
Effects of Selenium Nanoparticles on Preventing Patulin-Induced Liver, Kidney and Gastrointestinal Damage

Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals and Reagents
2.2. SeNPs Preparation and Characterization
2.3. In Vitro Antioxidant Activity
2.4. Cell Culture and Treatments
2.5. Animal Experiments
2.5.1. Animals and Treatments
2.5.2. Biochemical Parameters Determination
2.6. Statistical Analysis
3. Results
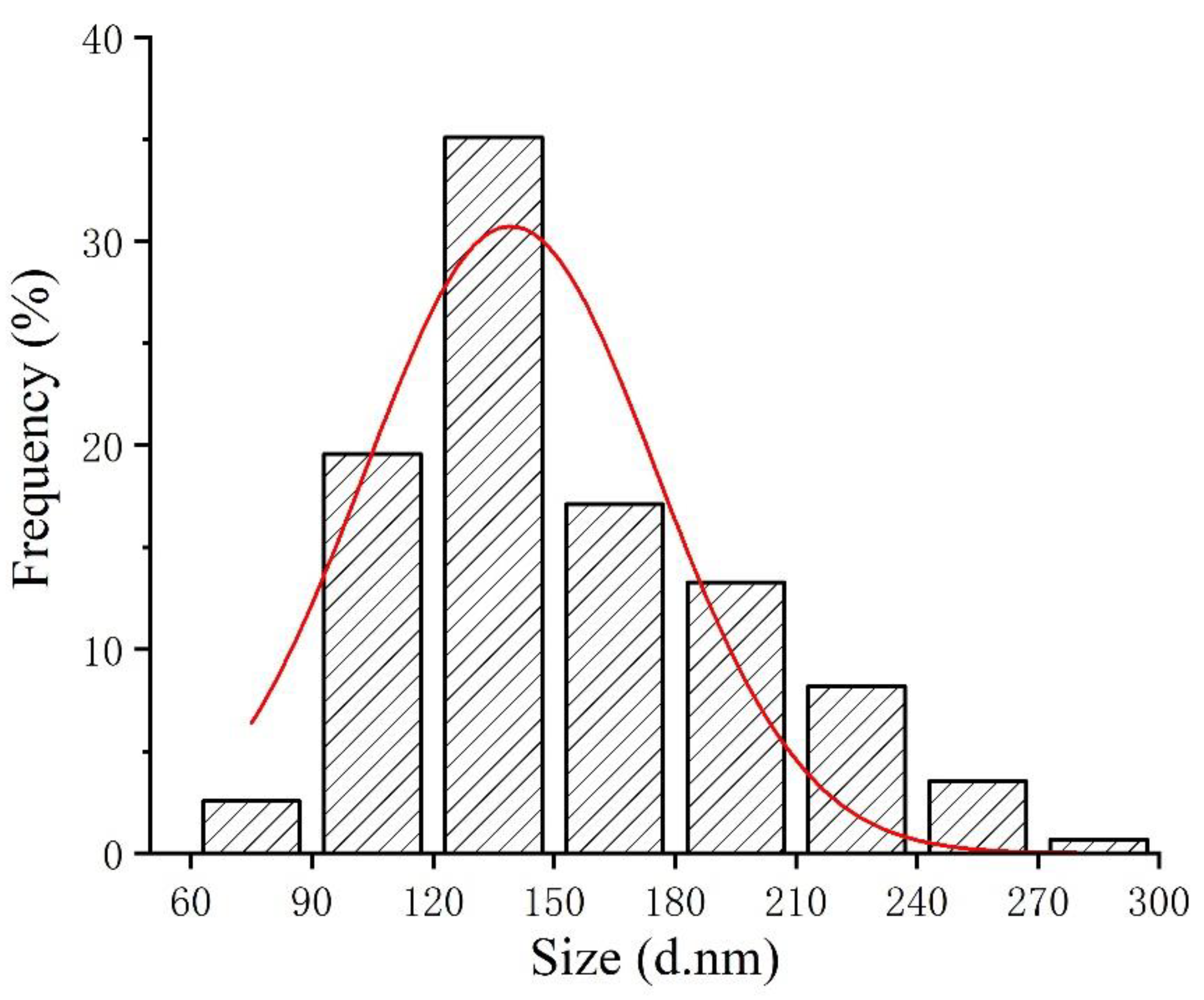
3.1. Characterization of SeNPs
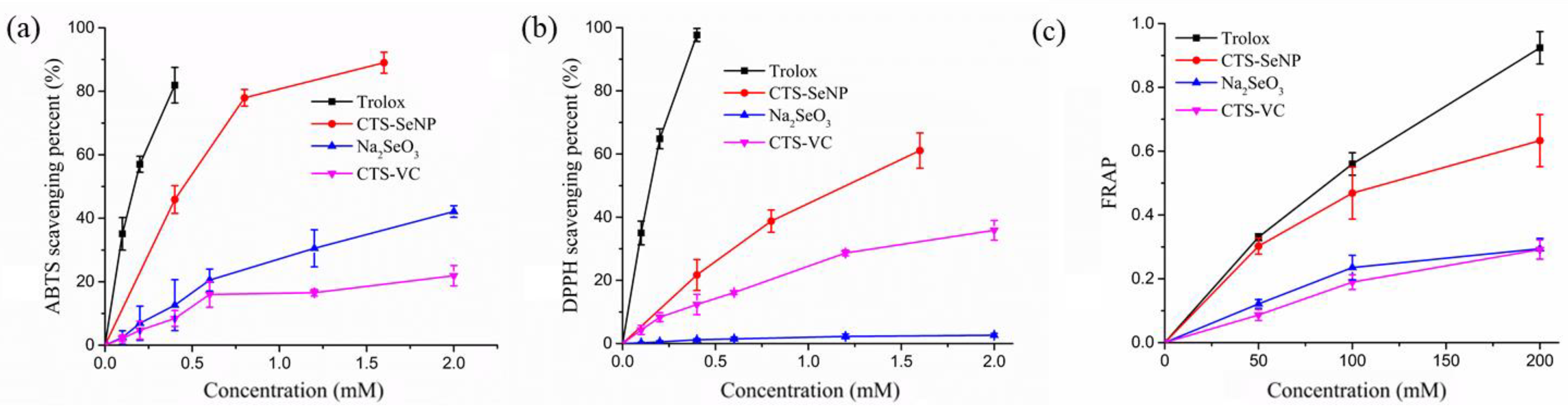
3.2. In Vitro SeNPs Antioxidant Activities
3.3. Effect of SeNPs on PAT-Induced Cytotoxicity
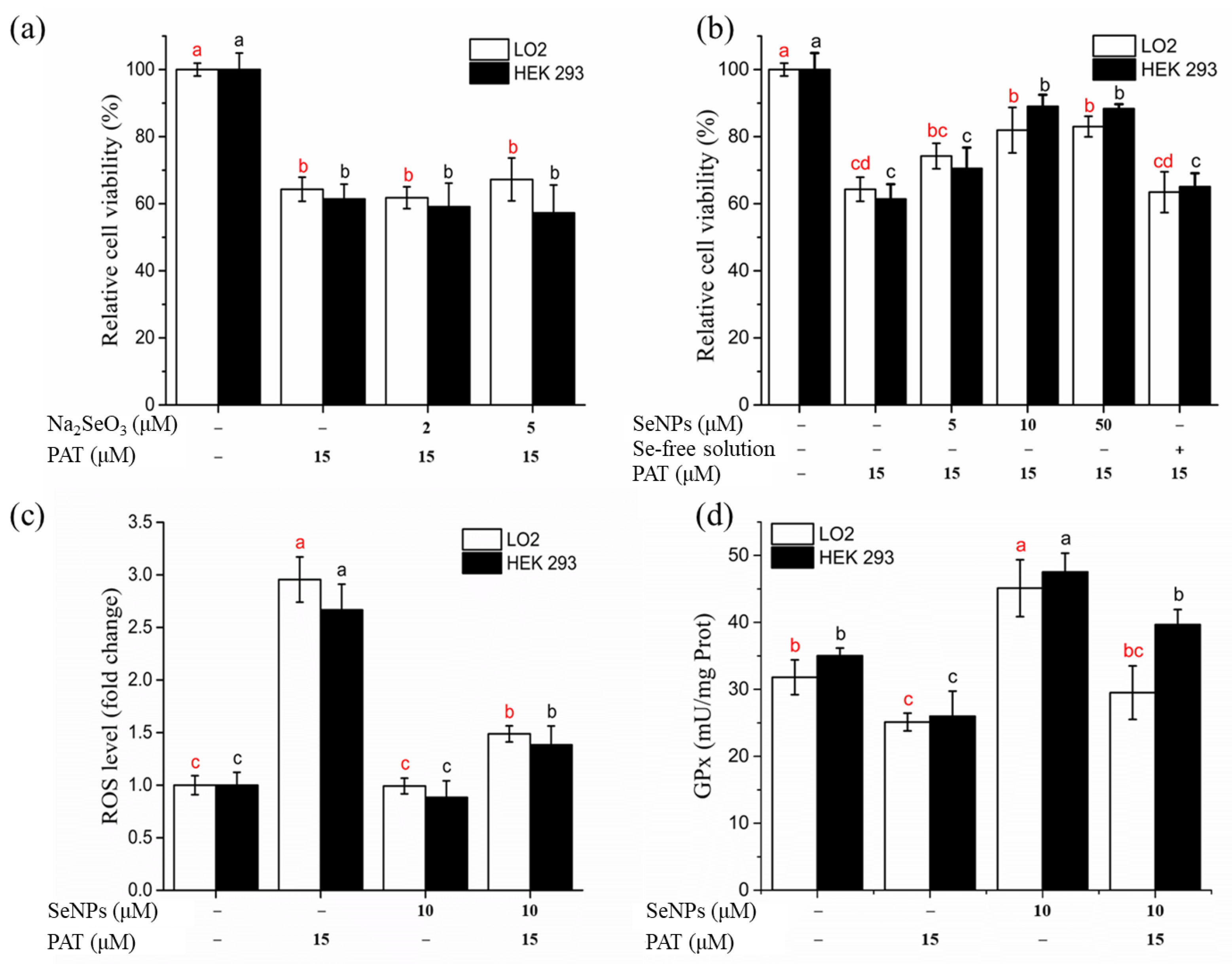
3.3.1. Cytotoxicity of Prepared SeNPs
3.3.2. Effect of SeNPs on PAT-Mediated Cytotoxicity
3.4. Effects of SeNPs on Liver and Kidney Injury Induced by PAT in Mouse Model
3.4.1. Serum Biochemistry Parameters
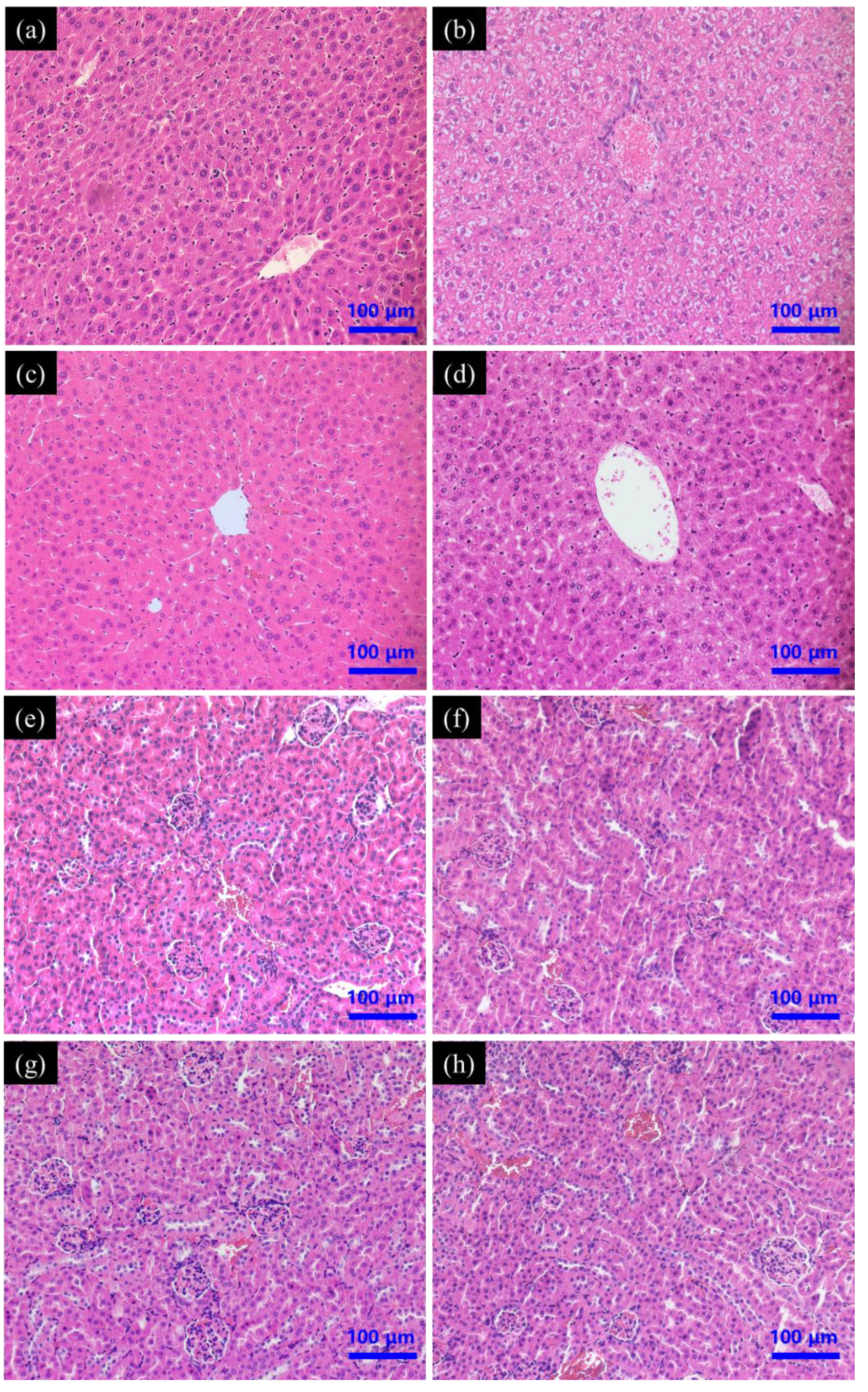
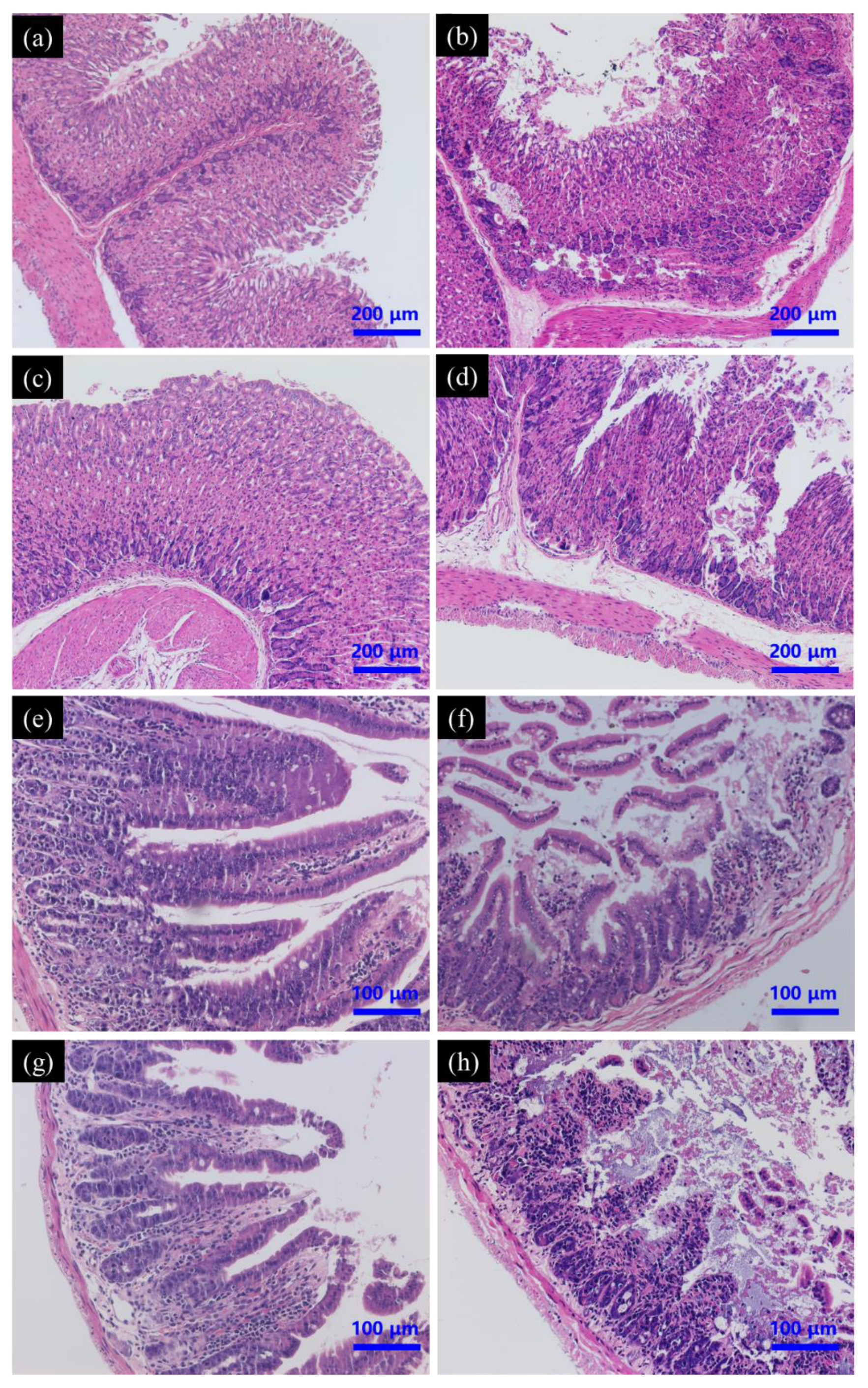
3.4.2. Histopathology of Liver and Kidney
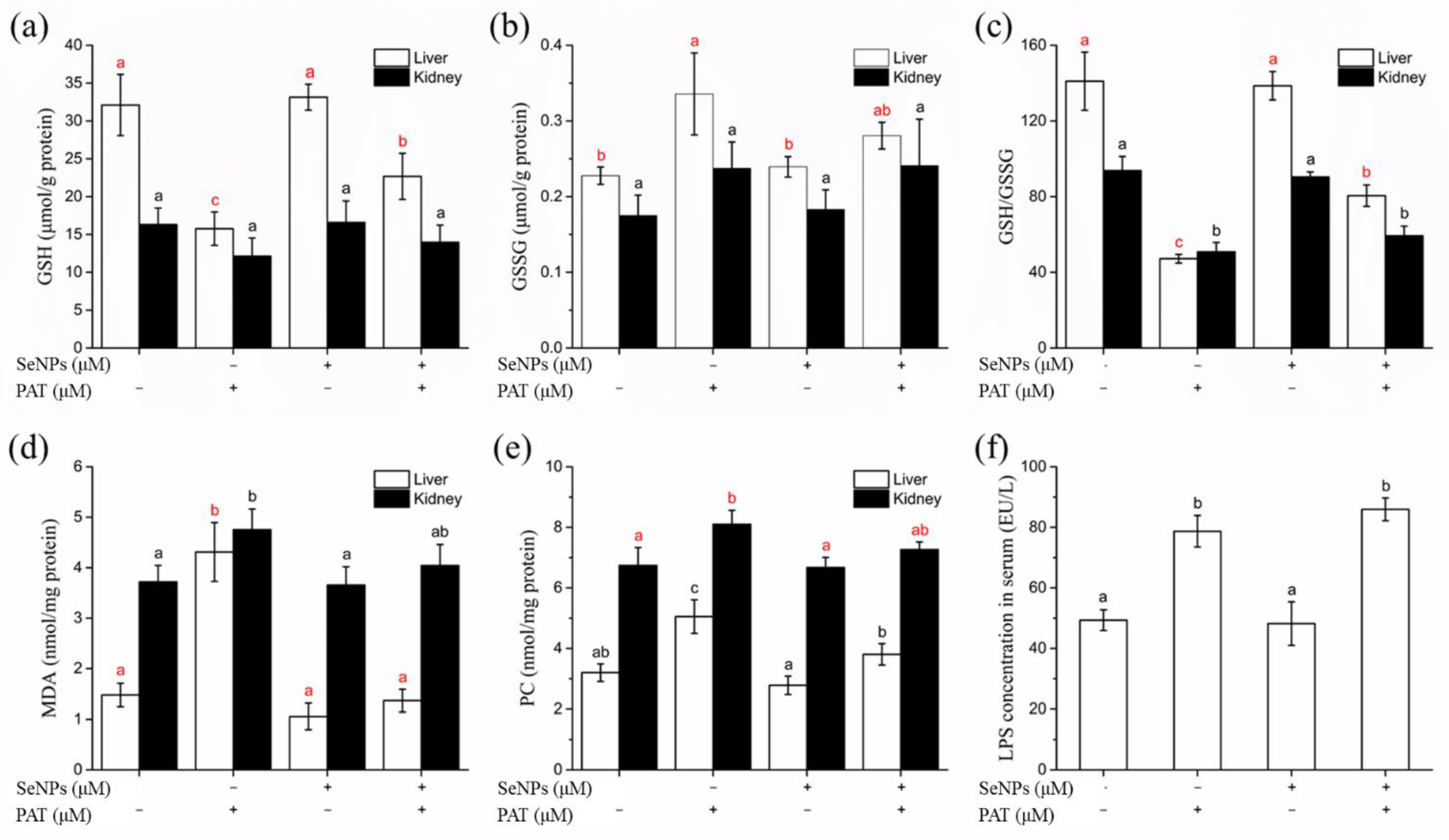
3.4.3. Oxidative Stress Biomarkers of Liver and Kidney
3.5. Effect of SeNPs on PAT-Induced Gastrointestinal Damage
3.5.1. Histopathology of Gastrointestinal Tract
3.5.2. LPS Level
3.5.3. Oxidative Stress Biomarkers of Gastrointestinal Tissues
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations and Acronyms
| BUN | urea nitrogen |
| CAT | catalase |
| CCK 8 | Cell Counting Kit-8 |
| CTS | chitosan |
| DPPH | 2,2-Diphenyl-1-picrylhydrazyl |
| FRAP | ferric reducing antioxidant potential |
| GPx | glutathione peroxidase |
| GR | glutathione reductase |
| GSH | glutathione |
| GSSG | glutathione disulfide |
| GST | glutathione S-transferase |
| LPS | lipopolysaccharides |
| MDA | malondialdehyde |
| MSeA | methylseleninic acid |
| Na2SeO3 | sodium selenite |
| NADPH | nicotinamide adenine dinucleotide phosphate |
| PAT | patulin |
| PC | protein carbonyls |
| ROS | reactive oxygen species |
| Se | selenium |
| SeMet | selenomethionine |
| SeNPs | selenium nanoparticles |
| SOD | dismutase |
| UA | uric acid |
| VC | Vitamin C/ascorbic acid |
References
- Ngolong Ngea, G.L.; Yang, Q.; Castoria, R.; Zhang, X.; Routledge, M.N.; Zhang, H. Recent trends in detecting, controlling, and detoxifying of patulin mycotoxin using biotechnology methods. Compr. Rev. Food Sci. F 2020, 19, 2447–2472. [Google Scholar] [CrossRef] [PubMed]
- Vidal, A.; Ouhibi, S.; Ghali, R.; Hedhili, A.; De Saeger, S.; De Boevre, M. The mycotoxin patulin: An updated short review on occurrence, toxicity and analytical challenges. Food Chem. Toxicol. 2019, 129, 249–256. [Google Scholar] [CrossRef] [PubMed]
- Sohrabi, H.; Arbabzadeh, O.; Khaaki, P.; Khataee, A.; Majidi, M.R.; Orooji, Y. Patulin and Trichothecene: Characteristics, occurrence, toxic effects and detection capabilities via clinical, analytical and nanostructured electrochemical sensing/biosensing assays in foodstuffs. Crit. Rev. Food Sci. 2021, 1–29. [Google Scholar] [CrossRef]
- Ramalingam, S.; Bahuguna, A.; Kim, M. The effects of mycotoxin patulin on cells and cellular components. Trends Food Sci. Technol. 2019, 83, 99–113. [Google Scholar] [CrossRef]
- Saleh, I.; Goktepe, I. The characteristics, occurrence, and toxicological effects of patulin. Food Chem. Toxicol. 2019, 129, 301–311. [Google Scholar] [CrossRef]
- Han, J.; Jin, C.; Zhong, Y.; Zhu, J.; Liu, Q.; Sun, D.; Feng, J.; Xia, X.; Peng, X. Involvement of NADPH oxidase in patulin-induced oxidative damage and cytotoxicity in HEK293 cells. Food Chem. Toxicol. 2021, 150, 112055. [Google Scholar] [CrossRef]
- Suzuki, T.; Iwahashi, Y. Gene expression profiles of yeast Saccharomyces cerevisiae sod1 caused by patulin toxicity and evaluation of recovery potential of ascorbic acid. J. Agric. Food Chem. 2011, 59, 7145–7154. [Google Scholar] [CrossRef] [PubMed]
- Ayed Boussema, I.; Abassi, H.; Bouaziz, C.; Hlima, W.B.; Ayed, Y.; Bacha, H. Antioxidative and antigenotoxic effect of vitamin E against patulin cytotoxicity and genotoxicity in HepG2 cells. Environ. Toxicol. 2013, 28, 299–306. [Google Scholar] [CrossRef]
- Yang, G.; Zhong, L.; Jiang, L.; Geng, C.; Cao, J.; Sun, X.; Liu, X.; Chen, M.; Ma, Y. 6-Gingerol prevents patulin-induced genotoxicity in HepG2 cells. Phytother. Res. 2011, 25, 1480–1485. [Google Scholar] [CrossRef]
- Jayashree, G.V.; Krupashree, K.; Rachitha, P.; Khanum, F. Patulin induced oxidative stress mediated apoptotic damage in mice, and its modulation by green tea leaves. J. Clin. Exp. Hepatol. 2017, 7, 127–134. [Google Scholar] [CrossRef]
- Zhong, Y.; Jin, C.; Gan, J.; Wang, X.; Shi, Z.; Xia, X.; Peng, X. Apigenin attenuates patulin-induced apoptosis in HEK293 cells by modulating ROS-mediated mitochondrial dysfunction and caspase signal pathway. Toxicon 2017, 137, 106–113. [Google Scholar] [CrossRef] [PubMed]
- Boussabbeh, M.; Salem, I.B.; Belguesmi, F.; Bacha, H.; Abid-Essefi, S. Tissue oxidative stress induced by patulin and protective effect of crocin. Neurotoxicology 2016, 53, 343–349. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Jin, C.; Zhong, Y.; Li, X.; Han, J.; Xue, W.; Wu, P.; Xia, X.; Peng, X. Glutathione reduction of patulin-evoked cytotoxicity in HEK293 cells by the prevention of oxidative damage and the mitochondrial apoptotic pathway. J. Agric. Food Chem. 2018, 66, 7775–7785. [Google Scholar] [CrossRef] [PubMed]
- Kieliszek, M.; Lipinski, B. Pathophysiological significance of protein hydrophobic interactions: An emerging hypothesis. Med. Hypotheses 2018, 110, 15–22. [Google Scholar] [CrossRef]
- Kuria, A.; Tian, H.; Li, M.; Wang, Y.; Aaseth, J.O.; Zang, J.; Cao, Y. Selenium status in the body and cardiovascular disease: A systematic review and meta-analysis. Crit. Rev. Food Sci. 2021, 61, 3616–3625. [Google Scholar] [CrossRef] [PubMed]
- Ren, Z.; He, H.; Fan, Y.; Chen, C.; Zuo, Z.; Deng, J. Research progress on the toxic antagonism of selenium against mycotoxins. Biol. Trace Elem. Res. 2019, 190, 273–280. [Google Scholar] [CrossRef]
- Lu, X.; Zhang, E.; Yin, S.; Fan, L.; Hu, H. Methylseleninic acid prevents patulin-induced hepatotoxicity and nephrotoxicity via the inhibition of oxidative stress and inactivation of p53 and MAPKs. J. Agric. Food Chem. 2017, 65, 5299–5305. [Google Scholar] [CrossRef]
- Song, E.; Su, C.; Fu, J.; Xia, X.; Yang, S.; Xiao, C.; Lu, B.; Chen, H.; Sun, Z.; Wu, S. Selenium supplementation shows protective effects against patulin-induced brain damage in mice via increases in GSH-related enzyme activity and expression. Life Sci. 2014, 109, 37–43. [Google Scholar] [CrossRef]
- Kieliszek, M. Selenium–fascinating microelement, properties and sources in food. Molecules 2019, 24, 1298. [Google Scholar] [CrossRef] [Green Version]
- Thiry, C.; Ruttens, A.; De Temmerman, L.; Schneider, Y.; Pussemier, L. Current knowledge in species-related bioavailability of selenium in food. Food Chem. 2012, 130, 767–784. [Google Scholar] [CrossRef]
- Hosnedlova, B.; Kepinska, M.; Skalickova, S.; Fernandez, C.; Ruttkay-Nedecky, B.; Peng, Q.; Baron, M.; Melcova, M.; Opatrilova, R.; Zidkova, J. Nano-selenium and its nanomedicine applications: A critical review. Int. J. Nanomed. 2018, 13, 2107. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gangadoo, S.; Dinev, I.; Willson, N.; Moore, R.J.; Chapman, J.; Stanley, D. Nanoparticles of selenium as high bioavailable and non-toxic supplement alternatives for broiler chickens. Environ. Sci. Pollut. Res. 2020, 27, 16159–16166. [Google Scholar] [CrossRef] [PubMed]
- Ringuet, M.T.; Hunne, B.; Lenz, M.; Bravo, D.M.; Furness, J.B. Analysis of bioavailability and induction of glutathione peroxidase by dietary nanoelemental, organic and inorganic selenium. Nutrients 2021, 13, 1073. [Google Scholar] [CrossRef] [PubMed]
- Song, X.; Chen, Y.; Sun, H.; Liu, X.; Leng, X. Physicochemical stability and functional properties of selenium nanoparticles stabilized by chitosan, carrageenan, and gum Arabic. Carbohyd. Polym. 2021, 255, 117379. [Google Scholar] [CrossRef] [PubMed]
- García, A.V.; Álvarez-Pérez, O.B.; Rojas, R.; Aguilar, C.N.; Garrigós, M.C. Impact of olive extract addition on corn starch-based active edible films properties for food packaging applications. Foods 2020, 9, 1339. [Google Scholar] [CrossRef]
- Hu, S.; Hu, W.; Li, Y.; Li, S.; Tian, H.; Lu, A.; Wang, J. Construction and structure-activity mechanism of polysaccharide nano-selenium carrier. Carbohyd. Polym. 2020, 236, 116052. [Google Scholar] [CrossRef]
- Peng, D.; Zhang, J.; Liu, Q.; Taylor, E.W. Size effect of elemental selenium nanoparticles (Nano-Se) at supranutritional levels on selenium accumulation and glutathione S-transferase activity. J. Inorg. Biochem. 2007, 101, 1457–1463. [Google Scholar] [CrossRef]
- Bai, K.; Hong, B.; He, J.; Hong, Z.; Tan, R. Preparation and antioxidant properties of selenium nanoparticles-loaded chitosan microspheres. Int. J. Nanomed. 2017, 12, 4527. [Google Scholar] [CrossRef] [Green Version]
- Liu, J.; Liu, Q.; Han, J.; Feng, J.; Guo, T.; Li, Z.; Min, F.; Jin, R.; Peng, X. N-Acetylcysteine Inhibits Patulin-Induced Apoptosis by Affecting ROS-Mediated Oxidative Damage Pathway. Toxins 2021, 13, 595. [Google Scholar] [CrossRef]
- Chu, Q.; Wang, S.; Jiang, L.; Jiao, Y.; Sun, X.; Li, J.; Yang, L.; Hou, Y.; Wang, N.; Yao, X. Patulin induces pyroptosis through the autophagic-inflammasomal pathway in liver. Food Chem. Toxicol. 2021, 147, 111867. [Google Scholar] [CrossRef]
- Wang, Z.; Gao, Y.; Huang, X.; Huang, S.; Yang, X.; Wang, J.; Zheng, N. Metabolomics analysis underlay mechanisms in the renal impairment of mice caused by combination of aflatoxin M1 and ochratoxin A. Toxicology 2021, 458, 152835. [Google Scholar] [CrossRef] [PubMed]
- Mézes, M.; Kovács, M.; Szabó, A. Mycotoxin exposure, oxidative stress, and lipid peroxidation. In Toxicology; Elsevier: Amsterdam, The Netherlands, 2021; pp. 191–200. [Google Scholar]
- Rašić, D.; Micek, V.; Klarić, M.S.; Peraica, M. Oxidative stress as a mechanism of combined OTA and CTN toxicity in rat plasma, liver and kidney. Hum. Exp. Toxicol. 2019, 38, 434–445. [Google Scholar] [CrossRef] [PubMed]
- Miller, C.G.; Schmidt, E.E. Sulfur metabolism under stress. Antioxid. Redox Sign 2020, 33, 1158–1173. [Google Scholar] [CrossRef]
- Guillin, O.M.; Vindry, C.; Ohlmann, T.; Chavatte, L. Selenium, selenoproteins and viral infection. Nutrients 2019, 11, 2101. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Horn, T.; Bettray, W.; Slusarenko, A.J.; Gruhlke, M.C. S-allylmercaptoglutathione is a substrate for glutathione reductase (EC 1.8. 1.7) from yeast (Saccharomyces cerevisiae). Antioxidants 2018, 7, 86. [Google Scholar] [CrossRef] [Green Version]
- Liu, N.; Li, J.; Lv, J.; Yu, J.; Xie, J.; Wu, Y.; Tang, Z. Melatonin alleviates imidacloprid phytotoxicity to cucumber (Cucumis sativus L.) through modulating redox homeostasis in plants and promoting its metabolism by enhancing glutathione dependent detoxification. Ecotox Environ. Safe 2021, 217, 112248. [Google Scholar] [CrossRef]
- Bartolini, D.; Torquato, P.; Piroddi, M.; Galli, F. Targeting glutathione S-transferase P and its interactome with selenium compounds in cancer therapy. Biochim. Biophys. Acta (BBA)-Gen. Subj. 2019, 1863, 130–143. [Google Scholar] [CrossRef]
- Ommati, M.M.; Farshad, O.; Niknahad, H.; Mousavi, K.; Moein, M.; Azarpira, N.; Mohammadi, H.; Jamshidzadeh, A.; Heidari, R. Oral administration of thiol-reducing agents mitigates gut barrier disintegrity and bacterial lipopolysaccharide translocation in a rat model of biliary obstruction. Curr. Res. Pharmacol. Drug Discov. 2020, 1, 10–18. [Google Scholar] [CrossRef] [PubMed]
- Zhai, X.; Zhang, C.; Zhao, G.; Stoll, S.; Ren, F.; Leng, X. Antioxidant capacities of the selenium nanoparticles stabilized by chitosan. J. Nanobiotechnol. 2017, 15, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Moreno, J.A.S.; Mendes, A.C.; Stephansen, K.; Engwer, C.; Goycoolea, F.M.; Boisen, A.; Nielsen, L.H.; Chronakis, I.S. Development of electrosprayed mucoadhesive chitosan microparticles. Carbohyd. Polym. 2018, 190, 240–247. [Google Scholar] [CrossRef]
- Yin, L.; Ding, J.; He, C.; Cui, L.; Tang, C.; Yin, C. Drug permeability and mucoadhesion properties of thiolated trimethyl chitosan nanoparticles in oral insulin delivery. Biomaterials 2009, 30, 5691–5700. [Google Scholar] [CrossRef] [PubMed]
- Zoidis, E.; Seremelis, I.; Kontopoulos, N.; Danezis, G.P. Selenium-dependent antioxidant enzymes: Actions and properties of selenoproteins. Antioxidants 2018, 7, 66. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sobolev, O.; Gutyj, B.; Petryshak, R.; Pivtorak, J.; Kovalskyi, Y.; Naumyuk, A.; Petryshak, O.; Semchuk, I.; Mateusz, V.; Shcherbatyy, A. Biological role of selenium in the organism of animals and humans. Ukr. J. Ecol. 2018, 8, 654–665. [Google Scholar] [CrossRef]
- Dailey, R.E.; Brouwer, E.; Blaschka, A.M.; Reynaldo, E.F.; Green, S.; Monlux, W.S.; Ruggles, D.I. Intermediate-duration toxicity study of patulin in rats. J. Toxicol. Environ. Health Part A Curr. Issues 1977, 2, 713–725. [Google Scholar] [CrossRef] [PubMed]
- McKinley, E.R.; Carlton, W.W. Patulin mycotoxicosis in Swiss ICR mice. Food Cosmet. Toxicol. 1980, 18, 181–187. [Google Scholar] [CrossRef]
- Mahfoud, R.; Maresca, M.; Garmy, N.; Fantini, J. The mycotoxin patulin alters the barrier function of the intestinal epithelium: Mechanism of action of the toxin and protective effects of glutathione. Toxicol. Appl. Pharm. 2002, 181, 209–218. [Google Scholar] [CrossRef]
- Zhai, Q.; Gong, X.; Wang, C.; Zhao, J.; Zhang, H.; Tian, F.; Chen, W. Food-borne patulin toxicity is related to gut barrier disruption and can be prevented by docosahexaenoic acid and probiotic supplementation. Food Funct. 2019, 10, 1330–1339. [Google Scholar] [CrossRef]
- Singh, N.; Bansal, M.; Pal, S.; Alam, S.; Jagdale, P.; Ayanur, A.; Ansari, K.M. COX-2/EP2-EP4/β-catenin signaling regulates patulin-induced intestinal cell proliferation and inflammation. Toxicol. Appl. Pharm. 2018, 356, 224–234. [Google Scholar] [CrossRef]
- Kumar, M.R.; Muzzarelli, R.A.; Muzzarelli, C.; Sashiwa, H.; Domb, A.J. Chitosan chemistry and pharmaceutical perspectives. Chem. Rev. 2004, 104, 6017–6084. [Google Scholar] [CrossRef]
- Pan, Y.; Li, Y.; Zhao, H.; Zheng, J.; Xu, H.; Wei, G.; Hao, J. Bioadhesive polysaccharide in protein delivery system: Chitosan nanoparticles improve the intestinal absorption of insulin in vivo. Int. J. Pharmaceut. 2002, 249, 139–147. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| ALT (U/L) | AST (U/L) | BUN (mmol/L) | UA (mmol/L) | |
|---|---|---|---|---|
| Control | 26.47 ± 2.98 a | 79.07 ± 7.39 a | 10.54 ± 1.05 a | 88.33 ± 10.62 a |
| PAT | 43.70 ± 4.64 b | 98.50 ± 8.00 b | 11.75 ± 0.77 a | 88.33 ± 15.63 a |
| SeNPs | 26.33 ± 1.30 a | 80.13 ± 5.73 a | 10.75 ± 0.88 a | 95.67 ± 10.66 a |
| SeNPs + PAT | 29.33 ± 3.91 a | 87.70 ± 3.36 ab | 11.22 ± 1.25 a | 94.00 ± 3.74 a |
| SOD (U/mg Prot) | CAT (U/mg Prot) | GPx (U/mg Prot) | GR (U/mg Prot) | GST (U/mg Prot) | |
|---|---|---|---|---|---|
| Control | 310.07 ± 31.07 a | 27.76 ± 2.49 a | 2.61 ± 0.31 b | 39.82 ± 3.99 a | 6.27 ± 0.53 b |
| PAT | 220.05 ± 25.00 b | 21.50 ± 1.75 b | 0.80 ± 0.19 c | 27.79 ± 3.48 b | 3.76 ± 0.49 c |
| SeNPs | 306.69 ± 15.62 a | 26.08 ± 2.73 ab | 3.37 ± 0.29 a | 38.04 ± 3.27 a | 8.57 ± 0.67 a |
| SeNPs + PAT | 288.75 ± 23.43 a | 22.17 ± 0.74 b | 2.88 ± 0.36 ab | 27.60 ± 1.28 b | 5.37 ± 0.61 b |
| SOD (U/mg Prot) | CAT (U/mg Prot) | GPx (U/mg Prot) | GR (U/mg Prot) | GST (U/mg Prot) | |
|---|---|---|---|---|---|
| Control | 217.00 ± 32.34 a | 16.88 ± 1.31 a | 2.13 ± 0.23 b | 38.57 ± 1.36 a | 4.09 ± 0.53 a |
| PAT | 177.90 ± 17.61 a | 17.44 ± 3.01 a | 1.48 ± 0.16 c | 36.61 ± 3.57 a | 3.82 ± 0.75 a |
| SeNPs | 169.80 ± 30.54 a | 16.30 ± 0.67 a | 2.64 ± 0.12 a | 36.10 ± 6.28 a | 5.14 ± 0.76 a |
| SeNPs + PAT | 190.58 ± 19.74 a | 15.78 ± 2.57 a | 2.18 ± 0.17 b | 33.30 ± 2.71 a | 4.59 ± 0.86 a |
| MDA (U/mg Prot) | GPx (U/mg Prot) | GR (U/mg Prot) | GST (U/mg Prot) | |
|---|---|---|---|---|
| Gastric | ||||
| Control | 1.75 ± 0.39 a | 2.31 ± 0.42 a | 200.41 ± 11.10 a | 1.94 ± 0.16 a |
| PAT | 4.49 ± 0.80 b | 4.33 ± 1.09 b | 160.80 ± 12.03 b | 1.57 ± 0.04 b |
| SeNPs | 1.52 ± 0.18 a | 2.26 ± 0.23 a | 220.85 ± 21.18 a | 1.94 ± 0.14 a |
| SeNPs + PAT | 3.96 ± 0.45 b | 4.32 ± 0.65 b | 153.18 ± 6.53 b | 1.64 ± 0.12 b |
| Intestinal | ||||
| Control | 1.29 ± 0.23 a | 2.83 ± 0.75 ab | 189.88 ± 21.00 a | 1.54 ± 0.25 a |
| PAT | 4.62 ± 0.84 c | 5.00 ± 1.12 c | 139.75 ± 15.69 b | 0.82 ± 0.19 b |
| SeNPs | 1.00 ± 0.38 a | 1.99 ± 0.59 a | 192.76 ± 16.80 a | 1.97 ± 0.22 a |
| SeNPs + PAT | 3.20 ± 0.67 b | 4.03 ± 0.78 bc | 143.26 ± 13.38 b | 1.00 ± 0.20 b |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Qiu, Y.; Chen, X.; Chen, Z.; Zeng, X.; Yue, T.; Yuan, Y. Effects of Selenium Nanoparticles on Preventing Patulin-Induced Liver, Kidney and Gastrointestinal Damage. Foods 2022, 11, 749. https://doi.org/10.3390/foods11050749
Qiu Y, Chen X, Chen Z, Zeng X, Yue T, Yuan Y. Effects of Selenium Nanoparticles on Preventing Patulin-Induced Liver, Kidney and Gastrointestinal Damage. Foods. 2022; 11(5):749. https://doi.org/10.3390/foods11050749
Chicago/Turabian StyleQiu, Yue, Xinlu Chen, Zhangxi Chen, Xuejun Zeng, Tianli Yue, and Yahong Yuan. 2022. "Effects of Selenium Nanoparticles on Preventing Patulin-Induced Liver, Kidney and Gastrointestinal Damage" Foods 11, no. 5: 749. https://doi.org/10.3390/foods11050749
APA StyleQiu, Y., Chen, X., Chen, Z., Zeng, X., Yue, T., & Yuan, Y. (2022). Effects of Selenium Nanoparticles on Preventing Patulin-Induced Liver, Kidney and Gastrointestinal Damage. Foods, 11(5), 749. https://doi.org/10.3390/foods11050749