
Effect of Pulsed Electric Fields on the Lipidomic Profile of Lipid Extracted from Hoki Fish Male Gonad


 ,
,  ,
, 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals and Sample Preparation
2.2. Measurements
2.2.1. Electrical Input
2.2.2. Temperature and Electrical Conductivity
2.3. Proximate Analysis
2.4. Total Lipid Extraction
2.5. Fatty Acid Methyl Ester (FAME) Analysis by GC-FID
2.5.1. Sample Preparation
2.5.2. GC-FID Operating Conditions
2.6. Phospholipid Analysis Using 31P NMR
2.7. Statistical Analysis
3. Results and Discussion
3.1. Proximate Analysis of HMG
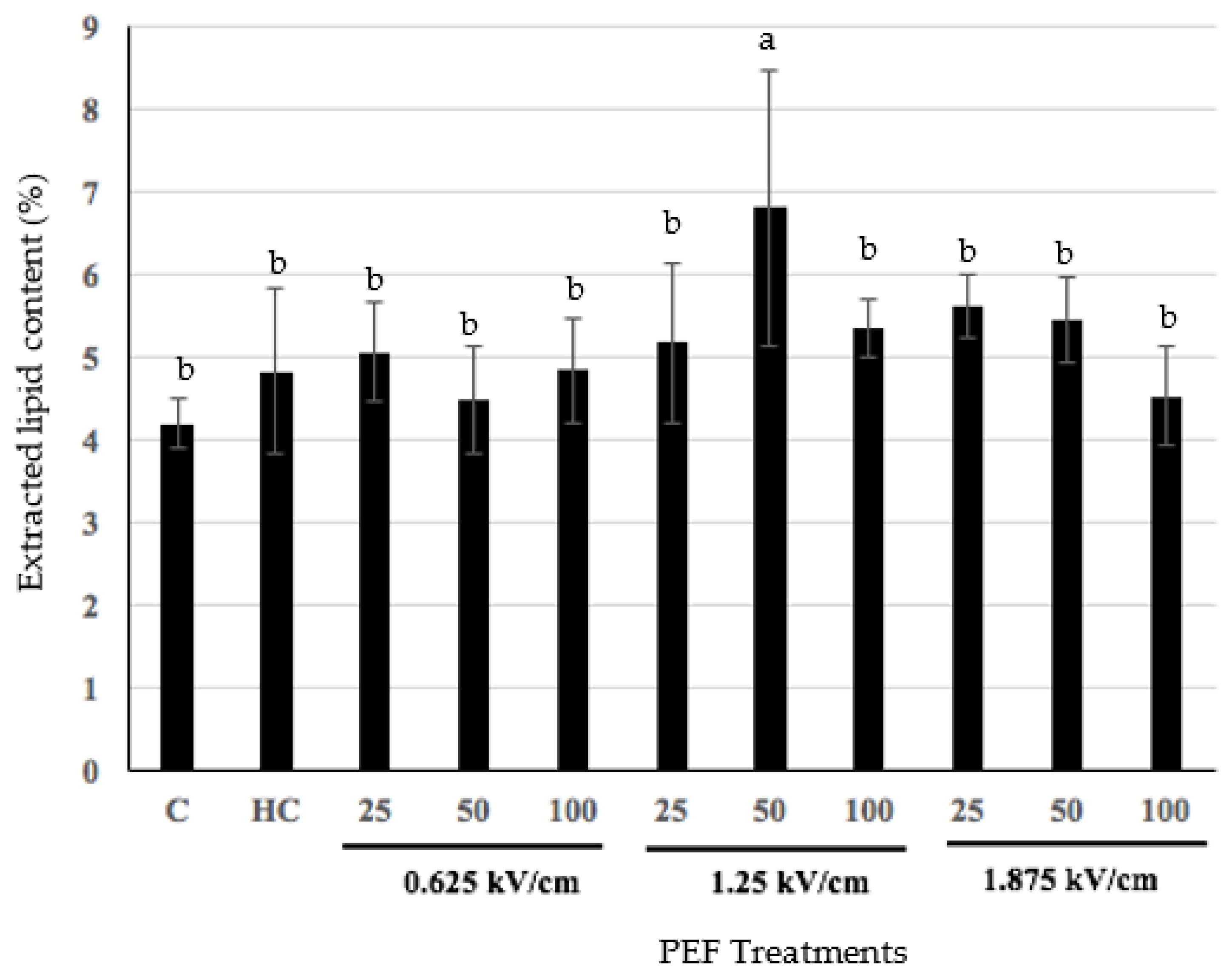
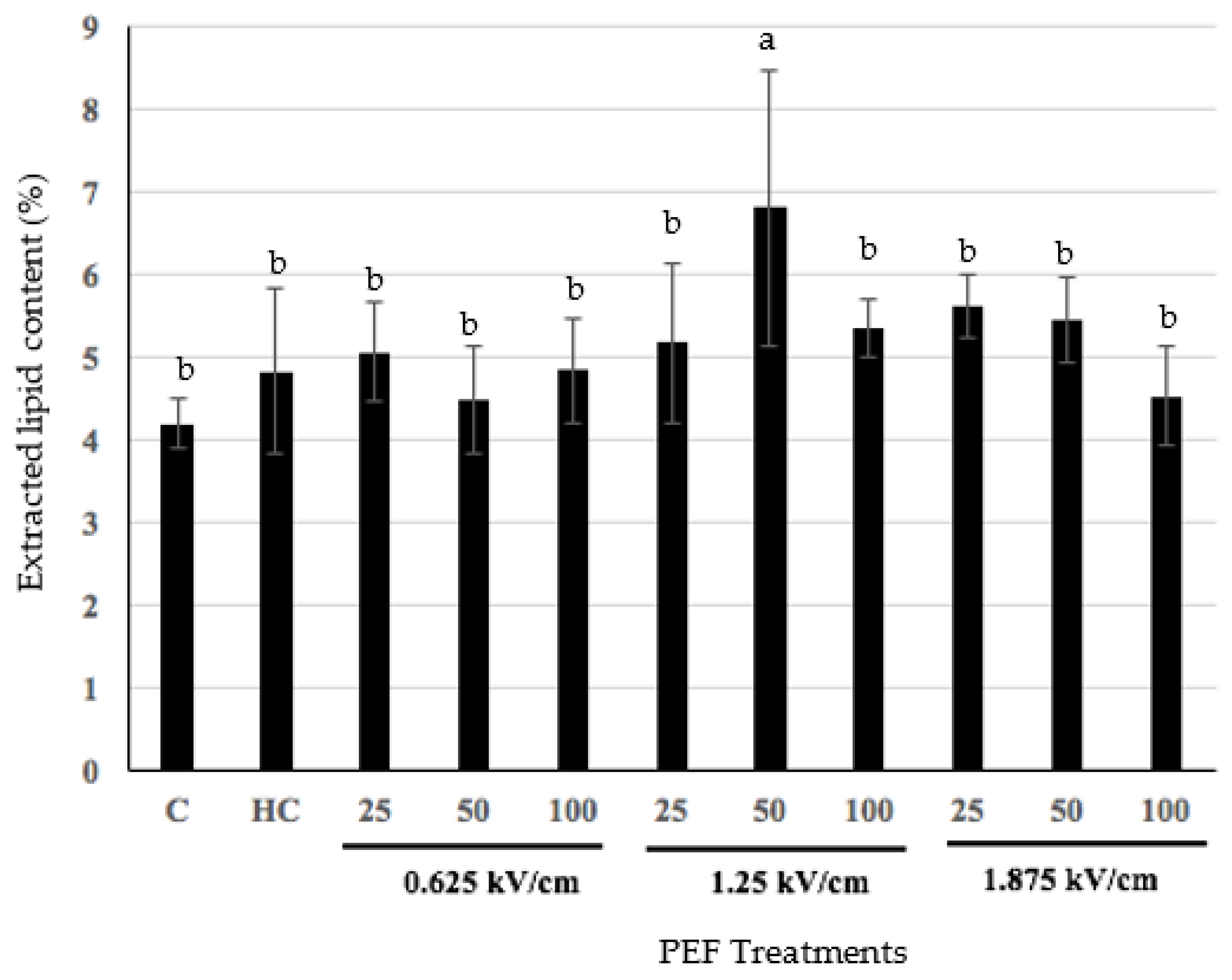
3.2. Effect of Pulsed Electric Fields on Hoki Roe
3.3. Effect of Heat Generated by PEF Treatment on the Composition of Lipid Extracted from HMG
3.3.1. Effect of Heat on Fatty Acid Composition
3.3.2. Effect of Heat on the Phospholipid Composition
3.4. Effect of PEF Treatment on the Lipid Composition of HMG
3.4.1. Effect on Fatty Acids
Effect of Frequency on Fatty Acid Composition
Effect of PEF Intensity on Fatty Acid Composition
Effect of the Interaction between Frequency and PEF Field Strength on Fatty Acid Composition of Extracted Lipid
3.4.2. Effect on Phospholipid Composition of Lipid Extracted from PEF Pre-Treated HMG
Effect of PEF Frequency on Phospholipid Composition
Effect of PEF Field Strength on Phospholipid Composition
Effect of the Interaction between PEF Frequency and PEF Field Strength on Phospholipid Composition
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Gil, A.; Gil, F. Fish, a Mediterranean source of n-3 PUFA: Benefits do not justify limiting consumption. Br. J. Nutr. 2015, 113, S58–S67. [Google Scholar] [CrossRef] [Green Version]
- Ahmmed, M.K.; Ahmmed, F.; Tian, H.; Carne, A.; Bekhit, A.E.D. Marine omega-3 (n-3) phospholipids: A comprehensive review of their properties, sources, bioavailability, and relation to brain health. Compr. Rev. Food Sci. Food Saf. 2020, 19, 64–123. [Google Scholar] [CrossRef] [Green Version]
- Masilan, K.; Neethiselvan, N.; Sivaraman, B.; Ravikumar, T.; Nagarajan, M.; Karthy, A.; Sukumar, D. Valorization of discarded industrial fish processing wastes for the extraction of gelatin to use as biodegradable fish bait matrix using RSM. PeerJ Mater. Sci. 2021, 3, e14. [Google Scholar] [CrossRef]
- Ghaly, A.E.; Ramakrishnan, V.V.; Brooks, M.S.; Budge, S.M.; Dave, D. Fish processing wastes as a potential source of proteins. Amino acids and oils: A critical review. J. Microb. Biochem. Technol. 2013, 5, 107–129. [Google Scholar]
- Ahmmed, M.K.; Bunga, S.; Stewart, I.; Tian, H.; Carne, A.; Bekhit, A.E.D.A. Simple and efficient one-pot extraction method for phospholipidomic profiling of total oil and lecithin by phosphorus-31 nuclear magnetic resonance measurements. J. Agric. Food Chem. 2020, 68, 14286–14296. [Google Scholar] [CrossRef] [PubMed]
- Ahmmed, M.K.; Ahmmed, F.; Stewart, I.; Carne, A.; Tian, H.S.; Bekhit, A.E.D.A. Omega-3 phospholipids in Pacific blue mackerel (Scomber australasicus) processing by-products. Food Chem. 2021, 353, 129451. [Google Scholar] [CrossRef]
- Ahmmed, M.K.; Carne, A.; Ahmmed, F.; Stewart, I.; Tian, H.S.; Bekhit, A.E.D.A. Positional distribution of fatty acids and phospholipid composition in King salmon (Oncorhynchus tshawytscha) head, roe and skin using nuclear magnetic resonance spectroscopy. Food Chem. 2021, 363, 130302. [Google Scholar] [CrossRef]
- Ahmmed, M.K.; Carne, A.; Bunga, S.; Tian, H.S.; Bekhit, A.E.D.A. Lipidomic signature of Pacific lean fish species head and skin using gas chromatography and nuclear magnetic resonance spectroscopy. Food Chem. 2021, 365, 130637. [Google Scholar] [CrossRef]
- Hanna, R.G. Proximate composition of certain Red Sea fishes. Mar. Fish. Rev. 1984, 46, 71–75. [Google Scholar]
- Ould El Kebir, M.V.; Barnathan, G.; Siau, Y.; Miralles, J.; Gaydou, E.M. Fatty acid distribution in muscle, liver, and gonads of rays (Dasyatis marmorata, Rhinobatos cemiculus, and Rhinoptera marginata) from the East Tropical Atlantic Ocean. J. Agric. Food Chem. 2003, 51, 1942–1947. [Google Scholar] [CrossRef]
- Urquidez-Bejarano, P.; Perez-Velazquez, M.; González-Félix, M.L.; Castro-Longoria, R. Fatty acid and proximate composition of wild male and female king angelfish (Holacanthus passer) gonads during the ripe and spent developmental stages. Anim. Reprod. 2018, 13, 820–829. [Google Scholar] [CrossRef]
- Jonsson, N.; Jonsson, B.; Hansen, L.P. Changes in proximate composition and estimates of energetic costs during upstream migration and spawning in Atlantic salmon Salmo salar. J. Anim. Ecol. 1997, 66, 425–436. [Google Scholar] [CrossRef]
- FAO. Global Production Statistics 1950–2019. 2021. Available online: https://www.fao.org/figis/servlet/TabSelector (accessed on 2 December 2021).
- Polikovsky, M.; Fernand, F.; Sack, M.; Frey, W.; Müller, G.; Golberg, A. Towards marine biorefineries: Selective proteins extractions from marine macroalgae Ulva with pulsed electric fields. Innov. Food Sci. Emerg. Technol. 2016, 37, 194–200. [Google Scholar] [CrossRef]
- Prabhu, M.S.; Levkov, K.; Livney, Y.D.; Israel, A.; Golberg, A. High-voltage pulsed electric field preprocessing enhances extraction of starch, proteins, and ash from marine macroalgae ulva ohnoi. ACS Sustain. Chem. Eng. 2019, 7, 17453–17463. [Google Scholar] [CrossRef]
- Gómez, B.; Munekata, P.E.; Gavahian, M.; Barba, F.J.; Martí-Quijal, F.J.; Bolumar, T.; Campagnol, P.C.B.; Tomasevic, I.; Lorenzo, J.M. Application of pulsed electric fields in meat and fish processing industries: An overview. Food Res. Int. 2019, 123, 95–105. [Google Scholar] [CrossRef]
- Bekhit, A.E.D.A.; van de Ven, R.; Fahri, F.; Hopkins, D.L. Effect of pulsed electric field treatment on cold-boned muscles of different potential tenderness. Food Bioprocess Technol. 2014, 7, 3136–3146. [Google Scholar] [CrossRef]
- He, G.; Yin, Y.; Yan, X.; Wang, Y. Semi-Bionic Extraction of Effective Ingredient from Fishbone by High Intensity Pulsed Electric Fields. J. Food Process Eng. 2017, 40, e12392. [Google Scholar] [CrossRef]
- Zhou, Y.; He, Q.; Zhou, D. Optimization extraction of protein from mussel by high-Intensity pulsed electric fields. J. Food Processing Preserv. 2017, 41, e12962. [Google Scholar] [CrossRef]
- Buchmann, L.; Mathys, A. Perspective on pulsed electric field treatment in the bio-based industry. Front. Bioeng. Biotechnol. 2019, 7, 265. [Google Scholar] [CrossRef]
- Folch, J.; Lees, M.; Stanley, G.S. A simple method for the isolation and purification of total lipides from animal tissues. J. Biol. Chem. 1957, 226, 497–509. [Google Scholar] [CrossRef]
- Bligh, G.E.; Dyer, W.J. A rapid method of total lipid extraction and purification. Can. J. Biochem. Physiol. 1959, 37, 911–917. [Google Scholar] [CrossRef] [PubMed]
- Bah, C.S.F.; Bekhit, A.E.D.A.; Fang, E.F.; Ng, T.B.; McConnell, M.A.; Bekhit, A.A.; Morton, J.D. Physicochemical properties and bioactivity of extracts from the roe of new zealand hoki and southern blue whiting. J. Aquat. Food Prod. Technol. 2016, 25, 1234–1248. [Google Scholar] [CrossRef]
- MacDonald, G.A.; Hall, B.I.; Vlieg, P. Seasonal Changes in Hoki (Macruronus novaezelandaie) Implications for Quality and Yield. J. Aquat. Food Prod. Technol. 2002, 11, 35–51. [Google Scholar] [CrossRef]
- Chvalová, D.; Špička, J. Identification of furan fatty acids in the lipids of common carp (Cyprinus carpio L.). Food Chem. 2016, 200, 183–188. [Google Scholar] [CrossRef] [PubMed]
- Pekkarinen, M. Seasonal variations in lipid content and fatty acids in the liver, muscle and gonads of the eel-pout, Zoarces viviparus (Teleostei) in brackish water. In Annales Zoologici Fennici; JSTOR: New York, NY, USA, 1980. [Google Scholar]
- Falch, E.; Størseth, T.R.; Aursand, M. Multi-component analysis of marine lipids in fish gonads with emphasis on phospholipids using high resolution NMR spectroscopy. Chem. Phys. Lipids 2006, 144, 4–16. [Google Scholar] [CrossRef] [PubMed]
- Hiratsuka, S.; Kitagawa, T.; Matsue, Y.; Hashidume, M.; Wada, S. Lipid class and fatty acid composition of phospholipids from the gonads of skipjack tuna. Fish. Sci. 2004, 70, 903–909. [Google Scholar] [CrossRef]
- Li, S.; Wen, W.; Gong, X.; Huang, X.; Chen, N. Variation of lipids and fatty acids composition in the tissues of wild devil stinger (Inimicus japonicas) during sexual maturation. Aquac. Fish. 2018, 3, 115–121. [Google Scholar] [CrossRef]
- Guderjan, M.; Elez-Martínez, P.; Knorr, D. Application of pulsed electric fields at oil yield and content of functional food ingredients at the production of rapeseed oil. Innov. Food Sci. Emerg. Technol. 2007, 8, 55–62. [Google Scholar] [CrossRef]
- Li, M.; Lin, J.I.E.; Chen, J.; Fang, T. Pulsed Electric Field-Assisted Enzymatic Extraction of Protein from Abalone (Haliotis Discus Hannai Ino) Viscera. J. Food Process Eng. 2016, 39, 702–710. [Google Scholar] [CrossRef]
- Bhardwaj, S.; Passi, S.J.; Misra, A.; Pant, K.K.; Anwar, K.; Pandey, R.M.; Kardam, V. Effect of heating/reheating of fats/oils, as used by Asian Indians, on trans fatty acid formation. Food Chem. 2016, 212, 663–670. [Google Scholar] [CrossRef]
- Watanabe, M.; Iida, T.; Inomata, H. Decomposition of a long chain saturated fatty acid with some additives in hot compressed water. Energy Convers. Manag. 2006, 47, 3344–3350. [Google Scholar] [CrossRef]
- Abd El-Hay, M.M. Processing and preparation of fish. In Postharvest and Postmortem Processing of Raw Food Materials; Elsevier: Amsterdam, The Netherlands, 2022; pp. 315–342. [Google Scholar]
- Tenyang, N. Effect of Cooking Methods on the Nutritive Value and Lipid Oxidation of Two Cricket Species Consumed In Cameroon. Eur. J. Nutr. Food Saf. 2021, 13, 11–23. [Google Scholar] [CrossRef]
- Bledsoe, G.; Bledsoe, C.; Rasco, B. Caviars and fish roe products. Crit. Rev. Food Sci. Nutr. 2003, 43, 317–356. [Google Scholar] [CrossRef]
- Simopoulos, A.P. An increase in the omega-6/omega-3 fatty acid ratio increases the risk for obesity. Nutrients 2016, 8, 128. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Prabhakara Rao, P.G.; Balaswamy, K.; Narsing Rao, G.; Jyothirmayi, T. Lipid classes, fatty acid and phospholipid composition of roe lipids from Catla catla and Cirrhinus mrigala. Int. Food Res. J. 2013, 20, 275–279. [Google Scholar]
- Chakrabarti, A. Phospholipid Asymmetry in Biological Membranes: Is the Role of Phosphatidylethanolamine Underappreciated? J. Membr. Biol. 2021, 254, 127–132. [Google Scholar] [CrossRef]
- Bazzi, M.D.; Youakim, M.A.; Nelsestuen, G.L. Importance of phosphatidylethanolamine for association of protein kinase C and other cytoplasmic proteins with membranes. Biochemistry 1992, 31, 1125–1134. [Google Scholar] [CrossRef]
- van der Veen, J.N.; Kennelly, J.P.; Wan, S.; Vance, J.E.; Vance, D.E.; Jacobs, R.L. The critical role of phosphatidylcholine and phosphatidylethanolamine metabolism in health and disease. Biochim. Biophys. Acta BBA-Biomembr. 2017, 1859, 1558–1572. [Google Scholar] [CrossRef]
- Takama, K.; Suzuki, T.; Yoshida, K.; Arai, H.; Mitsui, T. Phosphatidylcholine levels and their fatty acid compositions in teleost tissues and squid muscle. Comp. Biochem. Physiol. Part B Biochem. Mol. Biol. 1999, 124, 109–116. [Google Scholar] [CrossRef]
- Shen, Z.; Ye, C.; McCain, K.; Greenberg, M.L. The role of cardiolipin in cardiovascular health. BioMed Res. Int. 2015, 2015, 891707. [Google Scholar] [CrossRef] [Green Version]
- Body, D.R. The lipid composition of the roe tissues from four common New Zealand marine fish species. J. Food Compos. Anal. 1989, 2, 350–355. [Google Scholar] [CrossRef]
- Tilly, J.L.; Kolesnick, R.N. Sphingolipid signaling in gonadal development and function. Chem. Phys. Lipids 1999, 102, 149–155. [Google Scholar] [CrossRef]
- Tanaka, Y.; Matsuwaki, T.; Yamanouchi, K.; Nishihara, M. Increased lysosomal biogenesis in activated microglia and exacerbated neuronal damage after traumatic brain injury in progranulin-deficient mice. Neuroscience 2013, 250, 8–19. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
| Percentage | HMG (w/w) | HMG (d/w) |
|---|---|---|
| Moisture | 81.5 ± 0.53 | - |
| Protein a | 11.8 ± 0.78 | 63.8 ± 0.31 |
| Lipid | 4.1 ± 0.12 | 22.2 ± 1.11 |
| Ash | 1.3 ± 0.21 | 7.0 ± 0.13 |
| Carbohydrate b | 1.3 ± 0.62 | 7.0 ± 1.10 |
| Treatments | Temperature (°C) | Conductivity (S/m) | Specific Energy Input (kJ/kg) |
|---|---|---|---|
| Electric field strength (kV/cm) | |||
| 0.625 | 15.6 c | 16.7 | 90.2 c |
| 1.25 | 21.6 b | 17.4 | 222.4 b |
| 1.875 | 34.1 a | 16.9 | 313.6 a |
| SEM | 1.2 | 0.6 | 6.9 |
| p value | 0.001 | 0.72 | 0.001 |
| Frequency (Hz) | |||
| 25 | 18.0 c | 17.2 | 102.2 c |
| 50 | 23.0 b | 16.6 | 196.2 b |
| 100 | 30.3 a | 17.2 | 327.9 a |
| SEM | 1.2 | 0.6 | 6.9 |
| p value | 0.001 | 0.66 | 0.001 |
| Interaction | |||
| 0.625 × 25 | 15.1 d | 17.1 | 29.5 e |
| 0.625 × 50 | 16.9 d | 16.5 | 80.7 de |
| 0.625 × 100 | 17.8 cd | 16.6 | 160.6 c |
| 1.25 × 25 | 16.5 d | 17.9 | 99.3d |
| 1.25 × 50 | 20.2 cd | 16.5 | 181.3 c |
| 1.25 × 100 | 28.2 bc | 17.8 | 386.5 ab |
| 1.875 × 25 | 22.6 cd | 16.7 | 177.7 c |
| 1.875 × 50 | 34.9 ab | 16.9 | 326.7 b |
| 1.875 × 100 | 45.0 a | 17.2 | 436.5 a |
| SEM | 2.1 | 1.03 | 12.0 |
| p value | 0.004 | 0.94 | 0.001 |
| Fatty Acid | Un-Heated Control | Heat-Treated Control | p |
|---|---|---|---|
| C14:0 | 0.97 | 1.18 | 0.16 |
| C14:1 | 0.07 a | 0.00 b | 0.001 |
| C15:1 | 0.28 b | 0.32 a | 0.01 |
| C16:0 | 17.7 b | 19.5 a | 0.001 |
| C16:1 n7 | 1.97 b | 2.26 a | 0.02 |
| C17:0 | 0.30 | 0.30 | 0.14 |
| C17:1 | 0.37 a | 0.32 b | 0.001 |
| C18:0 | 4.51 | 4.52 | 0.82 |
| C18:1 n9 trans | 12.6 | 13.7 | 0.11 |
| C18:1 n9 cis | 3.71 b | 4.15 a | 0.04 |
| C18:2 n6 trans | 0.17 | 0.18 | 0.08 |
| C18:2 n6 cis | 0.60 | 0.64 | 0.09 |
| C18:3 n6 | 0.10 a | 0.00 b | 0.001 |
| C18:3 n3 | 0.19 | 0.22 | 0.12 |
| C20:1 | 0.20 b | 0.26 a | 0.03 |
| C20:2 n6 | 1.98 | 2.51 | 0.07 |
| C21:0 | 0.21 | 0.22 | 0.24 |
| C20:3 n3 | 2.38 a | 2.05 b | 0.01 |
| C20:4 n6 | 0.13 | 0.14 | 0.39 |
| C22:0 | 0.86 | 0.95 | 0.09 |
| C22:1 n9 | 0.25 | 0.22 | 0.24 |
| C20:5 n3 [EPA] | 8.01 | 7.87 | 0.54 |
| C23:0 | 0.12 | 0.16 | 0.34 |
| C24:0 | 0.39 a | 0.34 b | 0.001 |
| C24:1 | 0.13 | 0.18 | 0.12 |
| C22:5 n3 [DPA] | 2.53 | 2.50 | 0.57 |
| C22:6 n3 [DHA] | 35.2 | 33.9 | 0.31 |
| Unknown | 4.07 | 3.32 | 0.10 |
| MUFA | 19.6 | 21.5 | 0.06 |
| PUFA | 51.3 | 50.1 | 0.38 |
| SFA | 25.1 b | 27.2 a | 0.001 |
| n-3 | 48.3 | 46.6 | 0.27 |
| n-6 | 3.00 | 3.48 | 0.12 |
| n6/n3 | 0.06 | 0.07 | 0.12 |
| PL | Control | Heat-Treated Control | p |
|---|---|---|---|
| LDPG | 0.23 | 0.24 | 0.87 |
| PG | 0.13 b | 0.16 a | 0.02 |
| CL N-acyl PE | 0.86 a 0.32 a | 0.62 b 0.00 b | 0.00 0.00 |
| LPE | 0.10 a | 0.08 b | 0.00 |
| LPSP | 0.49 | 0.46 | 0.14 |
| LPS | 0.19 | 0.25 | 0.45 |
| SM | 0.38 | 0.38 | 0.75 |
| PE | 3.90 | 3.64 | 0.32 |
| LPC-1 | 0.57 a | 0.33 b | 0.00 |
| PS | 0.86 | 0.80 | 0.07 |
| LPC-2 | 0.09 a | 0.00 b | 0.00 |
| PI | 0.15 | 0.16 | 0.62 |
| PC | 7.78 | 8.09 | 0.56 |
| Fatty Acid | PEF Frequency | PEF Field Strength | PEF Interaction | p | |||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 25 | 50 | 100 | 0.625 | 1.25 | 1.875 | 25 × 0.625 | 25 × 1.25 | 25 × 1.875 | 50 × 0.625 | 50 × 1.25 | 50 × 1.875 | 100 × 0.625 | 100 × 1.25 | 100 × 1.875 | SEM | Freq | PEF | FRE*PEF | |
| C14:0 | 1.16 a | 1.11 ab | 1.02 b | 1.04 b | 1.28 a | 0.97 b | 1.08 cd | 1.37 a | 1.02 cd | 1.00 cd | 1.35 ab | 0.99 cd | 1.03 cd | 1.13 bc | 0.89 d | 0.03 | 0.004 | 0.001 | 0.13 |
| C15:1 | 0.30 | 0.30 | 0.29 | 0.29 b | 0.33 a | 0.27 c | 0.29 cde | 0.34 a | 0.27 de | 0.30 bcd | 0.33 ab | 0.27 de | 0.29 bcde | 0.31 abc | 0.26 e | 0.01 | 0.69 | 0.001 | 0.29 |
| C16:0 | 18.4 b | 19.2 a | 19.1 a | 19.9 a | 19.4 a | 17.3 b | 18.8 c | 19.1 bc | 17.3 d | 20.9 a | 19.3 bc | 17.4 d | 20.0 ab | 19.9 abc | 17.3 d | 0.23 | 0.002 | 0.001 | 0.002 |
| C16:1 n7 | 2.15 a | 2.12 a | 2.02 b | 2.03 b | 2.25 a | 2.01 b | 2.10 c | 2.32 a | 2.04 c | 1.99 c | 2.32 ab | 2.04 c | 2.00 c | 2.11 bc | 1.94 c | 0.04 | 0.002 | 0.001 | 0.10 |
| C17:0 | 0.29 | 0.30 | 0.30 | 0.32 a | 0.30 ab | 0.28 b | 0.30 ab | 0.30 ab | 0.28 b | 0.34 a | 0.29 ab | 0.28 b | 0.31 ab | 0.30 ab | 0.28 ab | 0.01 | 0.67 | 0.002 | 0.33 |
| C17:0 | 0.29 | 0.29 | 0.30 | 0.31 a | 0.30 a | 0.28 b | 0.30 abc | 0.30 ab | 0.28 cd | 0.31 ab | 0.29 abcd | 0.28 d | 0.31 a | 0.30 abc | 0.28 bcd | 0.01 | 0.29 | 0.001 | 0.55 |
| C17:1 | 0.36 a | 0.33 ab | 0.30 b | 0.31 | 0.32 | 0.35 | 0.35 a | 0.37 a | 0.35 a | 0.30 a | 0.32 a | 0.37 a | 0.29 a | 0.27 a | 0.34 a | 0.02 | 0.02 | 0.08 | 0.42 |
| C18:0 | 4.40 b | 4.43 b | 4.53 a | 4.54 a | 4.44 b | 4.38 b | 4.44 abc | 4.40 bc | 4.35 c | 4.58 ab | 4.38 bc | 4.33 c | 4.60 a | 4.55 ab | 4.46 abc | 0.04 | 0.001 | 0.001 | 0.22 |
| C18:1 n9 trans | 12.8 | 12.9 | 12.6 | 12.8 a | 13.1 a | 12.4 b | 12.8 ab | 13.3 a | 12.3 b | 12.9 ab | 13.3 a | 12.7 ab | 12.8 ab | 12.8 ab | 12.3 b | 0.17 | 0.11 | 0.001 | 0.40 |
| C18:1 n9 cis | 3.86 b | 4.00 a | 4.05 a | 4.16 a | 4.00 b | 3.74 c | 3.96 bc | 3.89 cd | 3.74 cd | 4.36 a | 3.94 bcd | 3.69 d | 4.17 ab | 4.17 ab | 3.80 cd | 0.05 | 0.001 | 0.001 | 0.002 |
| C18:3 n3 | 0.22 a | 0.20 ab | 0.20 b | 0.19 b | 0.22 a | 0.21 ab | 0.20 ab | 0.24 a | 0.21 ab | 0.18 b | 0.22 ab | 0.21 ab | 0.19 b | 0.20 b | 0.20 b | 0.01 | 0.004 | 0.01 | 0.23 |
| C20:1 | 0.23 a | 0.21 ab | 0.20 b | 0.19 b | 0.23 a | 0.21 a | 0.21 abc | 0.25 a | 0.22 abc | 0.19 bc | 0.24 ab | 0.22 abc | 0.19 c | 0.21 abc | 0.21 abc | 0.01 | 0.01 | 0.001 | 0.54 |
| C20:2 n6 | 2.32 a | 2.17 ab | 2.05 b | 2.13 | 2.18 | 2.23 | 2.29 ab | 2.31 ab | 2.38 a | 2.04 ab | 2.24 ab | 2.23 ab | 2.07 ab | 2.00 b | 2.07 ab | 0.07 | 0.001 | 0.33 | 0.58 |
| C21:0 | 0.21 | 0.20 | 0.20 | 0.20 | 0.21 | 0.20 | 0.20 | 0.22 | 0.20 | 0.20 | 0.21 | 0.20 | 0.20 | 0.20 a | 0.20 a | 0.01 | 0.30 | 0.21 | 0.24 |
| C20:3 n3 | 2.11 | 2.12 | 2.19 | 2.06 b | 2.13 ab | 2.22 a | 2.06 b | 2.08 b | 2.18 ab | 2.03 b | 2.18 ab | 2.16 ab | 2.09 b | 2.14 ab | 2.33 a | 0.05 | 0.07 | 0.002 | 0.26 |
| C20:4 n6 | 0.13 b | 0.15 a | 0.08 c | 0.13 a | 0.11 b | 0.13 a | 0.13 b | 0.14 b | 0.13 b | 0.13 b | 0.19 a | 0.13 b | 0.13 b | 0.00 c | 0.13 b | 0.00 | 0.001 | 0.001 | 0.001 |
| C22:0 | 0.92 a | 0.90 ab | 0.87 b | 0.85 b | 0.90 b | 0.94 a | 0.91 abc | 0.90 abc | 0.97 a | 0.83 bc | 0.93 abc | 0.95 ab | 0.83 c | 0.87 bc | 0.91 abc | 0.02 | 0.006 | 0.001 | 0.32 |
| C22:1 | 0.30 a | 0.24 ab | 0.23 b | 0.26 | 0.26 | 0.25 | 0.32 ab | 0.34 a | 0.24 ab | 0.22 ab | 0.24 ab | 0.27 ab | 0.23 ab | 0.21 b | 0.24 ab | 0.03 | 0.004 | 0.87 | 0.04 |
| C20:5 n3[EPA] | 8.03 | 8.04 | 7.99 | 7.69 b | 8.09 a | 8.27 a | 7.85 bcd | 7.97 abcd | 8.27 ab | 7.61 cd | 8.30 abc | 8.20 ab | 7.61 d | 8.00 abcd | 8.34 a | 0.09 | 0.76 | 0.001 | 0.08 |
| C23:0 | 0.17 a | 0.11 b | 0.10 b | 0.16 b | 0.05 c | 0.17 a | 0.17 abc | 0.16 cd | 0.18 ab | 0.15 cd | 0.00 e | 0.18 a | 0.15 d | 0.00 e | 0.16 bcd | 0.00 | 0.001 | 0.001 | 0.001 |
| C24:0 | 0.34 | 0.34 | 0.35 | 0.34 | 0.35 | 0.35 | 0.33 a | 0.36 a | 0.34 a | 0.33 a | 0.35 a | 0.34 a | 0.34 a | 0.34 a | 0.36 a | 0.01 | 0.25 | 0.12 | 0.05 |
| C24:1 | 0.16 a | 0.08 b | 0.08 b | 0.13 a | 0.06 b | 0.13 a | 0.15 a | 0.17 a | 0.15 a | 0.11 ab | 0.00 b | 0.13 a | 0.13 a | 0.00 b | 0.11 a | 0.02 | 0.001 | 0.001 | 0.001 |
| C22:5 n3[DPA] | 2.57 | 2.57 | 2.57 | 2.54 b | 2.49 b | 2.68 a | 2.60 abc | 2.42 d | 2.70 a | 2.51 abcd | 2.56 abcd | 2.66 ab | 2.51 bcd | 2.50 cd | 2.70 a | 0.03 | 0.99 | 0.001 | 0.04 |
| C22:6 n3[DHA] | 34.4 b | 34.9 ab | 35.1 a | 34.0 b | 34.6 b | 35.8 a | 34.1 cd | 33.8 d | 35.4 abc | 33.8 bcd | 35.3 abcd | 35.7 ab | 34.2 bcd | 34.8 abcd | 36.2 a | 0.36 | 0.06 | 0.001 | 0.28 |
| Unknown | 3.64 a | 2.86 b | 2.94 b | 3.17 ab | 2.66 b | 3.60 a | 3.47 ab | 3.56 a | 3.88 a | 3.04 abc | 2.10 bc | 3.44 ab | 3.01 abc | 2.33 c | 3.48 a | 0.22 | 0.001 | 0.001 | 0.14 |
| MUFA | 20.2 | 20.3 | 19.8 | 20.2 a | 20.6 a | 19.4 b | 20.2 abc | 21.0 a | 19.3 c | 20.3 abc | 20.7 ab | 19.7 bc | 20.0 abc | 20.1 abc | 19.2 c | 0.23 | 0.04 | 0.001 | 0.42 |
| PUFA | 50.7 | 50.3 | 50.9 | 49.5 b | 50.0 b | 52.4 a | 50.1 bc | 49.8 bc | 52.1 ab | 48.9 c | 49.8 bc | 52.2 ab | 49.5 c | 50.3 bc | 52.9 a | 0.50 | 0.34 | 0.001 | 0.59 |
| SFA | 25.9 b | 26.4 ab | 26.5 a | 27.2 a | 27.0 a | 24.6 b | 26.3 b | 26.9 ab | 24.7 c | 27.8 a | 26.7 ab | 24.7 c | 27.5 a | 27.3 ab | 24.6 c | 0.24 | 0.02 | 0.001 | 0.01 |
| n-3 | 47.4 | 47.2 | 48.0 | 46.4 b | 47.0 b | 49.2 a | 46.8 bc | 46.5 bc | 48.8 abc | 45.9 c | 46.8 bc | 48.9 ab | 46.6 bc | 47.7 abc | 49.8 a | 0.55 | 0.18 | 0.001 | 0.69 |
| n-6 | 3.31 a | 3.07 b | 2.91 b | 3.08 ab | 2.97 b | 3.24 a | 3.32 a | 3.29 a | 3.32 a | 2.95 ab | 2.99 ab | 3.27 a | 2.96 ab | 2.64 b | 3.12 a | 0.08 | 0.001 | 0.004 | 0.07 |
| n6/n3 | 0.07 a | 0.07 ab | 0.06 b | 0.07 | 0.06 | 0.07 | 0.07 a | 0.07 a | 0.07 a | 0.06 ab | 0.06 ab | 0.07 a | 0.06 ab | 0.06 b | 0.06 ab | 0.00 | 0.001 | 0.28 | 0.20 |
| PL | Freq | PEF | Interaction | SEM | P | ||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 25 | 50 | 100 | 0.625 | 1.25 | 1.875 | 25 × 0.625 | 25 × 1.25 | 25 × 1.875 | 50 × 0.625 | 50 × 1.25 | 50 × 1.875 | 100 × 0.625 | 100 × 1.25 | 100 × 1.875 | Freq | PEF | FRE*PEF | ||
| LDPG | 0.27 | 0.35 | 0.35 | 0.27 | 0.35 | 0.35 | 0.16 b | 0.29 ab | 0.37 ab | 0.33 ab | 0.39 a | 0.33 ab | 0.33 ab | 0.38 a | 0.34 ab | 0.04 | 0.06 | 0.06 | 0.15 |
| PG | 0.13 | 0.13 | 0.13 | 0.15 | 0.13 | 0.12 | 0.16 | 0.12 | 0.11 | 0.14 | 0.13 | 0.13 | 0.13 | 0.14 | 0.12 | 0.01 | 0.99 | 0.07 | 0.28 |
| CL | 0.85 | 0.81 | 0.79 | 0.85 | 0.84 | 0.76 | 0.99 | 0.79 | 0.79 | 0.79 | 0.85 | 0.80 | 0.77 | 0.89 | 0.69 | 0.08 | 0.61 | 0.41 | 0.32 |
| LPE | 0.10 | 0.09 | 0.09 | 0.10 | 0.09 | 0.09 | 0.11 | 0.09 | 0.09 | 0.09 | 0.10 | 0.10 | 0.09 | 0.09 | 0.08 | 0.01 | 0.44 | 0.92 | 0.59 |
| LPSP | 0.50 | 0.48 | 0.45 | 0.49 | 0.49 | 0.45 | 0.58 | 0.44 | 0.47 | 0.47 | 0.51 | 0.47 | 0.42 a | 0.5 | 0.41 | 0.04 | 0.42 | 0.41 | 0.07 |
| LPS | 0.29 b | 0.37 ab | 0.40 a | 0.28 b | 0.40 a | 0.38 a | 0.18 b | 0.33 ab | 0.38 ab | 0.28 ab | 0.46 a | 0.36 ab | 0.37 ab | 0.42 a | 0.41 a | 0.04 | 0.03 | 0.01 | 0.30 |
| SM | 0.38 | 0.45 | 0.36 | 0.40 | 0.43 | 0.37 | 0.37 | 0.39 | 0.39 | 0.48 | 0.48 | 0.39 | 0.34 | 0.41 | 0.33 | 0.07 | 0.30 | 0.61 | 0.94 |
| PE | 3.75 | 3.45 | 3.22 | 3.81 | 3.31 | 3.30 | 4.65 a | 3.32 ab | 3.29 ab | 3.37 ab | 3.35 ab | 3.62 ab | 3.43 ab | 3.26 ab | 2.98 b | 0.33 | 0.20 | 0.13 | 0.17 |
| LPC-1 | 0.53 | 0.49 | 0.49 | 0.51 | 0.52 a | 0.4 | 0.60 | 0.48 | 0.51 | 0.45 | 0.53 | 0.49 | 0.49 | 0.56 | 0.41 | 0.05 | 0.57 | 0.47 | 0.27 |
| PS | 0.78 | 0.74 | 0.72 | 0.80 | 0.75 | 0.69 | 0.94 | 0.72 | 0.67 | 0.73 | 0.75 | 0.74 | 0.74 | 0.76 | 0.66 | 0.07 | 0.64 | 0.19 | 0.26 |
| LPC-2 | 0.09 | 0.07 | 0.07 | 0.09 | 0.08 | 0.07 | 0.12 | 0.08 | 0.07 | 0.06 | 0.09 | 0.07 | 0.07 | 0.08 | 0.07 | 0.02 | 0.36 | 0.38 | 0.20 |
| PI | 0.13 | 0.12 | 0.12 | 0.13 | 0.12 | 0.12 | 0.15 | 0.09 | 0.09 | 0.12 | 0.12 | 0.13 | 0.11 | 0.14 | 0.14 | 0.02 | 0.58 | 0.90 | 0.20 |
| PC | 8.14 | 8.60 | 8.02 | 8.00 | 8.69 | 8.07 | 8.86 | 7.99 | 7.57 | 7.58 | 9.32 | 8.90 | 7.55 | 8.77 | 7.75 | 0.70 | 0.58 | 0.43 | 0.33 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Burnett, A.; Ahmmed, M.K.; Carne, A.; Tian, H.; Ahmed, I.A.M.; Al-Juhaimi, F.Y.; Bekhit, A.E.-D.A. Effect of Pulsed Electric Fields on the Lipidomic Profile of Lipid Extracted from Hoki Fish Male Gonad. Foods 2022, 11, 610. https://doi.org/10.3390/foods11040610
Burnett A, Ahmmed MK, Carne A, Tian H, Ahmed IAM, Al-Juhaimi FY, Bekhit AE-DA. Effect of Pulsed Electric Fields on the Lipidomic Profile of Lipid Extracted from Hoki Fish Male Gonad. Foods. 2022; 11(4):610. https://doi.org/10.3390/foods11040610
Chicago/Turabian StyleBurnett, Anna, Mirja Kaizer Ahmmed, Alan Carne, Hong (Sabrina) Tian, Isam A. Mohamed Ahmed, Fahad Y. Al-Juhaimi, and Alaa El-Din Ahmed Bekhit. 2022. "Effect of Pulsed Electric Fields on the Lipidomic Profile of Lipid Extracted from Hoki Fish Male Gonad" Foods 11, no. 4: 610. https://doi.org/10.3390/foods11040610
APA StyleBurnett, A., Ahmmed, M. K., Carne, A., Tian, H., Ahmed, I. A. M., Al-Juhaimi, F. Y., & Bekhit, A. E.-D. A. (2022). Effect of Pulsed Electric Fields on the Lipidomic Profile of Lipid Extracted from Hoki Fish Male Gonad. Foods, 11(4), 610. https://doi.org/10.3390/foods11040610