 1.05 | 57.97 a ± 1.11 | |
| Bulbs 10 µL/mL | 7.35 a ± 1.73 | 18.82 a ± 3.08 | 52.58 a ± 1.25 | 41.96 a ± 1.15 | |
| Bulbs 20 µL/mL | 52.83 a ± 1.14 | 45.95 a ± 0.81 | 63.24 a ± 1.72 | 50.17 a ± 0.82 | |
| ‘Contursi T.’ | Aerial parts 10 µL/mL | 0 | 47.55 a ± 3.51 | 0 | 50.54 a ± 0.71 |
| Aerial parts 20 µL/mL | 45.86 a ± 1.31 | 81.88 a ± 1.21 | 0 | 73.47 a ± 1.91 | |
| Bulbs 10 µL/mL | 0 | 0 | 20.65 a ± 3.2 | 11.11 a ± 1.8 | |
| Bulbs 20 µL/mL | 0 | 25.39 a ± 1.28 | 40.69 a ± 1,57 | 61.41 a ± 1.61 | |
| ‘Rosso di Sulmona’ | Aerial parts 10 µL/mL | 0 | 12.21 a ± 1.91 | 46.06 a ± 1.83 | 25.52 a ± 1.59 |
| Aerial parts 20 µL/mL | 61.76 a ± 3.17 | 36.31 a ± 1.47 | 64.11 a ± 0.74 | 33.48 a ± 2.16 | |
| Bulbs 10 µL/mL | 12.25 a ± 2.35 | 24.73 a ± 1.76 | 37.60 a ± 1.40 | 0 | |
| Bulbs 20 µL/mL | 48.55 a ± 1.52 | 41.20 a ± 3.37 | 42.03 a ± 0.54 | 1.12 ± 0.13 | |
| ‘Rosso di Spagna’ | Aerial parts 10 µL/mL | 46.08 a ± 2.56 | 41.56 a ± 3.34 | 40.98 a ± 4.32 | 9.47 a ± 0.67 |
| Aerial parts 20 µL/mL | 49.91 a ± 2.75 | 54.09 a ± 1.77 | 46.88 a ± 0.71 | 44.39 a ± 1.44 | |
| Bulbs 10 µL/mL | 26.62 a ± 3.02 | 0 | 44.84 a ± 4.64 | 15.25 a ± 0.38 | |
| Bulbs 20 µL/mL | 56.97 a ± 1.73 | 27.16 a ± 1.66 | 61.22 a ± 2.09 | 70.29 a ± 0.14 |
| A. baumannii | E. coli | L. monocytogenes | S. aureus | ||
|---|---|---|---|---|---|
| ‘Irsina’ | Aerial parts 10 µL/mL | 0 | 18.26 a ± 1.52 | 0 | 0 |
| Aerial parts 20 µL/mL | 30.13 a ± 0.28 | 71.08 a ± 2.47 | 31.43 a ± 0.31 | 55.99 a ± 1.1 | |
| Bulbs 10 µL/mL | 0 | 0 | 0 | 39.17 a ± 1.15 | |
| Bulbs 20 µL/mL | 14.48 a ± 0.13 | 0 | 30.25 a ± 0.68 | 47.81 a ± 0.82 | |
| ‘Contursi T.’ | Aerial parts 10 µL/mL | 76.15 a ± 0.91 | 63.86 a ± 2.13 | 8.65 a ± 0.68 | 6.55 a ± 0.23 |
| Aerial parts 20 µL/mL | 89.47 a ± 0.86 | 76.71 a ± 0.97 | 24.58 a ± 1.37 | 36.07 a ± 2.32 | |
| Bulbs 10 µL/mL | 69.28 a ± 1.37 | 65.08 a ± 0.99 | 8.46 a ± 0.55 | 7.97 a ± 0.94 | |
| Bulbs 20 µL/ml | 81.14 a ± 0.27 | 79.15 a ± 0.43 | 25.28 a ± 0.88 | 36.19 a ± 2.18 | |
| ‘Rosso di Sulmona’ | Aerial parts 10 µL/mL | 0 | 0 | 20.67 a ± 1.67 | 22.07 a ± 1.59 |
| Aerial parts 20 µL/mL | 16.94 a ± 1.13 | 0 | 68.57 a ± 0.89 | 30.34 a ± 2.16 | |
| Bulbs 10 µL/mL | 36.46 a ± 0.68 | 0 | 8.19 a ± 1.43 | 0 | |
| Bulbs 20 µL/mL | 65.16 a ± 0.85 | 0 | 11.13 a ± 1.65 | 0 | |
| ‘Rosso di Spagna’ | Aerial parts 10 µL/mL | 6.11 a ± 0.67 | 0 | 6.58 a ± 2.14 | 5.19 a ± 0.67 |
| Aerial parts 20 µL/mL | 25.75 a ± 0.69 | 18.27 a ± 0.66 | 60.20 a ± 2.06 | 41.77 a ± 1.44 | |
| Bulbs 10 µL/mL | 0 | 0 | 7.09 a ± 1.33 | 11.26 a ± 0.38 | |
| Bulbs 20 µL/mL | 45.88 a ± 1.50 | 4.27 a ± 0.69 | 13.61 a ± 1.65 | 68.89 a ± 1.45 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Polito, F.; Amato, G.; Caputo, L.; De Feo, V.; Fratianni, F.; Candido, V.; Nazzaro, F. Chemical Composition and Agronomic Traits of Allium sativum and Allium ampeloprasum Leaves and Bulbs and Their Action against Listeria monocytogenes and Other Food Pathogens. Foods 2022, 11, 995. https://doi.org/10.3390/foods11070995
Polito F, Amato G, Caputo L, De Feo V, Fratianni F, Candido V, Nazzaro F. Chemical Composition and Agronomic Traits of Allium sativum and Allium ampeloprasum Leaves and Bulbs and Their Action against Listeria monocytogenes and Other Food Pathogens. Foods. 2022; 11(7):995. https://doi.org/10.3390/foods11070995
Chicago/Turabian StylePolito, Flavio, Giuseppe Amato, Lucia Caputo, Vincenzo De Feo, Florinda Fratianni, Vincenzo Candido, and Filomena Nazzaro. 2022. "Chemical Composition and Agronomic Traits of Allium sativum and Allium ampeloprasum Leaves and Bulbs and Their Action against Listeria monocytogenes and Other Food Pathogens" Foods 11, no. 7: 995. https://doi.org/10.3390/foods11070995
APA StylePolito, F., Amato, G., Caputo, L., De Feo, V., Fratianni, F., Candido, V., & Nazzaro, F. (2022). Chemical Composition and Agronomic Traits of Allium sativum and Allium ampeloprasum Leaves and Bulbs and Their Action against Listeria monocytogenes and Other Food Pathogens. Foods, 11(7), 995. https://doi.org/10.3390/foods11070995