
Recent Advances in the Study of Wheat Protein and Other Food Components Affecting the Gluten Network and the Properties of Noodles



Abstract
1. Introduction
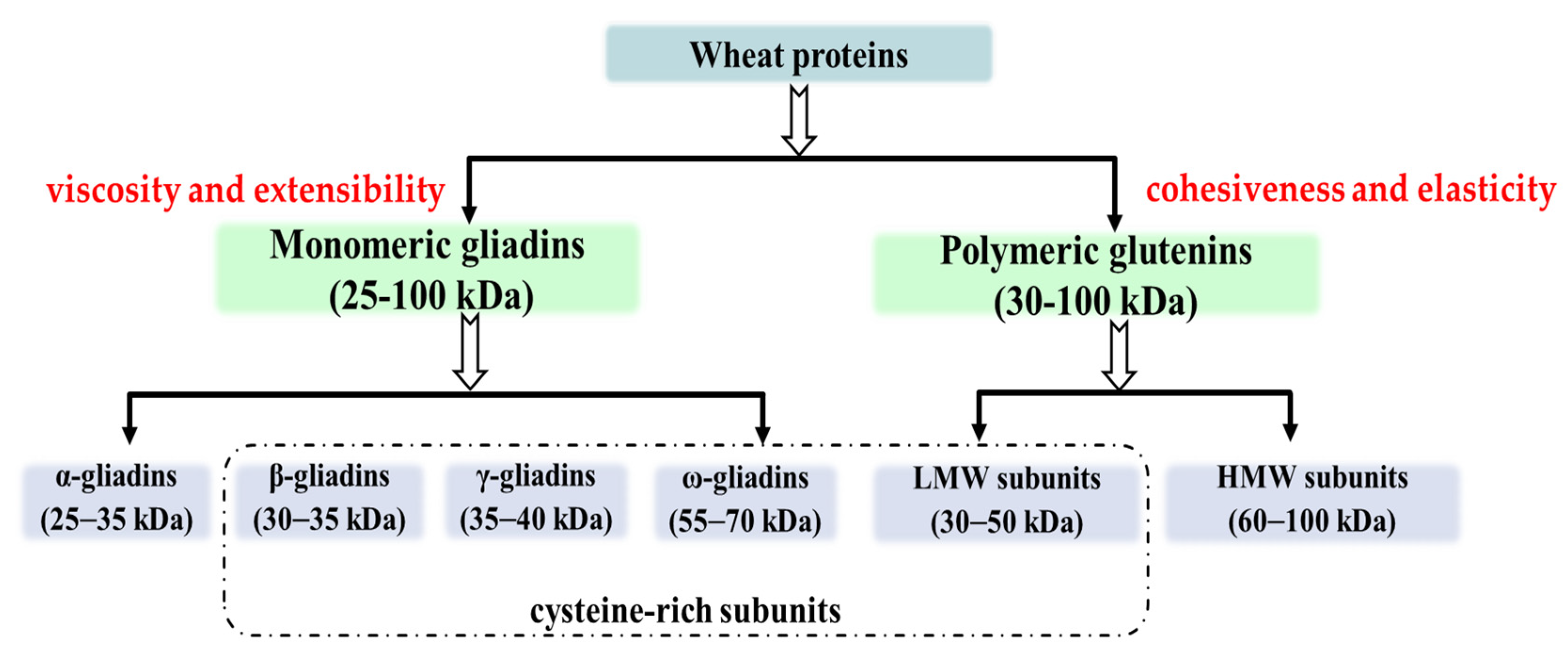
2. Structure and Function of Wheat Protein
2.1. Glutenin
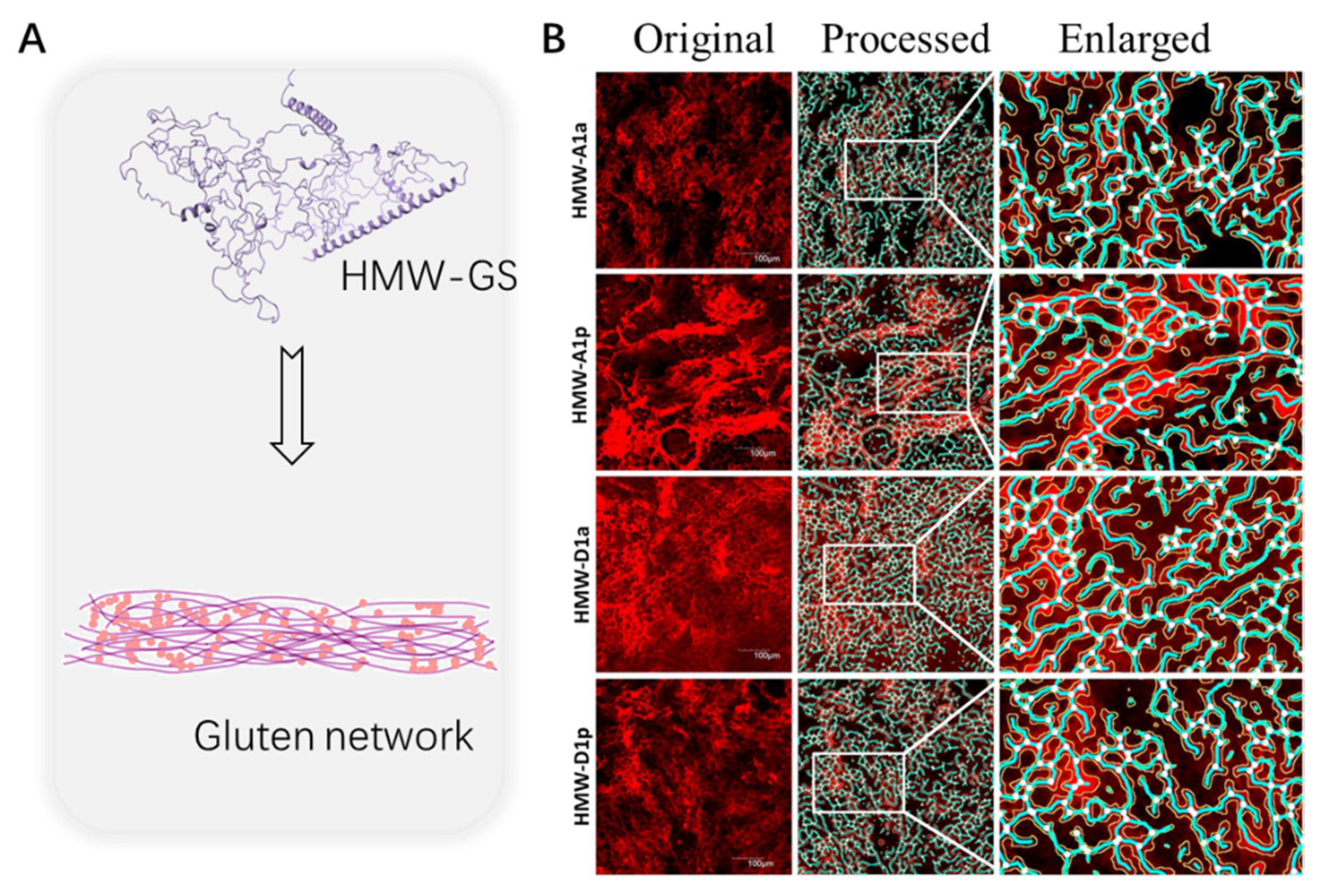
2.1.1. High-Molecular-Weight Glutenin Subunits
2.1.2. Low-Molecular-Weight Glutenin Subunits
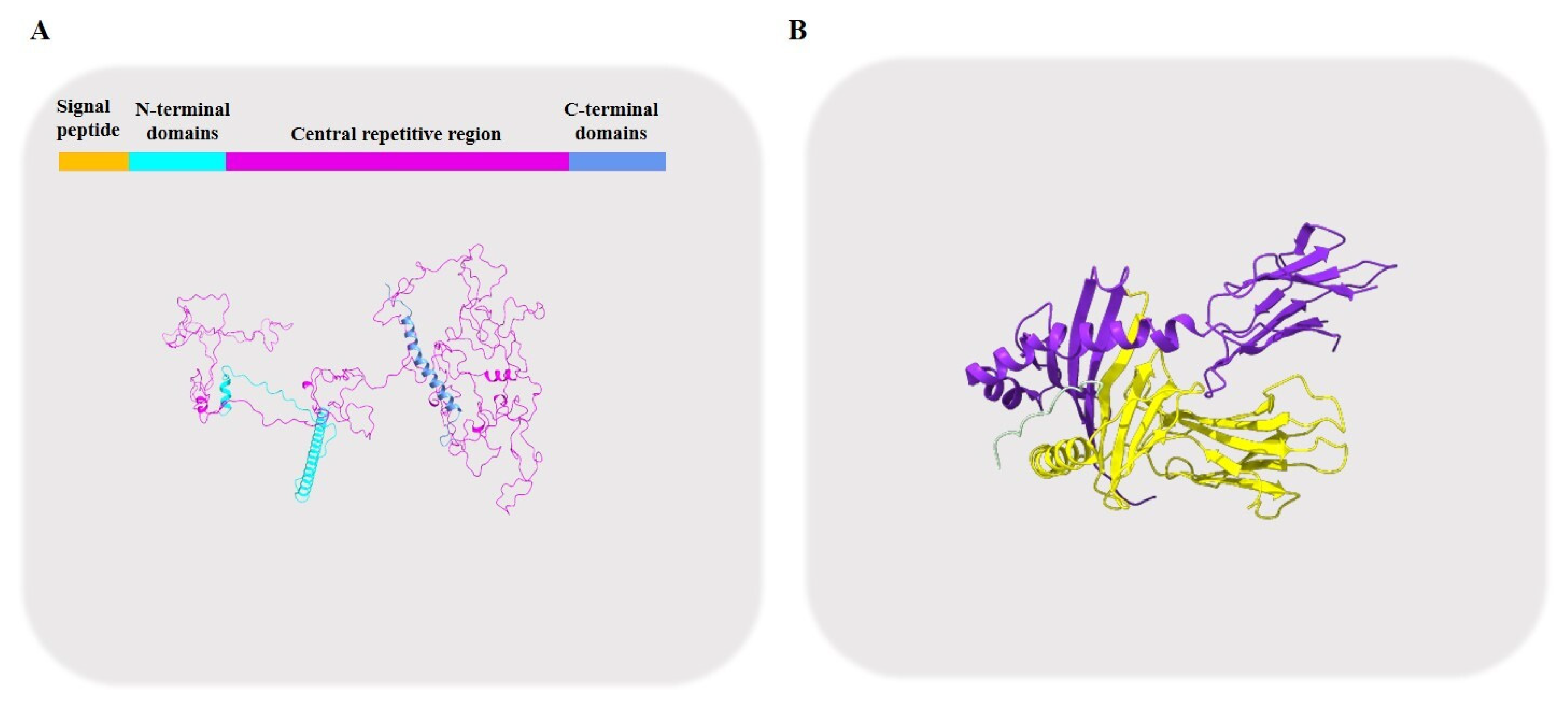
2.2. Gliadin
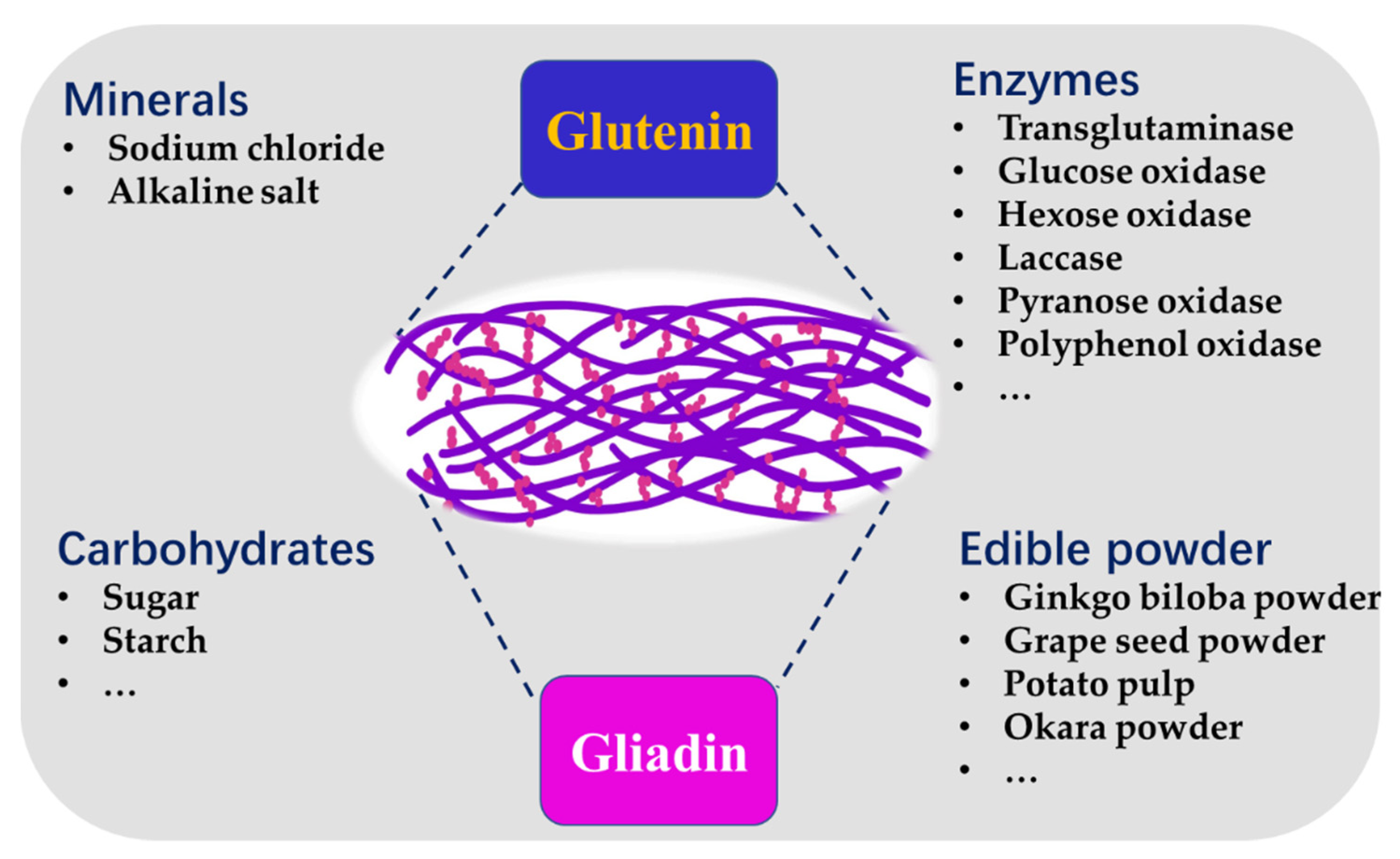
3. Food Components Affecting the Protein Network and the Properties of Noodles
3.1. Minerals
3.1.1. Sodium Chloride
3.1.2. Alkaline Salts
3.2. Carbohydrates
3.2.1. Starch
3.2.2. Sugar
3.3. Enzymes
3.3.1. Transglutaminase
3.3.2. Glucose Oxidase
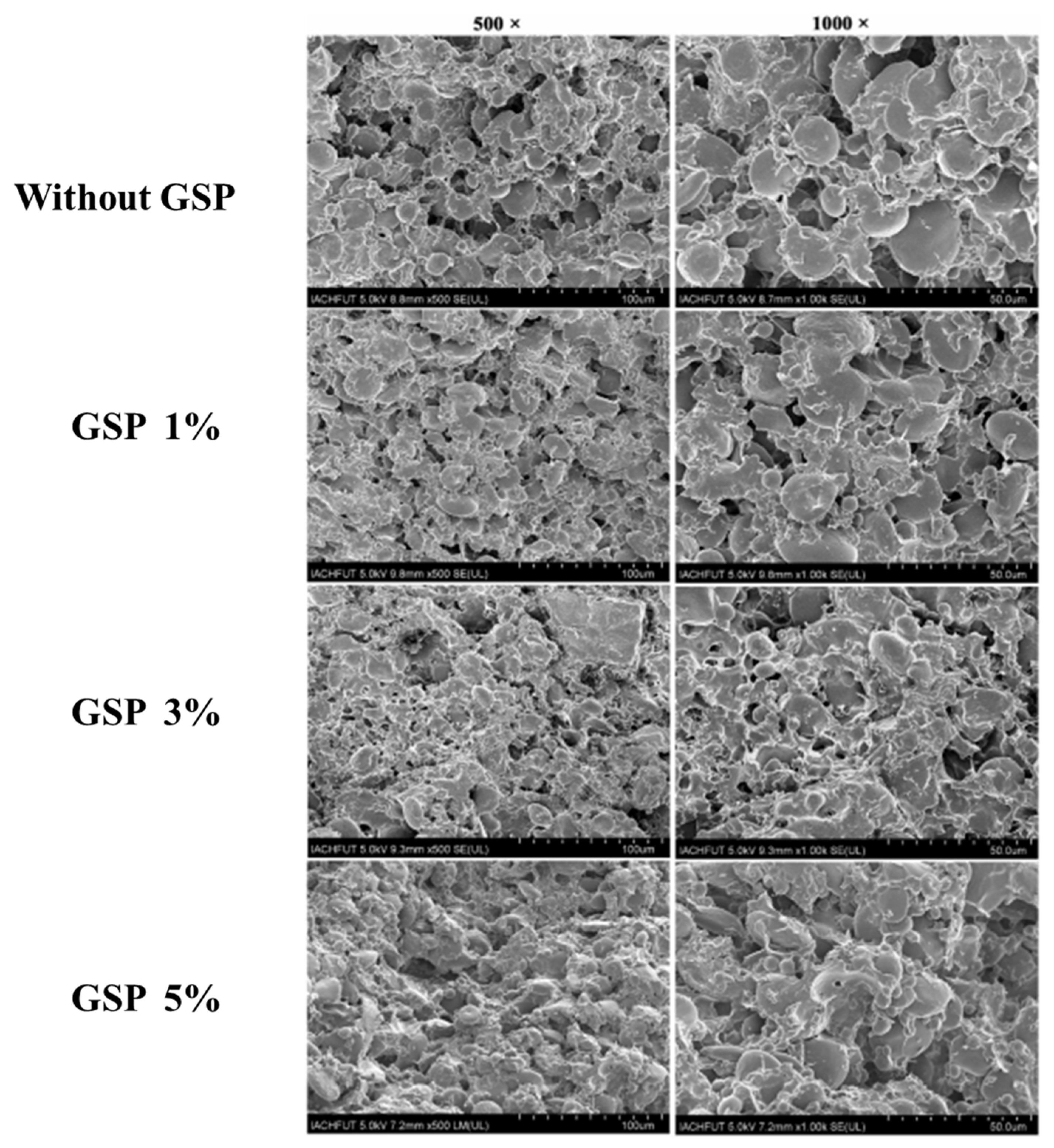
3.4. Edible Powder from Plant Sourcse
4. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Chen, S.-X.; Ni, Z.-J.; Thakur, K.; Wang, S.; Zhang, J.-G.; Shang, Y.-F.; Wei, Z.-J. Effect of Grape Seed Power on the Structural and Physicochemical Properties of Wheat Gluten in Noodle Preparation System. Food Chem. 2021, 355, 129500. [Google Scholar] [CrossRef]
- Obadi, M.; Zhang, J.; Shi, Y.; Xu, B. Factors Affecting Frozen Cooked Noodle Quality: A Review. Trends Food Sci. Technol. 2021, 109, 662–673. [Google Scholar] [CrossRef]
- Sun, K.-N.; Liao, A.-M.; Zhang, F.; Thakur, K.; Zhang, J.-G.; Huang, J.-H.; Wei, Z.-J. Microstructural, Textural, Sensory Properties and Quality of Wheat–Yam Composite Flour Noodles. Foods 2019, 8, 519. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.; Zhang, L.; Li, M.; Sun, Q. Inhibitory Effects of Sorbitol on the Collapse and Deterioration of Gluten Network in Fresh Noodles during Storage. Food Chem. 2021, 344, 128638. [Google Scholar] [CrossRef]
- Chen, G.; Ehmke, L.; Miller, R.; Faa, P.; Smith, G.; Li, Y. Effect of Sodium Chloride and Sodium Bicarbonate on the Physicochemical Properties of Soft Wheat Flour Doughs and Gluten Polymerization. J. Agric. Food Chem. 2018, 66, 6840–6850. [Google Scholar] [CrossRef] [PubMed]
- Yan, H.-L.; Lu, Q.-Y. Effect of A- and B-Granules of Wheat Starch on Chinese Noodle Quality. J. Cereal Sci. 2020, 91, 102860. [Google Scholar] [CrossRef]
- Liu, C.; Song, M.; Hong, J.; Li, L.; Zheng, X.; Bian, K.; Guan, E. Effects of Salt and Kansui on Rheological, Chemical and Structural Properties of Noodle Dough during Repeated Sheeting Process. Food Chem. 2021, 342, 128365. [Google Scholar] [CrossRef] [PubMed]
- Ma, S.; Han, W.; Li, L.; Zheng, X.; Wang, X. The Thermal Stability, Structural Changeability, and Aggregability of Glutenin and Gliadin Proteins Induced by Wheat Bran Dietary Fiber. Food Funct. 2019, 10, 172–179. [Google Scholar] [CrossRef] [PubMed]
- Lafiandra, D.; Shewry, P.R. Wheat Glutenin Polymers 2. The Role of Wheat Glutenin Subunits in Polymer Formation and Dough Quality. J. Cereal Sci. 2022, 106, 103487. [Google Scholar] [CrossRef]
- Veraverbeke, W.S.; Delcour, J.A. Wheat Protein Composition and Properties of Wheat Glutenin in Relation to Breadmaking Functionality. Crit. Rev. Food Sci. 2002, 42, 179–208. [Google Scholar] [CrossRef] [PubMed]
- He, Z.H.; Liu, L.; Xia, X.C.; Liu, J.J.; Peña, R.J. Composition of HMW and LMW Glutenin Subunits and Their Effects on Dough Properties, Pan Bread, and Noodle Quality of Chinese Bread Wheats. Cereal Chem. 2005, 82, 345–350. [Google Scholar] [CrossRef]
- Shewry, P.; Halford, N.; Tatham, A. High Molecular Weight Subunits of Wheat Glutenin. J. Cereal Sci. 1992, 15, 105–120. [Google Scholar] [CrossRef]
- Payne, P.I.; Holt, L.M.; Lawrence, G.J. Detection of a Novel High Molecular Weight Subunit of Glutenin in Some Japanese Hexaploid Wheats. J. Cereal Sci. 1983, 1, 3–8. [Google Scholar] [CrossRef]
- Tatham, A.S.; Drake, A.F.; Shewry, P.R. Conformational Studies of Synthetic Peptides Corresponding to the Repetitive Regions of the High Molecular Weight (HMW) Glutenin Subunits of Wheat. J. Cereal Sci. 1990, 11, 189–200. [Google Scholar] [CrossRef]
- Shewry, P.R.; Tatham, A.S.; Barro, F.; Barcelo, P.; Lazzeri, P. Biotechnology of Breadmaking: Unraveling and Manipulating the Multi-Protein Gluten Complex. J. Biotechnol. 1995, 13, 1185–1190. [Google Scholar] [CrossRef] [PubMed]
- Humphris, A.D.L.; McMaster, T.J.; Miles, M.J.; Gilbert, S.M.; Shewry, P.R.; Tatham, A.S. Atomic Force Microscopy (AFM) Study of Interactions of HMW Subunits of Wheat Glutenin. Cereal Chem. 2000, 77, 107–110. [Google Scholar] [CrossRef]
- Gilbert, S.M.; Wellner, N.; Belton, P.S.; Greenfield, J.A.; Siligardi, G.; Shewry, P.R.; Tatham, A.S. Expression and Characterisation of a Highly Repetitive Peptide Derived from a Wheat Seed Storage Protein. Biochim. Biophys. Acta (BBA)-Protein Struct. Mol. Enzymol. 2000, 1479, 135–146. [Google Scholar] [CrossRef]
- Gao, X.; Liu, T.; Yu, J.; Li, L.; Feng, Y.; Li, X. Influence of High-Molecular-Weight Glutenin Subunit Composition at Glu-B1 Locus on Secondary and Micro Structures of Gluten in Wheat (Triticum aestivum L.). Food Chem. 2016, 197, 1184–1190. [Google Scholar] [CrossRef]
- Wang, C.; Shen, X.; Wang, K.; Liu, Y.; Zhou, J.; Hu, Y.; Zeller, F.J.; Hsam, S.L.K.; Yan, Y. Molecular Characterization and Functional Properties of Two Novel X-Type HMW-GS from Wheat Line CNU608 Derived from Chinese Spring × Ae. Caudata Cross. J. Cereal Sci. 2016, 68, 16–24. [Google Scholar] [CrossRef]
- Jiang, P.; Xue, J.; Duan, L.; Gu, Y.; Mu, J.; Han, S.; Chen, L.; Li, Y.; Ma, W.; Yan, Y.; et al. Effects of High-Molecular-Weight Glutenin Subunit Combination in Common Wheat on the Quality of Crumb Structure: Effects of High-Molecular-Weight Glutenin Subunit Combination in Common Wheat. J. Sci. Food Agric. 2019, 99, 1501–1508. [Google Scholar] [CrossRef]
- Song, L.; Li, L.; Zhao, L.; Liu, Z.; Xie, T.; Li, X. Absence of Dx2 at Glu-D1 Locus Weakens Gluten Quality Potentially Regulated by Expression of Nitrogen Metabolism Enzymes and Glutenin-Related Genes in Wheat. Int. J. Mol. Sci. 2020, 21, 1383. [Google Scholar] [CrossRef] [PubMed]
- Gao, X.; Liu, T.; Ding, M.; Wang, J.; Li, C.; Wang, Z.; Li, X. Effects of HMW-GS Ax1 or Dx2 Absence on the Glutenin Polymerization and Gluten Micro Structure of Wheat (Triticum aestivum L.). Food Chem. 2018, 240, 626–633. [Google Scholar] [CrossRef] [PubMed]
- Lindsay, M.P.; Skerritt, J.H. The Glutenin Macropolymer of Wheat our Doughs: Structure±function Perspectives. Trends Food Sci. Technol. 1999, 10, 247–253. [Google Scholar] [CrossRef]
- Ibba, M.I.; Kiszonas, A.M.; Guzmán, C.; Morris, C.F. Definition of the Low Molecular Weight Glutenin Subunit Gene Family Members in a Set of Standard Bread Wheat (Triticum aestivum L.) Varieties. J. Cereal Sci. 2017, 74, 263–271. [Google Scholar] [CrossRef]
- Masci, S.; Lew, E.J.-L.; LaFiandra, D.; Porceddu, E.; Kasarda, D.D. Characterization of Low Molecular Weight Glutenin Subunits in Durum Wheat by Reversed-Phase High-Performance Liquid Chromatography and N-Terminal Sequencing. Cereal Chem. 1995, 72, 100–104. [Google Scholar]
- D’Ovidio, R.; Masci, S. The Low-Molecular-Weight Glutenin Subunits of Wheat Gluten. J. Cereal Sci. 2004, 39, 321–339. [Google Scholar] [CrossRef]
- Kasarda, D.D. Glutenin Structure in Relation to Wheat Quality. Wheat Unique 1989, 277, 302. [Google Scholar]
- Anderson, O.D.; Hsia, C.C.; Adalsteins, A.E.; Lew, E.-L.; Kasarda, D.D. Identification of Several New Classes of Low-Molecular-Weight Wheat Gliadin-Related Proteins and Genes. Theor. Appl. Genet. 2001, 103, 307–315. [Google Scholar] [CrossRef]
- Ikeda, T.M.; Araki, E.; Fujita, Y.; Yano, H. Characterization of Low-Molecular-Weight Glutenin Subunit Genes and Their Protein Products in Common Wheats. Theor. Appl. Genet. 2006, 112, 327–334. [Google Scholar] [CrossRef]
- Franaszek, S.; Salmanowicz, B. Composition of Low-Molecular-Weight Glutenin Subunits in Common Wheat (Triticum aestivum L.) and Their Effects on the Rheological Properties of Dough. Open Life Sci. 2021, 16, 641–652. [Google Scholar] [CrossRef] [PubMed]
- Ting, Y.T.; Dahal-Koirala, S.; Kim, H.S.K.; Qiao, S.-W.; Neumann, R.S.; Lundin, K.E.A.; Petersen, J.; Reid, H.H.; Sollid, L.M.; Rossjohn, J. A Molecular Basis for the T Cell Response in HLA-DQ2.2 Mediated Celiac Disease. Proc. Natl. Acad. Sci. USA 2020, 117, 3063–3073. [Google Scholar] [CrossRef]
- Byers, M.; Miflin, B.J.; Smith, S.J. A Quantitative Comparison of the Extraction of Protein Fractions from Wheat Grain by Different Solvents, and of the Polypeptide and Amino Acid Composition of the Alcohol-Soluble Proteins. J. Sci. Food Agric. 1983, 34, 447–462. [Google Scholar] [CrossRef] [PubMed]
- Elzoghby, A.O.; Elgohary, M.M.; Kamel, N.M. Implications of Protein-and Peptide-Based Nanoparticles as Potential Vehicles for Anticancer Drugs. Adv Protein Chem. 2015, 98, 169–221. [Google Scholar]
- Sardari, R.R.R.; Sutiono, S.; Azeem, H.A.; Galbe, M.; Larsson, M.; Turner, C.; Nordberg Karlsson, E. Evaluation of Sequential Processing for the Extraction of Starch, Lipids, and Proteins From Wheat Bran. Front. Bioeng. Biotechnol. 2019, 7, 413. [Google Scholar] [CrossRef] [PubMed]
- Mehanna, M.M.; Mneimneh, A.T. Updated but Not Outdated “Gliadin”: A Plant Protein in Advanced Pharmaceutical Nanotechnologies. Int. J. Pharm. 2020, 587, 119672. [Google Scholar] [CrossRef] [PubMed]
- García-Molina, M.D.; Barro, F. Characterization of Changes in Gluten Proteins in Low-Gliadin Transgenic Wheat Lines in Response to Application of Different Nitrogen Regimes. Front. Plant Sci. 2017, 8, 257. [Google Scholar] [CrossRef] [PubMed]
- Fu, B.X. Asian Noodles: History, Classification, Raw Materials, and Processing. Food Res. Int. 2008, 41, 888–902. [Google Scholar] [CrossRef]
- Rombouts, I.; Jansens, K.J.A.; Lagrain, B.; Delcour, J.A.; Zhu, K.-X. The Impact of Salt and Alkali on Gluten Polymerization and Quality of Fresh Wheat Noodles. J. Cereal Sci. 2014, 60, 507–513. [Google Scholar] [CrossRef]
- Wang, J.-R.; Guo, X.-N.; Xing, J.-J.; Zhu, K.-X. Revealing the Effect Mechanism of NaCl on the Rheological Properties of Dough of Chinese Traditional Hand-Stretched Dried Noodles. Food Chem. 2020, 320, 126606. [Google Scholar] [CrossRef]
- Chen, G.; Ehmke, L.; Sharma, C.; Miller, R.; Faa, P.; Smith, G.; Li, Y. Physicochemical Properties and Gluten Structures of Hard Wheat Flour Doughs as Affected by Salt. Food Chem. 2019, 275, 569–576. [Google Scholar] [CrossRef]
- Han, C.; Ma, M.; Li, M.; Sun, Q. Further Interpretation of the Underlying Causes of the Strengthening Effect of Alkali on Gluten and Noodle Quality: Studies on Gluten, Gliadin, and Glutenin. Food Hydrocoll. 2020, 103, 105661. [Google Scholar] [CrossRef]
- Chen, G.; Li, Y. Aggregation Behavior of Semolina Gluten during Dough Production and Fresh Pasta Cooking upon Kansui Treatment. Food Chem. 2019, 278, 579–586. [Google Scholar] [CrossRef]
- Wang, L.; Hou, G.G.; Hsu, Y.-H.; Zhou, L. Effect of Phosphate Salts on the Korean Non-Fried Instant Noodle Quality. J. Cereal Sci. 2011, 54, 506–512. [Google Scholar] [CrossRef]
- Rahimi, G.; Hedayati, S.; Mazloomi, S.M. Studies on Functional Properties of Wheat Starch in the Presence of Lepidium Perfoliatum and Alyssum homolocarpum Seed Gums. J. Food Sci. 2021, 86, 699–704. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Fitzgerald, M.A.; Prakash, S.; Nicholson, T.M.; Gilbert, R.G. The Molecular Structural Features Controlling Stickiness in Cooked Rice, a Major Palatability Determinant. Sci. Rep. 2017, 7, 43713. [Google Scholar] [CrossRef] [PubMed]
- Ghumman, A.; Kaur, A.; Singh, N. Functionality and Digestibility of Albumins and Globulins from Lentil and Horse Gram and Their Effect on Starch Rheology. Food Hydrocoll. 2016, 61, 843–850. [Google Scholar] [CrossRef]
- Doublier, J.-L.; Marzin, C.; Visdeloup, S.; Lefebvre, J. Effect of Sodium Caseinate on the Pasting Behaviour of Starches from Different Origins. Carbohydrate 1994, 25, 228–229. [Google Scholar] [CrossRef]
- Kumar, L.; Brennan, M.A.; Mason, S.L.; Zheng, H.; Brennan, C.S. Rheological, Pasting and Microstructural Studies of Dairy Protein-Starch Interactions and Their Application in Extrusion-Based Products: A Review: Dairy Protein-Starch Interactions. Starch-Stärke 2017, 69, 1600273. [Google Scholar] [CrossRef]
- Wang, J.; Zhao, S.; Min, G.; Qiao, D.; Zhang, B.; Niu, M.; Jia, C.; Xu, Y.; Lin, Q. Starch-Protein Interplay Varies the Multi-Scale Structures of Starch Undergoing Thermal Processing. Int. J. Biol. Macromol. 2021, 175, 179–187. [Google Scholar] [CrossRef] [PubMed]
- Li, M.; Yue, Q.; Liu, C.; Zheng, X.; Hong, J.; Li, L.; Bian, K. Effect of Gliadin/Glutenin Ratio on Pasting, Thermal, and Structural Properties of Wheat Starch. J. Cereal Sci. 2020, 93, 102973. [Google Scholar] [CrossRef]
- Yao, M.; Li, M.; Dhital, S.; Tian, Y.; Guo, B. Texture and Digestion of Noodles with Varied Gluten Contents and Cooking Time: The View from Protein Matrix and Inner Structure. Food Chem. 2020, 315, 126230. [Google Scholar] [CrossRef]
- Zhang, S.B.; Lu, Q.Y.; Yang, H.; Meng, D.D. Effects of Protein Content, Glutenin-to-Gliadin Ratio, Amylose Content, and Starch Damage on Textural Properties of Chinese Fresh White Noodles. Cereal Chem. 2011, 88, 296–301. [Google Scholar] [CrossRef]
- Yue, F.-L.; Guo, X.-N.; Zhu, K.-X. Impact of Characteristics of Different Wheat Flours on the Quality of Frozen Cooked Noodles. Cereal Chem. 2017, 94, 881–886. [Google Scholar] [CrossRef]
- Silvas-García, M.I.; Ramírez-Wong, B.; Torres-Chávez, P.I.; Bello-Pérez, L.A.; Carvajal-Millán, E.; Barrón-Hoyos, J.M.; Rodríguez-García, M.E.; Vázquez-Lara, F.; Quintero-Ramos, A. Effect of Freezing Rate and Storage on the Rheological, Thermal and Structural Properties of Frozen Wheat Dough Starch: Effect of Freezing Rate and Storage on the Rheological. Starch-Stärke 2016, 68, 1103–1110. [Google Scholar] [CrossRef]
- Roppongi, T.; Ogawa, T.; Adachi, S. Water Sorption Kinetics of Wheat Noodle with Different Diameters. FSTR 2014, 20, 241–246. [Google Scholar] [CrossRef]
- Li, Y.; Chen, Y.; Li, S.; Gao, A.; Dong, S. Structural Changes of Proteins in Fresh Noodles during Their Processing. Int. J. Food Prop. 2017, 20, S202–S213. [Google Scholar] [CrossRef]
- Ooms, N.; Delcour, J.A. How to Impact Gluten Protein Network Formation during Wheat Flour Dough Making. Curr. Opin. Food Sci. 2019, 25, 88–97. [Google Scholar] [CrossRef]
- Uedaira, H.; Uedaira, H. The Effect of Sugars on the Thermal Denaturation of Lysozyme. Bull. Chem. Soc. Jpn. 1980, 53, 2451–2455. [Google Scholar] [CrossRef]
- Pareyt, B.; Brijs, K.; Delcour, J.A. Sugar-Snap Cookie Dough Setting: The Impact of Sucrose on Gluten Functionality. J. Agric. Food Chem. 2009, 57, 7814–7818. [Google Scholar] [CrossRef] [PubMed]
- Joye, I.J.; Lagrain, B.; Delcour, J.A. Use of Chemical Redox Agents and Exogenous Enzymes to Modify the Protein Network during Breadmaking—A Review. J. Cereal Sci. 2009, 50, 11–21. [Google Scholar] [CrossRef]
- Motoki, M.; Kumazawa, Y. Recent Research Trends in Transglutaminase Technology for Food Processing. FSTR 2000, 6, 151–160. [Google Scholar] [CrossRef]
- Huang, W.N.; Yuan, Y.L.; Kim, Y.S.; Chung, O.K. Effects of Transglutaminase on Rheology, Microstructure, and Baking Properties of Frozen Dough. Cereal Chem. 2008, 85, 301–306. [Google Scholar] [CrossRef]
- Wu, J.; Corke, H. Quality of Dried White Salted Noodles Affected by Microbial Transglutaminase. J. Sci. Food Agric. 2005, 85, 2587–2594. [Google Scholar] [CrossRef]
- Yokoyama, K.; Nio, N.; Kikuchi, Y. Properties and Applications of Microbial Transglutaminase. Appl. Microbiol. Biot. 2004, 64, 447–454. [Google Scholar] [CrossRef] [PubMed]
- Bauer, N.; Koehler, P.; Wieser, H.; Schieberle, P. Studies on Effects of Microbial Transglutaminase on Gluten Proteins of Wheat. I. Biochemical Analysis. Cereal Chem. 2003, 80, 781–786. [Google Scholar] [CrossRef]
- Autio, K.; Kruus, K.; Knaapila, A.; Gerber, N.; Flander, L.; Buchert, J. Kinetics of Transglutaminase-Induced Cross-Linking of Wheat Proteins in Dough. J. Agric. Food Chem. 2005, 53, 1039–1045. [Google Scholar] [CrossRef] [PubMed]
- Primo-Martín, C.; Valera, R.; Martínez-Anaya, M.A. Effect of Pentosanase and Oxidases on the Characteristics of Doughs and the Glutenin Macropolymer (GMP). J. Agric. Food Chem. 2003, 51, 4673–4679. [Google Scholar] [CrossRef]
- Steffolani, M.E.; Ribotta, P.D.; Pérez, G.T.; León, A.E. Effect of Glucose Oxidase, Transglutaminase, and Pentosanase on Wheat Proteins: Relationship with Dough Properties and Bread-Making Quality. J. Cereal Sci. 2010, 51, 366–373. [Google Scholar] [CrossRef]
- Bonet, A.; Rosell, C.M.; Caballero, P.A.; Gómez, M.; Pérez-Munuera, I.; Lluch, M.A. Glucose Oxidase Effect on Dough Rheology and Bread Quality: A Study from Macroscopic to Molecular Level. Food Chem. 2006, 99, 408–415. [Google Scholar] [CrossRef]
- Decamps, K.; Joye, I.J.; Rakotozafy, L.; Nicolas, J.; Courtin, C.M.; Delcour, J.A. The Bread Dough Stability Improving Effect of Pyranose Oxidase from Trametes Multicolor and Glucose Oxidase from Aspergillus Niger: Unraveling the Molecular Mechanism. J. Agric. Food Chem. 2013, 61, 7848–7854. [Google Scholar] [CrossRef] [PubMed]
- Morris, C.F. Determinants of Wheat Noodle Color. J. Sci. Food Agric. 2018, 98, 5171–5180. [Google Scholar] [CrossRef] [PubMed]
- Gujral, H.S.; Guardiola, I.; Carbonell, J.V.; Rosell, C.M. Effect of Cyclodextrinase on Dough Rheology and Bread Quality from Rice Flour. J. Agric. Food Chem. 2003, 51, 3814–3818. [Google Scholar] [CrossRef] [PubMed]
- Choo, C.L.; Aziz, N.A.A. Effects of Banana Flour and β-Glucan on the Nutritional and Sensory Evaluation of Noodles. Food Chem. 2010, 119, 34–40. [Google Scholar] [CrossRef]
- Li, L.; Zhou, W.; Wu, A.; Qian, X.; Xie, L.; Zhou, X.; Zhang, L. Effect of Ginkgo Biloba Powder on the Physicochemical Properties and Quality Characteristics of Wheat Dough and Fresh Wet Noodles. Foods 2022, 11, 698. [Google Scholar] [CrossRef] [PubMed]
- Zeeshan, M. Utilization of Mango Peels and Seed Kernels Powders as Sources of Phytochemicals in Biscuit. World J. Dairy Food Sci. 2011, 6, 35–42. [Google Scholar]
- Yu, K.; Zhou, H.-M.; Zhu, K.-X.; Guo, X.-N.; Peng, W. Water Cooking Stability of Dried Noodles Enriched with Different Particle Size and Concentration Green Tea Powders. Foods 2020, 9, 298. [Google Scholar] [CrossRef]
- Iuga, M.; Mironeasa, S. Potential of Grape Byproducts as Functional Ingredients in Baked Goods and Pasta. Compr. Rev. Food Sci. F. 2020, 19, 2473–2505. [Google Scholar] [CrossRef]
- Cao, Y.; Zhang, M.; Dong, S.; Guo, P.; Li, H. Impact of Potato Pulp on the Processing Characteristics and Gluten Structures of Wheat Flour Dough. J. Food Process. Preserv. 2020, 44, e14473. [Google Scholar] [CrossRef]
- Yan, Z.; Yuping, G.; Wenjia, H.; Yuanxia, S. Antioxidant Peptides from Freshwater Clam Extract Using Enzymatic Hydrolysis. Afr. J. Food Sci. 2014, 8, 148–154. [Google Scholar] [CrossRef]
- Liu, J.; Meenu, M.; Xu, B. Effect of Unripe Banana Flour and Wheat Gluten on Physicochemical Characteristics and Sensory Properties of White Salted Noodles. J. Food Process. Preserv. 2020, 44, e14513. [Google Scholar] [CrossRef]
- Tao, C.; Wang, K.; Liu, X.; Gou, E. Effects of Potato Starch on the Properties of Wheat Dough and the Quality of Fresh Noodles. CyTA J. Food 2020, 18, 427–434. [Google Scholar] [CrossRef]
- Wang, L.; Tian, Y.; Chen, Z.; Chen, J. Effects of Hericium Erinaceus Powder on the Digestion, Gelatinization of Starch, and Quality Characteristics of Chinese Noodles. Cereal Chem. 2021, 98, 482–491. [Google Scholar] [CrossRef]
- Shiau, S.; Li, G.; Pan, W.; Xiong, C. Effect of Pitaya Peel Powder Addition on the Phytochemical and Textural Properties and Sensory Acceptability of Dried and Cooked Noodles. J. Food Process. Preserv. 2020, 44, e14491. [Google Scholar] [CrossRef]
- Zhang, N.; Gao, Y.; Tong, L.; Li, Z. Superheated Steam Processing Improved the Qualities of Oats Flour and Noodles. J. Cereal Sci. 2018, 83, 96–100. [Google Scholar] [CrossRef]
- Ma, Y.J.; Guo, X.D.; Liu, H.; Xu, B.N.; Wang, M. Cooking, Textural, Sensorial, and Antioxidant Properties of Common and Tartary Buckwheat Noodles. Food Sci. Biotechnol. 2013, 22, 153–159. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Type | Predicted Structure |
|---|---|
| α/β-gliadins |  |
| γ-gliadins |  |
| ω-gliadins |  |
| Enzymes | Effects | References |
|---|---|---|
| Hexose oxidase (HOX) | Due to the formation of hydrogen peroxide, HOX causes S-S linkages to form between proteins and boosts the gelation of arabinoxylans. This increases the strength of dough and the volume of bread. | [60] |
| Laccase (LAC) | LAC treatment reduces dough extensibility and promotes dough stiffness. | [67] |
| Pyranose oxidase (P2O) | The molecular mechanism of improving dough stability by P2O involves the cross-linking of gluten proteins and arabinoxylan with formation of H2O2. | [70] |
| Polyphenol oxidase (PPO) | PPO would cause discoloration of the noodles. | [71] |
| Cyclodextrin Glycosyl transferase (CGT) | CGT decreases dough consistency but provides a good specific volume and a very soft crumb texture to bread. | [72] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zang, P.; Gao, Y.; Chen, P.; Lv, C.; Zhao, G. Recent Advances in the Study of Wheat Protein and Other Food Components Affecting the Gluten Network and the Properties of Noodles. Foods 2022, 11, 3824. https://doi.org/10.3390/foods11233824
Zang P, Gao Y, Chen P, Lv C, Zhao G. Recent Advances in the Study of Wheat Protein and Other Food Components Affecting the Gluten Network and the Properties of Noodles. Foods. 2022; 11(23):3824. https://doi.org/10.3390/foods11233824
Chicago/Turabian StyleZang, Peng, Yang Gao, Pu Chen, Chenyan Lv, and Guanghua Zhao. 2022. "Recent Advances in the Study of Wheat Protein and Other Food Components Affecting the Gluten Network and the Properties of Noodles" Foods 11, no. 23: 3824. https://doi.org/10.3390/foods11233824
APA StyleZang, P., Gao, Y., Chen, P., Lv, C., & Zhao, G. (2022). Recent Advances in the Study of Wheat Protein and Other Food Components Affecting the Gluten Network and the Properties of Noodles. Foods, 11(23), 3824. https://doi.org/10.3390/foods11233824