
Colostrum-Induced Temporary Changes in the Expression of Proteins Regulating the Epithelial Barrier Function in the Intestine

 ,
,  , , ,
, , ,  , and
, and
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals and Procedures
2.2. Tissues
2.3. Quantitative Real-Time Reverse Transcription PCR (qRT-PCR) Analysis
2.4. Histological and Immunohistological Methods
2.5. Statistic Analysis
3. Results
3.1. Small Intestine
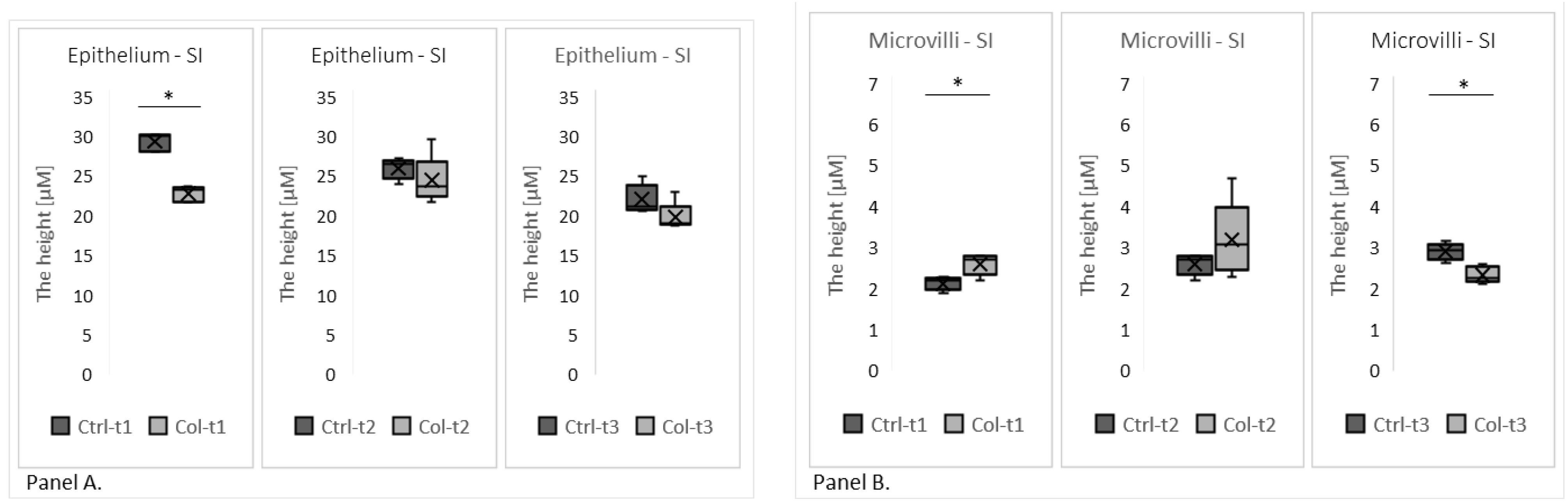
3.1.1. Effect of Col Supplementation on the Morphology of the Small Intestine
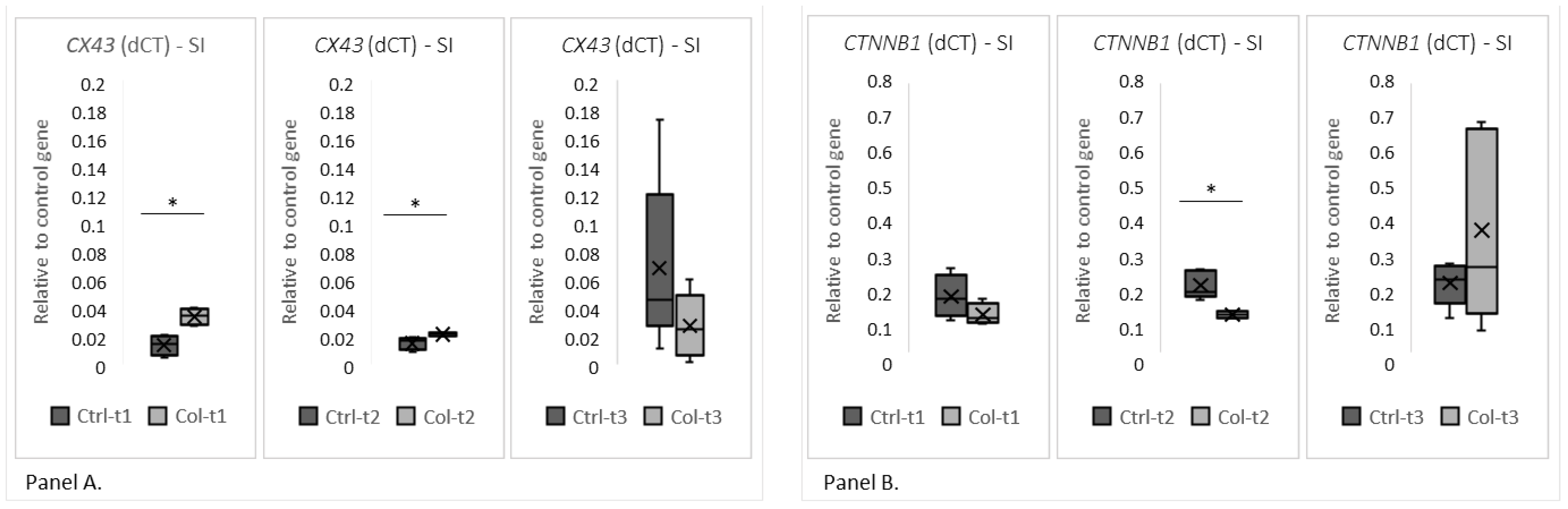
3.1.2. Effects of Col Supplementation on the Expression of Genes Forming the Epithelial Barrier of the Small Intestine
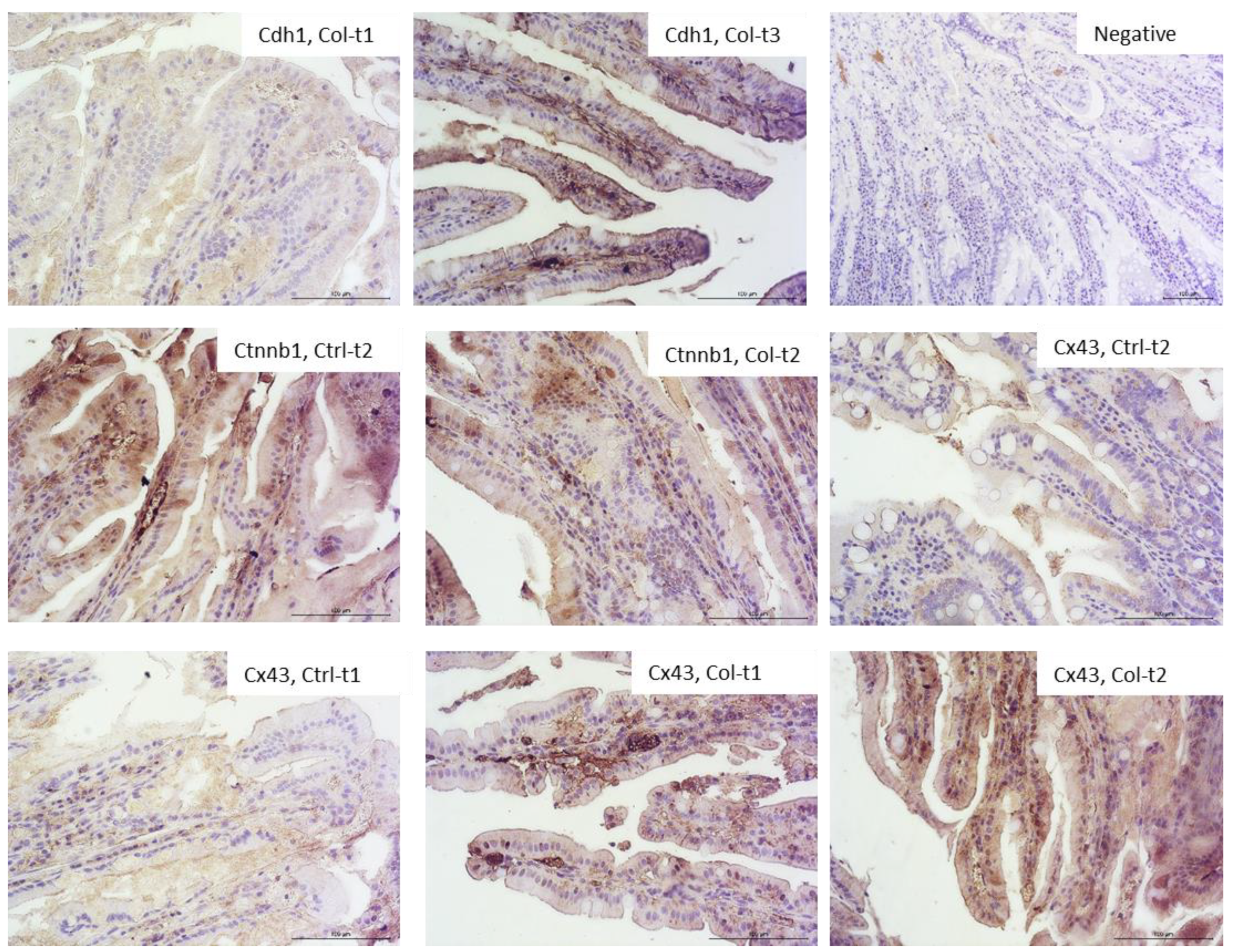
3.1.3. Effects of Col Supplementation on the Expression of Proteins Forming the Epithelial Barrier of the Small Intestine
3.1.4. Correlations between Gene Expression and Small Intestine Epithelium and Microvilli Morphology
3.2. Large Intestine
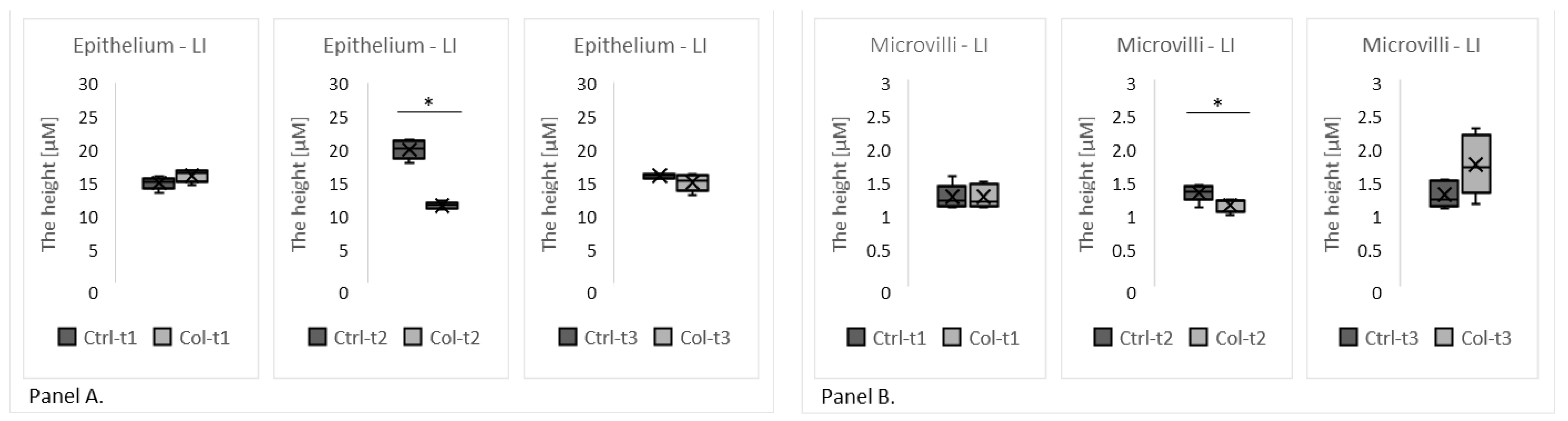
3.2.1. Effect of Col Supplementation on the Morphology of the Large Intestine
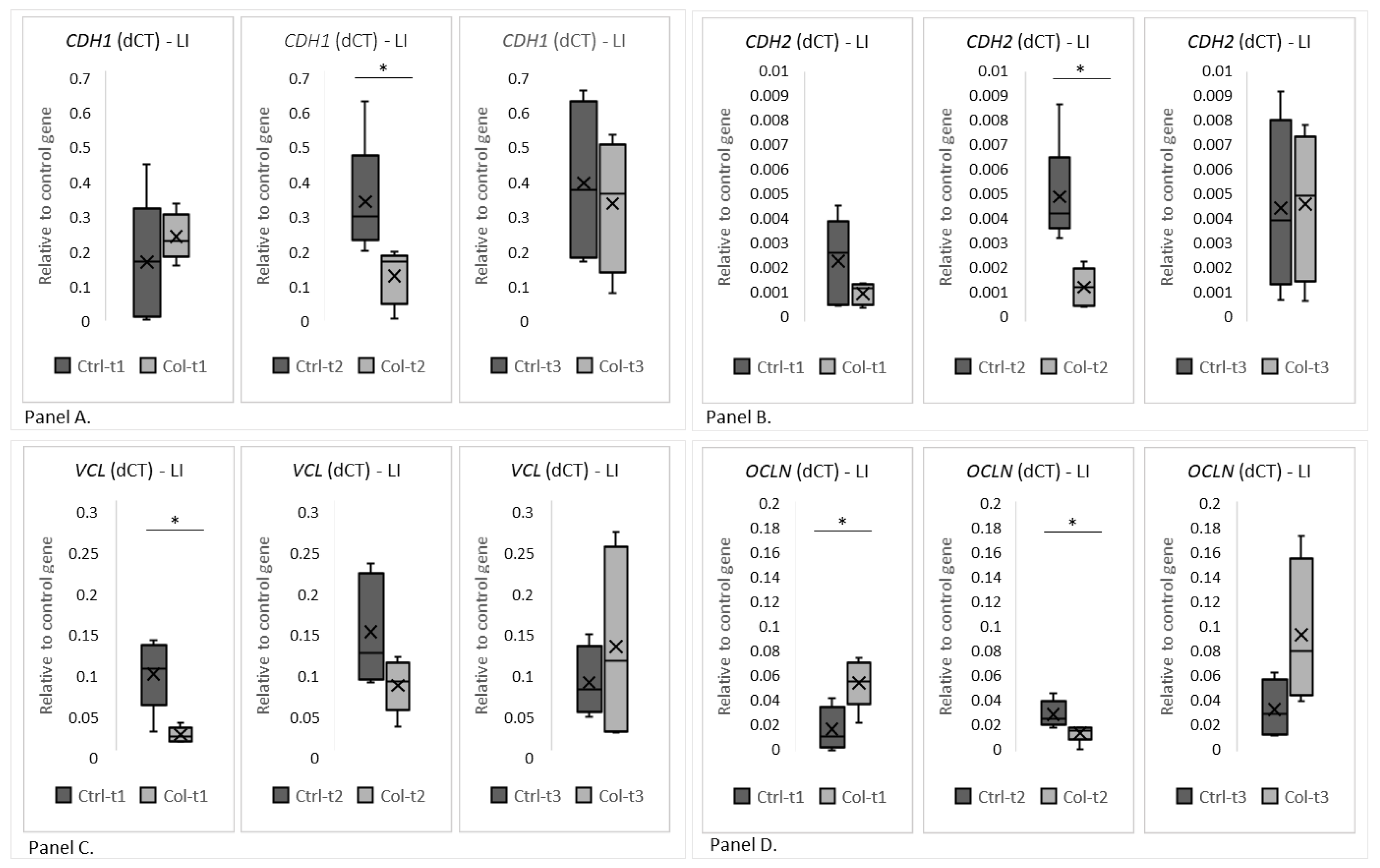
3.2.2. Effects of Col Supplementation on the Expression of Genes Forming the Epithelial Barrier of the Large Intestine
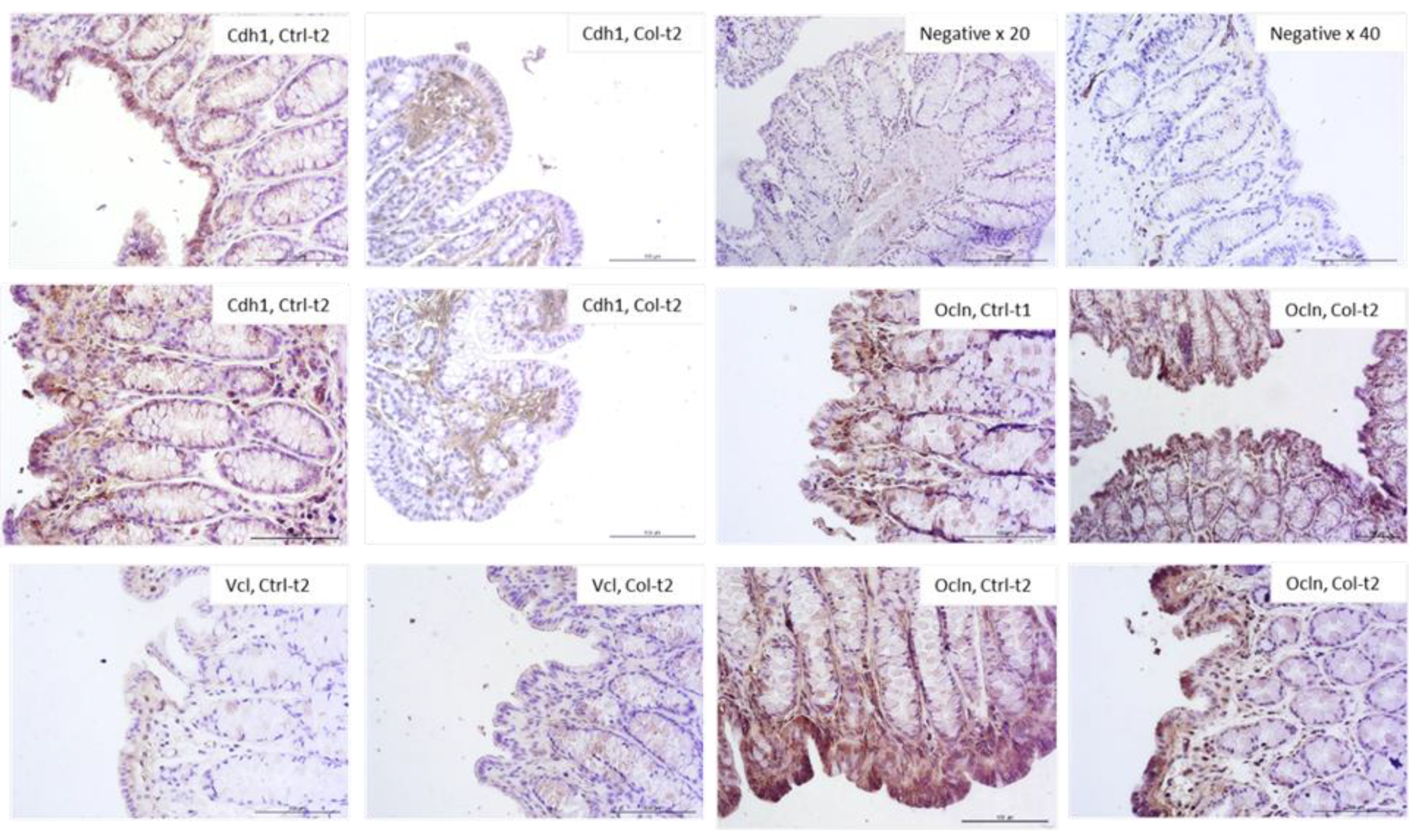
3.2.3. Effects of Col Supplementation on the Expression of Proteins Forming the Epithelial Barrier of the Large Intestine
3.2.4. Correlations between Gene Expression and Epithelium or Microvilli in the Large Intestine
4. Discussion
4.1. Effects of Col Supplementation on Cellular Junction Genes and Protein Expression
4.1.1. Cdh1 and Cdh2
4.1.2. Ctnnb1
4.1.3. Cx43
4.1.4. Vcl
4.1.5. Ocln
4.1.6. Cytoskeleton Components
4.2. Relationships in the Expression of Cell Adhesion Molecules
4.3. The Limitation of the Present Study
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Lee, B.; Moon, K.M.; Kim, C.Y. Tight junction in the intestinal epithelium: Its association with diseases and regulation by phytochemicals. J. Immunol. Res. 2018, 2018, 2645465. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, T. Regulation of the intestinal barrier by nutrients: The role of tight junctions. Anim. Sci. J. 2020, 91, e13357. [Google Scholar] [CrossRef]
- Lepanto, M.S.; Rosa, L.; Paesano, R.; Valenti, P.; Cutone, A. Lactoferrin in aseptic and septic inflammation. Molecules 2019, 24, 1323. [Google Scholar] [CrossRef] [PubMed]
- Santos-Pereira, C.; Rocha, J.F.; Fernandes, H.S.; Rodrigues, L.R.; Côrte-Real, M.; Sousa, S.F. The milk-derived lactoferrin inhibits V-ATPase activity by targeting its V1 domain. Int. J. Biol. Macromol. 2021, 186, 54–70. [Google Scholar] [CrossRef] [PubMed]
- Alexander, M.R.; Norlander, A.; Elijovich, F.; Atreya, R.V.; Gaye, A.; Gnecco, J.S.; Laffer, C.L.; Galindo, C.L.; Madhur, M.S. Human monocyte transcriptional profiling identifies IL-18 receptor accessory protein and lactoferrin as novel immune targets in hypertension. Br. J. Pharmacol. 2019, 176, 2015–2027. [Google Scholar] [CrossRef]
- Bagwe-Parab, S.; Yadav, P.; Kaur, G.; Tuli, H.S.; Buttar, H.S. Therapeutic applications of human and bovine colostrum in the treatment of gastrointestinal diseases and distinctive cancer types: The current evidence. Front. Pharmacol. 2020, 11, 01100. [Google Scholar] [CrossRef]
- Huang, L.; Chen, R.; Liu, L.; Zhou, Y.; Chen, Z. Lactoferrin ameliorates pathological cardiac hypertrophy related to mitochondrial quality control in aged mice. Food Funct. 2021, 12, 7514–7526. [Google Scholar] [CrossRef]
- Trybek, G.; Metlerski, M.; Szumilas, K.; Aniko-Włodarczyk, M.; Preuss, O.; Grocholewicz, K.; Wiszniewska, B. The biological properties of lactoferrin. Cent. Eur. J. Sport Sci. Med. 2016, 15, 15–25. [Google Scholar] [CrossRef][Green Version]
- Trybek, G.; Jedliński, M.; Jaroń, A.; Preuss, O.; Mazur, M.; Grzywacz, A. Impact of lactoferrin on bone regenerative processes and its possible implementation in oral surgery—A systematic review of novel studies with metanalysis and metaregression. BMC Oral Health 2020, 20, 232. [Google Scholar] [CrossRef]
- Cutone, A.; Rosa, L.; Ianiro, G.; Lepanto, M.S.; Di Patti, M.C.B.; Valenti, P.; Musci, G. Lactoferrin’s anti-cancer properties: Safety, selectivity, and wide range of action. Biomolecules 2020, 10, 456. [Google Scholar] [CrossRef]
- Zhao, X.; Xu, X.-X.; Liu, Y.; Xi, E.-Z.; An, J.-J.; Tabys, D.; Liu, N. The in vitro protective role of bovine lactoferrin on intestinal epithelial barrier. Molecules 2019, 24, 148. [Google Scholar] [CrossRef] [PubMed]
- Kur, P.; Kolasa-Wołosiuk, A.; Grabowska, M.; Kram, A.; Tarnowski, M.; Baranowska-Bosiacka, I.; Rzeszotek, S.; Piasecka, M.; Wiszniewska, B. The postnatal offspring of finasteride-treated male rats shows hyperglycaemia, elevated hepatic glycogen storage and altered GLUT2, IR, and AR expression in the liver. Int. J. Mol. Sci. 2021, 22, 1242. [Google Scholar] [CrossRef]
- Liu, H.-M.; Yang, D.; Liu, Z.-F.; Hu, S.-Z.; Yan, S.-H.; He, X.-W. Density distribution of gene expression profiles and evaluation of using maximal information coefficient to identify differentially expressed genes. PLoS ONE 2019, 14, e0219551. [Google Scholar] [CrossRef]
- Schneider, M.R.; Dahlhoff, M.; Horst, D.; Hirschi, B.; Trülzsch, K.; Müller-Höcker, J.; Vogelmann, R.; Allgäuer, M.; Gerhard, M.; Steininger, S.; et al. A key role for e-cadherin in intestinal homeostasis and paneth cell maturation. PLoS ONE 2010, 5, e14325. [Google Scholar] [CrossRef] [PubMed]
- Nelson, W.J.; Nusse, R. Convergence of Wnt, beta-catenin, and cadherin pathways. Science 2004, 303, 1483–1487. [Google Scholar] [CrossRef]
- Zadvornyi, T.V.; Lukianova, N.Y.; Borikun, T.V.; Chekhun, V.F. Effects of exogenous lactoferrin on phenotypic profile and inva-siveness of human prostate cancer cells (DU145 and LNCaP) in vitro. Exp. Oncol. 2018, 40, 184–189. [Google Scholar] [CrossRef]
- Kline, K.T.; Lian, H.; Zhong, X.S.; Luo, X.; Winston, J.H.; Cong, Y.; Savidge, T.C.; Dashwood, R.H.; Powell, D.W.; Li, Q. Neonatal injury increases gut permeability by epigenetically suppressing E-cadherin in adulthood. J. Immunol. 2020, 204, 980–989. [Google Scholar] [CrossRef] [PubMed]
- Cepek, K.L.; Shaw, S.K.; Parker, C.M.; Russell, G.J.; Morrow, J.S.; Rimm, D.L.; Brenner, M.B. Adhesion between epithelial cells and T lymphocytes mediated by E-cadherin and the αEβ7 integrin. Nature 1994, 372, 190–193. [Google Scholar] [CrossRef]
- Grabowska, M.M.; Day, M.L. Soluble E-cadherin: More than a symptom of disease. Front. Biosci. 2012, 17, 1948–1964. [Google Scholar] [CrossRef]
- Trentini, A.; Maritati, M.; Cervellati, C.; Manfrinato, M.C.; Gonelli, A.; Volta, C.A.; Vesce, F.; Greco, P.; Dallocchio, F.; Bellini, T.; et al. Vaginal lactoferrin modulates PGE2, MMP-9, MMP-2, and TIMP-1 amniotic fluid concentrations. Mediat. Inflamm. 2016, 2016, 3648719. [Google Scholar] [CrossRef]
- Libusova, L.; Stemmler, M.P.; Hierholzer, A.; Schwarz, H.; Kemler, R. N-cadherin can structurally substitute for E-cadherin during intestinal development but leads to polyp formation. Development 2010, 137, 2297–2305. [Google Scholar] [CrossRef]
- Loh, C.-Y.; Chai, J.Y.; Tang, T.F.; Wong, W.F.; Sethi, G.; Shanmugam, M.K.; Chong, P.P.; Looi, C.Y. The E-cadherin and N-cadherin switch in epithelial-to-mesenchymal transition: Signaling, therapeutic implications, and challenges. Cells 2019, 8, 1118. [Google Scholar] [CrossRef]
- Takayama, Y.; Mizumachi, K. Inhibitory effect of lactoferrin on hypertrophic differentiation of ATDC5 mouse chondroprogenitor cells. BioMetals 2010, 23, 477–484. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Li, B.; Lee, C.; Zhu, H.; Zheng, S.; Pierro, A. Protective effects of lactoferrin on injured intestinal epithelial cells. J. Pediatr. Surg. 2019, 54, 2509–2513. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.; Chen, C.Y.; Hu, Y.; Lyu, B. The relationship between adherens junction and tight junction and clinical symptoms in patients with diarrhea predominant irritable bowel syndrome. Zhonghua Nei Ke Za Zhi 2020, 59, 40–46. [Google Scholar]
- Maes, M.; Yanguas, S.C.; Willebrords, J.; Cogliati, B.; Vinken, M. Connexin and pannexin signaling in gastrointestinal and liver disease. Transl. Res. 2015, 166, 332–343. [Google Scholar] [CrossRef] [PubMed]
- Al-Ghadban, S.; Kaissi, S.; Homaidan, F.R.; Naim, H.Y.; El-Sabban, M.E. Cross-talk between intestinal epithelial cells and immune cells in inflammatory bowel disease. Sci. Rep. 2016, 6, 29783. [Google Scholar] [CrossRef]
- Kotini, M.; Barriga, E.H.; Leslie, J.; Gentzel, M.; Rauschenberger, V.; Schambony, A.; Mayor, R. Gap junction protein connexin-43 is a direct transcriptional regulator of N-cadherin in vivo. Nat. Commun. 2018, 9, 3846. [Google Scholar] [CrossRef]
- Yang, S.-C.; Hung, C.-F.; Aljuffali, I.A.; Fang, J.-Y. The roles of the virulence factor IpaB in Shigella spp. in the escape from immune cells and invasion of epithelial cells. Microbiol. Res. 2015, 181, 43–51. [Google Scholar] [CrossRef]
- Vandrovcova, M.; Douglas, T.E.L.; Heinemann, S.; Scharnweber, D.; Dubruel, P.; Bacakova, L. Collagen-lactoferrin fibrillar coatings enhance osteoblast proliferation and differentiation. J. Biomed. Mater. Res. Part A 2015, 103, 525–533. [Google Scholar] [CrossRef]
- Adler, B.L.; McMahan, Z. Anti-vinculin autoantibodies in systemic sclerosis: A step toward a novel biomarker? Clin. Rheumatol. 2021, 40, 809–811. [Google Scholar] [CrossRef] [PubMed]
- Pimentel, M.; Morales, W.; Pokkunuri, V.; Brikos, C.; Kim, S.M.; Kim, S.E.; Triantafyllou, K.; Weitsman, S.; Marsh, Z.; Marsh, E.; et al. Autoimmunity links vinculin to the pathophysiology of chronic functional bowel changes following campylobacter jejuni infection in a rat model. Am. J. Dig. Dis. 2014, 60, 1195–1205. [Google Scholar] [CrossRef]
- Mierke, C.T.; Kollmannsberger, P.; Zitterbart, D.P.; Diez, G.; Koch, T.M.; Marg, S.; Ziegler, W.H.; Goldmann, W.H.; Fabry, B. Vinculin Facilitates Cell Invasion into Three-dimensional Collagen Matrices. J. Biol. Chem. 2010, 285, 13121–13130. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.H.; Geng, J.S. Expression of paxillin and vinculin in gastric carcinoma and precancerous lesion and their effects on prognosis of gastric carcinoma. Chin. J. Diagn. Pathol. 2007, 14, 377–380. [Google Scholar]
- Oh-Oka, K.; Kono, H.; Ishimaru, K.; Miyake, K.; Kubota, T.; Ogawa, H.; Okumura, K.; Shibata, S.; Nakao, A. Expressions of tight junction proteins occludin and claudin-1 are under the circadian control in the mouse large intestine: Implications in intestinal permeability and susceptibility to colitis. PLoS ONE 2014, 9, e98016. [Google Scholar] [CrossRef]
- Hu, P.; Zhao, F.; Wang, J.; Zhu, W. Early-life lactoferrin intervention modulates the colonic microbiota, colonic microbial metabolites and intestinal function in suckling piglets. Appl. Microbiol. Biotechnol. 2020, 104, 6185–6197. [Google Scholar] [CrossRef] [PubMed]
- Olivier, S.; Leclerc, J.; Grenier, A.; Foretz, M.; Tamburini, J.; Viollet, B. AMPK activation promotes tight junction assembly in intestinal epithelial caco-2 cells. Int. J. Mol. Sci. 2019, 20, 5171. [Google Scholar] [CrossRef] [PubMed]
- Kitakaze, T.; Oshimo, M.; Kobayashi, Y.; Ryu, M.; Suzuki, Y.A.; Inui, H.; Harada, N.; Yamaji, R. Lactoferrin promotes murine C2C12 myoblast proliferation and differentiation and myotube hypertrophy. Mol. Med. Rep. 2018, 17, 5912–5920. [Google Scholar] [CrossRef] [PubMed]
- Vanuytsel, T.; Vermeire, S.; Cleynen, I. The role of haptoglobin and its related protein, zonulin, in inflammatory bowel disease. Tissue Barriers 2013, 1, e27321. [Google Scholar] [CrossRef]
- Wall, M.E.; Otey, C.; Qi, J.; Banes, A.J. Connexin 43 is localized with actin in tenocytes. Cell Motil. Cytoskelet. 2007, 64, 121–130. [Google Scholar] [CrossRef]
- Ionta, M.; Ferreira, R.A.S.; Pfister, S.C.; Machado-Santelli, G.M. Exogenous Cx43 expression decrease cell proliferation rate in rat hepatocarcinoma cells independently of functional gap junction. Cancer Cell Int. 2009, 9, 22. [Google Scholar] [CrossRef] [PubMed]
- Batra, N.; Burra, S.; Siller-Jackson, A.J.; Gu, S.; Xia, X.; Weber, G.F.; DeSimone, D.; Bonewald, L.F.; Lafer, E.M.; Sprague, E.; et al. Mechanical stress-activated integrin 5 1 induces opening of connexin 43 hemichannels. Proc. Natl. Acad. Sci. USA 2012, 109, 3359–3364. [Google Scholar] [CrossRef] [PubMed]
- Waxse, B.J.; Sengupta, P.; Hesketh, G.G.; Lippincott-Schwartz, J.; Buss, F. Myosin VI facilitates connexin 43 gap junction accretion. J. Cell Sci. 2017, 130, 827–840. [Google Scholar] [CrossRef] [PubMed]
- Montecino-Rodriguez, E.; Leathers, H.; Dorshkind, K. Expression of connexin 43 (Cx43) is critical for normal hematopoiesis. Blood 2000, 96, 917–924. [Google Scholar] [CrossRef] [PubMed]
- Garibay-Cerdenares, O.L.; I Hernández-Ramírez, V.; Osorio-Trujillo, J.C.; Gallardo-Rincón, D.; Talamás-Rohana, P. Haptoglobin and CCR2 receptor expression in ovarian cancer cells that were exposed to ascitic fluid: Exploring a new role of haptoglobin in the tumoral microenvironment. Cell Adhes. Migr. 2015, 9, 394–405. [Google Scholar] [CrossRef] [PubMed]
- Nichols, L.A.; Grunz-Borgmann, E.A.; Wang, X.; Parrish, A.R. A role for the age-dependent loss of α(E)-catenin in regulation of N-cadherin expression and cell migration. Physiol. Rep. 2014, 2, e12039. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| CDH1 | CDH2 | CTNNB1 | ACTG2 | VCL | CX43 | HP | OCLN | MYH9 | Epithelium | Microvilli | |
|---|---|---|---|---|---|---|---|---|---|---|---|
| CDH1 | r = 0.07 p = 0.71 | r = 0.61 p < 0.01 * | r = 0.04 p = 0.85 | r = 0.15 p = 0.48 | r = 0.03 p = 0.87 | r = −0.08 p = 0.70 | r = 0.36 p = 0.08 | r = 0.48 p < 0.05 * | r = −0.11 p = 0.57 | r = −0.12 p = 0.57 | |
| CDH2 | r = 0.07 p = 0.71 | r = 0.28 p = 0.18 | r = 0.74 p < 0.01 * | r = 0.62 p < 0.01* | r = 0.76 p < 0.01 * | r = 0.65 p < 0.01 * | r = 0.11 p = 0.64 | r = 0.61 p < 0.01 * | r = −0.25 p = 0.22 | r = 0.13 p = 0.52 | |
| CTNNB1 | r = 0.61 p < 0.01 * | r = 0.28 p = 0.18 | r = 0.52 p < 0.01 * | r = 0.56 p < 0.01 * | r = 0.42 p < 0.05 * | r = 0.46 p < 0.05 * | r = 0.44 p < 0.05 * | r = 0.72 p < 0.01 * | r = −0.10 p = 0.63 | r = −0.25 p = 0.22 | |
| ACTG2 | r = 0.04 p = 0.85 | r = 0.74 p < 0.01 * | r = 0.52 p < 0.01 * | r = 0.74 p < 0.01 * | r = 0.91 p < 0.01 * | r = 0.81 p < 0.01 * | r = 0.35 p = 0.09 | r = 0.76 p < 0.01 * | r = −0.35 P = 0.09 | r = 0.12 p = 0.56 | |
| VCL | r = 0.15 p = 0.48 | r = 0.62 p < 0.01 * | r = 0.56 p < 0.01 * | r = 0.74 p < 0.01 * | r = 0.61 p < 0.01 * | r = 0.61 p < 0.01 * | r = 0.18 p = 0.38 | r = 0.46 p < 0.05 * | r = 0.03 p = 0.89 | r = −0.33 p = 0.10 | |
| CX43 | r = 0.03 p = 0.87 | r = 0.76 p < 0.01 * | r = 0.42 p < 0.05 * | r = 0.91 p < 0.01 * | r = 0.61 p < 0.01 * | r = 0.83 p < 0.01 | r = 0.34 p = 0.10 | r = 0.77 p < 0.01 * | r = −0.40 p < 0.05 * | r = 0.27 p = 0.20 | |
| HP | r = −0.08 p = 0.70 | r = 0.65 p < 0.01 * | r = 0.46 p < 0.05 * | r = 0.81 p < 0.01 * | r = 0.61 p < 0.01 * | r = 0.83 p < 0.01 | r = 0.23 p = 0.28 | r = 0.66 p < 0.01 | r = −0.19 p = 0.38 | r = 0.14 p = 0.51 | |
| OCLN | r = 0.36 p = 0.08 | r = 0.11 p = 0.64 | r = 0.44 p < 0.05 * | r = 0.35 p = 0.09 | r = 0.18 p = 0.38 | r = 0.34 p = 0.10 | r = 0.23 p = 0.28 | r = 0.53 p < 0.01 * | r = −0.31 p = 0.12 | r = 0.21 p = 0.33 | |
| MYH9 | r = 0.48 p < 0.05 * | r = 0.61 p < 0.01 * | r = 0.72 p < 0.01 * | r = 0.76 p < 0.01 * | r = 0.46 p < 0.05 * | r = 0.77 p < 0.01 * | r = 0.66 p < 0.01 * | r = 0.53 p < 0.01 * | r = −0.39 p < 0.05 * | r = 0.20 p = 0.33 | |
| Epithelium | r = −0.11 p = 0.57 | r = −0.25 p = 0.22 | r = −0.10 p = 0.63 | r = −0.35 P = 0.09 | r = 0.03 p = 0.89 | r = −0.40 p < 0.05 * | r = −0.19 p = 0.38 | r = −0.31 p = 0.12 | r = −0.39 p < 0.05 * | r = −0.32 p = 0.11 | |
| Microvilli | r = −0.12 p = 0.57 | r = 0.13 p = 0.52 | r = −0.25 p = 0.22 | r = 0.12 p = 0.56 | r = −0.33 p = 0.10 | r = 0.27 p = 0.20 | r = 0.14 p = 0.51 | r = 0.21 p = 0.33 | r = 0.20 p = 0.33 | r = −0.32 p = 0.11 |
| CDH1 | CDH2 | CTNNB1 | ACTG2 | VCL | CX43 | HP | OCLN | Epithelium | Microvilli | |
|---|---|---|---|---|---|---|---|---|---|---|
| CDH1 | r = 0.55 p < 0.01* | r = 0.49 p = 0.01* | r = 0.24 p = 0.22 | r = 0.26 p =0.18 | r = 0.25 p = 0.20 | r = −0.13 p = 0.51 | r = 0.67 p < 0.01* | r = 0.42 p = 0.02* | r = 0.28 p = 0.15 | |
| CDH2 | r = 0.55 p < 0.01* | r = 0.38 p = 0.06 | r = 0.56 p < 0.01* | r = 0.75 p < 0.01* | r = 0.49 p < 0.01* | r = 0.19 p = 0.33 | r = 0 p > 0.99 | r = 0.47 p = 0.01* | r = 0.13 p = 0.56 | |
| CTNNB1 | r = 0.49 p = 0.01* | r = 0.38 p = 0.06 | r = 0.28 p = 0.15 | r = 0.45 p = 0.02 | r = 0.36 p = 0.06 | r = 0.22 p = 0.27 | r = 0.08 p = 0.68 | r = 0.03 p = 0.85 | r = 0.03 p = 0.86 | |
| ACTG2 | r = 0.24 p = 0.22 | r = 0.56 p < 0.01* | r = 0.28 p = 0.15 | r = 0.62 p < 0.01* | r = 0.21 p = 0.31 | r = 0.04 p = 0.81 | r = −0.27 p = 0.16 | r = 0.06 p = 0.74 | r = 0.03 p = 0.86 | |
| VCL | r = 0.26 p =0.18 | r = 0.75 p < 0.01* | r = 0.45 p = 0.02* | r = 0.62 p < 0.01* | r = 0.30 p =0.12 | r = 0.15 p = 0.42 | r = −0.20 p = 0.31 | r = 0.27 p = 0.16 | r = 0.13 p = 0.52 | |
| CX43 | r = 0.25 p = 0.20 | r = 0.49 p < 0.01* | r = 0.36 p = 0.06 | r = 0.21 p = 0.31 | r = 0.30 p =0.12 | r = 0.71 p < 0.01* | r = −0.22 p = 0.27 | r = 0.15 p = 0.42 | r = 0.19 p = 0.33 | |
| HP | r = −0.13 p = 0.51 | r = 0.19 p = 0.33 | r = 0.22 p = 0.27 | r = 0.04 p = 0.81 | r = 0.15 p = 0.42 | r = 0.71 p < 0.01* | r = −0.35 p = 0.06 | r = −0.29 p = 0.13 | r < 0.01 p = 0.99 | |
| OCLN | r = 0.67 p < 0.01* | r = 0 p > 0.99 | r = 0.08 p = 0.68 | r = −0.27 p = 0.16 | r = −0.20 p = 0.31 | r = −0.22 p = 0.27 | r =−0.35 p = 0.06 | r = 0.41 p = 0.03* | r = 0.44 p = 0.02* | |
| Epithelium | r = 0.42 p = 0.02* | r = 0.47 p = 0.01* | r = 0.03 p = 0.85 | r = 0.06 p = 0.74 | r = 0.27 p = 0.16 | r = 0.15 p = 0.42 | r = −0.29 p = 0.13 | r = 0.41 p = 0.03* | r = 0.33 p = 0.08 | |
| Microvilli | r = 0.28 p = 0.15 | r = 0.13 p = 0.56 | r = 0.03 p = 0.86 | r < 0.01 p = 0.99 | r = 0.13 p = 0.52 | r = 0.19 p = 0.33 | r < 0.01 p = 0.99 | r = 0.44 p = 0.02* | r = 0.33 p = 0.08 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rzeszotek, S.; Trybek, G.; Tarnowski, M.; Serwin, K.; Jaroń, A.; Schneider, G.; Kolasa, A.; Wiszniewska, B. Colostrum-Induced Temporary Changes in the Expression of Proteins Regulating the Epithelial Barrier Function in the Intestine. Foods 2022, 11, 685. https://doi.org/10.3390/foods11050685
Rzeszotek S, Trybek G, Tarnowski M, Serwin K, Jaroń A, Schneider G, Kolasa A, Wiszniewska B. Colostrum-Induced Temporary Changes in the Expression of Proteins Regulating the Epithelial Barrier Function in the Intestine. Foods. 2022; 11(5):685. https://doi.org/10.3390/foods11050685
Chicago/Turabian StyleRzeszotek, Sylwia, Grzegorz Trybek, Maciej Tarnowski, Karol Serwin, Aleksandra Jaroń, Gabriela Schneider, Agnieszka Kolasa, and Barbara Wiszniewska. 2022. "Colostrum-Induced Temporary Changes in the Expression of Proteins Regulating the Epithelial Barrier Function in the Intestine" Foods 11, no. 5: 685. https://doi.org/10.3390/foods11050685
APA StyleRzeszotek, S., Trybek, G., Tarnowski, M., Serwin, K., Jaroń, A., Schneider, G., Kolasa, A., & Wiszniewska, B. (2022). Colostrum-Induced Temporary Changes in the Expression of Proteins Regulating the Epithelial Barrier Function in the Intestine. Foods, 11(5), 685. https://doi.org/10.3390/foods11050685